
Neurotropic Astroviruses in Animals


Abstract
:1. Introduction
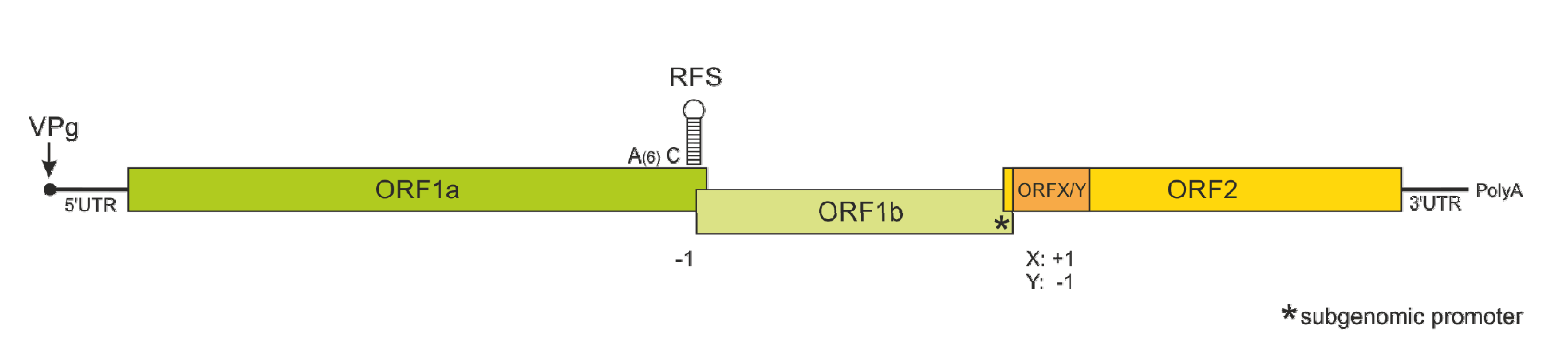
2. Molecular Biology of Astroviruses
3. Neuropathology of Astrovirus-Associated Encephalitis
4. Neurotropic Astroviruses in Different Animal Species
4.1. Neurotropic Astrovirus in Minks
4.2. Neurotropic Astroviruses in Pigs
4.3. Neurotropic Astroviruses in Cattle

{kind=link}
{kind=link}
{kind=link}
| Report Country/Reference | No. of Cases | Respiratory Tract 1 | Lymphatic Tissue | Serum | Liver | GIT 2 | Heart |
|---|---|---|---|---|---|---|---|
| Hungary [20] | 8 | 4/4 | 3/4 | 2/2 | 2/2 | 2/4 | 2/2 |
| USA [36] | 4 | 0/4 | nd | 0/4 | 0/4 | 0/4 | nd |
| USA [26] | 22 | 0/19 | 1/12 | nd | 0/18 | 2/15 | 0/16 |
| USA [38] | 3 | 0/2 | 0/2 | nd | 0/2 | 0/2 | 0/2 |
| Host | Report Country [References] | No. of Cases | Histopathology | ISH/IHC | Strain | GenBank Accession Number 1 |
|---|---|---|---|---|---|---|
| Cattle | USA [9] | 4 1 | + | + | BoAstV NeuroS1 | KF233994 |
| Cattle | Switzerland [10,28,31,46] | 47 1,2 | + | + | BoAstV CH13 | KM03579/KX266901 to KX266908 |
| Cattle | Switzerland [49] | 2 2 | + | + | BoAstV CH15 | KT956903 |
| Cattle | Uruguay [43] | 1 | + | + | BoAstV-Neuro-Uy | MK386569 |
| Cattle | Canada [27,47] | 6 1 | + | + | BoAstV NeuroS1/CH13 | KY614055/KY614056 |
| Cattle | Italy [45] | 1 | + | + | BoAstV PE3373/2019/Italy | MN46146 |
| Cattle | Japan [29] | 1 | + | nd | KagoshimaSR28-462Brain | LC341267 |
| Cattle | Germany [48] | 1 | + | + | BoAstV BH89/14 | LN879482 |
| Sheep | Switzerland [22,52] | 2 | + | + | OvAstV CH16 OvAstV CH17 | KY859988 MK286562 |
| Sheep | United Kingdom [53] | 2 | + | + | OvAstV UK/2013/ewe/lib01454 OvAstV UK/2014/lamb/lib01455 | LT706531 LT706530 |
| Muskox | Switzerland [21] | 1 | + | + | MOxAstV CH18 | MK211323 |
| Alpaca | Switzerland [24] | 1 | + | + | BoAstV-CH13/NeuroS1 | - |
4.4. Neurotropic Astroviruses in Sheep
4.5. Neurotropic Astrovirus in a Muskox
4.6. Neurotropic Astrovirus in an Alpaca
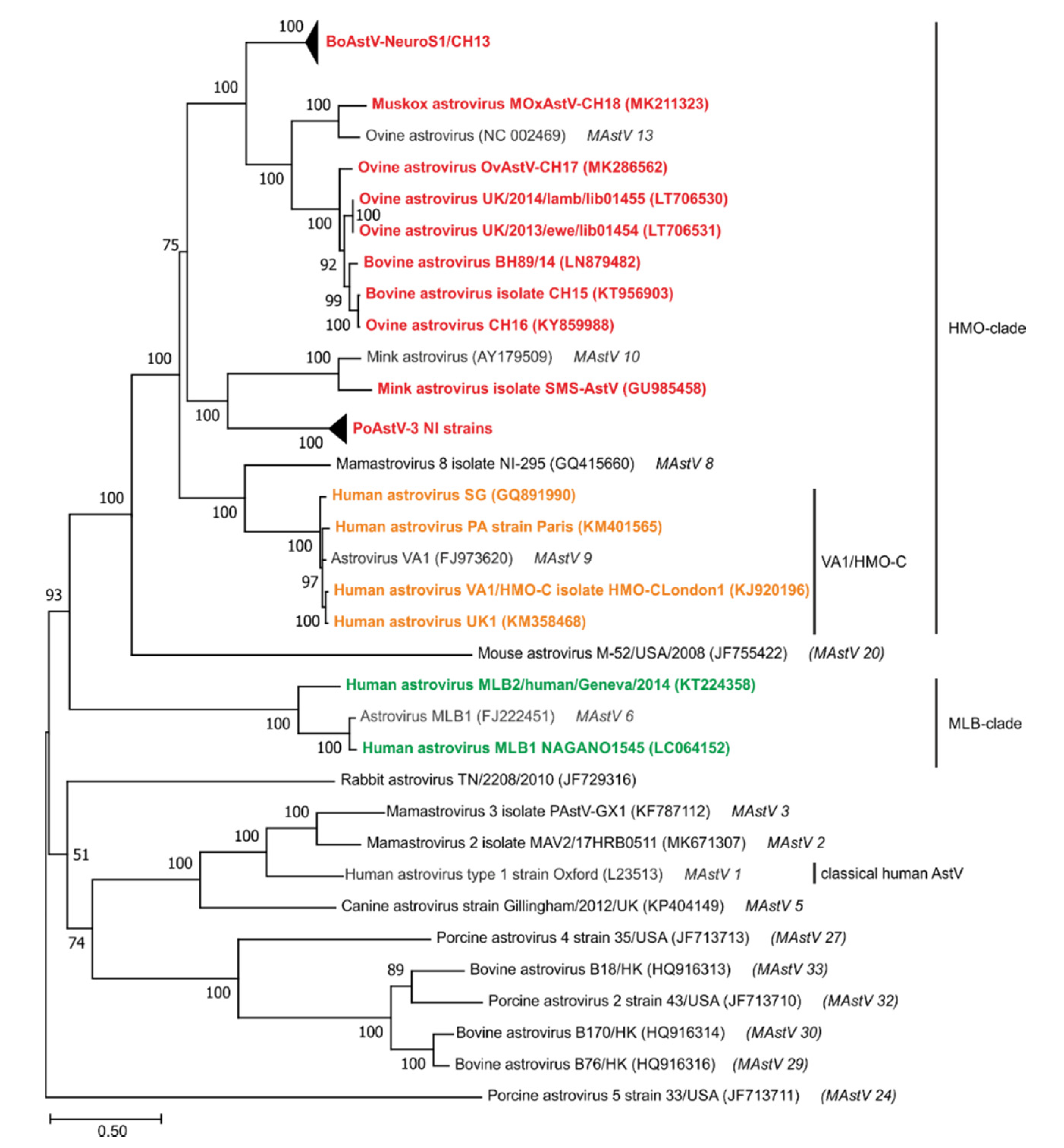
5. Phylogeny

6. Prophylaxis and Therapeutics
7. Pathogenesis
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- De Blauw, D.; Bruning, A.H.L.; Busch, C.B.E.; Kolodziej, L.M.; Jansen, N.J.G.; Van Woensel, J.B.M.; Pajkrt, D. Epidemiology and Etiology of Severe Childhood Encephalitis in the Netherlands. Pediatr. Infect. Dis. J. 2020, 39, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Granerod, J.; Tam, C.C.; Crowcroft, N.S.; Davies, N.W.S.; Borchert, M.; Thomas, S.L. Challenge of the unknown: A systematic review of acute encephalitis in non-outbreak situations. Neurology 2010, 75, 924–932. [Google Scholar] [CrossRef] [PubMed]
- Theil, D.; Fatzer, R.; Schiller, I.; Caplazi, P.; Zurbriggen, A.; Vandevelde, M. Neuropathological and aetiological studies of sporadic non-suppurative meningoencephalomyelitis of cattle. Vet. Rec. 1998, 143, 244–249. [Google Scholar] [CrossRef]
- Sánchez, S.; Clark, E.G.; Wobeser, G.A.; Janzen, E.D.; Philibert, H. A retrospective study of non-suppurative encephalitis in beef cattle from western Canada. Can. Vet. J. Rev. Vet. Can. 2013, 54, 1127–1132. [Google Scholar]
- Benjamin, L.A.; Lewthwaite, P.; Vasanthapuram, R.; Zhao, G.; Sharp, C.; Simmonds, P.; Wang, D.; Solomon, T. Human Parvovirus 4 as Potential Cause of Encephalitis in Children, India. Emerg. Infect. Dis. 2011, 17, 1484–1487. [Google Scholar] [CrossRef] [PubMed]
- Niller, H.H.; Angstwurm, K.; Rubbenstroth, D.; Schlottau, K.; Ebinger, A.; Giese, S.; Wunderlich, S.; Banas, B.; Forth, L.F.; Hoffmann, D.; et al. Zoonotic spillover infections with Borna disease virus 1 leading to fatal human encephalitis, 1999–2019: An epidemiological investigation. Lancet Infect. Dis. 2020, 20, 467–477. [Google Scholar] [CrossRef]
- Hierweger, M.M.; Werder, S.; Seuberlich, T. Parainfluenza Virus 5 Infection in Neurological Disease and Encephalitis of Cattle. Int. J. Mol. Sci. 2020, 21, 498. [Google Scholar] [CrossRef] [Green Version]
- Blomström, A.L.; Widén, F.; Hammer, A.S.; Belák, S.; Berg, M. Detection of a novel astrovirus in brain tissue of mink suffering from shaking mink syndrome by use of viral metagenomics. J. Clin. Microbiol. 2010, 48, 4392–4396. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Diab, S.; McGraw, S.; Barr, B.; Traslavina, R.; Higgins, R.; Talbot, T.; Blanchard, P.; Rimoldi, G.; Fahsbender, E.; et al. Divergent astrovirus associated with neurologic disease in cattle. Emerg. Infect. Dis. 2013, 19, 1385–1392. [Google Scholar] [CrossRef]
- Bouzalas, I.G.; Wüthrich, D.; Walland, J.; Drögemüller, C.; Zurbriggen, A.; Vandevelde, M.; Oevermann, A.; Bruggmann, R.; Seuberlich, T. Neurotropic Astrovirus in Cattle with Nonsuppurative Encephalitis in Europe. J. Clin. Microbiol. 2014, 52, 3318–3324. [Google Scholar] [CrossRef] [Green Version]
- Quan, P.L.; Wagner, T.A.; Briese, T.; Torgerson, T.R.; Hornig, M.; Tashmukhamedova, A.; Firth, C.; Palacios, G.; Baisre-de-Leon, A.; Paddock, C.D.; et al. Astrovirus encephalitis in boy with X-linked agammaglobulinemia. Emerg. Infect. Dis. 2010, 16, 918–925. [Google Scholar] [CrossRef] [PubMed]
- Appleton, H.; Higgins, P.G. Viruses and gastroenteritis in infants. Lancet 1975, 305, 1297. [Google Scholar] [CrossRef]
- Astroviridae-Positive Sense RNA Viruses-Positive Sense RNA Viruses (2011)—ICTV. Available online: https://talk.ictvonline.org/ictv-reports/ictv_9th_report/positive-sense-rna-viruses-2011/w/posrna_viruses/247/astroviridae (accessed on 31 May 2021).
- Shi, M.; Lin, X.D.; Chen, X.; Tian, J.H.; Chen, L.J.; Li, K.; Wang, W.; Eden, J.S.; Shen, J.J.; Liu, L.; et al. The evolutionary history of vertebrate RNA viruses. Nature 2018, 556, 197–202. [Google Scholar] [CrossRef]
- Cavanagh, D. Innovation and discovery: The application of nucleic acid-based technology to avian virus detection and characterization. Avian Pathol. 2001, 30, 581–598. [Google Scholar] [CrossRef]
- Chen, Q.; Xu, X.; Yu, Z.; Sui, C.; Zuo, K.; Zhi, G.; Ji, J.; Yao, L.; Kan, Y.; Bi, Y.; et al. Characterization and genomic analysis of emerging astroviruses causing fatal gout in goslings. Transbound. Emerg. Dis. 2020, 67, 865–876. [Google Scholar] [CrossRef]
- Bosch, A.; Pintó, R.M.; Guix, S. Human astroviruses. Clin. Microbiol. Rev. 2014, 27, 1048–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lulla, V.; Firth, A.E. A hidden gene in astroviruses encodes a viroporin. Nat. Commun. 2020, 11, 4070. [Google Scholar] [CrossRef] [PubMed]
- Reuter, G.; Pankovics, P.; Boros, Á. Nonsuppurative (Aseptic) Meningoencephalomyelitis Associated with Neurovirulent Astrovirus Infections in Humans and Animals. Clin. Microbiol. Rev. 2018, 31, e00040-18. [Google Scholar] [CrossRef] [Green Version]
- Boros, Á.; Albert, M.; Pankovics, P.; Bíró, H.; Pesavento, P.A.; Phan, T.G.; Delwart, E.; Reuter, G. Outbreaks of neuroinvasive astrovirus associated with encephalomyelitis, weakness, and paralysis among weaned pigs, Hungary. Emerg. Infect. Dis. 2017, 23, 1999–2010. [Google Scholar] [CrossRef]
- Boujon, C.L.; Koch, M.C.; Kauer, R.V.; Keller-Gautschi, E.; Hierweger, M.M.; Hoby, S.; Seuberlich, T. Novel encephalomyelitis-associated astrovirus in a muskox (Ovibos moschatus): A surprise from the archives. Acta Vet. Scand. 2019, 61, 31. [Google Scholar] [CrossRef]
- Küchler, L.; Koch, M.C.; Seuberlich, T.; Boujon, C.L. Archive mining brings to light a 25-year old astrovirus encephalitis case in a sheep. Front. Vet. Sci. 2019, 6, 51. [Google Scholar] [CrossRef]
- Dolores, G.-W.; Caroline, B.; Hans, H.D.; Englund, L.; Anne, S.H.; Hedlund, K.-O.; Carl Hård af, S.; Nilsson, K.; Nowotny, N.; Puurula, V.; et al. Investigations into Shaking Mink Syndrome: An Encephalomyelitis of Unknown Cause in Farmed Mink (Mustela Vison) Kits in Scandinavia. J. Vet. Diagn. Investig. 2004, 16, 305–312. [Google Scholar] [CrossRef]
- Küchler, L.; Rüfli, I.; Koch, M.C.; Hierweger, M.M.; Kauer, R.V.; Boujon, C.L.; Hilbe, M.; Oevermann, A.; Zanolari, P.; Seuberlich, T.; et al. Astrovirus-Associated Polioencephalomyelitis in an Alpaca. Viruses 2020, 13, 50. [Google Scholar] [CrossRef]
- Vandevelde, M.; Higgins, R.J.; Oevermann, A. Veterinary Neuropathology: Essentials of Theory and Practice, 1st ed.; John Wiley & Sons, Incorporated: Chichester, UK, 2012. [Google Scholar]
- Ferreyra, F.S.M.; Bradner, L.K.; Burrough, E.R.; Cooper, V.L.; Derscheid, R.J.; Gauger, P.C.; Harmon, K.M.; Madson, D.; Piñeyro, P.E.; Schwartz, K.J.; et al. Polioencephalomyelitis in Domestic Swine Associated with Porcine Astrovirus Type 3. Vet. Pathol. 2020, 57, 82–89. [Google Scholar] [CrossRef]
- Selimovic-Hamza, S.; Sanchez, S.; Philibert, H.; Clark, E.G.; Seuberlich, T. Bovine astrovirus infection in feedlot cattle with neurological disease in western Canada. Can. Vet. J. 2017, 58, 601–603. [Google Scholar] [CrossRef]
- Selimovic-Hamza, S.; Bouzalas, I.G.; Vandevelde, M.; Oevermann, A.; Seuberlich, T. Detection of astrovirus in historical cases of European sporadic bovine encephalitis, Switzerland 1958–1976. Front. Vet. Sci. 2016, 3, 91. [Google Scholar] [CrossRef] [Green Version]
- Hirashima, Y.; Okada, D.; Shibata, S.; Yoshida, S.; Fujisono, S.; Omatsu, T.; Mizutani, T.; Nagai, M. Whole genome analysis of a novel neurotropic bovine astrovirus detected in a Japanese black steer with non-suppurative encephalomyelitis in Japan. Arch. Virol. 2018, 163, 2805–2810. [Google Scholar] [CrossRef]
- Naccache, S.N.; Peggs, K.S.; Mattes, F.M.; Phadke, R.; Garson, J.A.; Grant, P.; Samayoa, E.; Federman, S.; Miller, S.; Lunn, M.P.; et al. Diagnosis of neuroinvasive astrovirus infection in an immunocompromised adult with encephalitis by unbiased next-generation sequencing. Clin. Infect. Dis. 2015, 60, 919–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selimovic-Hamza, S.; Boujon, C.L.; Hilbe, M.; Oevermann, A.; Seuberlich, T. Frequency and pathological phenotype of bovine astrovirus CH13/neuroS1 infection in neurologically-diseased cattle: Towards assessment of causality. Viruses 2017, 9, 12. [Google Scholar] [CrossRef]
- Boujon, C.L.; Selimovic-Hamza, S.; Bouzalas, I.; Seuberlich, T. Development and validation of an immunohistochemistry procedure for the detection of a neurotropic bovine astrovirus. J. Virol. Methods 2017, 239, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Englund, L.; Chriél, M.; Dietz, H.H.; Hedlund, K.O. Astrovirus epidemiologically linked to pre-weaning diarrhoea in mink. Vet. Microbiol. 2002, 85, 1–11. [Google Scholar] [CrossRef]
- Jakubczak, A.; Kowalczyk, M.; Mazurkiewicz, I.; Kondracki, M. Detection of mink astrovirus in Poland and further phylogenetic comparison with other European and Canadian astroviruses. Virus Genes 2021, 57, 258–265. [Google Scholar] [CrossRef]
- Blomström, A.L.; Ley, C.; Jacobson, M. Astrovirus as a possible cause of congenital tremor type AII in piglets? Acta Vet. Scand. 2014, 56, 82. [Google Scholar] [CrossRef] [Green Version]
- Arruda, B.; Arruda, P.; Hensch, M.; Chen, Q.; Zheng, Y.; Yang, C.; Gatto, I.R.H.; Ferreyra, F.M.; Gauger, P.; Schwartz, K.; et al. Porcine astrovirus type 3 in central nervous system of swine with polioencephalomyelitis. Emerg. Infect. Dis. 2017, 23, 2097–2100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, C.-T.; Halbur, P.G.; Opriessnig, T. Complete Genome Sequence of a Newly Identified Porcine Astrovirus Genotype 3 Strain US-MO123. J. Virol. 2012, 86, 13126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawal, G.; Ferreyra, F.M.; Macedo, N.R.; Bradner, L.K.; Harmon, K.M.; Allison, G.; Linhares, D.C.L.; Arruda, B.L. Ecology of Porcine Astrovirus Type 3 in a Herd with Associated Neurologic Disease. Viruses 2020, 12, 992. [Google Scholar] [CrossRef] [PubMed]
- Goecke, N.B.; Hjulsager, C.K.; Kongsted, H.; Boye, M.; Rasmussen, S.; Granberg, F.; Fischer, T.K.; Midgley, S.E.; Rasmussen, L.D.; Angen, O.; et al. No evidence of enteric viral involvement in the new neonatal porcine diarrhoea syndrome in Danish pigs. BMC Vet. Res. 2017, 13, 315. [Google Scholar] [CrossRef] [Green Version]
- Rawal, G.; Ferreyra, F.M.; MacEdo, N.R.; Bradner, L.K.; Harmon, K.M.; Mueller, A.; Allison, G.; Linhares, D.C.L.; Arruda, B.L. Detection and cellular tropism of porcine astrovirus type 3 on breeding farms. Viruses 2019, 11, 1051. [Google Scholar] [CrossRef] [Green Version]
- Xiao, C.T.; Giménez-Lirola, L.G.; Gerber, P.F.; Jiang, Y.H.; Halbur, P.G.; Opriessnig, T. Identification and characterization of novel porcine astroviruses (PAstVs) with high prevalence and frequent co-infection of individual pigs with multiple PAstV types. J. Gen. Virol. 2013, 94, 570–582. [Google Scholar] [CrossRef] [PubMed]
- Brnić, D.; Jemeršić, L.; Keros, T.; Prpić, J. High prevalence and genetic heterogeneity of porcine astroviruses in domestic pigs. Vet. J. 2014, 202, 390–392. [Google Scholar] [CrossRef] [PubMed]
- Giannitti, F.; Caffarena, R.D.; Pesavento, P.; Uzal, F.A.; Maya, L.; Fraga, M.; Colina, R.; Castells, M. The first case of bovine astrovirus-associated encephalitis in the Southern hemisphere (Uruguay), uncovers evidence of viral introduction to the Americas from Europe. Front. Microbiol. 2019, 10, 1240. [Google Scholar] [CrossRef] [Green Version]
- Truchet, L.; Walland, J.; Wüthrich, D.; Boujon, C.L.; Posthaus, H.; Bruggmann, R.; Schüpbach-Regula, G.; Oevermann, A.; Seuberlich, T. Neuropathological survey reveals underestimation of the prevalence of neuroinfectious diseases in cattle in Switzerland. Vet. Microbiol. 2017, 208, 137–145. [Google Scholar] [CrossRef]
- Zaccaria, G.; Lorusso, A.; Hierweger, M.M.; Malatesta, D.; Defourny, S.V.P.; Ruggeri, F.; Cammà, C.; Ricci, P.; Di Domenico, M.; Rinaldi, A.; et al. Detection of astrovirus in a cow with neurological signs by nanopore technology, Italy. Viruses 2020, 12, 530. [Google Scholar] [CrossRef] [PubMed]
- Wüthrich, D.; Boujon, C.L.; Truchet, L.; Selimovic-Hamza, S.; Oevermann, A.; Bouzalas, I.G.; Bruggmann, R.; Seuberlich, T. Exploring the virome of cattle with non-suppurative encephalitis of unknown etiology by metagenomics. Virology 2016, 493, 22–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spinato, M.T.; Vince, A.; Cai, H.; Ojkic, D. Identification of bovine astrovirus in cases of bovine non-suppurative encephalitis in eastern Canada. Can. Vet. J. 2017, 58, 607–609. [Google Scholar] [PubMed]
- Schlottau, K.; Schulze, C.; Bilk, S.; Hanke, D.; Höper, D.; Beer, M.; Hoffmann, B. Detection of a Novel Bovine Astrovirus in a Cow with Encephalitis. Transbound. Emerg. Dis. 2016, 63, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Seuberlich, T.; Wüthrich, D.; Selimovic-Hamza, S.; Drögemüller, C.; Oevermann, A.; Bruggmann, R.; Bouzalas, I. Identification of a second encephalitis-associated astrovirus in cattle. Emerg. Microbes Infect. 2016, 5, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Jonassen, C.M.; Jonassen, T.; Sveen, T.M.; Grinde, B. Complete genomic sequences of astroviruses from sheep and turkey: Comparison with related viruses. Virus Res. 2003, 91, 195–201. [Google Scholar] [CrossRef]
- Deiss, R.; Selimovic-Hamza, S.; Seuberlich, T.; Meylan, M. Neurologic Clinical Signs in Cattle With Astrovirus-Associated Encephalitis. J. Vet. Intern. Med. 2017, 31, 1209–1214. [Google Scholar] [CrossRef] [Green Version]
- Boujon, C.L.; Koch, M.C.; Wüthrich, D.; Werder, S.; Jakupovic, D.; Bruggmann, R.; Seuberlich, T. Indication of Cross-Species Transmission of Astrovirus Associated with Encephalitis in Sheep and Cattle. Emerg. Infect. Dis. 2017, 23, 1604–1608. [Google Scholar] [CrossRef] [Green Version]
- Pfaff, F.; Schlottau, K.; Scholes, S.; Courtenay, A.; Hoffmann, B.; Höper, D.; Beer, M. A novel astrovirus associated with encephalitis and ganglionitis in domestic sheep. Transbound. Emerg. Dis. 2017, 64, 677–682. [Google Scholar] [CrossRef]
- Snodgrass, D.R.; Gray, E.W. Detection and transmission of 30 nm virus particles (astroviruses) in faeces of lambs with diarrhoea. Arch. Virol. 1977, 55, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Jonassen, C.M.; Jonassen, T.; Saif, Y.M.; Snodgrass, D.R.; Ushijima, H.; Shimizu, M.; Grinde, B. Comparison of capsid sequences from human and animal astroviruses. J. Gen. Virol. 2001, 82, 1061–1067. [Google Scholar] [CrossRef]
- Koukou, G.; Niendorf, S.; Hornei, B.; Schlump, J.U.; Jenke, A.C.; Jacobsen, S. Human astrovirus infection associated with encephalitis in an immunocompetent child: A case report. J. Med. Case Rep. 2019, 13, 341. [Google Scholar] [CrossRef] [Green Version]
- Wunderli, W.; Meerbach, A.; Güngör, T.; Berger, C.; Greiner, O.; Caduff, R.; Trkola, A.; Bossart, W.; Gerlach, D.; Schibler, M.; et al. Astrovirus Infection in Hospitalized Infants with Severe Combined Immunodeficiency after Allogeneic Hematopoietic Stem Cell Transplantation. PLoS ONE 2011, 6, e27483. [Google Scholar] [CrossRef] [Green Version]
- Finkbeiner, S.R.; Li, Y.; Ruone, S.; Conrardy, C.; Gregoricus, N.; Toney, D.; Virgin, H.W.; Anderson, L.J.; Vinjé, J.; Wang, D.; et al. Identification of a Novel Astrovirus (Astrovirus VA1) Associated with an Outbreak of Acute Gastroenteritis. J. Virol. 2009, 83, 10836–10839. [Google Scholar] [CrossRef] [Green Version]
- Donato, C.; Vijaykrishna, D. The Broad Host Range and Genetic Diversity of Mammalian and Avian Astroviruses. Viruses 2017, 9, 102. [Google Scholar] [CrossRef] [Green Version]
- Boujon, C.L.; Koch, M.C.; Seuberlich, T. The Expanding Field of Mammalian Astroviruses: Opportunities and Challenges in Clinical Virology. Adv. Virus Res. 2017, 99, 109–137. [Google Scholar] [CrossRef] [PubMed]
- Guix, S.; Bosch, A.; Pintó, R.M. Astrovirus Research, 1st ed.; Schultz-Cherry, S., Ed.; Springer: New York, NY, USA, 2013. [Google Scholar]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press, Incorporated: Cary, NC, USA, 2000. [Google Scholar]
- Koci, M.D.; Schultz-Cherry, S. Avian astroviruses. Avian Pathol. 2002, 31, 213–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abad, F.X.; Pintó, R.M.; Villena, C.; Gajardo, R.; Bosch, A. Astrovirus survival in drinking water. Appl. Environ. Microbiol. 1997, 63, 3119–3122. [Google Scholar] [CrossRef] [Green Version]
- Kurtz, J.B.; Lee, T.W.; Parsons, A.J. The action of alcohols on rotavirus, astrovirus and enterovirus. J. Hosp. Infect. 1980, 1, 321–325. [Google Scholar] [CrossRef]
- Marvin, S.A.; Huerta, C.T.; Sharp, B.; Freiden, P.; Cline, T.D.; Schultz-Cherry, S. Type I Interferon Response Limits Astrovirus Replication and Protects against Increased Barrier Permeability In Vitro and In Vivo. J. Virol. 2016, 90, 1988–1996. [Google Scholar] [CrossRef] [Green Version]
- Janowski, A.B.; Dudley, H.; Wang, D. Antiviral activity of ribavirin and favipiravir against human astroviruses. J. Clin. Virol. 2020, 123, 104247. [Google Scholar] [CrossRef] [PubMed]
- Janowski, A.B.; Bauer, I.K.; Holtz, L.R.; Wang, D. Propagation of Astrovirus VA1, a Neurotropic Human Astrovirus, in Cell Culture. J. Virol. 2017, 91, e00740-17. [Google Scholar] [CrossRef] [Green Version]
- Superti, F.; Seganti, L.; Orsi, N.; Desideri, N.; Stein, M.L.; Tinari, A.; Marziano, M.L.; Donelli, G. In vitro effect of synthetic flavanoids on astrovirus infection. Antivir. Res. 1990, 13, 201–208. [Google Scholar] [CrossRef]
- Guix, S.; Pérez-Bosque, A.; Miró, L.; Moretó, M.; Bosch, A.; Pintó, R.M. Type I interferon response is delayed in human astrovirus infections. PLoS ONE 2015, 10, e0123087. [Google Scholar] [CrossRef] [PubMed]
- Hargest, V.; Sharp, B.; Livingston, B.; Cortez, V.; Schultz-Cherry, S. Astrovirus replication is inhibited by nitazoxanide in vitro and in vivo. J. Virol. 2020, 94. [Google Scholar] [CrossRef]
- Hargest, V.; Davis, A.E.; Tan, S.; Cortez, V.; Schultz-Cherry, S. Human astroviruses: A tale of two strains. Viruses 2021, 13, 376. [Google Scholar] [CrossRef]
- Brown, J.R.; Morfopoulou, S.; Hubb, J.; Emmett, W.A.; Ip, W.; Shah, D.; Brooks, T.; Paine, S.M.L.; Anderson, G.; Virasami, A.; et al. Astrovirus VA1/HMO-C: An increasingly recognized neurotropic pathogen in immunocompromised patients. Clin. Infect. Dis. 2015, 60, 881–888. [Google Scholar] [CrossRef]
- Lum, S.H.; Turner, A.; Guiver, M.; Bonney, D.; Martland, T.; Davies, E.; Newbould, M.; Brown, J.; Morfopoulou, S.; Breuer, J.; et al. An emerging opportunistic infection: Fatal astrovirus (VA1/HMO-C) encephalitis in a pediatric stem cell transplant recipient. Transpl. Infect. Dis. 2016, 18, 960–964. [Google Scholar] [CrossRef] [PubMed]
- Kauer, R.V.; Koch, M.C.; Schönecker, L.; Becker, J.; Holwerda, M.; Glaus, A.N.; Hierweger, M.M.; Werder, S.; Dijkman, R.; Meylan, M.; et al. Fecal Shedding of Bovine Astrovirus CH13/NeuroS1 in Veal Calves. J. Clin. Microbiol. 2020, 58, e01964-19. [Google Scholar] [CrossRef] [PubMed]
- Cordey, S.; Brito, F.; Vu, D.-L.; Turin, L.; Kilowoko, M.; Kyungu, E.; Genton, B.; Zdobnov, E.M.; D’Acremont, V.; Kaiser, L. Astrovirus VA1 identified by next-generation sequencing in a nasopharyngeal specimen of a febrile Tanzanian child with acute respiratory disease of unknown etiology. Emerg. Microbes Infect. 2016, 5, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Pérot, P.; Lecuit, M.; Eloit, M. Astrovirus Diagnostics. Viruses 2017, 9, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aroonprasert, D.; Fagerland, J.A.; Kelso, N.E.; Zheng, S.; Woode, G.N. Cultivation and partial characterization of bovine astrovirus. Vet. Microbiol. 1989, 19, 113–125. [Google Scholar] [CrossRef]
- Woode, G.N.; Bridger, J.C. Isolation of small viruses resembling astroviruses and caliciviruses from acute enteritis of calves. J. Med. Microbiol. 1978, 11, 441–452. [Google Scholar] [CrossRef] [Green Version]
- Harbour, D.A.; Ashley, C.R.; Williams, P.D.; Gruffydd-Jones, T.J. Natural and experimental astrovirus infection of cats. Vet. Rec. 1987, 120, 555–557. [Google Scholar] [CrossRef]
- Shimizu, M.; Shirai, J.; Narita, M.; Yamane, T. Cytopathic astrovirus isolated from porcine acute gastroenteritis in an established cell line derived from porcine embryonic kidney. J. Clin. Microbiol. 1990, 28, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Kolawole, A.O.; Mirabelli, C.; Hill, D.R.; Svoboda, S.A.; Janowski, A.B.; Passalacqua, K.D.; Rodriguez, B.N.; Dame, M.K.; Freiden, P.; Berger, R.P.; et al. Astrovirus replication in human intestinal enteroids reveals multi-cellular tropism and an intricate host innate immune landscape. PLoS Pathog. 2019, 15, e1008057. [Google Scholar] [CrossRef] [Green Version]
- Geigenmüller, U.; Ginzton, N.H.; Matsui, S.M. Construction of a genome-length cDNA clone for human astrovirus serotype 1 and synthesis of infectious RNA transcripts. J. Virol. 1997, 71, 1713–1717. [Google Scholar] [CrossRef] [Green Version]
- Wohlgemuth, N.; Honce, R.; Schultz-Cherry, S. Astrovirus evolution and emergence. Infect. Genet. Evol. 2019, 69, 30–37. [Google Scholar] [CrossRef]
- De Benedictis, P.; Schultz-Cherry, S.; Burnham, A.; Cattoli, G. Astrovirus infections in humans and animals—Molecular biology, genetic diversity, and interspecies transmissions. Infect. Genet. Evol. 2011, 11, 1529–1544. [Google Scholar] [CrossRef] [PubMed]
- Hata, A.; Kitajima, M.; Haramoto, E.; Lee, S.; Ihara, M.; Gerba, C.P.; Tanaka, H. Next-generation amplicon sequencing identifies genetically diverse human astroviruses, including recombinant strains, in environmental waters. Sci. Rep. 2018, 8, 11837. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wildi, N.; Seuberlich, T. Neurotropic Astroviruses in Animals. Viruses 2021, 13, 1201. https://doi.org/10.3390/v13071201
Wildi N, Seuberlich T. Neurotropic Astroviruses in Animals. Viruses. 2021; 13(7):1201. https://doi.org/10.3390/v13071201
Chicago/Turabian StyleWildi, Nicole, and Torsten Seuberlich. 2021. "Neurotropic Astroviruses in Animals" Viruses 13, no. 7: 1201. https://doi.org/10.3390/v13071201
APA StyleWildi, N., & Seuberlich, T. (2021). Neurotropic Astroviruses in Animals. Viruses, 13(7), 1201. https://doi.org/10.3390/v13071201