
HIV-1 Persistence in Children during Suppressive ART

{kind=link}
{kind=link}
Abstract
1. Introduction
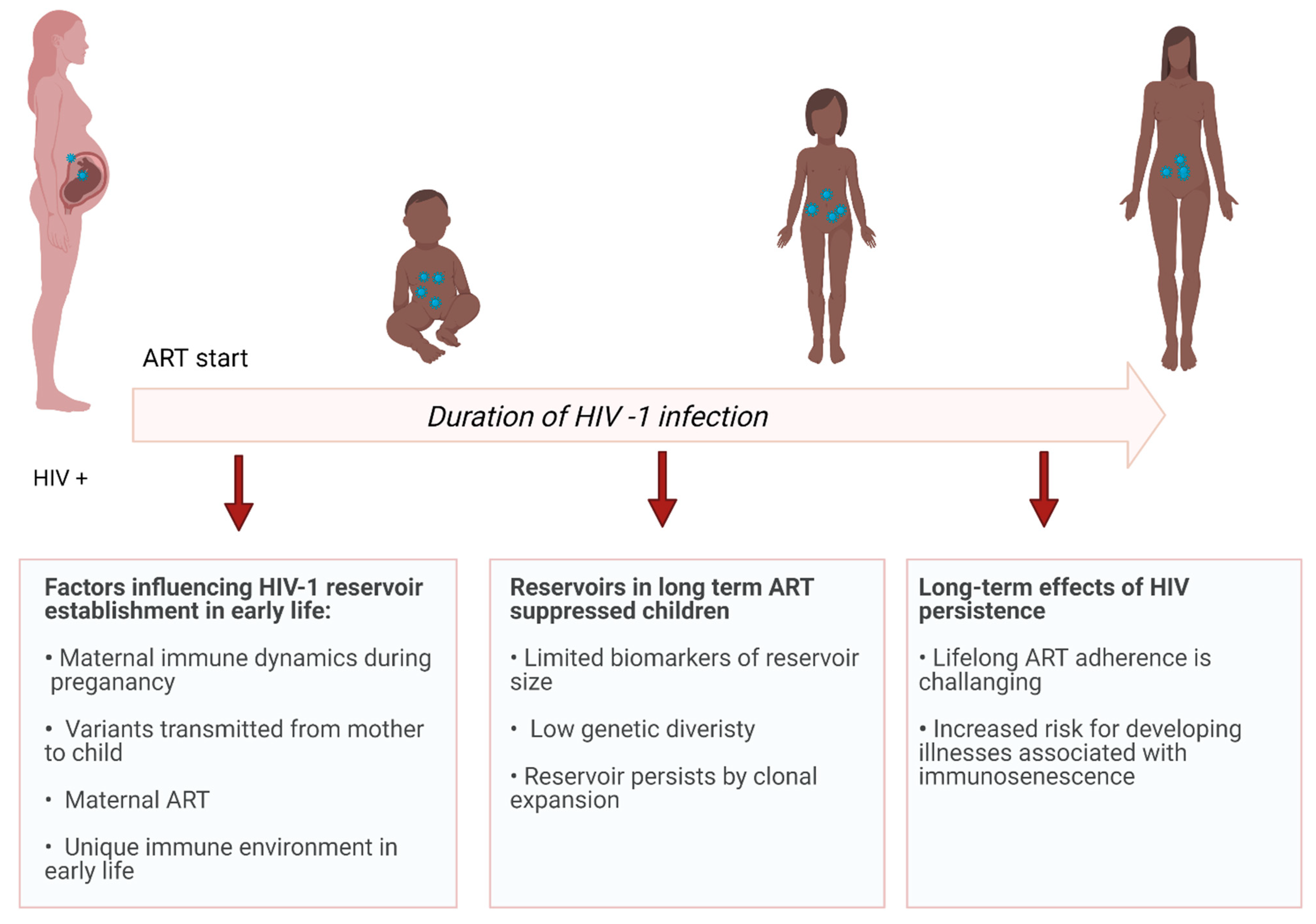
2. Reservoir Establishment in Early Life
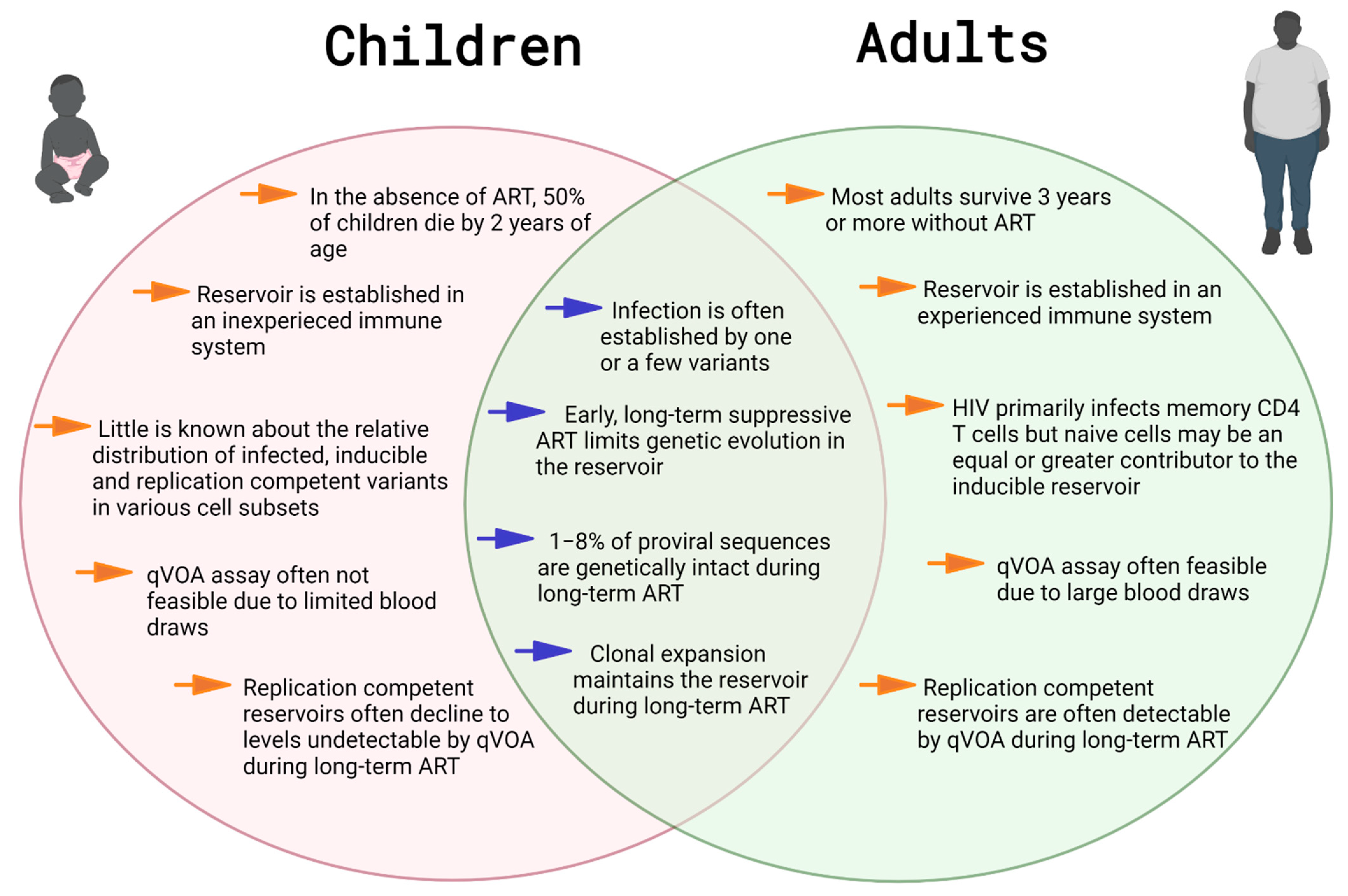
3. Reservoirs in Early-Treated, Long-Term Suppressed Children
3.1. HIV-1 Cell-Associated DNA: A Biomarker of the Reservoir in Long-Term-Treated Children
3.2. Measuring Infectious Virus in Long-Term Treated Children
3.3. Post-ART Control of Viral Replication
3.4. Genetic Diversity during Long-Term ART
3.5. Reservoir Decay Kinetics and the Proviral Landscape during Long-Term ART
4. Clonal Expansion Maintains the Reservoir in Treated Children
5. Long-Term Effects of HIV Persistence in Perinatally Infected Individuals
6. Conclusions
Funding
Conflicts of Interest
References
- UNAIDS. Global HIV & AIDS Statistics–2020 Fact Sheet; UNAIDS: Geneva, Switzerland, 2020. [Google Scholar]
- Myer, L.; Essajee, S.; Broyles, L.N.; Watts, D.H.; Lesosky, M.; El-Sadr, W.M.; Abrams, E.J. Pregnant and breastfeeding women: A priority population for HIV viral load monitoring. PLoS Med. 2017, 14, e1002375. [Google Scholar] [CrossRef]
- Moland, K.M.I.; De Paoli, M.M.; Sellen, D.W.; Van Esterik, P.; Leshabari, S.C.; Blystad, A. Breastfeeding and HIV: Experiences from a decade of prevention of postnatal HIV transmission in sub-Saharan Africa. Int. Breastfeed. J. 2010, 5, 10. [Google Scholar] [CrossRef]
- Myburgh, D.; Rabie, H.; Slogrove, A.; Edson, C.; Cotton, M.; Dramowski, A. Horizontal HIV transmission to children of HIV-uninfected mothers: A case series and review of the global literature. Int. J. Infect. Dis. 2020, 98, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Scott, G.B.; Brogly, S.B.; Muenz, D.; Stek, A.M.; Read, J.S.; for the International Maternal Pediatric Adolescent AIDS Clinical Trials Group (IMPAACT) P1025 Study Team. Missed Opportunities for Prevention of Mother-to-Child Transmission of Human Immunodeficiency Virus. Obstet. Gynecol. 2017, 129, 621–628. [Google Scholar] [CrossRef] [PubMed]
- WHO. Global Health Sector Strategy on HIV 2016–2021 towards Ending AIDS. 2016. Available online: https://apps.who.int/iris/bitstream/handle/10665/246178/WHO-HIV-2016.05-eng.pdf?sequence=1 (accessed on 2 February 2021).
- Jewell, B.L.; Mudimu, E.; Stover, J.; Brink, D.T.; Phillips, A.N.; Smith, J.A.; Martin-Hughes, R.; Teng, Y.; Glaubius, R.; Mahiane, S.G.; et al. Potential effects of disruption to HIV programmes in sub-Saharan Africa caused by COVID-19: Results from multiple mathematical models. Lancet HIV 2020, 7, e629–e640. [Google Scholar] [CrossRef]
- Van Elsland, S.L.; Peters, R.P.H.; Grobbelaar, N.; Ketelo, P.; Kok, M.O.; Cotton, M.F.; Van Furth, A.M. Paediatric ART Adherence in South Africa: A comprehensive analysis. AIDS Behav. 2019, 23, 475–488. [Google Scholar] [CrossRef] [PubMed]
- Ammon, N.; Mason, S.; Corkery, J. Factors impacting antiretroviral therapy adherence among human immunodeficiency virus–positive adolescents in Sub-Saharan Africa: A systematic review. Public Health 2018, 157, 20–31. [Google Scholar] [CrossRef]
- Brown, T.T.; Tassiopoulos, K.; Bosch, R.J.; Shikuma, C.; Mccomsey, G.A. Association Between Systemic Inflammation and Incident Diabetes in HIV-Infected Patients After Initiation of Antiretroviral Therapy. Diabetes Care 2010, 33, 2244–2249. [Google Scholar] [CrossRef]
- Nansseu, J.R.; Bigna, J.J.; Kaze, A.D.; Noubiap, J.J. Incidence and risk factors for prediabetes and diabetes mellitus among HIV-infected adults on antiretroviral therapy: A systematic review and meta-analysis. Epidemiology 2018, 29, 431–441. [Google Scholar] [CrossRef]
- Van Zoest, R.A.; van den Born, B.-J.H.; Reiss, P. Hypertension in people living with HIV. Curr. Opin. HIV AIDS 2017, 12, 513–522. [Google Scholar] [CrossRef]
- Sarfo, F.S.; Singh, A.; Tagge, R.; Mensah, G.; Ovbiagele, B. Duration of antiretroviral therapy among people living with HIV and incidence of hypertension in Ghana. J. Clin. Hypertens. 2020, 22, 2361–2371. [Google Scholar] [CrossRef]
- Dimala, C.A.; Blencowe, H.; Choukem, S.P. The association between antiretroviral therapy and selected cardiovascular disease risk factors in sub-Saharan Africa: A systematic review and meta-analysis. PLoS ONE 2018, 13, e0201404. [Google Scholar] [CrossRef]
- Sabin, C.A. Do people with HIV infection have a normal life expectancy in the era of combination antiretroviral therapy? BMC Med. 2013, 11, 251. [Google Scholar] [CrossRef] [PubMed]
- Brooks, D.G.; Kitchen, S.G.; Kitchen, C.M.R.; Scripture-Adams, D.D.; Zack, J.A. Generation of HIV latency during thymopoiesis. Nat. Med. 2001, 7, 459–464. [Google Scholar] [CrossRef]
- Siliciano, R.F.; Greene, W.C. HIV Latency. Cold Spring Harb. Perspect. Med. 2011, 1, a007096. [Google Scholar] [CrossRef]
- Chun, T.-W.; Stuyver, L.; Mizell, S.B.; Ehler, L.A.; Mican, J.A.M.; Baseler, M.; Lloyd, A.; Nowak, M.A.; Fauci, A.S. Presence of an inducible HIV-1 latent reservoir during highly active antiretroviral therapy. Proc. Natl. Acad. Sci. USA 1997, 94, 13193–13197. [Google Scholar] [CrossRef]
- Newell, M.-L.; Coovadia, H.; Cortina-Borja, M.; Rollins, N.; Gaillard, P.; Dabis, F. Mortality of infected and uninfected infants born to HIV-infected mothers in Africa: A pooled analysis. Lancet 2004, 364, 1236–1243. [Google Scholar] [CrossRef]
- Patrikar, S.; Basannar, D.; Bhatti, V.; Kotwal, A.; Gupta, R.; Grewal, R. Rate of decline in CD4 count in HIV patients not on antiretroviral therapy. Med. J. Armed Forces India 2014, 70, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Shearer, W.; Quinn, T.C.; LaRussa, P.; Lew, J.F.; Mofenson, L.; Almy, S.; Rich, K.; Handelsman, E.; Diaz, C.; Pagano, M.; et al. Viral load and disease progression in infants infected with human immunodeficiency virus type. N. Engl. J. Med. 1997, 336, 1337–1342. [Google Scholar] [CrossRef]
- Lilic, D.; Cant, A.J.; Abinun, M.; Calvert, J.E.; Spickett, G.P. Cytokine production differs in children and adults. Pediatr. Res. 1997, 42, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Siegrist, C.-A.; Aspinall, R. B-cell responses to vaccination at the extremes of age. Nat. Rev. Immunol. 2009, 9, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Payne, H.; Mkhize, N.N.; Otwombe, K.; Lewis, J.; Panchia, R.; Callard, R.E.; Morris, L.; Babiker, A.; Violari, A.; Cotton, M.F.; et al. Reactivity of routine HIV antibody tests in children who initiated antiretroviral therapy in early infancy as part of the Children with HIV Early Antiretroviral Therapy (CHER) trial: A retrospective analysis. Lancet Infect. Dis. 2015, 15, 803–809. [Google Scholar] [CrossRef]
- Shalekoff, S.; Gray, G.E.; Tiemessen, C.T. Age-related changes in expression of CXCR4 and CCR5 on peripheral blood leukocytes from uninfected infants born to human immunodeficiency virus type 1-infected mothers. Clin. Diagn. Lab. Immunol. 2004, 11, 229–234. [Google Scholar] [CrossRef][Green Version]
- Tuttle, D.L.; Coberley, C.R.; Xie, X.; Kou, Z.C.; Sleasman, J.W.; Goodenow, M.M. Effects of human immunodeficiency virus type 1 infection on CCR5 and CXCR4 coreceptor expression on CD4 T lymphocyte subsets in infants and adolescents. AIDS Res. Hum. Retroviruses 2004, 20, 305–313. [Google Scholar] [CrossRef]
- Shearer, W.; Rosenblatt, H.M.; Gelman, R.S.; Oyomopito, R.; Plaeger, S.; Stiehm, E.; Wara, D.W.; Douglas, S.D.; Luzuriaga, K.; McFarland, E.J.; et al. Lymphocyte subsets in healthy children from birth through 18 years of age: The pediatric aids clinical trials group P1009 study. J. Allergy Clin. Immunol. 2003, 112, 973–980. [Google Scholar] [CrossRef]
- Schatorjé, E.J.; Gemen, E.F.; Driessen, G.J.; Leuvenink, J.; Van Hout, R.W.; De Vries, E. Paediatric reference values for the peripheral T cell compartment. Scand. J. Immunol. 2012, 75, 436–444. [Google Scholar] [CrossRef]
- Lynch, H.E.; Goldberg, G.L.; Chidgey, A.; Van den Brink, M.R.; Boyd, R.; Sempowski, G.D. Thymic involution and immune reconstitution. Trends Immunol. 2009, 30, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.C.; Boechat, M.I.; Belzer, M.; Church, J.A.; De Ville, J.; Nielsen, K.; Weston, S.; Geng, Y.; Dunaway, T.; Kitchen, C.; et al. Thymic volume, T-cell populations, and parameters of thymopoiesis in adolescent and adult survivors of HIV infection acquired in infancy. AIDS 2006, 20, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Franco, J.M.; León-Leal, J.A.; Leal, M.; Cano-Rodriguez, A.; Pineda, J.A.; Macías, J.; Rubio, A.; Rey, C.; Sanchez, B.; Lissen, E. CD4+ and CD8+ T lymphocyte regeneration after anti-retroviral therapy in HIV-1-infected children and adult patients. Clin. Exp. Immunol. 2000, 119, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, S.; Graf, E.H.; Dahl, V.; Strain, M.C.; Yukl, S.A.; Lysenko, E.S.; Bosch, R.J.; Lai, J.; Chioma, S.; Emad, F.; et al. Comparative analysis of measures of viral reservoirs in HIV-1 eradication studies. PLoS Pathog. 2013, 9, e1003174. [Google Scholar] [CrossRef]
- Zerbato, J.M.; Serrao, E.; Lenzi, G.; Kim, B.; Ambrose, Z.; Watkins, S.C.; Engelman, A.N.; Sluis-Cremer, N. Establishment and reversal of HIV-1 latency in naive and central memory CD4+T cells in vitro. J. Virol. 2016, 90, 8059–8073. [Google Scholar] [CrossRef] [PubMed]
- Zerbato, J.M.; McMahon, D.K.; Sobolewski, M.D.; Mellors, J.W.; Sluis-Cremer, N. Naive CD4+ T cells harbor a large inducible reservoir of latent, replication-competent human immunodeficiency virus type. Clin. Infect. Dis. 2019, 69, 1919–1925. [Google Scholar] [CrossRef] [PubMed]
- Rullo, E.V.; Cannon, L.; Pinzone, M.R.; Ceccarelli, M.; Nunnari, G.; O’Doherty, U. Genetic evidence that naive T cells can contribute significantly to the human immunodeficiency virus intact reservoir: Time to re-evaluate their role. Clin. Infect. Dis. 2019, 69, 2236–2237. [Google Scholar] [CrossRef]
- Chomont, N.; El-Far, M.; Ancuta, P.; Trautmann, L.; Procopio, F.A.; Yassine-Diab, B.; Boucher, G.; Boulassel, M.-R.; Ghattas, G.; Brenchley, J.M.; et al. HIV reservoir size and persistence are driven by T cell survival and homeostatic proliferation. Nat. Med. 2009, 15, 893–900. [Google Scholar] [CrossRef]
- Luzuriaga, K.; Tabak, B.; Garber, M.; Chen, Y.H.; Ziemniak, C.; McManus, M.M.; Murray, D.; Strain, M.C.; Richman, D.D.; Chun, T.-W.; et al. HIV type 1 (HIV-1) proviral reservoirs decay continuously under sustained virologic control in HIV-1-infected children who received early treatment. J. Infect. Dis. 2014, 210, 1529–1538. [Google Scholar] [CrossRef]
- Veldsman, K.A.; Maritz, J.; Isaacs, S.; Katusiime, M.G.; Van Rensburg, A.J.; Laughton, B.; Mellors, J.W.; Cotton, M.; van Zyl, G. Rapid decline of HIV-1 DNA and RNA in infants starting very early antiretroviral therapy may pose a diagnostic challenge. AIDS (Lond. Engl.) 2018, 32, 629–634. [Google Scholar] [CrossRef]
- Bachmann, N.; Von Siebenthal, C.; Vongrad, V.; Turk, T.; Neumann, K.; Beerenwinkel, N.; Bogojeska, J.; Fellay, J.; Roth, V.; Kok, Y.L.; et al. Determinants of HIV-1 reservoir size and long-term dynamics during suppressive ART. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef]
- Hill, A.L.; Rosenbloom, D.I.S.; Goldstein, E.; Hanhauser, E.; Kuritzkes, D.R.; Siliciano, R.F.; Henrich, T.J. Real-time predictions of reservoir size and rebound time during antiretroviral therapy interruption trials for HIV. PLoS Pathog. 2016, 12, e1005535. [Google Scholar] [CrossRef] [PubMed]
- Ioannidis, J.P.A.; Tatsioni, A.; Abrams, E.J.; Bulterys, M.; Coombs, R.W.; Goedert, J.J.; Korber, B.T.; Mayaux, M.J.; Mofenson, L.M.; Moye, J.J.; et al. Maternal viral load and rate of disease progression among vertically HIV-1-infected children: An international meta-analysis. AIDS 2004, 18, 99–108. [Google Scholar] [CrossRef]
- Abrams, E.J.; Wiener, J.; Carter, R.; Kuhn, L.; Palumbo, P.; Nesheim, S.; Lee, F.; Vink, P.; Bulterys, M. Maternal health factors and early pediatric antiretroviral therapy influence the rate of perinatal HIV-1 disease progression in children. AIDS 2003, 17, 867–877. [Google Scholar] [CrossRef] [PubMed]
- Obimbo, E.M.; Wamalwa, D.; Richardson, B.; Mbori-Ngacha, D.; Overbaugh, J.; Emery, S.; Otieno, P.; Farquhar, C.; Bosire, R.; Payne, B.L.; et al. Pediatric HIV-1 in Kenya: Pattern and correlates of viral load and association with mortality. JAIDS J. Acquir. Immune Defic. Syndr. 2009, 51, 209–215. [Google Scholar] [CrossRef]
- Russell, E.S.; Kwiek, J.J.; Keys, J.; Barton, K.; Mwapasa, V.; Montefiori, D.C.; Meshnick, S.R.; Swanstrom, R. The genetic bottleneck in vertical transmission of subtype C HIV-1 is not driven by selection of especially neutralization-resistant virus from the maternal viral population. J. Virol. 2011, 85, 8253–8262. [Google Scholar] [CrossRef] [PubMed]
- Derdeyn, C.A.; Decker, J.M.; Bibollet-Ruche, F.; Mokili, J.L.; Muldoon, M.; Denham, S.A.; Heil, M.L.; Kasolo, F.; Musonda, R.; Hahn, B.H.; et al. Envelope-constrained neutralization-sensitive HIV-1 after heterosexual transmission. Science 2004, 303, 2019–2022. [Google Scholar] [CrossRef]
- Kishko, M.; Somasundaran, M.; Brewster, F.; Sullivan, J.L.; Clapham, P.R.; Luzuriaga, K. Genotypic and functional properties of early infant HIV-1 envelopes. Retrovirology 2011, 8, 67. [Google Scholar] [CrossRef]
- Kumar, A.; Smith, C.E.P.; Giorgi, E.E.; Eudailey, J.; Martinez, D.R.; Yusim, K.; Douglas, A.O.; Stamper, L.; McGuire, E.; Labranche, C.C.; et al. Infant transmitted/founder HIV-1 viruses from peripartum transmission are neutralization resistant to paired maternal plasma. PLoS Pathog. 2018, 14, e1006944. [Google Scholar] [CrossRef] [PubMed]
- Martinez, D.R.; Tu, J.; Kumar, A.; Mangold, J.F.; Mangan, R.J.; Goswami, R.; Giorgi, E.E.; Chen, J.; Mengual, M.; Douglas, A.O.; et al. Maternal broadly neutralizing antibodies can select for neutralization-resistant, infant-transmitted/founder HIV variants. mBio 2020, 11, e00176-20. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Parast, A.B.; Richardson, B.A.; Nduati, R.; John-Stewart, G.; Mbori-Ngacha, D.; Rainwater, S.M.J.; Overbaugh, J. Neutralization escape variants of human immunodeficiency virus type 1 are transmitted from mother to infant. J. Virol. 2006, 80, 835–844. [Google Scholar] [CrossRef]
- Ananworanich, J.; Puthanakit, T.; Suntarattiwong, P.; Chokephaibulkit, K.; Kerr, S.J.; Fromentin, R.; Bakeman, W.; Intasan, J.; Mahanontharit, A.; Sirivichayakul, S.; et al. Reduced markers of HIV persistence and restricted HIV-specific immune responses after early antiretroviral therapy in children. AIDS 2014, 28, 1015–1020. [Google Scholar] [CrossRef]
- Lilian, R.R.; Kalk, E.; Technau, K.-G.; Sherman, G.G. Birth diagnosis of HIV infection in infants to reduce infant mortality and monitor for elimination of mother-to-child transmission. Pediatr. Infect. Dis. J. 2013, 32, 1080–1085. [Google Scholar] [CrossRef]
- Cotton, M.F.; Violari, A.; Otwombe, K.; Panchia, R.; Dobbels, E.; Rabie, H.; Josipovic, D.; Liberty, A.; Lazarus, E.; Innes, S.; et al. Early time-limited antiretroviral therapy versus deferred therapy in South African infants infected with HIV: Results from the children with HIV early antiretroviral (CHER) randomised trial. Lancet 2013, 382, 1555–1563. [Google Scholar] [CrossRef]
- Van Zyl, G.; Bedison, M.A.; Van Rensburg, A.J.; Laughton, B.; Cotton, M.F.; Mellors, J.W. Early antiretroviral therapy in South African children reduces HIV-1-infected cells and cell-associated HIV-1 RNA in blood mononuclear cells. J. Infect. Dis. 2015, 212, 39–43. [Google Scholar] [CrossRef]
- De Oliveira, M.F.; Gianella, S.; Letendre, S.; Scheffler, K.; Pond, S.L.K.; Smith, D.M.; Strain, M.; Ellis, R.J. Comparative analysis of cell-associated HIV DNA levels in cerebrospinal fluid and peripheral blood by droplet digital PCR. PLoS ONE 2015, 10, e0139510. [Google Scholar] [CrossRef]
- Henrich, T.J.; Gallien, S.; Li, J.Z.; Pereyra, F.; Kuritzkes, D.R. Low-level detection and quantitation of cellular HIV-1 DNA and 2-LTR circles using droplet digital PCR. J. Virol. Methods 2012, 186, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Kiselinova, M.; Pasternak, A.; De Spiegelaere, W.; Vogelaers, D.; Berkhout, B.; Vandekerckhove, L. Comparison of droplet digital PCR and seminested real-time PCR for quantification of cell-associated HIV-1 RNA. PLoS ONE 2014, 9, e85999. [Google Scholar] [CrossRef]
- Strain, M.C.; Lada, S.M.; Luong, T.; Rought, S.E.; Gianella, S.; Terry, V.H.; Spina, C.A.; Woelk, C.H.; Richman, D.D. Highly precise measurement of HIV DNA by droplet digital PCR. PLoS ONE 2013, 8, e55943. [Google Scholar] [CrossRef] [PubMed]
- Archin, N.M.; Vaidya, N.K.; Kuruc, J.D.; Liberty, A.L.; Wiegand, A.; Kearney, M.F.; Cohen, M.S.; Coffin, J.M.; Bosch, R.J.; Gay, C.; et al. Immediate antiviral therapy appears to restrict resting CD4+ cell HIV-1 infection without accelerating the decay of latent infection. Proc. Natl. Acad. Sci. USA 2012, 109, 9523–9528. [Google Scholar] [CrossRef]
- Martínez-Bonet, M.; Puertas, M.C.; Fortuny, C.; Ouchi, D.; Mellado, M.J.; Rojo, P.; Noguera-Julian, A.; Muñoz-Fernández, M.A.; Martinez-Picado, J. Establishment and replenishment of the viral reservoir in perinatally HIV-1-infected children initiating very early antiretroviral therapy. Clin. Infect. Dis. 2015, 61, 1169–1178. [Google Scholar] [CrossRef]
- Foster, C.; Pace, M.; Kaye, S.; Hopkins, E.; Jones, M.; Robinson, N.; Mant, C.; Cason, J.; Fidler, S.; Frater, J. Early antiretroviral therapy reduces HIV DNA following perinatal HIV infection. AIDS 2017, 31, 1847–1851. [Google Scholar] [CrossRef]
- Van Zyl, G.U.; Katusiime, M.G.; Wiegand, A.; McManus, W.R.; Bale, M.; Halvas, E.K.; Luke, B.; Boltz, V.F.; Spindler, J.; Laughton, B.; et al. No evidence of HIV replication in children on antiretroviral therapy. J. Clin. Investig. 2017, 127, 3827–3834. [Google Scholar] [CrossRef] [PubMed]
- Persaud, D.; Patel, K.; Karalius, B.; Rainwater-Lovett, K.; Ziemniak, C.; Ellis, A.; Chen, Y.H.; Richman, U.; Siberry, G.K.; Van Dyke, R.B.; et al. Influence of age at virologic control on peripheral blood human immunodeficiency virus reservoir size and serostatus in perinatally infected adolescents. JAMA Pediatr. 2014, 168, 1138–1146. [Google Scholar] [CrossRef]
- Luzuriaga, K. Early combination antiretroviral therapy limits HIV-1 persistence in children. Annu. Rev. Med. 2016, 67, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, L.; Paximadis, M.; Da Costa Dias, B.; Loubser, S.; Strehlau, R.; Patel, F.; Shiau, S.; Coovadia, A.; Abrams, E.J.; Tiemessen, C.T. Age at antiretroviral therapy initiation and cell-associated HIV-1 DNA levels in HIV-1-infected children. PLoS ONE 2018, 13, e0195514. [Google Scholar] [CrossRef]
- Tagarro, A.; Chan, M.; Zangari, P.; Ferns, B.; Foster, C.; De Rossi, A.; Nastouli, E.; Muñoz-Fernández, M.A.; Gibb, D.; Rossi, P.; et al. Early and highly suppressive antiretroviral therapy are main factors associated with low viral reservoir in European perinatally HIV-infected children. JAIDS J. Acquir. Immune Defic. Syndr. 2018, 79, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Ajibola, G.; Garcia-Broncano, P.; Maswabi, K.; Bennett, K.; Hughes, M.D.; Moyo, S.; Mohammed, T.; Jean-Philippe, P.; Sakoi, M.; Batlang, O.; et al. Viral reservoir in early-treated HIV-infected children and markers for sustained viral suppression. Clin. Infect. Dis. 2021. [Google Scholar] [CrossRef] [PubMed]
- Massanella, M.; Puthanakit, T.; Leyre, L.; Jupimai, T.; Sawangsinth, P.; De Souza, M.; Suntarattiwong, P.; Kosalarksa, P.; Borkird, T.; Kanjanavanit, S.; et al. Continuous prophylactic antiretrovirals/antiretroviral therapy since birth reduces seeding and persistence of the viral reservoir in children vertically infected with human immunodeficiency virus. Clin. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- McManus, M.; Mick, E.; Hudson, R.; Mofenson, L.M.; Sullivan, J.L.; Somasundaran, M.; Luzuriaga, K. PACTG 356 investigators early combination antiretroviral therapy limits exposure to HIV-1 replication and cell-associated HIV-1 DNA levels in infants. PLoS ONE 2016, 11, e0154391. [Google Scholar] [CrossRef]
- Persaud, D.; Palumbo, P.E.; Ziemniak, C.; Hughes, M.D.; Alvero, C.G.; Luzuriaga, K.; Yogev, R.; Capparelli, E.V.; Chadwick, E.G. Dynamics of the resting CD4+ T-cell latent HIV reservoir in infants initiating HAART less than 6 months of age. AIDS 2012, 26, 1483–1490. [Google Scholar] [CrossRef]
- Ho, Y.-C.; Shan, L.; Hosmane, N.N.; Wang, J.; Laskey, S.B.; Rosenbloom, D.I.; Lai, J.; Blankson, J.N.; Siliciano, J.D.; Siliciano, R.F. Replication-competent noninduced proviruses in the latent reservoir increase barrier to HIV-1 cure. Cell 2013, 155, 540–551. [Google Scholar] [CrossRef]
- Siliciano, J.D.; Siliciano, R.F. Enhanced culture assay for detection and quantitation of latently infected, resting CD4+ T-cells carrying replication-competent virus in HIV-1-infected individuals. Hum. Retrovir. Protoc. 2005, 304, 003–016. [Google Scholar] [CrossRef]
- Rosenbloom, D.I.S.; Elliott, O.; Hill, A.L.; Henrich, T.J.; Siliciano, J.M.; Siliciano, R.F. Designing and interpreting limiting dilution assays: General principles and applications to the latent reservoir for human immunodeficiency virus-1. Open Forum Infect. Dis. 2015, 2, ofv123. [Google Scholar] [CrossRef] [PubMed]
- Bruner, K.M.; Wang, Z.; Simonetti, F.R.; Bender, A.M.; Kwon, K.J.; Sengupta, S.; Fray, E.J.; Beg, S.A.; Antar, A.; Jenike, K.M.; et al. A quantitative approach for measuring the reservoir of latent HIV-1 proviruses. Nat. Cell Biol. 2019, 566, 120–125. [Google Scholar] [CrossRef]
- Chun, T.-W.; Carruth, L.; Finzi, D.; Shen, X.; DiGiuseppe, J.A.; Taylor, H.; Hermankova, M.; Chadwick, K.; Margolick, J.; Quinn, T.C.; et al. Quantification of latent tissue reservoirs and total body viral load in HIV-1 infection. Nat. Cell Biol. 1997, 387, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Bruner, K.M.; Murray, A.J.; Pollack, R.A.; Soliman, M.G.; Laskey, S.B.; Capoferri, A.A.; Lai, J.; Strain, M.C.; Lada, S.M.; Hoh, R.; et al. Defective proviruses rapidly accumulate during acute HIV-1 infection. Nat. Med. 2016, 22, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Hosmane, N.N.; Kwon, K.J.; Bruner, K.M.; Capoferri, A.A.; Beg, S.; Rosenbloom, D.I.; Keele, B.F.; Ho, Y.-C.; Siliciano, J.D.; Siliciano, R.F. Proliferation of latently infected CD4+ T cells carrying replication-competent HIV-1: Potential role in latent reservoir dynamics. J. Exp. Med. 2017, 214, 959–972. [Google Scholar] [CrossRef] [PubMed]
- Rainwater-Lovett, K.; Ziemniak, C.; Watson, D.; Luzuriaga, K.; Siberry, G.; Petru, A.; Chen, Y.; Uprety, P.; McManus, M.; Ho, Y.-C.; et al. Paucity of intact non-induced provirus with early, long-term antiretroviral therapy of perinatal HIV infection. PLoS ONE 2017, 12, e0170548. [Google Scholar] [CrossRef]
- Dhummakupt, A.; Rubens, J.H.; Anderson, T.; Powell, L.; Nonyane, B.A.; Siems, L.V.; Collinson-Streng, A.; Nilles, T.; Jones, R.B.; Tepper, V.; et al. Differences in inducibility of the latent HIV reservoir in perinatal and adult infection. JCI Insight 2020, 5, e134105. [Google Scholar] [CrossRef]
- Hill, A.L.; Rosenbloom, D.I.S.; Fu, F.; Nowak, M.A.; Siliciano, R.F. Predicting the outcomes of treatment to eradicate the latent reservoir for HIV. Proc. Natl. Acad. Sci. USA 2014, 111, 13475–13480. [Google Scholar] [CrossRef]
- Persaud, D.; Gay, H.; Ziemniak, C.; Chen, Y.H.; Piatak, M.; Chun, T.-W.; Strain, M.; Richman, D.; Luzuriaga, K. Absence of detectable HIV-1 viremia after treatment cessation in an infant. N. Engl. J. Med. 2013, 369, 1828–1835. [Google Scholar] [CrossRef]
- Luzuriaga, K.; Gay, H.; Ziemniak, C.; Sanborn, K.B.; Somasundaran, M.; Rainwater-Lovett, K.; Mellors, J.W.; Rosenbloom, D.I.; Persaud, D. Viremic relapse after HIV-1 remission in a perinatally infected child. N. Engl. J. Med. 2015, 372, 786–788. [Google Scholar] [CrossRef]
- Frange, P.; Faye, A.; Avettand-Fenoël, V.; Bellaton, E.; Descamps, D.; Angin, M.; David, A.; Caillat-Zucman, S.; Peytavin, G.; Dollfus, C.; et al. HIV-1 virological remission lasting more than 12 years after interruption of early antiretroviral therapy in a perinatally infected teenager enrolled in the French ANRS EPF-CO10 paediatric cohort: A case report. Lancet HIV 2016, 3, e49–e54. [Google Scholar] [CrossRef]
- Violari, A.; Cotton, M.F.; Kuhn, L.; Schramm, D.B.; Paximadis, M.; Loubser, S.; Shalekoff, S.; Da Costa Dias, B.; Otwombe, K.; Liberty, A.; et al. A child with perinatal HIV infection and long-term sustained virological control following antiretroviral treatment cessation. Nat. Commun. 2019, 10, 412. [Google Scholar] [CrossRef] [PubMed]
- Colby, D.J.; Trautmann, L.; Pinyakorn, S.; Leyre, L.; Pagliuzza, A.; Kroon, E.; Rolland, M.; Takata, H.; Buranapraditkun, S.; Intasan, J.; et al. Rapid HIV RNA rebound after antiretroviral treatment interruption in persons durably suppressed in Fiebig I acute HIV infection. Nat. Med. 2018, 24, 923–926. [Google Scholar] [CrossRef] [PubMed]
- Sáez-Cirión, A.; Bacchus, C.; Hocqueloux, L.; Avettand-Fenoel, V.; Girault, I.; Lecuroux, C.; Potard, V.; Versmisse, P.; Melard, A.; Prazuck, T.; et al. Post-treatment HIV-1 controllers with a long-term virological remission after the interruption of early initiated antiretroviral therapy ANRS VISCONTI study. PLoS Pathog. 2013, 9, e1003211. [Google Scholar] [CrossRef]
- Trautmann, L. How to generate good killers by initiating ART (not too) early? In Proceedings of the Conference on Retroviruses and Oppourtunisitc Infections, Virtual, 6–10 March 2021; Available online: https://www.croiconference.org/abstract/how-to-generate-good-killers-by-initiating-art-not-too-early/ (accessed on 12 March 2021).
- Wagner, T.A.; McKernan, J.L.; Tobin, N.H.; Tapia, K.A.; Mullins, J.I.; Frenkel, L.M. An increasing proportion of monotypic HIV-1 DNA sequences during antiretroviral treatment suggests proliferation of HIV-infected cells. J. Virol. 2012, 87, 1770–1778. [Google Scholar] [CrossRef] [PubMed]
- Castro-Nallar, E.; Pérez-Losada, M.; Burton, G.F.; Crandall, K.A. The evolution of HIV: Inferences using phylogenetics. Mol. Phylogenetics Evol. 2012, 62, 777–792. [Google Scholar] [CrossRef]
- Hammer, S.M.; Ribaudo, H.; Bassett, R.; Mellors, J.W.; Demeter, L.M.; Coombs, R.W.; Currier, J.; Morse, G.D.; Gerber, J.G.; Martinez, A.I.; et al. A randomized, placebo-controlled trial of abacavir intensification in HIV-1-infected adults with virologic suppression on a protease inhibitor-containing regimen. HIV Clin. Trials 2010, 11, 312–324. [Google Scholar] [CrossRef]
- Persaud, D.; Ray, S.C.; Kajdas, J.; Ahonkhai, A.; Siberry, G.K.; Ferguson, K.; Ziemniak, C.; Quinn, T.C.; Casazza, J.P.; Zeichner, S.; et al. Slow human immunodeficiency virus type 1 evolution in viral reservoirs in infants treated with effective antiretroviral therapy. AIDS Res. Hum. Retrovir. 2007, 23, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Van Zyl, G.; Bale, M.J.; Kearney, M.F. HIV evolution and diversity in ART-treated patients. Retrovirology 2018, 15, 1–12. [Google Scholar] [CrossRef]
- Sanchez-Merino, V.; Nie, S.; Luzuriaga, K. HIV-1-specific CD8+T cell responses and viral evolution in women and infants. J. Immunol. 2005, 175, 6976–6986. [Google Scholar] [CrossRef]
- Kearney, M.F.; Wiegand, A.; Shao, W.; Coffin, J.M.; Mellors, J.W.; Lederman, M.; Gandhi, R.T.; Keele, B.F.; Li, J.Z. Origin of rebound plasma HIV includes cells with identical proviruses that are transcriptionally active before stopping of antiretroviral therapy. J. Virol. 2016, 90, 1369–1376. [Google Scholar] [CrossRef]
- Kearney, M.F.; Spindler, J.; Shao, W.; Yu, S.; Anderson, E.M.; O’Shea, A.; Rehm, C.; Poethke, C.; Kovacs, N.; Mellors, J.W.; et al. Lack of detectable HIV-1 molecular evolution during suppressive antiretroviral therapy. PLoS Pathog. 2014, 10, e1004010. [Google Scholar] [CrossRef]
- Veldsman, K.A.; Van Rensburg, A.J.; Isaacs, S.; Naidoo, S.; Laughton, B.; Lombard, C.; Cotton, M.F.; Mellors, J.W.; Van Zyl, G.U. HIV-1DNA decay is faster in children who initiate ART shortly after birth than later. J. Int. AIDS Soc. 2019, 22, e25368. [Google Scholar] [CrossRef] [PubMed]
- Uprety, P.; Chadwick, E.G.; Rainwater-Lovett, K.; Ziemniak, C.; Luzuriaga, K.; Capparelli, E.V.; Yenokyan, G.; Persaud, D. Cell-associated HIV-1 DNA and RNA decay dynamics during early combination antiretroviral therapy in HIV-1-infected infants. Clin. Infect. Dis. 2015, 61, 1862–1870. [Google Scholar] [CrossRef]
- Ananworanich, J.; Chomont, N.; Eller, L.A.; Kroon, E.; Tovanabutra, S.; Bose, M.; Nau, M.; Fletcher, J.L.; Tipsuk, S.; Vandergeeten, C.; et al. HIV DNA set point is rapidly established in acute HIV infection and dramatically reduced by early ART. EBioMedicine 2016, 11, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Besson, G.J.; Lalama, C.M.; Bosch, R.J.; Gandhi, R.T.; Bedison, M.A.; Aga, E.; Riddler, S.A.; McMahon, D.K.; Hong, F.; Mellors, J.W. HIV-1 DNA decay dynamics in blood during more than a decade of suppressive antiretroviral therapy. Clin. Infect. Dis. 2014, 59, 1312–1321. [Google Scholar] [CrossRef] [PubMed]
- Buzon, M.J.; Martin-Gayo, E.; Pereyra, F.; Ouyang, Z.; Sun, H.; Li, J.Z.; Piovoso, M.; Shaw, A.; Dalmau, J.; Zangger, N.; et al. Long-term antiretroviral treatment initiated at primary HIV-1 infection affects the size, composition, and decay kinetics of the reservoir of HIV-1-infected CD4 T cells. J. Virol. 2014, 88, 10056–10065. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.; Joos, B.; Niederöst, B.; Kaiser, P.; Hafner, R.; Von Wyl, V.; Ackermann, M.; Weber, R.; Günthard, H.F. Biphasic decay kinetics suggest progressive slowing in turnover of latently HIV-1 infected cells during antiretroviral therapy. Retrovirology 2008, 5, 107. [Google Scholar] [CrossRef] [PubMed]
- Crooks, A.M.; Bateson, R.; Cope, A.B.; Dahl, N.P.; Griggs, M.K.; Kuruc, J.D.; Gay, C.; Eron, J.J.; Margolis, D.M.; Bosch, R.J.; et al. Precise quantitation of the latent HIV-1 reservoir: Implications for eradication strategies. J. Infect. Dis. 2015, 212, 1361–1365. [Google Scholar] [CrossRef]
- Siliciano, J.D.; Kajdas, J.; Finzi, D.; Quinn, T.C.; Chadwick, K.; Margolick, J.B.; Kovacs, C.; Gange, S.; Siliciano, R.F. Long-term follow-up studies confirm the stability of the latent reservoir for HIV-1 in resting CD4+ T cells. Nat. Med. 2003, 9, 727–728. [Google Scholar] [CrossRef]
- Antar, A.; Jenike, K.M.; Jang, S.; Rigau, D.N.; Reeves, D.B.; Hoh, R.; Krone, M.R.; Keruly, J.C.; Moore, R.D.; Schiffer, J.T.; et al. Longitudinal study reveals HIV-1-infected CD4+ T cell dynamics during long-term antiretroviral therapy. J. Clin. Investig. 2020, 130, 3543–3559. [Google Scholar] [CrossRef]
- Peluso, M.J.; Bacchetti, P.; Ritter, K.D.; Beg, S.; Lai, J.; Martin, J.N.; Hunt, P.W.; Henrich, T.J.; Siliciano, J.D.; Siliciano, R.F.; et al. Differential decay of intact and defective proviral DNA in HIV-1-infected individuals on suppressive antiretroviral therapy. JCI Insight 2020, 5. [Google Scholar] [CrossRef]
- Garcia-Broncano, P.; Maddali, S.; Einkauf, K.B.; Jiang, C.; Gao, C.; Chevalier, J.; Chowdhury, F.Z.; Maswabi, K.; Ajibola, G.; Moyo, S.; et al. Early antiretroviral therapy in neonates with HIV-1 infection restricts viral reservoir size and induces a distinct innate immune profile. Sci. Transl. Med. 2019, 11, eaax7350. [Google Scholar] [CrossRef] [PubMed]
- Hiener, B.; Horsburgh, B.A.; Eden, J.-S.; Barton, K.; Schlub, T.E.; Lee, E.; von Stockenstrom, S.; Odevall, L.; Milush, J.M.; Liegler, T.; et al. Identification of genetically intact HIV-1 proviruses in specific CD4+ T cells from effectively treated participants. Cell Rep. 2017, 21, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Bebenek, K.; Abbotts, J.; Roberts, J.D.; Wilson, S.; Kunkel, T.A. Specificity and mechanism of error-prone replication by human immunodeficiency virus-1 reverse transcriptase. J. Biol. Chem. 1989, 264, 16948–16956. [Google Scholar] [CrossRef]
- Kieffer, T.L.; Kwon, P.; Nettles, R.E.; Han, Y.; Ray, S.C.; Siliciano, R.F.G. A hypermutation in protease and reverse transcriptase regions of human immunodeficiency virus type 1 residing in resting CD4+ T cells in vivo. J. Virol. 2005, 79, 1975–1980. [Google Scholar] [CrossRef]
- Sanchez, G.; Xu, X.; Chermann, J.C.; Hirsch, I. Accumulation of defective viral genomes in peripheral blood mononuclear cells of human immunodeficiency virus type 1-infected individuals. J. Virol. 1997, 71, 2233–2240. [Google Scholar] [CrossRef]
- Simonetti, F.R.; White, J.A.; Tumiotto, C.; Ritter, K.D.; Cai, M.; Gandhi, R.T.; Deeks, S.G.; Howell, B.J.; Montaner, L.J.; Blankson, J.N.; et al. Intact proviral DNA assay analysis of large cohorts of people with HIV provides a benchmark for the frequency and composition of persistent proviral DNA. Proc. Natl. Acad. Sci. USA 2020, 117, 18692–18700. [Google Scholar] [CrossRef]
- Katusiime, M.G.; Halvas, E.K.; Wright, I.; Joseph, K.; Bale, M.J.; Kirby-McCullough, B.; Engelbrecht, S.; Shao, W.; Hu, W.-S.; Cotton, M.F.; et al. Intact HIV proviruses persist in children seven to nine years after initiation of antiretroviral therapy in the first year of life. J. Virol. 2020, 94, e01519-19. [Google Scholar] [CrossRef]
- Anderson, J.A.; Archin, N.M.; Ince, W.; Parker, D.; Wiegand, A.; Coffin, J.M.; Kuruc, J.; Eron, J.; Swanstrom, R.; Margolis, D.M. Clonal sequences recovered from plasma from patients with residual HIV-1 viremia and on intensified antiretroviral therapy are identical to replicating viral RNAs recovered from circulating resting CD4+ T cells. J. Virol. 2011, 85, 5220–5223. [Google Scholar] [CrossRef]
- Bailey, J.R.; Sedaghat, A.R.; Kieffer, T.; Brennan, T.; Lee, P.K.; Wind-Rotolo, M.; Haggerty, C.M.; Kamireddi, A.R.; Liu, Y.; Lee, J.; et al. Residual human immunodeficiency virus type 1 viremia in some patients on antiretroviral therapy is dominated by a small number of invariant clones rarely found in circulating CD4+ T cells. J. Virol. 2006, 80, 6441–6457. [Google Scholar] [CrossRef]
- Evering, T.H.; Mehandru, S.; Racz, P.; Tenner-Racz, K.; Poles, M.; Figueroa, A.; Mohri, H.; Markowitz, M. Absence of HIV-1 evolution in the gut-associated lymphoid tissue from patients on combination antiviral therapy initiated during primary infection. PLoS Pathog. 2012, 8, e1002506. [Google Scholar] [CrossRef] [PubMed]
- Bull, M.; Learn, G.; Genowati, I.; McKernan, J.; Hitti, J.; Lockhart, D.; Tapia, K.; Holte, S.; Dragavon, J.; Coombs, R.; et al. Compartmentalization of HIV-1 within the female genital tract is due to monotypic and low-diversity variants not distinct viral populations. PLoS ONE 2009, 4, e7122. [Google Scholar] [CrossRef] [PubMed]
- Wagner, T.A.; McLaughlin, S.; Garg, K.; Cheung, C.Y.K.; Larsen, B.B.; Styrchak, S.; Huang, H.C.; Edlefsen, P.T.; Mullins, J.I.; Frenkel, L.M. Proliferation of cells with HIV integrated into cancer genes contributes to persistent infection. Science 2014, 345, 570–573. [Google Scholar] [CrossRef]
- Patro, S.C.; Brandt, L.D.; Bale, M.; Halvas, E.K.; Joseph, K.W.; Shao, W.; Wu, X.; Guo, S.; Murrell, B.; Wiegand, A.; et al. Combined HIV-1 sequence and integration site analysis informs viral dynamics and allows reconstruction of replicating viral ancestors. Proc. Natl. Acad. Sci. USA 2019, 116, 25891–25899. [Google Scholar] [CrossRef] [PubMed]
- Simonetti, F.R.; Sobolewski, M.D.; Fyne, E.; Shao, W.; Spindler, J.; Hattori, J.; Anderson, E.M.; Watters, S.A.; Hill, S.; Wu, X.; et al. Clonally expanded CD4+ T cells can produce infectious HIV-1 in vivo. Proc. Natl. Acad. Sci. USA 2016, 113, 1883–1888. [Google Scholar] [CrossRef]
- Einkauf, K.B.; Lee, G.Q.; Gao, C.; Sharaf, R.; Sun, X.; Hua, S.; Chen, S.M.; Jiang, C.; Lian, X.; Chowdhury, F.Z.; et al. Intact HIV-1 proviruses accumulate at distinct chromosomal positions during prolonged antiretroviral therapy. J. Clin. Investig. 2019, 129, 988–998. [Google Scholar] [CrossRef]
- Maldarelli, F.; Wu, X.; Su, L.; Simonetti, F.R.; Shao, W.; Hill, S.; Spindler, J.; Ferris, A.L.; Mellors, J.W.; Kearney, M.F.; et al. Specific HIV integration sites are linked to clonal expansion and persistence of infected cells. Research Support, N.I.H.; Extramural Research Support, Non-U.S. Gov’t. Science 2014, 345, 179–183. [Google Scholar] [CrossRef]
- Bui, J.K.; Sobolewski, M.D.; Keele, B.F.; Spindler, J.; Musick, A.; Wiegand, A.; Luke, B.T.; Shao, W.; Hughes, S.H.; Coffin, J.M.; et al. Proviruses with identical sequences comprise a large fraction of the replication-competent HIV reservoir. PLoS Pathog. 2017, 13, e1006283. [Google Scholar] [CrossRef]
- Halvas, E.K.; Joseph, K.W.; Brandt, L.D.; Guo, S.; Sobolewski, M.D.; Jacobs, J.L.; Tumiotto, C.; Bui, J.K.; Cyktor, J.C.; Keele, B.F.; et al. HIV-1 viremia not suppressible by antiretroviral therapy can originate from large T cell clones producing infectious virus. J. Clin. Investig. 2020, 130, 5847–5857. [Google Scholar] [CrossRef]
- Lorenzi, J.C.C.; Cohen, Y.Z.; Cohn, L.B.; Kreider, E.; Barton, J.P.; Learn, G.H.; Oliveira, T.; Lavine, C.L.; Horwitz, J.A.; Settler, A.; et al. Paired quantitative and qualitative assessment of the replication-competent HIV-1 reservoir and comparison with integrated proviral DNA. Proc. Natl. Acad. Sci. USA 2016, 113, E7908–E7916. [Google Scholar] [CrossRef]
- Cohn, L.B.; Silva, I.T.; Oliveira, T.Y.; Rosales, R.A.; Parrish, E.H.; Learn, G.H.; Hahn, B.H.; Czartoski, J.L.; McElrath, M.J.; Lehmann, C.; et al. HIV-1 integration landscape during latent and active infection. Cell 2015, 160, 420–432. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Siliciano, R.F. Reservoir expansion by T-cell proliferation may be another barrier to curing HIV infection. Proc. Natl. Acad. Sci. USA 2016, 113, 1692–1694. [Google Scholar] [CrossRef]
- Pinzone, M.R.; VanBelzen, D.J.; Weissman, S.; Bertuccio, M.P.; Cannon, L.; Hwang, W.-T.; Sherman, B.; O’Doherty, U. Proviral sequencing suggests the majority of the HIV reservoir is expressed over time but significant decay is obscured by clonal expansion. bioRxiv 2018, 348409. [Google Scholar] [CrossRef]
- Bale, M.J.; Katusiime, M.G.; Wells, D.; Wu, X.; Spindler, J.; Halvas, E.K.; Cyktor, J.C.; Wiegand, A.; Shao, W.; Cotton, M.F.; et al. Early emergence and long-term persistence of HIV-infected T-cell clones in children. mBio 2021, 12, e00568-21. [Google Scholar] [CrossRef] [PubMed]
- Coffin, J.M.; Wells, D.W.; Zerbato, J.M.; Kuruc, J.D.; Guo, S.; Luke, B.T.; Eron, J.J.; Bale, M.; Spindler, J.; Simonetti, F.R.; et al. Clones of infected cells arise early in HIV-infected individuals. JCI Insight 2019, 4, e128432. [Google Scholar] [CrossRef]
- Boshier, F.A.T.; Reeves, D.B.; Duke, E.R.; Swan, D.A.; Prlic, M.; Cardozo-Ojeda, E.F.; Schiffer, J.T. Blind uneven proliferation of CD4+ T cells during primary infection generates the majority of the HIV reservoir. medRxiv 2020. [Google Scholar] [CrossRef]
- Cesana, D.; De Sio, F.R.S.; Rudilosso, L.; Gallina, P.; Calabria, A.; Beretta, S.; Merelli, I.; Bruzzesi, E.; Passerini, L.; Nozza, S.; et al. HIV-1-mediated insertional activation of STAT5B and BACH2 trigger viral reservoir in T regulatory cells. Nat. Commun. 2017, 8, 1–11. [Google Scholar] [CrossRef]
- Ikeda, T.; Shibata, J.; Yoshimura, K.; Koito, A.; Matsushita, S. Recurrent HIV-1 integration at the BACH2 locus in resting CD4+ T cell populations during effective highly active antiretroviral therapy. J. Infect. Dis. 2007, 195, 716–725. [Google Scholar] [CrossRef]
- Wang, Z.; Gurule, E.E.; Brennan, T.P.; Gerold, J.M.; Kwon, K.J.; Hosmane, N.N.; Kumar, M.R.; Beg, S.A.; Capoferri, A.A.; Ray, S.C.; et al. Expanded cellular clones carrying replication-competent HIV-1 persist, wax, and wane. Proc. Natl. Acad. Sci. USA 2018, 115, E2575–E2584. [Google Scholar] [CrossRef]
- Simonetti, F.R.; Zhang, H.; Soroosh, G.P.; Duan, J.; Rhodehouse, K.; Hill, A.L.; Beg, S.A.; McCormick, K.; Raymond, H.E.; Nobles, C.L.; et al. Antigen-driven clonal selection shapes the persistence of HIV-1-infected CD4+ T cells in vivo. J. Clin. Investig. 2021, 131. [Google Scholar] [CrossRef]
- Mendoza, P.; Jackson, J.R.; Oliveira, T.Y.; Gaebler, C.; Ramos, V.; Caskey, M.; Jankovic, M.; Nussenzweig, M.C.; Cohn, L.B. Antigen-responsive CD4+ T cell clones contribute to the HIV-1 latent reservoir. J. Exp. Med. 2020, 217. [Google Scholar] [CrossRef]
- Miles, D.J.C.; van der Sande, M.; Jeffries, D.; Kaye, S.; Ismaili, J.; Ojuola, O.; Sanneh, M.; Touray, E.S.; Waight, P.; Rowland-Jones, S.; et al. Cytomegalovirus infection in gambian infants leads to profound CD8 T-cell differentiation. J. Virol. 2007, 81, 5766–5776. [Google Scholar] [CrossRef]
- Yindom, L.-M.; Simms, V.; Majonga, E.D.; McHugh, G.; Dauya, E.; Bandason, T.; Vincon, H.; Rylance, J.; Munyati, S.; Ferrand, R.A.; et al. Unexpectedly high prevalence of cytomegalovirus DNA emia in older children and adolescents with perinatally acquired human immunodeficiency virus infection. Clin. Infect. Dis. 2019, 69, 580–587. [Google Scholar] [CrossRef]
- Horvat, T.; Adel, N.G.; Dang, T.-O.; Momtaz, P.; Postow, M.A.; Callahan, M.K.; Carvajal, R.D.; Dickson, M.A.; D’Angelo, S.P.; Woo, K.M.; et al. Immune-related adverse events, need for systemic immunosuppression, and effects on survival and time to treatment failure in patients with melanoma treated with ipilimumab at memorial sloan kettering cancer center. J. Clin. Oncol. 2015, 33, 3193–3198. [Google Scholar] [CrossRef]
- Cotugno, N.; Douagi, I.; Rossi, P.; Palma, P. Suboptimal immune reconstitution in vertically HIV infected children: A view on how HIV replication and timing of HAART initiation can impact on T and B-cell compartment. Clin. Dev. Immunol. 2012, 2012, 805151. [Google Scholar] [CrossRef]
- Bekker, V.; Westerlaken, G.H.A.; Scherpbier, H.J.; Alders, S.; Zaaijer, H.L.; Van Baarle, D.; Kuijpers, T. Varicella vaccination in HIV-1-infected children after immune reconstitution. AIDS 2006, 20, 2321–2329. [Google Scholar] [CrossRef] [PubMed]
- Gasper, M.A.; Hesseling, A.C.; Mohar, I.; Myer, L.; Azenkot, T.; Passmore, J.-A.S.; Hanekom, W.; Cotton, M.F.; Crispe, I.N.; Sodora, N.L.; et al. BCG vaccination induces HIV target cell activation in HIV-exposed infants in a randomized trial. JCI Insight 2017, 2, e91963. [Google Scholar] [CrossRef] [PubMed]
- Hesseling, A.C.; Caldwell, J.; Cotton, M.F.; Eley, B.S.; Jaspan, H.B.; Jennings, K.; Marais, B.J.; Nuttall, J.; Rabie, H.; Roux, P.; et al. BCG vaccination in South African HIV-exposed infants—Risks and benefits. S. Afr. Med. J. 2009, 99, 88–91. [Google Scholar] [PubMed]
- Fastenackels, S.; Sauce, D.; Vigouroux, C.; Avettand-Fènoël, V.; Bastard, J.-P.; Fellahi, S.; Nailler, L.; Arezes, E.; Rouzioux, C.; Warszawski, J.; et al. HIV-mediated immune aging in young adults infected perinatally or during childhood. AIDS 2019, 33, 1705–1710. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.W.; Elahi, S. Profound immune consequences for young adults infected with HIV perinatally or during childhood: A cautionary tale regarding adherence to antiretroviral therapy. AIDS 2019, 33, 2251–2252. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katusiime, M.G.; Van Zyl, G.U.; Cotton, M.F.; Kearney, M.F. HIV-1 Persistence in Children during Suppressive ART. Viruses 2021, 13, 1134. https://doi.org/10.3390/v13061134
Katusiime MG, Van Zyl GU, Cotton MF, Kearney MF. HIV-1 Persistence in Children during Suppressive ART. Viruses. 2021; 13(6):1134. https://doi.org/10.3390/v13061134
Chicago/Turabian StyleKatusiime, Mary Grace, Gert U. Van Zyl, Mark F. Cotton, and Mary F. Kearney. 2021. "HIV-1 Persistence in Children during Suppressive ART" Viruses 13, no. 6: 1134. https://doi.org/10.3390/v13061134
APA StyleKatusiime, M. G., Van Zyl, G. U., Cotton, M. F., & Kearney, M. F. (2021). HIV-1 Persistence in Children during Suppressive ART. Viruses, 13(6), 1134. https://doi.org/10.3390/v13061134