
Anti-SARS-CoV-2 Vaccines and Monoclonal Antibodies Facing Viral Variants



Abstract
1. Introduction
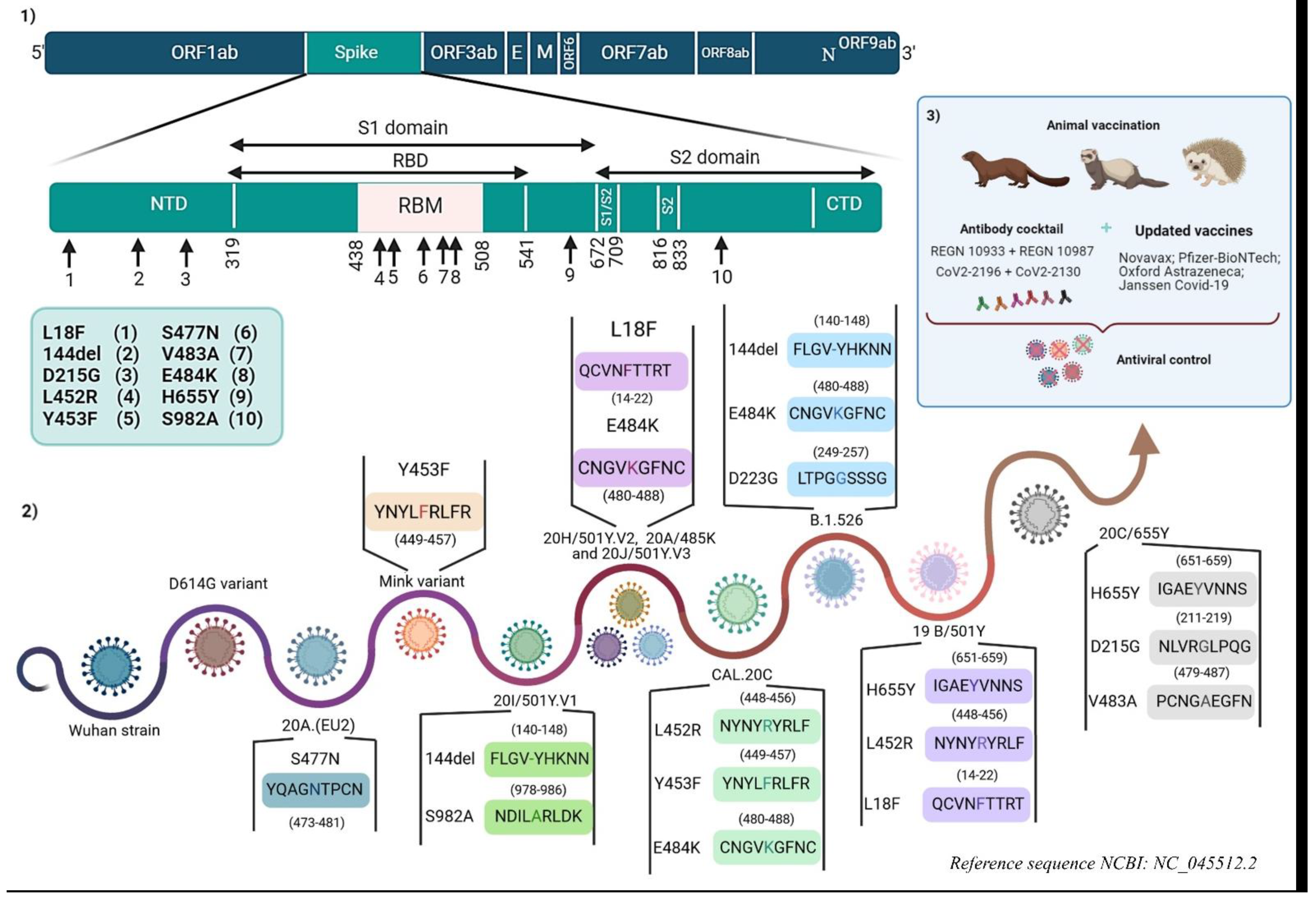
2. Genetic Variability
2.1. D614G Variant
2.2. 20I/501Y.V1 Variant (Lineage B.1.1.7)
2.3. 20H/501Y.V2 Variant (Lineage B.1.351)
2.4. 20J/501Y.V3 Variant (Lineage P.1)
2.5. Mink and CAL.20C Variants
2.6. 19B/501Y Variant (Lineage A.27)
2.7. 20C/655Y Variant (Lineage B.1.616)
2.8. 20C/477N and 20C/484K (Lineage B.1.526)
2.9. 20C/484K.V3 Variant (Lineage B.1.525)
2.10. 20B/484K (P.2 Lineage)
2.11. 20B/501Y Variant (P.3 Lineage)
2.12. 20A/214Ins Variant (B.1.214.2 Lineage)
2.13. 20A/484Q Variant (Lineage B.1.617)
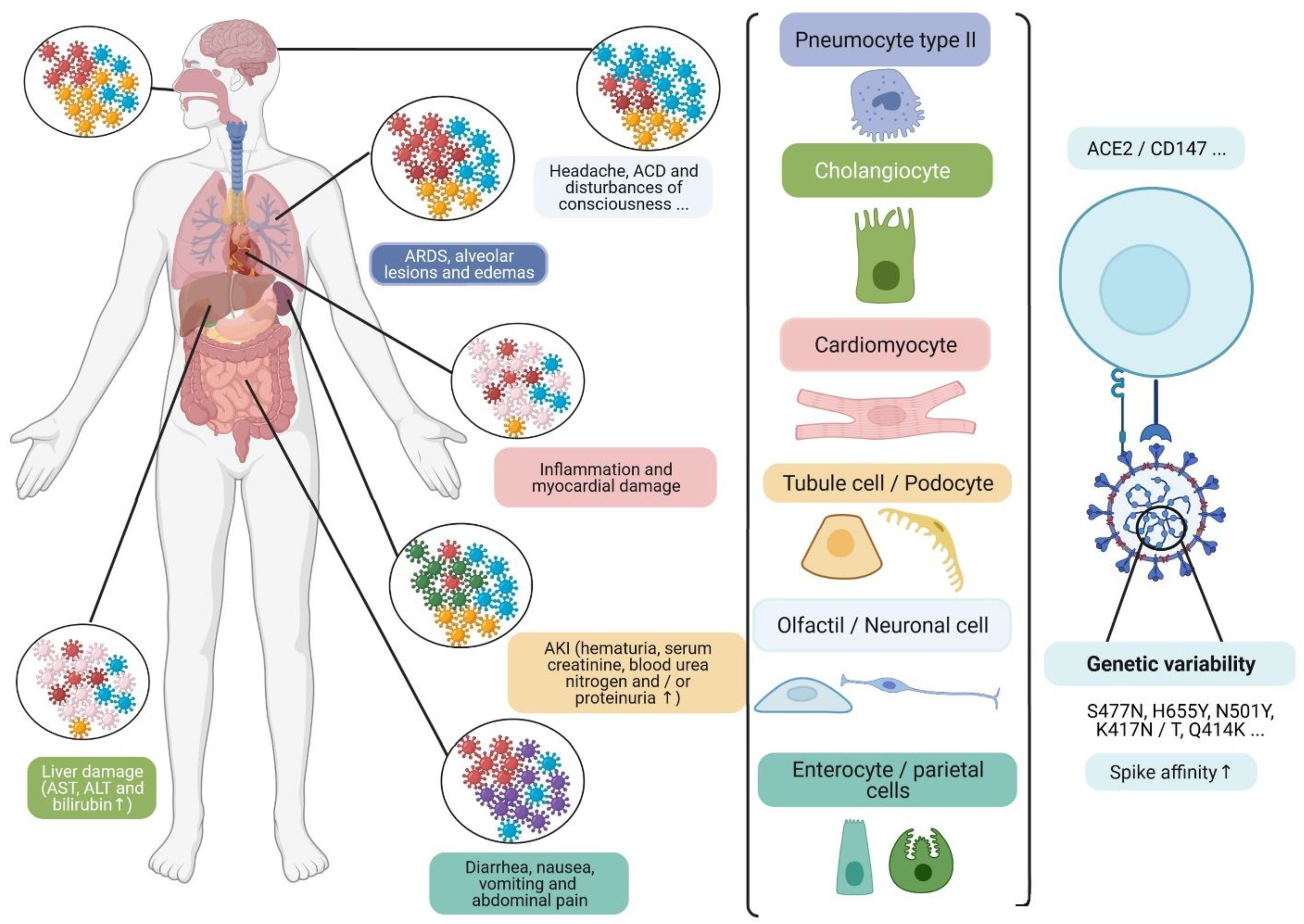
3. Viral Quasi-Species and Tissue Tropism
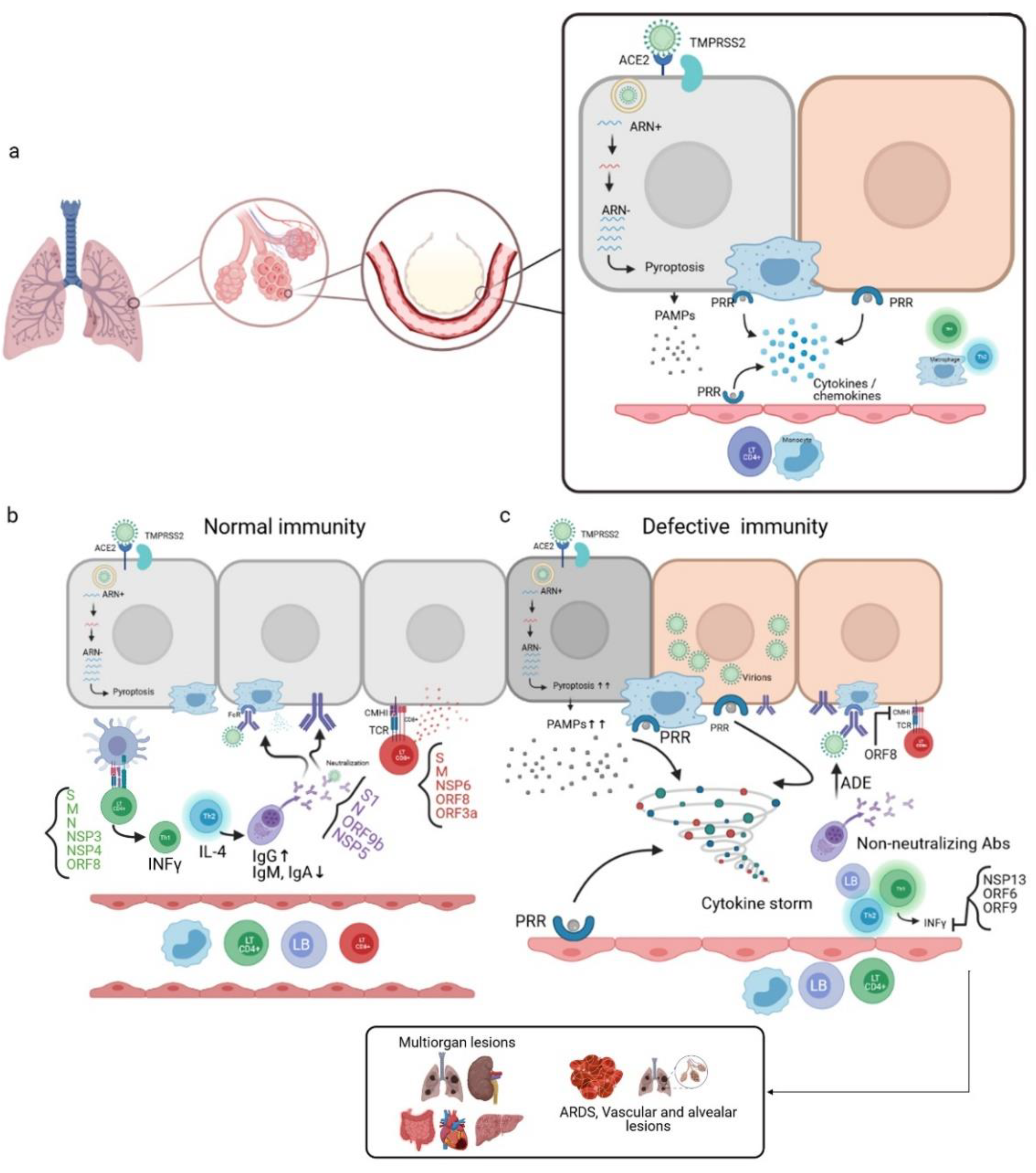
4. SARS-CoV-2 and Immunity
4.1. Natural and Adaptive Immunity
4.2. Vaccinable and Therapeutic Immunity
5. SARS-CoV-2 Variants Facing Vaccination and Monoclonal Antibodies
5.1. Variants and Monoclonal Antibodies
5.2. Variants and Vaccines
6. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef]
- Astuti, I. Ysrafil Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2): An Overview of Viral Structure and Host Response. Diabetes Metab. Syndr. Clin. Res. Rev. 2020, 14, 407–412. [Google Scholar] [CrossRef]
- Zheng, J. SARS-CoV-2: An Emerging Coronavirus That Causes a Global Threat. Int. J. Biol. Sci. 2020, 16, 1678–1685. [Google Scholar] [CrossRef] [PubMed]
- Zhai, X.; Sun, J.; Yan, Z.; Zhang, J.; Zhao, J.; Zhao, Z.; Gao, Q.; He, W.-T.; Veit, M.; Su, S. Comparison of Severe Acute Respiratory Syndrome Coronavirus 2 Spike Protein Binding to ACE2 Receptors from Human, Pets, Farm Animals, and Putative Intermediate Hosts. J. Virol. 2020, 94, e00831-20. [Google Scholar] [CrossRef] [PubMed]
- Creech, C.B.; Walker, S.C.; Samuels, R.J. SARS-CoV-2 Vaccines. JAMA 2021, 325, 1318–1320. [Google Scholar] [CrossRef]
- Dong, Y.; Dai, T.; Wei, Y.; Zhang, L.; Zheng, M.; Zhou, F. A Systematic Review of SARS-CoV-2 Vaccine Candidates. Signal Transduct. Target. Ther. 2020, 5, 237. [Google Scholar] [CrossRef] [PubMed]
- Shah, V.K.; Firmal, P.; Alam, A.; Ganguly, D.; Chattopadhyay, S. Overview of Immune Response During SARS-CoV-2 Infection: Lessons From the Past. Front. Immunol. 2020, 11, 1949. [Google Scholar] [CrossRef] [PubMed]
- Tay, M.Z.; Poh, C.M.; Rénia, L.; MacAry, P.A.; Ng, L.F.P. The Trinity of COVID-19: Immunity, Inflammation and Intervention. Nat. Rev. Immunol. 2020, 20, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zhou, X.; Qiu, Y.; Feng, F.; Feng, J.; Jia, Y.; Zhu, H.; Hu, K.; Liu, J.; Liu, Z.; et al. Clinical Characteristics of 82 Death Cases with COVID-19. medRxiv 2020. [Google Scholar] [CrossRef]
- Korber, B.; Fischer, W.M.; Gnanakaran, S.; Yoon, H.; Theiler, J.; Abfalterer, W.; Hengartner, N.; Giorgi, E.E.; Bhattacharya, T.; Foley, B.; et al. Tracking Changes in SARS-CoV-2 Spike: Evidence That D614G Increases Infectivity of the COVID-19 Virus. Cell 2020, 182, 812–827.e19. [Google Scholar] [CrossRef]
- Davies, N.G.; Abbott, S.; Barnard, R.C.; Jarvis, C.I.; Kucharski, A.J.; Munday, J.; Pearson, C.A.B.; Russell, T.W.; Tully, D.C.; Washburne, A.D.; et al. Estimated Transmissibility and Severity of Novel SARS-CoV-2 Variant of Concern 202012/01 in England. medRxiv 2021. [Google Scholar] [CrossRef]
- Tang, J.W.; Toovey, O.T.R.; Harvey, K.N.; Hui, D.D.S. Introduction of the South African SARS-CoV-2 Variant 501Y.V2 into the UK. J. Infect. 2021, 82, e8–e10. [Google Scholar] [CrossRef]
- Naveca, F.; da Costa, C.; Nascimento, V. SARS-CoV-2 Reinfection by the New Variant of Concern (VOC) P.1 in Amazonas, Brazil-SARS-CoV-2 Coronavirus/NCoV-2019 Genomic Epidemiology. Available online: https://virological.org/t/sars-cov-2-reinfection-by-the-new-variant-of-concern-voc-p-1-in-amazonas-brazil/596 (accessed on 14 March 2021).
- Santé Publique France. Analyse de Risque Liée Auxvariants Émergents de SARS-CoV-2 Réalisée Conjointement Par Le CNR Des Virus Des Infections Respiratoires et Santé Publique France; Report; Santé Publique France: Saint-Maurice, France, 2021; pp. 1–8.
- Sosin, D.V.; Tchurikov, N.A. Molecular Mechanisms of HIV-1 Genetic Diversity. Mol. Biol. 2017, 51, 547–560. [Google Scholar] [CrossRef]
- Al-Mawsawi, L.Q.; Wu, N.C.; Olson, C.A.; Shi, V.C.; Qi, H.; Zheng, X.; Wu, T.-T.; Sun, R. High-Throughput Profiling of Point Mutations across the HIV-1 Genome. Retrovirology 2014, 11, 124. [Google Scholar] [CrossRef]
- Dunfee, R.L.; Thomas, E.R.; Gorry, P.R.; Wang, J.; Taylor, J.; Kunstman, K.; Wolinsky, S.M.; Gabuzda, D. The HIV Env Variant N283 Enhances Macrophage Tropism and Is Associated with Brain Infection and Dementia. Proc. Natl. Acad. Sci. USA 2006, 103, 15160–15165. [Google Scholar] [CrossRef]
- Agence Nationale de Sécurité Sanitaire de l’Alimentation, de l’Environnement et du Travail. Saisine N° 2020-SA-0158 Relatif à La Surveillance Sanitaire à Mettre En Œuvre Pour Le SARS-CoV-2 Au Sein Des Élevages de Visons et Lien Avec La Santé Des Travailleurs Auprès Des Visons; Report; Agence Nationale de Sécurité Sanitaire de l’Alimentation, de l’Environnement et du Travail: Maisons-Alfort, France, 2021; pp. 1–26. [Google Scholar]
- Patiño-Galindo, J.Á.; Filip, I.; Chowdhury, R.; Maranas, C.D.; Sorger, P.K.; AlQuraishi, M.; Rabadan, R. Recombination and Lineage-Specific Mutations Linked to the Emergence of SARS-CoV-2. bioRxiv 2021. [Google Scholar] [CrossRef]
- Singh, D.; Yi, S.V. On the Origin and Evolution of SARS-CoV-2. Exp. Mol. Med. 2021, 53, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zai, J.; Zhao, Q.; Nie, Q.; Li, Y.; Foley, B.T.; Chaillon, A. Evolutionary History, Potential Intermediate Animal Host, and Cross-Species Analyses of SARS-CoV-2. J. Med Virol. 2020, 92, 602–611. [Google Scholar] [CrossRef] [PubMed]
- Chaw, S.-M.; Tai, J.-H.; Chen, S.-L.; Hsieh, C.-H.; Chang, S.-Y.; Yeh, S.-H.; Yang, W.-S.; Chen, P.-J.; Wang, H.-Y. The Origin and Underlying Driving Forces of the SARS-CoV-2 Outbreak. J. BioMed Sci. 2020, 27, 1–12. [Google Scholar] [CrossRef]
- Callaway, E. The Coronavirus Is Mutating—Does It Matter? Nature 2020, 585, 174–177. [Google Scholar] [CrossRef] [PubMed]
- Eskier, D.; Suner, A.; Oktay, Y.; Karakülah, G. Mutations of SARS-CoV-2 Nsp14 Exhibit Strong Association with Increased Genome-Wide Mutation Load. PeerJ 2020, 8, e10181. [Google Scholar] [CrossRef] [PubMed]
- Banoun, H. Évolution du SARS-CoV-2: Reveiw of Mutations, Role of the Host Immune System. Nephron 2021, 145, 392–403. [Google Scholar] [CrossRef]
- Yurkovetskiy, L.; Wang, X.; Pascal, K.E.; Tomkins-Tinch, C.; Nyalile, T.P.; Wang, Y.; Baum, A.; Diehl, W.E.; Dauphin, A.; Carbone, C.; et al. Structural and Functional Analysis of the D614G SARS-CoV-2 Spike Protein Variant. Cell 2020, 183, 739–751.e8. [Google Scholar] [CrossRef] [PubMed]
- Becerra-Flores, M.; Cardozo, T. SARS-CoV-2 Viral Spike G614 Mutation Exhibits Higher Case Fatality Rate. Int. J. Clin. Pract. 2020, 74, e13525. [Google Scholar] [CrossRef] [PubMed]
- Santé Publique France. COVID-19: Point Epidémiologique du 1er avril 2021; Report; Santé Publique France: Saint-Maurice, France, 2021; pp. 1–68.
- COVID-19 Scientific Council-France. Situation épidemiologique en Inde: Le variant B.1.617; Report; COVID-19 Scientific Council-France: Paris, France, 2021; Volume 9, pp. 1–36. [Google Scholar]
- WHO. SARS-CoV-2 Mink-Associated Variant Strain–Denmark. Available online: http://www.who.int/csr/don/06-november-2020-mink-associated-sars-cov2-denmark/en/ (accessed on 15 March 2021).
- OMS. Variant Du SARS-CoV-2–Royaume-Uni. Available online: http://www.who.int/csr/don/21-december-2020-sars-cov2-variant-united-kingdom/fr/ (accessed on 15 March 2021).
- Rambaut, A.; Loman, N.; Pybus, O.; Barclay, W. Preliminary Genomic Characterisation of an Emergent SARS-CoV-2 Lineage in the UK Defined by a Novel Set of Spike Mutations-SARS-CoV-2 Coronavirus/NCoV-2019 Genomic Epidemiology. Available online: https://virological.org/t/preliminary-genomic-characterisation-of-an-emergent-sars-cov-2-lineage-in-the-uk-defined-by-a-novel-set-of-spike-mutations/563 (accessed on 15 March 2021).
- Zhang, Y.; Zhang, J.; Chen, Y.; Luo, B.; Yuan, Y.; Huang, F.; Yang, T.; Yu, F.; Liu, J.; Liu, B.; et al. The ORF8 Protein of SARS-CoV-2 Mediates Immune Evasion through Potently Downregulating MHC-I. bioRxiv 2020, 111823. [Google Scholar] [CrossRef]
- Santé Publique France. COVID-19: Point Epidémiologique du 8 avril 2021; Report; Santé Publique France: Saint-Maurice, France, 2021; pp. 1–60.
- Tegally, H.; Wilkinson, E.; Giovanetti, M.; Iranzadeh, A.; Fonseca, V.; Giandhari, J.; Doolabh, D.; Pillay, S.; San, E.J.; Msomi, N.; et al. Emergence and Rapid Spread of a New Severe Acute Respiratory Syndrome-Related Coronavirus 2 (SARS-CoV-2) Lineage with Multiple Spike Mutations in South Africa. medRxiv 2020. [Google Scholar] [CrossRef]
- Hammer, A.S.; Quaade, M.L.; Rasmussen, T.B.; Fonager, J.; Rasmussen, M.; Mundbjerg, K.; Lohse, L.; Strandbygaard, B.; Jørgensen, C.S.; Alfaro-Núñez, A.; et al. SARS-CoV-2 Transmission between Mink (Neovison Vison) and Humans, Denmark. Emerg. Infect. Dis. 2021, 27, 547–551. [Google Scholar] [CrossRef]
- Bal, A.; Destras, G.; Gaymard, A.; Stefic, K.; Marlet, J.; Eymieux, S.; Regue, H.; Semanas, Q.; d’Aubarede, C.; Billaud, G.; et al. Two-Step Strategy for the Identification of SARS-CoV-2 Variant of Concern 202012/01 and Other Variants with Spike Deletion H69–V70, France, August to December 2020. Eurosurveillance 2021, 26, 2100008. [Google Scholar] [CrossRef]
- Koyama, T. Review 1: “Emergence of a Novel SARS-CoV-2 Strain in Southern California, USA”. Rapid Rev. Covid-19 2021, 20. [Google Scholar] [CrossRef]
- Mascola, J.R.; Graham, B.S.; Fauci, A.S. SARS-CoV-2 Viral Variants—Tackling a Moving Target. JAMA 2021, 325, 1261–1262. [Google Scholar] [CrossRef]
- Grabowski, F.; Kochańczyk, M.; Lipniacki, T. L18F Substrain of SARS-CoV-2 VOC-202012/01 Is Rapidly Spreading in England. medRxiv 2021. [Google Scholar] [CrossRef]
- Tchesnokova, V.; Kulakesara, H.; Larson, L.; Bowers, V.; Rechkina, E.; Kisiela, D.; Sledneva, Y.; Choudhury, D.; Maslova, I.; Deng, K.; et al. Acquisition of the L452R Mutation in the ACE2-Binding Interface of Spike Protein Triggers Recent Massive Expansion of SARS-Cov-2 Variants. bioRxiv 2021. [Google Scholar] [CrossRef]
- Van Dorp, L.; Tan, C.C.; Lam, S.D.; Richard, D.; Owen, C.; Berchtold, D.; Orengo, C.; Balloux, F. Recurrent Mutations in SARS-CoV-2 Genomes Isolated from Mink Point to Rapid Host-Adaptation. bioRxiv 2020, 1–27. [Google Scholar] [CrossRef]
- Baum, A.; Fulton, B.O.; Wloga, E.; Copin, R.; Pascal, K.E.; Russo, V.; Giordano, S.; Lanza, K.; Negron, N.; Ni, M.; et al. Antibody Cocktail to SARS-CoV-2 Spike Protein Prevents Rapid Mutational Escape Seen with Individual Antibodies. Science 2020, 369, 1014–1018. [Google Scholar] [CrossRef]
- Hodcroft, E.B.; Domman, D.B.; Oguntuyo, K.; Snyder, D.J.; Diest, M.V.; Densmore, K.H.; Schwalm, K.C.; Femling, J.; Carroll, J.L.; Scott, R.S.; et al. Emergence in Late 2020 of Multiple Lineages of SARS-CoV-2 Spike Protein Variants Affecting Amino Acid Position 677. medRxiv 2021. [Google Scholar] [CrossRef]
- West, A.P.; Barnes, C.O.; Yang, Z.; Bjorkman, P.J. SARS-CoV-2 Lineage B.1.526 Emerging in the New York Region Detected by Software Utility Created to Query the Spike Mutational Landscape. bioRxiv 2021. [Google Scholar] [CrossRef]
- Hou, Y.J.; Okuda, K.; Edwards, C.E.; Martinez, D.R.; Asakura, T.; Dinnon, K.H.; Kato, T.; Lee, R.E.; Yount, B.L.; Mascenik, T.M.; et al. SARS-CoV-2 Reverse Genetics Reveals a Variable Infection Gradient in the Respiratory Tract. Cell 2020, 182, 429–446.e14. [Google Scholar] [CrossRef]
- Zou, X.; Chen, K.; Zou, J.; Han, P.; Hao, J.; Han, Z. Single-Cell RNA-Seq Data Analysis on the Receptor ACE2 Expression Reveals the Potential Risk of Different Human Organs Vulnerable to 2019-NCoV Infection. Front. Med. 2020, 14, 185–192. [Google Scholar] [CrossRef]
- Lebeau, G.; Vagner, D.; Frumence, É.; Ah-Pine, F.; Guillot, X.; Nobécourt, E.; Raffray, L.; Gasque, P. Deciphering SARS-CoV-2 Virologic and Immunologic Features. Int. J. Mol. Sci 2020, 21, 5932. [Google Scholar] [CrossRef] [PubMed]
- Gabarre, P.; Dumas, G.; Dupont, T.; Darmon, M.; Azoulay, E.; Zafrani, L. Acute Kidney Injury in Critically Ill Patients with COVID-19. Intensive Care Med. 2020, 46, 1339–1348. [Google Scholar] [CrossRef]
- Hodcroft, E.B.; Zuber, M.; Nadeau, S.; Vaughan, T.G.; Crawford, K.H.D.; Althaus, C.L.; Reichmuth, M.L.; Bowen, J.E.; Walls, A.C.; Corti, D.; et al. Emergence and Spread of a SARS-CoV-2 Variant through Europe in the Summer of 2020. medRxiv 2021. [Google Scholar] [CrossRef]
- Global Initiative on Sharing Avian Influenza Data. Available online: https://www.epicov.org/epi3/frontend#598730 (accessed on 31 May 2021).
- Zhang, Y.; Chen, C.; Zhu, S.; Shu, C.; Wang, D.; Song, J.; Song, Y.; Zhen, W.; Feng, Z.; Wu, G.; et al. Isolation of 2019-NCoV from a Stool Specimen of a Laboratory-Confirmed Case of the Coronavirus Disease 2019 (COVID-19). CCDCW 2020, 2, 123–124. [Google Scholar] [CrossRef]
- Sbaoui, Y.; Bennis, F.; Chegdani, F. SARS-CoV-2 as Enteric Virus in Wastewater: Which Risk on the Environment and Human Behavior? Microbiol. Insights 2021, 14, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Sun, J.; Xu, Y.; Li, F.; Huang, X.; Li, H.; Zhao, J.; Huang, J.; Zhao, J. Infectious SARS-CoV-2 in Feces of Patient with Severe COVID-19. Emerg. Infect. Dis. J. CDC 2021, 26, 1920–1922. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.W.; Kim, S.-M.; Kim, H.-S.; Kim, Y.-I.; Kim, J.H.; Cho, J.Y.; Kim, S.; Kang, H.; Kim, S.-G.; Park, S.-J.; et al. Viable SARS-CoV-2 in Various Specimens from COVID-19 Patients. Clin. Microbio. Infect. 2020, 26, 1520–1524. [Google Scholar] [CrossRef]
- Poland, G.A.; Ovsyannikova, I.G.; Kennedy, R.B. SARS-CoV-2 Immunity: Review and Applications to Phase 3 Vaccine Candidates. Lancet 2020, 396, 1595–1606. [Google Scholar] [CrossRef]
- Iwasaki, A.; Yang, Y. The Potential Danger of Suboptimal Antibody Responses in COVID-19. Nat. Rev. Immunol. 2020, 20, 339–341. [Google Scholar] [CrossRef] [PubMed]
- Azkur, A.K.; Akdis, M.; Azkur, D.; Sokolowska, M.; van de Veen, W.; Brüggen, M.-C.; O’Mahony, L.; Gao, Y.; Nadeau, K.; Akdis, C.A. Immune Response to SARS-CoV-2 and Mechanisms of Immunopathological Changes in COVID-19. Allergy 2020, 75, 1564–1581. [Google Scholar] [CrossRef]
- Jackson, L.A.; Anderson, E.J.; Rouphael, N.G.; Roberts, P.C.; Makhene, M.; Coler, R.N.; McCullough, M.P.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; et al. An MRNA Vaccine against SARS-CoV-2—Preliminary Report. N. Engl. J. Med. 2020, 383, 1920–1931. [Google Scholar] [CrossRef]
- WHO. Draft Landscape and Tracker of COVID-19 Candidate Vaccines. Available online: https://www.who.int/publications/m/item/draft-landscape-of-covid-19-candidate-vaccines (accessed on 27 April 2021).
- La plateforme d’information sur les vaccinations. Vaccins contre le COVID-19: Liste des Essais Cliniques. Available online: www.infovac.ch/fr/infovac/actualites/955-vaccins-contre-le-covid-19-liste-des-essais-cliniques (accessed on 7 April 2021).
- Tuccori, M.; Ferraro, S.; Convertino, I.; Cappello, E.; Valdiserra, G.; Blandizzi, C.; Maggi, F.; Focosi, D. Anti-SARS-CoV-2 Neutralizing Monoclonal Antibodies: Clinical Pipeline. MAbs 2020, 12, 1854149. [Google Scholar] [CrossRef]
- Clinical Trials Arena. Bamlanivimab (LY-Cov555) for the Treatment of COVID-19, USA. Available online: https://www.clinicaltrialsarena.com/projects/bamlanivimab-ly-cov555-for-the-treatment-of-covid-19/ (accessed on 12 April 2021).
- Weinreich, D.M.; Sivapalasingam, S.; Norton, T.; Ali, S.; Gao, H.; Bhore, R.; Musser, B.J.; Soo, Y.; Rofail, D.; Im, J.; et al. REGN-COV2, a Neutralizing Antibody Cocktail, in Outpatients with Covid-19. N. Engl. J. Med. 2021, 384, 238–251. [Google Scholar] [CrossRef]
- Malik, J.A.; Mulla, A.H.; Farooqi, T.; Pottoo, F.H.; Anwar, S.; Rengasamy, K.R.R. Targets and Strategies for Vaccine Development against SARS-CoV-2. Biomed. Pharm. 2021, 137, 111254. [Google Scholar] [CrossRef]
- Diamond, M.; Chen, R.; Xie, X.; Case, J.; Zhang, X.; VanBlargan, L.; Liu, Y.; Liu, J.; Errico, J.; Winkler, E.; et al. SARS-CoV-2 Variants Show Resistance to Neutralization by Many Monoclonal and Serum-Derived Polyclonal Antibodies. Res. Sq. 2021. [Google Scholar] [CrossRef]
- Wang, P.; Liu, L.; Iketani, S.; Luo, Y.; Guo, Y.; Wang, M.; Yu, J.; Zhang, B.; Kwong, P.D.; Graham, B.S.; et al. Increased Resistance of SARS-CoV-2 Variants B.1.351 and B.1.1.7 to Antibody Neutralization. bioRxiv 2021. [Google Scholar] [CrossRef]
- Moore, J.P.; Offit, P.A. SARS-CoV-2 Vaccines and the Growing Threat of Viral Variants. JAMA 2021, 325, 821–822. [Google Scholar] [CrossRef] [PubMed]
- Forcelloni, S.; Benedetti, A.; Dilucca, M.; Giansanti, A. Identification of Conserved Epitopes in SARS-CoV-2 Spike and Nucleocapsid Protein. bioRxiv 2020. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variant | Date/Country of the 1st Detection | Key Mutations | Location | Potential Role/Major Impact |
|---|---|---|---|---|
| D614G [10] | End of January 2020/Germany and China | D614G 1 | CTD-S | Fitness/transmission advantage Correlation with lethality |
| Mink [30] | Denmark/ 7 November 2020 | Y453F 4 | RBM-S | Not associated to severity/death |
| N501T 2 | ||||
| 20I/501Y.V1 (B.1.1.7) [14] (VOC) | United Kingdom/ End of December 2020 | 69/70 Del 1,3 | NTD-S | up to 75% higher transmissibility Death (up to 82%)/severity increase Impact of S diagnosis kit |
| 144Y Del 4 | NTD-S | |||
| N501Y 2 | RBM-S | |||
| A570D | S1 region-Spike | |||
| D614G 1 | CTD-S | |||
| P681H 7 | Furin cleavage site | |||
| Q27stop 5 | ORF8 | |||
| 20I/484K/Q (B.1.1.7) [14] (VOC) | United Kingdom/ January 2021 | E484K 4/Q | RBD-S | Potential immune escape for the 20I/484K/Q variant |
| 20H/501Y.V2 (B.1.351) [14,28] (VOC) | South Africa/ December 2020 | K417N 2 | RBD-S | Highly transmissible (50%) Death (20%)/severity increase Post vaccine and infection immune escape |
| E484K 4 | RBD-S | |||
| N501Y 2/D614G 1 | Spike | |||
| L18F 4 | NTD-S | |||
| 20J/501Y.V3 (P.1) [14,28] (VOC) | Brazil/January 2021 | E484K 4, N501Y 2, D614G 1 | Spike | Transmissibility increase (up to 120%) Post vaccine and infection immune escape |
| K417N/T 2 | RBD-S | |||
| H655Y 2,4,6 | Near the furin cleavage site | |||
| 20A/484K (B.1.525) [14,28] (VOI) | United States and Nigeria/ December 2020 | 69/70 Del 1,3, 144-145del 4, E484K 4, Q52R, A677H, F888L | Spike | Sporadic detection in France No evidence of health impact |
| CAL.20C [14,28] (VOI) | Southern California/May 2020 | L452R 4,6 | RBD-S | Slight increase in transmissibility (20%) |
| 20C/477N and 20C/484K (B.1.526) [14] (VOI) | New York/February 2021 | D253G 4 | NTD-S | Some mutations may be involved in immune escape |
| D614G 1, E484K 4 | CTD-S | |||
| 19B/501Y(A27) [14] (VOI) | Henri Mondor Hospital-France/January 2021 | L18F 4, L452R 4,6, N501Y 2, H655Y 2,4,6 | Spike | Detection in the context of clusters Being evaluated Some mutations may be involved in immune escape |
| 20C/655Y (B.1.616) [14] (VOI) | Brittany-France/January 2021 | H66D | NTD-S | Detection in the context of clusters Being evaluated Some mutations may be involved in immune escape |
| 144-145del 4, H655Y 2,4,6 | Spike | |||
| D215G 4 | NTD | |||
| V483A 4 | RBD-S | |||
| 20A/214Ins (B.1.214.2) [14] (VUS) | Swiss and recently in Belgium/November 2020 | Ins214TDR | Spike | Being evaluated Some mutations may be involved in immune escape, transmissibility and ACE2R-S affinity increase |
| Q414K 2, N450K 4,2, D614G 1 | ||||
| T716I 2 | ||||
| 20A.(EU2) [14,28] | Europe/Summer 2020 | S477N 2,4 | RBM-S | Majority of sequences in Europe in autumn 2020 Some mutations may be involved in immune escape, and ACE2R-S affinity increase |
| D614G 1 | CTD-S | |||
| 20B/484K (P.2) [14] (VOI) | Brazil/April 2021 | E484K 4, D614G 1, | Spike | Being evaluated Detected sporadically in France |
| V1176F 2 | S2-Domain | |||
| 20B/501Y (P.3) [14,28] (VOI) | Philippines/January 2021 | E484K 4, N501Y 2, D614G 1, | Spike | Being evaluated Not detected in France |
| P681H 7, | Furin cleavage site | |||
| V1176F 2 | S2-Domain | |||
| 20A/484Q (B.1.617) [29] (VOI) | India (Maharashtra state (Mumbai))/October 2020 | E484Q, L452R 4,6 | Spike | Under investigation |
| Vaccine Type | Vaccine/mAb and Sponsor Names | Description of the Vaccine/mAb | Type of Test | Number of Participant/Phase |
|---|---|---|---|---|
| Inactivated vaccine [61] | Coronavac (Sinovac and Butantan Institute) | Inactivated SARS-CoV-2 virus, with aluminum salts | phase I/II + phase III | 144 (phase I); 600 (phase II) and 8870 (phase III) |
| Inactivated SARS-CoV-2 Vaccine (Chinese Academy of Medical Sciences) | Inactivated SARS-CoV-2 virus | phase I/II | 942 | |
| BBIBP-CorV (Beijing Institute of Biological Products Co., Ltd. and Laboratorio Elea Phoenix S.A.) | phase I/II and phase III | 448 (phase I); 1412 (phase II) and 3000 (phase III) | ||
| Subunit/purified vaccines [61] | NVX-CoV2373 (Novavax) | Nanoparticles containing Spike trimers with Matrix-M adjuvant | phase I/II, phase II in South Africa, and phase III (UK) + phase III (USA, Mexico and Puerto Rico) | 1631; 2904; 15,000 and 30,000 |
| UB-612 (United Biomedical, COVAXX) | S1 and S2 subunits of the Spike protein and M and N proteins of SARS-CoV-2 | phase I + phase II/III | 60 and 7320 | |
| SCB-2019 (Clover Biopharmaceuticals Australia) | SARS-CoV-2 Spike protein subunit in trimer form, with or without adjuvants (adjuvant 1: AS03/adjuvant 2: oligonucleotide “CpG 1018” + aluminum salts) | phase I + phase II/III | 150 and 34,000 | |
| Vector vaccines [61] | AZD1222 (ChAdOx1 nCoV-19) (University of Oxford and Astra Zeneca) | Chimpanzee Adenovirus expressing the SARS-CoV-2 Spike protein | phase I/II + phaseIIb/III + phase III | 1090; 10,260 and 40,050 |
| GRAd-COV2 (ReiThera Srl) | Inactivated gorilla Adenovirus, expressing the SARS-CoV-2 Spike protein | phase I | 90 | |
| Ad26.COV2.S (Johnson & Johnson) | Inactivated human Adenovirus type 26 expressing the SARS-CoV-2 Spike | phase I/II + phase III | 1045 and 60,000 | |
| Genetic vaccines (DNA, RNA) [61] | BNT162 (Pfizer-BioNTech) | mRNA encoding the SARS-CoV-2 Spike protein, encapsulated in a lipid nanoparticle | phase I/II+ phase I/II + phase III | 196; 7600 and 30,000 |
| mRNA-1273 (Moderna) | mRNA encoding the SARS-CoV-2 Spike | phase I + phase II + III | 105,600 and 30,000 | |
| AG0301-COVID19 (AnGes, Inc., Japan Agency for Medical Research and Development) | Spike encoding DNA plasmid | phase I/II | 30 | |
| Monoclonal antibodies [62] | Bamlanivimab (LY-CoV555)/(Eli Lilly and Company) [63] | recombinant neutralizing human IgG1κ | Authorized by US Food and Drug Administration (FDA) in November 2020 | 465 (phase II) |
| Casirivimab (REGN10933) + Imdevimab (REGN10987) [64] | Cocktail of Spike neutralization antibodies | 799 (Phase I-III) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaqroun, A.; Hartard, C.; Schvoerer, E. Anti-SARS-CoV-2 Vaccines and Monoclonal Antibodies Facing Viral Variants. Viruses 2021, 13, 1171. https://doi.org/10.3390/v13061171
Chaqroun A, Hartard C, Schvoerer E. Anti-SARS-CoV-2 Vaccines and Monoclonal Antibodies Facing Viral Variants. Viruses. 2021; 13(6):1171. https://doi.org/10.3390/v13061171
Chicago/Turabian StyleChaqroun, Ahlam, Cédric Hartard, and Evelyne Schvoerer. 2021. "Anti-SARS-CoV-2 Vaccines and Monoclonal Antibodies Facing Viral Variants" Viruses 13, no. 6: 1171. https://doi.org/10.3390/v13061171
APA StyleChaqroun, A., Hartard, C., & Schvoerer, E. (2021). Anti-SARS-CoV-2 Vaccines and Monoclonal Antibodies Facing Viral Variants. Viruses, 13(6), 1171. https://doi.org/10.3390/v13061171