
Detection of Laryngotracheitis Virus in Poultry Flocks with Respiratory Disorders in Slovenia

Abstract
1. Introduction
2. Materials and Methods
2.1. Affected Poultry Flocks
2.2. Molecular Analyses
3. Results
3.1. Detection of Respiratory Pathogens by PCR
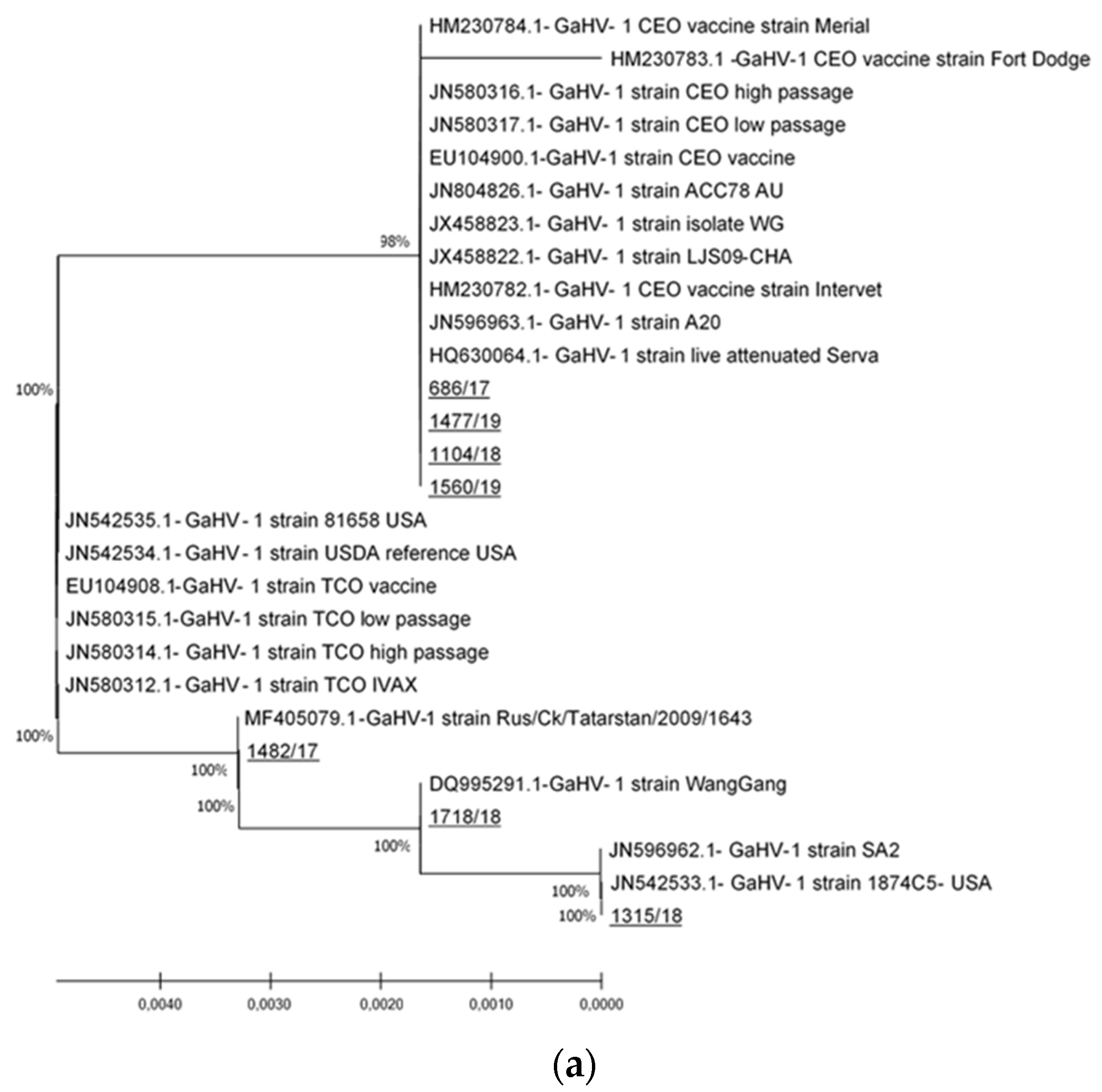
3.2. ICP4 Sequencing
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- García, M.; Spatz, S. Infectious Laryngotracheitis. In Diseases of Poultry, 14th ed.; Swayne, D.E., Boulianne, M., Logue, C.M., McDougald, L.R., Nair, V., Suarez, D.L., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2020; Volume 1, pp. 189–203. [Google Scholar]
- Gowthaman, V.; Kumar, S.; Koul, M.; Dave, U.; Murthy, T.R.G.K.; Munuswamy, P.; Tiwari, R.; Karthik, K.; Dhama, K.; Michalak, I.; et al. Infectious laryngotracheitis: Etiology, epidemiology, pathobiology, and advances in diagnosis and control—A comprehensive review. Vet. Q. 2020, 40, 140–161. [Google Scholar] [CrossRef]
- Menendez, K.R.; Garcia, M.; Spatz, S.; Tablante, N.L. Molecular epidemiology of infectious laryngotracheitis: A review. Avian Pathol. 2014, 43, 108–117. [Google Scholar] [CrossRef]
- Kirkpatrick, N.C.; Mahmoudian, A.; Colson, C.A.; Devlin, J.M.; Noormohammadi, A.H. Relationship between mortality, clinical signs and tracheal pathology in infectious laryngotracheitis. Avian Pathol. 2006, 35, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Oldoni, I.; Rodriguez-Avila, A.; Riblet, S.; Garcia, M. Characterization of infectious laryngotracheitis virus (ILTV) isolates from commercial poultry by polymerase chain reaction and restriction fragment length polymorphism (PCR-RFLP). Avian Dis. 2008, 52, 59–63. [Google Scholar] [CrossRef]
- Williams, R.A.; Bennett, M.; Bradbury, J.M.; Gaskell, R.M.; Jones, R.C.; Jordan, F.T.W. Demonstration of sites of latency of infectious laryngotracheitis virus using the polymerase chain reaction. J. Gen. Virol. 1992, 73 Pt 9, 2415–2420. [Google Scholar] [CrossRef] [PubMed]
- Hughes, C.S.; Gaskell, R.M.; Jones, R.C.; Bradbury, J.M.; Jordan, F.T. Effects of certain stress factors on the re-excretion of infectious laryngotracheitis virus from latently infected carrier birds. Res. Vet. Sci. 1989, 46, 274–276. [Google Scholar] [CrossRef]
- Hughes, C.S.; Williams, R.A.; Gaskell, R.M.; Jordan, F.T.W.; Bradbury, J.M.; Bennett, M.; Jones, R.C. Latency and reactivation of infectious laryngotracheitis vaccine virus. Arch. Virol. 1991, 121, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Bagust, T.J.; Johnson, M.A. Avian infectious laryngotracheitis: Virus-host interactions in relation to prospects for eradication. Avian Pathol. 1995, 24, 373–391. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M. Current and future vaccines and vaccination strategies against infectious laryngotracheitis (ILT) respiratory disease of poultry. Vet. Microbiol. 2017, 206, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.; Zavala, G. Commercial Vaccines and Vaccination Strategies Against Infectious Laryngotracheitis: What We Have Learned and Knowledge Gaps That Remain. Avian Dis. 2019, 63, 325–334. [Google Scholar] [CrossRef]
- Bayoumi, M.; El-Saied, M.; Amer, H.; Bastami, M.; Sakr, E.E.; El-Mahdy, M. Molecular characterization and genetic diversity of the infectious laryngotracheitis virus strains circulating in Egypt during the outbreaks of 2018 and 2019. Arch. Virol. 2020, 165, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Ojkic, D.; Swinton, J.; Vallieres, M.; Martin, E.; Shapiro, J.; Sanei, B.; Binnington, B. Characterization of field isolates of infectious laryngotracheitis virus from Ontario. Avian Pathol. 2006, 35, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.A.; Tyack, S.G.; Prideaux, C.; Kongsuwan, K.; Sheppard, M. Nucleotide sequence of infectious laryngotracheitis virus (gallid herpesvirus 1) ICP4 gene. Virus Res. 1995, 35, 193–204. [Google Scholar] [CrossRef]
- Chacon, J.L.; Ferreira, A.J.P. Differentiation of field isolates and vaccine strains of infectious laryngotracheitis virus by DNA sequencing. Vaccine 2009, 27, 6731–6738. [Google Scholar] [CrossRef]
- Madsen, J.M.; Zimmermann, N.G.; Timmons, J.; Tablante, N.L. Prevalence and differentiation of diseases in Maryland backyard flocks. Avian Dis. 2013, 57, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Blakey, J.; Stoute, S.; Crossley, B.; Mete, A. Retrospective analysis of infectious laryngotracheitis in backyard chicken flocks in California, 2007–2017, and determination of strain origin by partial ICP4 sequencing. J. Vet. Diagn. Investig. 2019, 31, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Spackman, E.; Senne, D.A.; Myers, T.J.; Bulaga, L.L.; Garber, L.P.; Perdue, M.L.; Lohman, K.; Daum, L.T.; Suarez, D.L. Development of a real-time reverse transcriptase PCR assay for type A influenza virus and the avian H5 and H7 hemagglutinin subtypes. J. Clin. Microbiol. 2002, 40, 3256–3260. [Google Scholar] [CrossRef]
- Wise, M.G.; Suarez, D.L.; Seal, B.S.; Pedersen, J.C.; Senne, D.A.; King, D.J.; Kapczynski, D.R.; Spackman, E. Development of a real-time reverse-transcription PCR for detection of newcastle disease virus RNA in clinical samples. J. Clin. Microbiol. 2004, 42, 329–338. [Google Scholar] [CrossRef]
- Muradrasoli, S.; Mohamed, N.; Hornyak, A.; Fohlman, J.; Olsen, B.; Belak, S.; Blomberg, J. Broadly targeted multiprobe QPCR for detection of coronaviruses: Coronavirus is common among mallard ducks (Anas platyrhynchos). J. Virol. Methods 2009, 159, 277–287. [Google Scholar] [CrossRef]
- VanDevanter, D.R.; Warrener, P.; Bennett, L.; Schultz, E.R.; Coulter, S.; Garber, R.L.; Rose, T.M. Detection and analysis of diverse herpesviral species by consensus primer PCR. J. Clin. Microbiol. 1996, 34, 1666–1671. [Google Scholar] [CrossRef]
- Manarolla, G.; Pisoni, G.; Sironi, G.; Rampin, T. Molecular biological characterization of avian poxvirus strains isolated from different avian species. Vet. Microbiol. 2010, 140, 1–8. [Google Scholar] [CrossRef]
- Raviv, Z.; Kleven, S.H. The development of diagnostic real-time TaqMan PCRs for the four pathogenic avian mycoplasmas. Avian Dis. 2009, 53, 103–107. [Google Scholar] [CrossRef]
- Kastelic, S.; Bercic, R.L.; Cizelj, I.; Bencina, M.; Makrai, L.; Zorman-Rojs, O.; Narat, M.; Bisgaard, M.; Christensen, H.; Bencina, D. Ornithobacterium rhinotracheale has neuraminidase activity causing desialylation of chicken and turkey serum and tracheal mucus glycoproteins. Vet. Microbiol. 2013, 162, 707–712. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Piccirillo, A.; Lavezzo, E.; Niero, G.; Moreno, A.; Massi, P.; Franchin, E.; Toppo, S.; Salata, C.; Palu, G. Full Genome Sequence-Based Comparative Study of Wild-Type and Vaccine Strains of Infectious Laryngotracheitis Virus from Italy. PLoS ONE 2016, 11, e0149529. [Google Scholar] [CrossRef]
- Moreno, A.; Piccirillo, A.; Mondin, A.; Morandini, E.; Gavazzi, L.; Cordioli, P. Epidemic of infectious laryngotracheitis in Italy: Characterization of virus isolates by PCR-restriction fragment length polymorphism and sequence analysis. Avian Dis. 2010, 54, 1172–1177. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.C.; Lee, Y.L.; Shien, J.H.; Shieh, H.K. Rapid differentiation of vaccine strains and field isolates of infectious laryngotracheitis virus by restriction fragment length polymorphism of PCR products. J. Virol. Methods 1997, 66, 179–186. [Google Scholar] [CrossRef]
- Graham, D.A.; McLaren, I.E.; Calvert, V.; Torrens, D.; Meehan, B.M. RFLP analysis of recent Northern Ireland isolates of infectious laryngotracheitis virus: Comparison with vaccine virus and field isolates from England, Scotland and the Republic of Ireland. Avian Pathol. 2000, 29, 57–62. [Google Scholar] [CrossRef]
- Kirkpatrick, N.C.; Mahmoudian, A.; O’Rourke, D.; Noormohammadi, A.H. Differentiation of infectious laryngotracheitis virus isolates by restriction fragment length polymorphic analysis of polymerase chain reaction products amplified from multiple genes. Avian Dis. 2006, 50, 28–34. [Google Scholar] [CrossRef]
- Loncoman, C.A.; Vaz, P.K.; Coppo, M.J.; Hartley, C.A.; Morera, F.J.; Browning, G.F.; Devlin, J.M. Natural recombination in alphaherpesviruses: Insights into viral evolution through full genome sequencing and sequence analysis. Infect. Genet. Evol. 2017, 49, 174–185. [Google Scholar] [CrossRef] [PubMed]
- Derksen, T.; Lampron, R.; Hauck, R.; Pitesky, M.; Gallardo, R.A. Biosecurity Assessment and Seroprevalence of Respiratory Diseases in Backyard Poultry Flocks Located Close to and Far from Commercial Premises. Avian Dis. 2018, 62, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Couto, R.M.; Braga, J.F.V.; Gomes, S.Y.M.; Resende, M.; Martins, N.R.S.; Ecco, R. Natural concurrent infections associated with infectious laryngotracheitis in layer chickens. J. Appl. Poult. Res. 2016, 25, 113–128. [Google Scholar] [CrossRef]
- Brochu, N.M.; Guerin, M.T.; Varga, C.; Lillie, B.N.; Brash, M.L.; Susta, L. A two-year prospective study of small poultry flocks in Ontario, Canada, part 1: Prevalence of viral and bacterial pathogens. J. Vet. Diagn. Investig. 2019, 31, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Slavec, B.; Sernel, T.; Krapež, U.; Zorman-Rojs, O. Longitudinal serological investigation of Mycoplasma synoviae infection in brolier breeder’s flocks in Slovenia. In Proceedings of the International Symposium on Avian Mycoplasmosis and Infectious Coryza, Leusden, The Netherlands, 13–15 November 2019; p. 27. [Google Scholar]
- Zorman-Rojs, O.; Slavec, B. Zdravstvena problematika v manjših rejah perutnine v Sloveniji. Vestn. Vet. Zb. Slov. 2020, 3, 179–184. [Google Scholar]
- Council Directive 2009/158/EC of 30 November 2009 on animal health conditions governing intra-Community trade in, and imports from third countries of, poultry and hatching eggs (Text with EEA relevance). OJ 2012, L343, 74–113.
- Guzman, M.; Hidalgo, H. Live Attenuated Infectious Bronchitis Virus Vaccines in Poultry: Modifying Local Viral Populations Dynamics. Animals 2020, 10, 2058. [Google Scholar] [CrossRef]
- Krapez, U.; Slavec, B.; Barlic-Maganja, D.; Rojs, O.Z. Molecular analysis of infectious bronchitis viruses isolated in Slovenia between 1990 and 2005: A retrospective study. Virus Genes 2010, 41, 414–416. [Google Scholar] [CrossRef]
- Krapez, U.; Slavec, B.; Rojs, O.Z. Circulation of infectious bronchitis virus strains from Italy 02 and QX genotypes in Slovenia between 2007 and 2009. Avian Dis. 2011, 55, 155–161. [Google Scholar] [CrossRef]
- Barbosa, E.V.; Cardoso, C.V.; Silva, R.C.F.; Cerqueira, A.M.F.; Liberal, M.H.T.; Castro, H.C. Ornithobacterium rhinotracheale: An Update Review about An Emerging Poultry Pathogen. Vet. Sci. 2020, 7, 3. [Google Scholar] [CrossRef]
- Fatunmbi, O.O.; Reed, W.M.; Schwartz, D.L.; Tripathy, D.N. Dual infection of chickens with pox and infectious laryngotracheitis (ILT) confirmed with specific pox and ILT DNA dot-blot hybridization assays. Avian Dis. 1995, 39, 925–930. [Google Scholar] [CrossRef] [PubMed]
- Diallo, I.S.; Taylor, J.; Gibson, J.; Hoad, J.; De Jong, A.; Hewitson, G.; Corney, B.G.; Rodwell, B.J. Diagnosis of a naturally occurring dual infection of layer chickens with fowlpox virus and gallid herpesvirus 1 (infectious laryngotracheitis virus). Avian Pathol. 2010, 39, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Tadese, T.; Potter, A.E.; Fitzgerald, S.; Reed, W.M. Concurrent infection in chickens with fowlpox virus and infectious laryngotracheitis virus as detected by immunohistochemistry and a multiplex polymerase chain reaction technique. Avian Dis. 2007, 51, 719–724. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Case ID/ Year of Collection | Flock Description | Age | Clinical Signs |
|---|---|---|---|
| 686/17 | Backyard flock, 25 hens, different ages | Not known | Severe respiratory disorders, purulent conjunctivitis, diarrhea |
| 1482/17 | Backyard flock, 80 broilers and 45 hens | 4 weeks, <1 year | Severe respiratory disorders and mortality seen in broilers, no clinical signs in adult hens |
| 1104/18 | Commercial layers, 6000 hens, farm with outdoor access 1 | 50 weeks | Mild respiratory disorders, drop in egg production (3%), higher daily mortality (0.81% per day) |
| 1315/18 | Backyard flock, 400 hens, two ages | >1 year, 20 weeks | Severe respiratory disorders and conjunctivitis observed only in younger hens |
| 1718/18 | Hobby flock, 420 hens, different ages | Not known | Nasal discharge, mild to severe respiratory disorders seen in 35% of birds |
| 1477/19 | Commercial layers, kept on multi-age farm, 6300 hens 1,2 | 25 weeks | Respiratory disorders, drop in egg production (c. 5%), higher daily mortality (0.24% per day) |
| 1560/19 | Commercial layers, 14,000 hens, multi-age farm 1 | 39 weeks | Respiratory disorders, slight drop in egg production (4.0%), higher daily mortality (0.12% per day) |
| Pathogen | Primer/Probe | Sequence (5′-3′) | Reference |
|---|---|---|---|
| AIV | M + 25 | AGATGAGTCTTCTAACCGAGGTCG | [18] |
| M − 124 | TGCAAAAACATCTTCAAGTCTCTG | ||
| M + 64 | FAM-TCAGGCCCCCTCAAAGCCGA-TAMRA | ||
| NCD (APMV1) | M + F4100 | AGTGATGTGCTCGGACCTTC | [19] |
| M − R4220 | CCTGAGGAGAGGCATTTGCTA | ||
| M + 4169 | FAM-TTCTCTAGCAGTGGGACAGCCTGC-TAMRA | ||
| Coronaviruses (IBV) | 11-FW | TGATGATGSNGTTGTNTGYTAYAA | [20] |
| 13-RV | GCATWGTRTGYTGNGARCARAATTC | ||
| probe III | FAM-TCTAARTGTTGGGTDGA-EDQ | ||
| Herpesvirus | DFA | GAYTTYGCNAGYYTNTAYCC | [21] |
| KG1 | TCCTGGACAAGCAGCARNYSGCNMTNAA | ||
| ILK | GTCTTGCTCACCAGNTCNACNCCYTT | ||
| TGV | TGTAACTCGGTGTAYGGNTTYACNGGNGT | ||
| IYG | CACAGAGTCCGTRTCNCCRTADAT | ||
| APV | HP444F | CAGCAGGTGCTAAACAACAA | [22] |
| HP444R | CGGTAGCTTAACGCCGAATA | ||
| MG | F | TTGGGTTTAGGGATTGGGATT | [23] |
| R | CCAAGGGATTCAACCATCTT | ||
| TaqMan probe | FAM-TGATGATCCAAGAACGTGAAGAACACC-BHQ2 | ||
| MS | F | CTAAATACAATAGCCCAAGGCAA | |
| R | CCTCCTTTCTTACGGAGTACA | ||
| TaqMan probe | FAM-AGCGATACACAACCGCTTTTAGAAT-BHQ1 | ||
| ORT | Ort 2F | GTCGCCGTAGTCATTAACCTCGTA | [24] |
| Ort 2R | CTGTAGCCGATCAGCGTTTGAATG | ||
| ILT | ICP4-1F | ACTGATAGCTTTTCGTACAGCACG | [15] |
| ICP4-1R | CATCGGGACATTCTCCAGGTAGCA | ||
| ICP4-2F | CTTCAGACTCCAGCTCATCTG | ||
| ICP4-2R | AGTCATGCGTCTATGGCGTTGAC |
| Case ID | No Examined Birds | Main Pathological Findings 1 | Results of PCR Testing 2 | |||||
|---|---|---|---|---|---|---|---|---|
| ILTV | IBV | APV | MG | MS | ORT | |||
| 686/17 | 2 | Edema of the conjunctivae and severe fibrinous conjunctivitis, sinusitis, tracheitis and pneumonia, fibrinous oophoritis, infestation with Ascaridia galli, Capillaria sp., and Dermanyssus gallinae (2/2) | + | + | + | + | + | + |
| 1482/17 | 2 | Hyperemia of the conjunctivae, serofibrinous sinusitis, hollow caseous cast in larynx and upper trachea, fibrinous to caseous tracheitis, fibrinous pneumonia (2/2) | + | + | nd | + | + | + |
| 1104/18 | 6 | Mild conjunctivitis (6/6), fibrinous oophoritis and peritonitis (egg peritonitis) (3/6), infestation with Ascaridia galli (6/6) | + | + | nd | + | + | − |
| 1315/18 | 5 | Conjunctivitis (5/5), serofibrinous sinusitis (5/5), fibrinous to caseous tracheitis (3/5), serofibrinous airsacculitis, peritonitis and oophoritis (3/5), infestation with Ascaridia galli and Capillaria sp. (2/5) | + | nd | − | − | + | nd |
| 1718/18 | 1 | Severe fibrinous conjunctivitis, yellow caseous exudate in beak cavity, hemorrhagic to caseous laryngitis and tracheitis, serofibrinous sinusitis (1/1) | + | nd | nd | + | + | + |
| 1477/19 | 4 | Hyperemia of conjunctivae (4/4), serofibrinous sinusitis (4/5), hollow caseous cast in larynx and upper trachea (3/4), fibrinous to caseous tracheitis (3/4) | + | + | nd | − | + | nd |
| 1560/19 | 6 | Conjunctivitis (5/6), serofibrinous sinusitis (5/6), fibrinous to caseous tracheitis (4/6), serofibrinous airsacculitis (5/6), fibrinous peritonitis and oophoritis (4/6), infestation with Ascaridia galli (3/6) | + | + | nd | − | + | nd |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zorman Rojs, O.; Dovč, A.; Krapež, U.; Žlabravec, Z.; Račnik, J.; Slavec, B. Detection of Laryngotracheitis Virus in Poultry Flocks with Respiratory Disorders in Slovenia. Viruses 2021, 13, 707. https://doi.org/10.3390/v13040707
Zorman Rojs O, Dovč A, Krapež U, Žlabravec Z, Račnik J, Slavec B. Detection of Laryngotracheitis Virus in Poultry Flocks with Respiratory Disorders in Slovenia. Viruses. 2021; 13(4):707. https://doi.org/10.3390/v13040707
Chicago/Turabian StyleZorman Rojs, Olga, Alenka Dovč, Uroš Krapež, Zoran Žlabravec, Joško Račnik, and Brigita Slavec. 2021. "Detection of Laryngotracheitis Virus in Poultry Flocks with Respiratory Disorders in Slovenia" Viruses 13, no. 4: 707. https://doi.org/10.3390/v13040707
APA StyleZorman Rojs, O., Dovč, A., Krapež, U., Žlabravec, Z., Račnik, J., & Slavec, B. (2021). Detection of Laryngotracheitis Virus in Poultry Flocks with Respiratory Disorders in Slovenia. Viruses, 13(4), 707. https://doi.org/10.3390/v13040707