
Mastomys natalensis Has a Cellular Immune Response Profile Distinct from Laboratory Mice

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Reagents and Antibodies
2.3. Tissue Preparation
2.4. T Cell Proliferation
2.5. Extracellular Staining for Flow Cytometry
2.6. Intracellular Staining for Flow Cytometry
2.7. Cytometric Bead Array (CBA)
2.8. Software and Statistical Analysis
3. Results
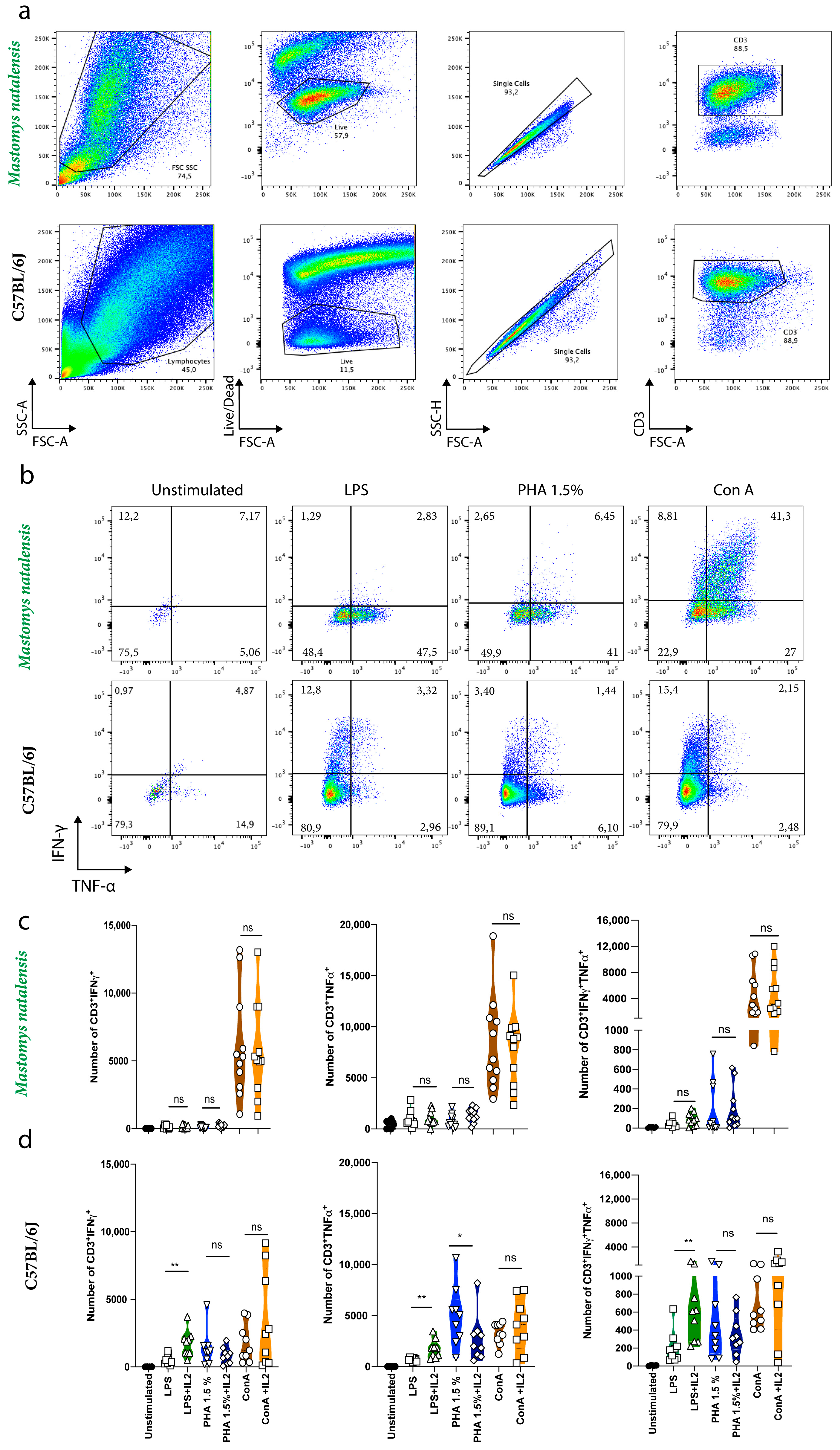
3.1. Commercial Rat and Mouse Antibodies Cross-React with M. natalensis T Cell Receptors and Intracellular Cytokines
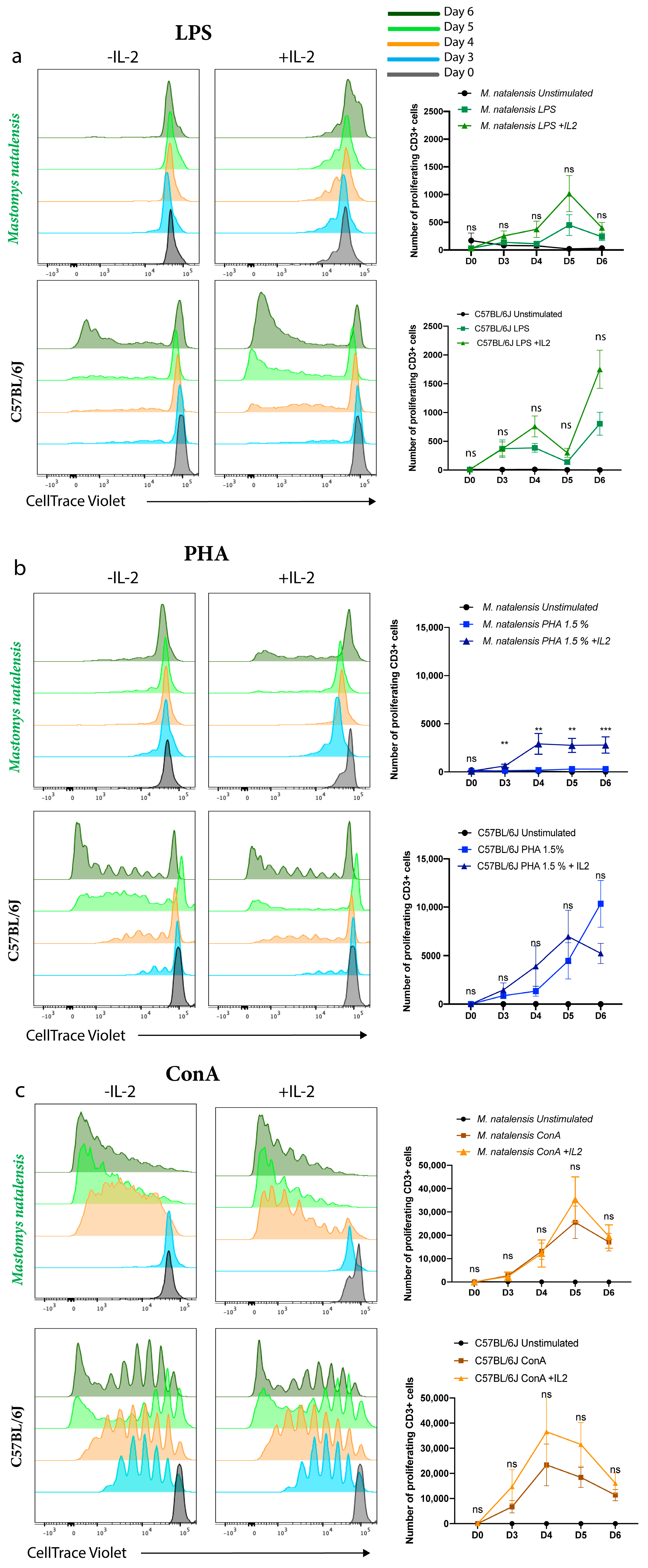
3.2. ConA Mitogen Efficiently Induced M. natalensis T Cell Proliferation In Vitro
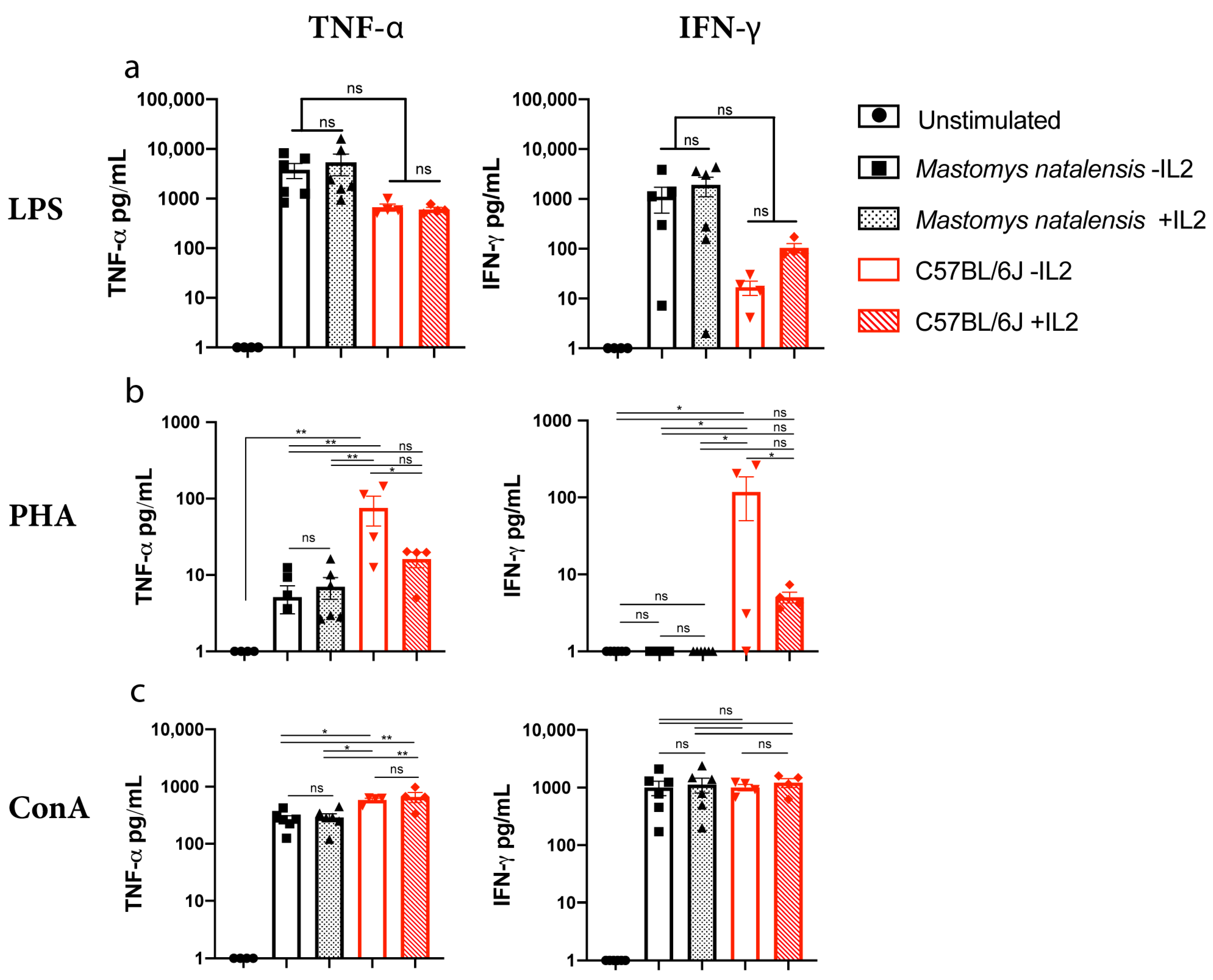
3.3. Comparative Secretion of Effector Molecules in M. natalensis in Response to Stimuli
3.4. Comparative Expression of Intracellular Cytokines in M. natalensis in Response to Different Stimuli
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hasche, D.; Rösl, F. Mastomys Species as Model Systems for Infectious Diseases. Viruses 2019, 11, 182. [Google Scholar] [CrossRef]
- Jansa, S.A.; Giarla, T.C.; Lim, B.K. The Phylogenetic Position of the Rodent GenusTyphlomysand the Geographic Origin of Muroidea. J. Mammal. 2009, 90, 1083–1094. [Google Scholar] [CrossRef]
- Lecompte, E.; Granjon, L.; Denys, C. The phylogeny of the Praomys complex (Rodentia: Muridae) and its phylogeographic implications. J. Zool. Syst. Evol. Res. 2002, 40, 8–25. [Google Scholar] [CrossRef]
- Chevret, P.; Granjon, L.; Duplantier, J.-M.; Denys, C.; Catzeflis, F.M. Molecular phylogeny of the Praomys complex (Rodentia: Murinae): A study based on DNA/DNA hybridization experiments. Zool. J. Linn. Soc. 1994, 112, 425–442. [Google Scholar] [CrossRef]
- Walker, D.H.; Wulff, H.; Lange, J.V.; Murphy, F.A. Comparative pathology of Lassa virus infection in monkeys, guinea-pigs, and Mastomys natalensis. Bull. World Health Organ. 1975, 52, 523–534. [Google Scholar] [PubMed]
- Olayemi, A.; Cadar, D.; Magassouba, N.; Obadare, A.; Kourouma, F.; Oyeyiola, A.; Fasogbon, S.; Igbokwe, J.; Rieger, T.; Bockholt, S.; et al. New Hosts of The Lassa Virus. Sci. Rep. 2016, 6, 25280. [Google Scholar] [CrossRef]
- Monath, T.P.; Newhouse, V.F.; Kemp, G.E.; Setzer, H.W.; Cacciapuoti, A. Lassa Virus Isolation from Mastomys natalensis Rodents during an Epidemic in Sierra Leone. Science 1974, 185, 263–265. [Google Scholar] [CrossRef]
- Sadlova, J.; Vojtkova, B.; Hrncirova, K.; Lestinova, T.; Spitzova, T.; Becvar, T.; Votypka, J.; Bates, P.; Volf, P. Host competence of African rodents Arvicanthis neumanni, A. niloticus and Mastomys natalensis for Leishmania major. Int. J. Parasitol. Parasites Wildl. 2019, 8, 118–126. [Google Scholar] [CrossRef]
- Boardman, K.; Rosenke, K.; Safronetz, D.; Feldmann, H.; Schwan, T.G. Host Competency of the Multimammate Rat Mastomys natalensis Demonstrated by Prolonged Spirochetemias with the African Relapsing Fever Spirochete Borrelia crocidurae. Am. J. Trop. Med. Hyg. 2019, 101, 1272–1275. [Google Scholar] [CrossRef]
- Zumpt, F. Is the Multimammate Rat a Natural Reservoir of Borrelia Duttoni? Nat. Cell Biol. 1959, 184, 793–794. [Google Scholar] [CrossRef]
- Schwan, T.G.; Anderson, J.M.; Lopez, J.E.; Fischer, R.J.; Raffel, S.J.; McCoy, B.N.; Safronetz, D.; Sogoba, N.; Maïga, O.; Traoré, S.F. Endemic Foci of the Tick-Borne Relapsing Fever Spirochete Borrelia crocidurae in Mali, West Africa, and the Potential for Human Infection. PLoS Negl. Trop. Dis. 2012, 6, e1924. [Google Scholar] [CrossRef]
- Green, C.A.; Gordon, D.H.; Lyons, N.F. Biological Species in Praomys (Mastomys) Natalensis (Smith), a Rodent Carrier of Lassa Virus and Bubonic Plague in Africa. Am. J. Trop. Med. Hyg. 1978, 27, 627–629. [Google Scholar] [CrossRef] [PubMed]
- Crawford, S.E.; Patel, D.G.; Cheng, E.; Berkova, Z.; Hyser, J.M.; Ciarlet, M.; Finegold, M.J.; Conner, M.E.; Estes, M.K. Rotavirus Viremia and Extraintestinal Viral Infection in the Neonatal Rat Model. J. Virol. 2006, 80, 4820–4832. [Google Scholar] [CrossRef]
- Kesavalu, L.; Sathishkumar, S.; Bakthavatchalu, V.; Matthews, C.; Dawson, D.; Steffen, M.; Ebersole, J.L. Rat Model of Polymicrobial Infection, Immunity, and Alveolar Bone Resorption in Periodontal Disease. Infect. Immun. 2007, 75, 1704–1712. [Google Scholar] [CrossRef]
- Wang, G.; Ojaimi, C.; Wu, H.; Saksenberg, V.; Iyer, R.; Liveris, D.; McClain, S.A.; Wormser, G.P.; Schwartz, I. Disease severity in a murine model of lyme borreliosis is associated with the genotype of the infecting Borrelia burgdorferi sensu stricto strain. J. Infect. Dis. 2002, 186, 782–791. [Google Scholar] [CrossRef]
- Loria-Cervera, E.N.; Andrade-Narvaez, F.J. Animal models for the study of leishmaniasis immunology. Revista do Instituto de Medicina Tropical de São Paulo 2014, 56, 1–11. [Google Scholar] [CrossRef]
- Yun, N.E.; Ronca, S.; Tamura, A.; Koma, T.; Seregin, A.V.; Dineley, K.T.; Miller, M.; Cook, R.; Shimizu, N.; Walker, A.G.; et al. Animal Model of Sensorineural Hearing Loss Associated with Lassa Virus Infection. J. Virol. 2016, 90, 2920–2927. [Google Scholar] [CrossRef]
- Goicochea, M.A.; Zapata, J.C.; Bryant, J.; Davis, H.; Salvato, M.S.; Lukashevich, I.S. Evaluation of Lassa virus vaccine immunogenicity in a CBA/J-ML29 mouse model. Vaccine 2012, 30, 1445–1452. [Google Scholar] [CrossRef]
- Zhu, J.; Yamane, H.; Paul, W.E. Differentiation of Effector CD4 T Cell Populations. Annu. Rev. Immunol. 2010, 28, 445–489. [Google Scholar] [CrossRef]
- Battegay, M.; Moskophidis, D.; Rahemtulla, A.; Hengartner, H.; Mak, T.W.; Zinkernagel, R.M. Enhanced establishment of a virus carrier state in adult CD4+ T-cell-deficient mice. J. Virol. 1994, 68, 4700–4704. [Google Scholar] [CrossRef]
- Matloubian, M.; Concepcion, R.J.; Ahmed, R. CD4+ T cells are required to sustain CD8+ cytotoxic T-cell responses during chronic viral infection. J. Virol. 1994, 68, 8056–8603. [Google Scholar] [CrossRef]
- Demers, K.R.; Reuter, M.A.; Betts, M.R. CD8+T-cell effector function and transcriptional regulation during HIV pathogenesis. Immunol. Rev. 2013, 254, 190–206. [Google Scholar] [CrossRef]
- Darrah, P.A.; Patel, D.T.; De Luca, P.M.; Lindsay, R.W.B.; Davey, D.F.; Flynn, B.J.; Hoff, S.T.; Andersen, P.; Reed, S.G.; Morris, S.L.; et al. Multifunctional TH1 cells define a correlate of vaccine-mediated protection against Leishmania major. Nat. Med. 2007, 13, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, V.; Cordeiro-Da-Silva, A.; LaForge, M.; Ouaissi, A.; Akharid, K.; Silvestre, R.; Estaquier, J. Impairment of T Cell Function in Parasitic Infections. PLoS Negl. Trop. Dis. 2014, 8, e2567. [Google Scholar] [CrossRef] [PubMed]
- Schussek, S.; Trieu, A.; Apte, S.H.; Sidney, J.; Sette, A.; Doolan, D.L. Immunization with Apical Membrane Antigen 1 Confers Sterile Infection-Blocking Immunity against Plasmodium Sporozoite Challenge in a Rodent Model. Infect. Immun. 2013, 81, 3586–3599. [Google Scholar] [CrossRef]
- Egui, A.; Ledesma, D.; Pérez-Antón, E.; Montoya, A.; Gómez, I.; Robledo, S.M.; Infante, J.J.; Vélez, I.D.; López, M.C.; Thomas, M.C. Phenotypic and Functional Profiles of Antigen-Specific CD4+ and CD8+ T Cells Associated With Infection Control in Patients With Cutaneous Leishmaniasis. Front. Cell. Infect. Microbiol. 2018, 8, 393. [Google Scholar] [CrossRef]
- Qiu, Z.; Khairallah, C.; Sheridan, B.S. Listeria Monocytogenes: A Model Pathogen Continues to Refine Our Knowledge of the CD8 T Cell Response. Pathogens 2018, 7, 55. [Google Scholar] [CrossRef] [PubMed]
- Thakur, A.; Mikkelsen, H.; Jungersen, G. Intracellular Pathogens: Host Immunity and Microbial Persistence Strategies. J. Immunol. Res. 2019, 2019, 1–24. [Google Scholar] [CrossRef]
- Elsaesser, H.; Sauer, K.; Brooks, D.G. IL-21 Is Required to Control Chronic Viral Infection. Science 2009, 324, 1569–1572. [Google Scholar] [CrossRef]
- Martin, M.D.; Badovinac, V.P. Antigen-dependent and -independent contributions to primary memory CD8 T cell activation and protection following infection. Sci. Rep. 2015, 5, 18022. [Google Scholar] [CrossRef] [PubMed]
- Gideon, H.P.; Phuah, J.; Myers, A.J.; Bryson, B.D.; Rodgers, M.A.; Coleman, M.T.; Maiello, P.; Rutledge, T.; Marino, S.; Fortune, S.M.; et al. Variability in Tuberculosis Granuloma T Cell Responses Exists, but a Balance of Pro- and Anti-inflammatory Cytokines Is Associated with Sterilization. PLoS Pathog. 2015, 11, e1004603. [Google Scholar] [CrossRef]
- Li, B.; Du, C.; Zhou, L.; Bi, Y.; Wang, X.; Wen, L.; Guo, Z.; Song, Z.; Yang, R. Humoral and Cellular Immune Responses to Yersinia pestis Infection in Long-Term Recovered Plague Patients. Clin. Vaccine Immunol. 2011, 19, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Boussoffara, T.; Chelif, S.; Ben Ahmed, M.; Mokni, M.; Ben Salah, A.; Dellagi, K.; Louzir, H. Immunity Against Leishmania major Infection: Parasite-Specific Granzyme B Induction as a Correlate of Protection. Front. Cell. Infect. Microbiol. 2018, 8, 397. [Google Scholar] [CrossRef]
- Campos, T.M.; Costa, R.; Passos, S.; Carvalho, L.P. Cytotoxic activity in cutaneous leishmaniasis. Memórias Inst. Oswaldo Cruz 2017, 112, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Fahey, L.M.; Wilson, E.B.; Elsaesser, H.; Fistonich, C.D.; McGAVERN, D.B.; Brooks, D.G. Viral persistence redirects CD4 T cell differentiation toward T follicular helper cells. J. Exp. Med. 2011, 208, 987–999. [Google Scholar] [CrossRef]
- Crotty, S. T Follicular Helper Cell Differentiation, Function, and Roles in Disease. Immunity 2014, 41, 529–542. [Google Scholar] [CrossRef] [PubMed]
- McKisic, M.D.; Barthold, S.W. T-Cell-Independent Responses to Borrelia burgdorferi Are Critical for Protective Immunity and Resolution of Lyme Disease. Infect. Immun. 2000, 68, 5190–5197. [Google Scholar] [CrossRef]
- Lakhal-Naouar, I.; Boussoffara, T.; Meddeb-Garnaoui, A.; Ben Achour-Chenik, Y.; Louzir, H.; Chenik, M. Cellular and Humoral Responses to Leishmania major Virulence Factors in Healed Cutaneous Leishmaniasis and Mediterranean Visceral Leishmaniasis Patients. Clin. Vaccine Immunol. 2009, 16, 956–958. [Google Scholar] [CrossRef]
- Goncalves, R.; Christensen, S.M.; Mosser, D.M. Humoral immunity in leishmaniasis—Prevention or promotion of parasite growth? Cytokine X 2020, 2, 100046. [Google Scholar] [CrossRef]
- Safronetz, D.; Rosenke, K.; Fischer, R.J.; LaCasse, R.A.; Scott, D.P.; Saturday, G.; Hanley, P.W.; Maiga, O.; Sogoba, N.; Schwan, T.G.; et al. Establishment of a Genetically Confirmed Breeding Colony of Mastomys natalensis from Wild-Caught Founders from West Africa. Viruses 2021, 13, 590. [Google Scholar] [CrossRef]
- Shinohara, Y.; Tsukimoto, M. Adenine Nucleotides Attenuate Murine T Cell Activation Induced by Concanavalin A or T Cell Receptor Stimulation. Front. Pharmacol. 2018, 8, 986. [Google Scholar] [CrossRef] [PubMed]
- Palacios, R. Concanavalin A triggers T lymphocytes by directly interacting with their receptors for activation. J. Immunol. 1982, 128, 337–342. [Google Scholar]
- Yamamura, Y.; Tanaka, J.L.; Madyastha, K.R.; Fudenberg, H.H.; Proctor, J.W. Differences in Mitogenic Responses of Murine T Cells to Two Distinct Phytohemagglutinin (Pha) Subcomponents. Immunol. Commun. 1981, 10, 9–20. [Google Scholar] [CrossRef]
- Ceuppens, J.L.; Baroja, M.L.; Lorre, K.; Van Damme, J.; Billiau, A. Human T cell activation with phytohemagglutinin. The function of IL-6 as an accessory signal. J. Immunol. 1988, 141, 3868–3874. [Google Scholar]
- Katzen, D.; Chu, E.; Terhost, C.; Leung, D.Y.; Gesner, M.; Miller, R.A.; Geha, R.S. Mechanisms of human T cell response to mitogens: IL 2 induces IL 2 receptor expression and proliferation but not IL 2 synthesis in PHA-stimulated T cells. J. Immunol. 1985, 135, 1840–1845. [Google Scholar]
- Tough, D.F.; Sun, S.; Sprent, J. T Cell Stimulation In Vivo by Lipopolysaccharide (LPS). J. Exp. Med. 1997, 185, 2089–2094. [Google Scholar] [CrossRef]
- McAleer, J.P.; Vella, A.T. Understanding how lipopolysaccharide impacts CD4 T cell immunity. Crit. Rev. Immunol. 2008, 28, 281–299. [Google Scholar]
- Vogel, S.N.; Hilfiker, M.L.; Caulfield, M.J. Endotoxin-induced T lymphocyte proliferation. J. Immunol. 1983, 130, 1774–1779. [Google Scholar] [PubMed]
- Brown, A.F.; Murphy, A.G.; Lalor, S.J.; Leech, J.M.; O’Keeffe, K.M.; Mac Aogáin, M.; O’Halloran, D.P.; Lacey, K.A.; Tavakol, M.; Hearnden, C.H.; et al. Memory Th1 Cells Are Protective in Invasive Staphylococcus aureus Infection. PLoS Pathog. 2015, 11, e1005226. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, J.; Ge, S.; Chen, C.; Li, S.; Wu, X.; Feng, X.; Wang, Y.; Cai, D. Saikosaponin A Inhibits Breast Cancer by Regulating Th1/Th2 Balance. Front. Pharmacol. 2019, 10, 624. [Google Scholar] [CrossRef]
- Benihoud, K.; Esselin, S.; Descamps, D.; Jullienne, B.; Salone, B.; Bobé, P.; Bonardelle, D.; Connault, E.; Opolon, P.; Saggio, I.; et al. Erratum: Respective roles of TNF-α and IL-6 in the immune response-elicited by adenovirus-mediated gene transfer in mice. Gene Ther. 2007, 14, 551. [Google Scholar] [CrossRef]
- Schroder, K.; Hertzog, P.J.; Ravasi, T.; Hume, D.A. Interferon-γ: An overview of signals, mechanisms and functions. J. Leukoc. Biol. 2003, 75, 163–189. [Google Scholar] [CrossRef]
- Westermann, J.; van Lessen, A.; Schlimper, C.; Baskaynak, G.; le Coutre, P.; Dörken, B.; Pezzutto, A. Simultaneous cytokine analysis by cytometric bead array for the detection of leukaemia-reactive T cells in patients with chronic myeloid leukaemia. Br. J. Haematol. 2006, 132, 32–35. [Google Scholar] [CrossRef]
- Karlsson, A.C.; Martin, J.N.; Younger, S.R.; Bredt, B.M.; Epling, L.; Ronquillo, R.; Varma, A.; Deeks, S.G.; McCune, J.M.; Nixon, D.F.; et al. Comparison of the ELISPOT and cytokine flow cytometry assays for the enumeration of antigen-specific T cells. J. Immunol. Methods 2003, 283, 141–153. [Google Scholar] [CrossRef]
- Morgan, E.; Varro, R.; Sepulveda, H.; Ember, J.A.; Apgar, J.; Wilson, J.; Lowe, L.; Chen, R.; Shivraj, L.; Agadir, A.; et al. Cytometric bead array: A multiplexed assay platform with applications in various areas of biology. Clin. Immunol. 2004, 110, 252–266. [Google Scholar] [CrossRef]
- Hardin, A.; Nevonen, K.A.; Eckalbar, W.L.; Carbone, L.; Ahituv, N. Comparative Genomic Characterization of the Multimammate Mouse Mastomys coucha. Mol. Biol. Evol. 2019, 36, 2805–2812. [Google Scholar] [CrossRef]
- Masopust, D.; Sivula, C.P.; Jameson, S.C. Of Mice, Dirty Mice, and Men: Using Mice To Understand Human Immunology. J. Immunol. 2017, 199, 383–388. [Google Scholar] [CrossRef]
- Rosshart, S.P.; Herz, J.; Vassallo, B.G.; Hunter, A.; Wall, M.K.; Badger, J.H.; McCulloch, J.A.; Anastasakis, D.G.; Sarshad, A.A.; Leonardi, I.; et al. Laboratory mice born to wild mice have natural microbiota and model human immune responses. Science 2019, 365, eaaw4361. [Google Scholar] [CrossRef] [PubMed]
- Komai-Koma, M.; Gilchrist, D.S.; Xu, D. Direct recognition of LPS by human but not murine CD8+T cellsviaTLR4 complex. Eur. J. Immunol. 2009, 39, 1564–1572. [Google Scholar] [CrossRef]
- Chien, E.J.; Chien, C.-H.; Chen, J.-J.; Wang, S.-W.; Hsieh, D.J.-Y. Bacterial lipopolysaccharide activates protein kinase C, but not intracellular calcium elevation, in human peripheral T cells. J. Cell. Biochem. 2000, 76, 404–410. [Google Scholar] [CrossRef]
- Ulmer, A.J.; Flad, H.-D.; Rietschel, T.; Mattern, T. Induction of proliferation and cytokine production in human T lymphocytes by lipopolysaccharide (LPS). Toxicology 2000, 152, 37–45. [Google Scholar] [CrossRef]
- Wolk, K.; Döcke, W.D.; von Baehr, V.; Volk, H.D.; Sabat, R. Impaired antigen presentation by human monocytes during endotoxin tolerance. Blood 2000, 96, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Mattern, T.; Thanhäuser, A.; Reiling, N.; Toellner, K.M.; Duchrow, M.; Kusumoto, S.; Rietschel, E.T.; Ernst, M.; Brade, H.; Flad, H.D. Endotoxin and lipid A stimulate proliferation of human T cells in the presence of autologous monocytes. J. Immunol. 1994, 153, 2996–3004. [Google Scholar]
- O’Neil-Andersen, N.J.; Lawrence, D.A. Differential Modulation of Surface and Intracellular Protein Expression by T Cells after Stimulation in the Presence of Monensin or Brefeldin A. Clin. Vaccine Immunol. 2002, 9, 243–250. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Guy, C.S.; Vignali, K.M.; Temirov, J.; Bettini, M.L.; Overacre, A.E.; Smeltzer, M.; Zhang, H.; Huppa, J.B.; Tsai, Y.-H.; Lobry, C.; et al. Distinct TCR signaling pathways drive proliferation and cytokine production in T cells. Nat. Immunol. 2013, 14, 262–270. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Mechanical and Tissue Dissociation | ||
| Reagents | References | Vendors |
| Gibco Fetal Bovine Serum | 16000044 | ThermoFisher |
| RPMI | R8758 | Sigma-Aldrich |
| Pencillin/Streptomycin | 15070063 | ThermoFisher |
| L-glutamine | 25030164 | ThermoFisher |
| β-Mercaptoethanol | M3148-25ML | Sigma-Aldrich |
| Miltenyi dissociator C tubes | 130-096-334 | Miltenyi |
| Cell strainer (40 μm) | 22-363-547 | Fisherscientific |
| Red blood cell lysis (1× RBC Lysis Buffer) | 00-4333-57 | ThermoFisher |
| In vitro Lymphocytes stimulation and proliferation | ||
| Reagents | References | Vendors |
| Concanavalin A | 00-4978-03 | ThermoFisher |
| Phytohaemagglutinin P | 10576015 | ThermoFisher |
| Lipopolysaccharide | L2630-10MG | Sigma-Aldrich |
| Mouse interleukin (IL)-2 | 130-120-331 | Miltenyi |
| CellTrace Violet | C34557 | ThermoFisher |
| Reagents and antibodies for flow cytometry | ||
| Reagents | References | Vendors |
| Phorbol 12-myristate 13-acetate | P8139-1MG | Sigma-Aldrich |
| Ionomycin | 407950-1MG | Merck Calbiochem |
| Brefeldin A | 00-4506-51 | ThermoFisher |
| TruStain FcX | 101320 | BioLegend |
| Fixable Viability Dye eFluor™ 780 | 65-0865-14 | ThermoFisher |
| Intracellular Fixation & Permeabilization Buffer Set | 88-8824-00 | ThermoFisher |
| Rat anti Human CD3 FITC | MCA1477F | Bio-Rad |
| Anti-mouse TNF-α Brilliant Violet 785 | 506341 | BioLegend |
| Mouse Anti-Rat IFN-γ PE | 559499 | BDbiosciences |
| Reagents | Source | Catalog Number | Cross-React with Mastomys | |
|---|---|---|---|---|
| Cytometric Bead Array (CBA) | Rat IFN-γ FlexSet | BD | 558305 | Yes |
| Mouse TNF-α Flex Set | BD | 558299 | Yes | |
| Mouse/Rat Soluble Protein Master Buffer Kit | BD | 558266 | Yes | |
| ELISPOT | Rat IFN-γ Single color | ImmunoSpot | No | |
| Mouse TNF-α Single color | ImmunoSpot | Yes | ||
| ELISA | Mouse TNF-α ELISA MAX™Standard Set | BioLegend | 430901 | No |
| Mouse IFN-γ ELISA MAX™Standard Set | BioLegend | 430801 | No | |
| Purified Rat Anti-Mouse IFN-γ | BD | 551309 | No | |
| Biotin Anti-Mouse IFN-γ | BD | 551506 | No | |
| Biotin Rat Anti-Mouse IFN-γ | BD | 554410 | No | |
| Recombinant Rat IFN-γ | BD | 550072 | No |
| Specificity | Antibody | Conjugate | Clone | Reference | Vendor | Cross-React with Mastomys |
|---|---|---|---|---|---|---|
| Mouse | Purified anti-mouse CD4 | N/A | GK1.5 | 100401 | Biolegend | No |
| CD3e | PE | 145-2C11 | 100307 | Biolegend | No | |
| CD8a | APC | 53.6.7 | 100711 | Biolegend | No | |
| IFN-γ | APC | XMG1.2 | 505809 | Biolegend | No | |
| TNF-α | Brilliant Violet 785 | MP6-XT22 | 506341 | Biolegend | Yes | |
| TNF-α | PE | MP6-XT22 | 12-7321-41 | eBioscience | Yes | |
| Rat/Human/Mouse | CD3 | FITC | CD3-12 | MCA1477F | Bio-Rad | Yes |
| Rat | CD3 | FITC | G4.18 | 559975 | BD | No |
| Purified anti-rat CD8b | N/A | 341 | 200702 | Biolegend | Yes | |
| CD8b | PE | eBio341 | 12-0080-82 | ThermoFisher | No | |
| CD8a | APC | G28 | 200609 | Biolegend | No | |
| CD8a | BV421 | OX-8 | 740041 | BD | No | |
| CD8b | BV421 | 341 | 742915 | BD | No | |
| Purified anti-rat CD8a | N/A | OX-8 | 201701 | Biolegend | No | |
| Purified anti-rat CD4 | N/A | W3/25 | 201501 | Biolegend | No | |
| CD4 | BV786 | OX-35 | 740912 | BD | No | |
| CD4 | BV786 | OX-38 | 743093 | BD | No | |
| CD4 | APC | W3/25 | 201509 | Biolegend | No | |
| IFN-γ | AF647 | DB-1 | 562213 | BD | Yes | |
| IFN-γ | PE | DB-1 | 559499 | BD | Yes | |
| Rat/mouse/rabbit | TNF-α | PE | TN3-19.12 | 559503 | BD | No |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang-Huau, T.-L.; Rosenke, K.; Meade-White, K.; Carmody, A.; Smith, B.J.; Bosio, C.M.; Jarvis, M.A.; Feldmann, H. Mastomys natalensis Has a Cellular Immune Response Profile Distinct from Laboratory Mice. Viruses 2021, 13, 729. https://doi.org/10.3390/v13050729
Tang-Huau T-L, Rosenke K, Meade-White K, Carmody A, Smith BJ, Bosio CM, Jarvis MA, Feldmann H. Mastomys natalensis Has a Cellular Immune Response Profile Distinct from Laboratory Mice. Viruses. 2021; 13(5):729. https://doi.org/10.3390/v13050729
Chicago/Turabian StyleTang-Huau, Tsing-Lee, Kyle Rosenke, Kimberly Meade-White, Aaron Carmody, Brian J. Smith, Catharine M. Bosio, Michael A. Jarvis, and Heinz Feldmann. 2021. "Mastomys natalensis Has a Cellular Immune Response Profile Distinct from Laboratory Mice" Viruses 13, no. 5: 729. https://doi.org/10.3390/v13050729
APA StyleTang-Huau, T.-L., Rosenke, K., Meade-White, K., Carmody, A., Smith, B. J., Bosio, C. M., Jarvis, M. A., & Feldmann, H. (2021). Mastomys natalensis Has a Cellular Immune Response Profile Distinct from Laboratory Mice. Viruses, 13(5), 729. https://doi.org/10.3390/v13050729