
The Bank Vole (Clethrionomys glareolus)—Small Animal Model for Hepacivirus Infection

 , , , , , , and
, , , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Bank Voles
2.2. BvHV Inocula
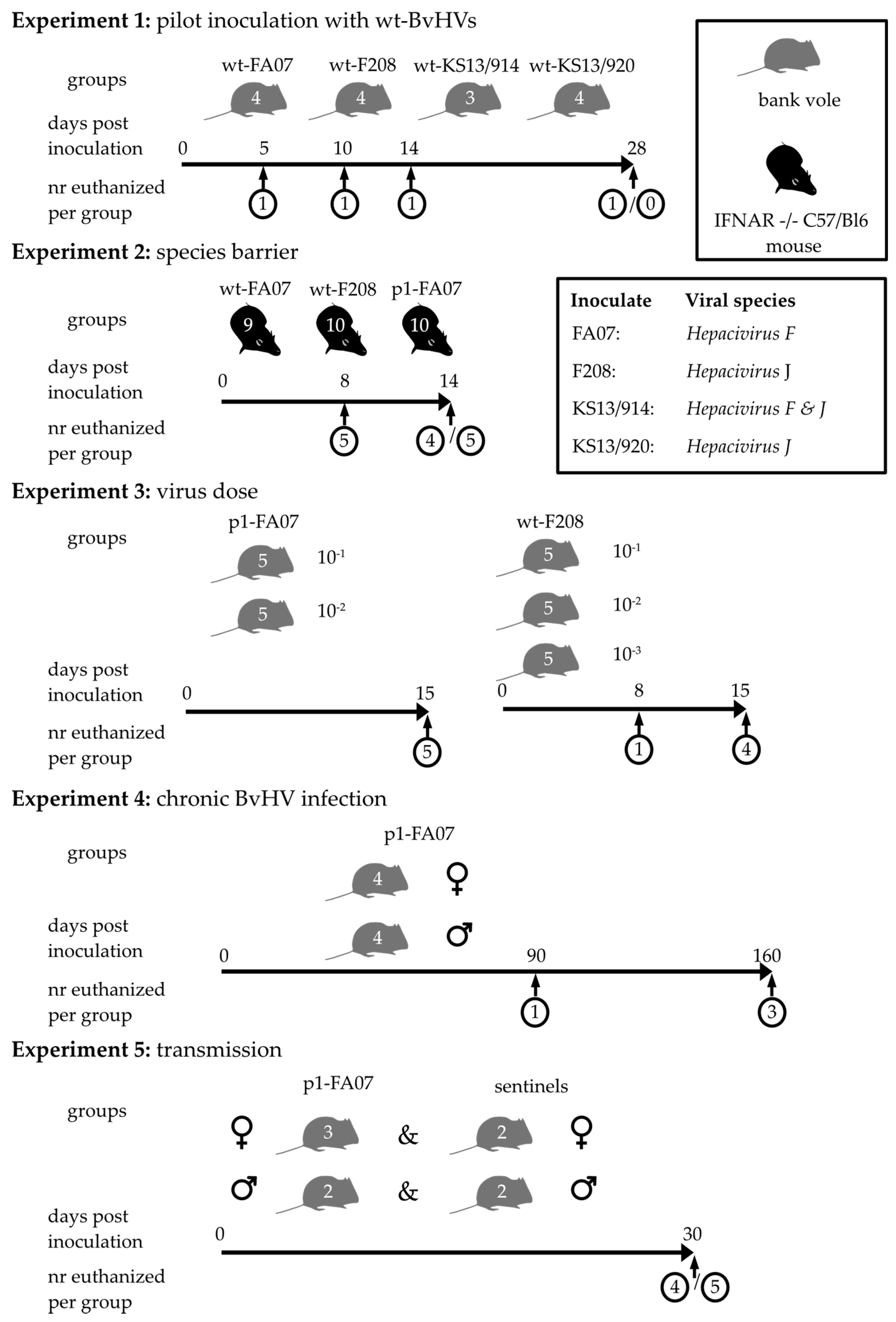
2.3. Experimental Design of Infection Studies
2.4. Sampling Procedure and RNA Extraction
2.5. RT-qPCR Detection of BvHV RNA
2.6. Virus Genome Sequencing
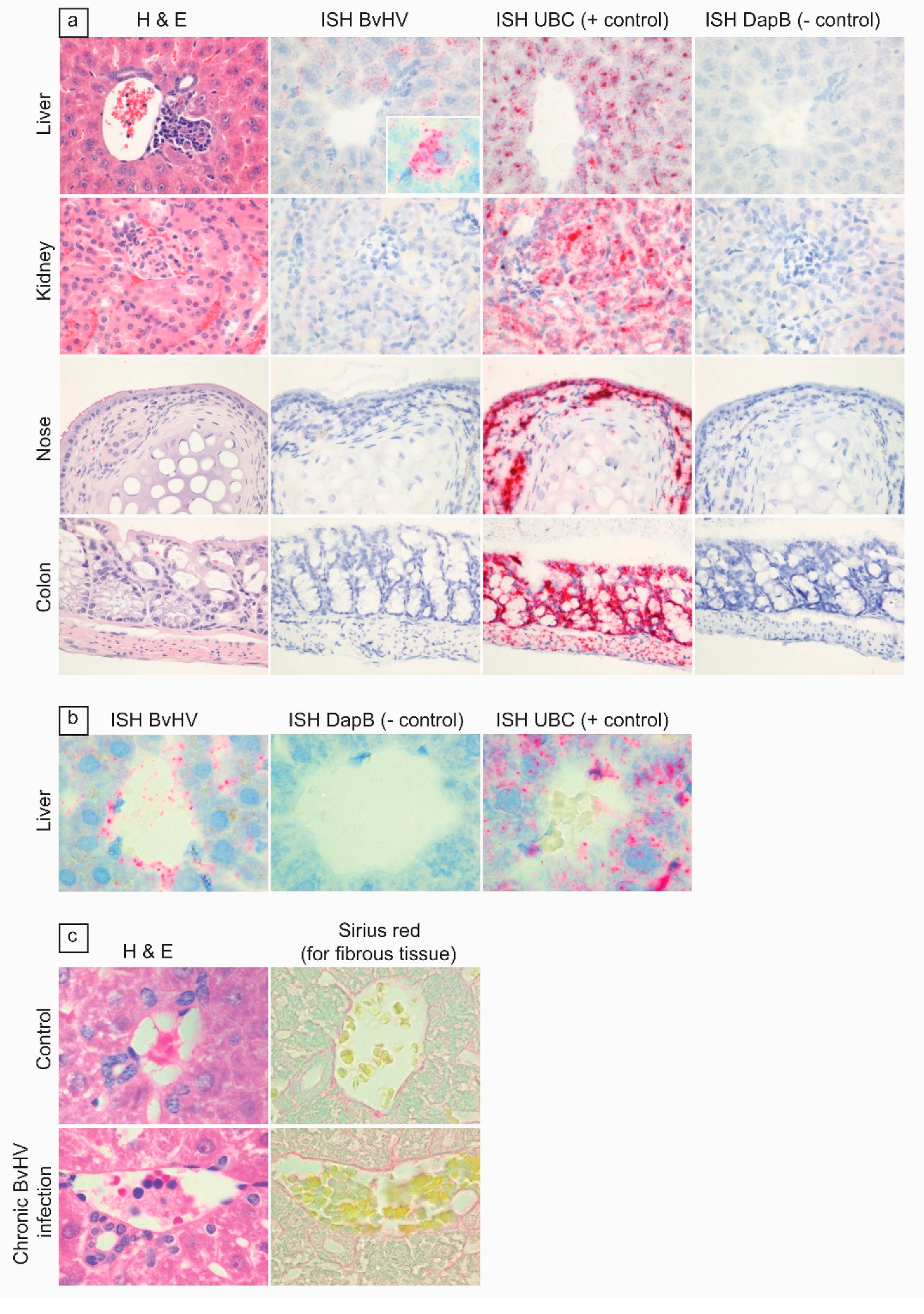
2.7. In Situ Hybridization and Microscopic Evaluation
2.8. Transmission Electron Microscopy
3. Results
3.1. Collection of BvHV-Positive Animals
3.2. Inoculation of Bank Voles with Blood and Visceral Cavity Lavage of Wild-Trapped Bank Voles—Inoculation Experiment 1
3.3. Inoculation of IFNAR-/- Mice with Liver Suspension of Bank Voles—Inoculation Experiment 2
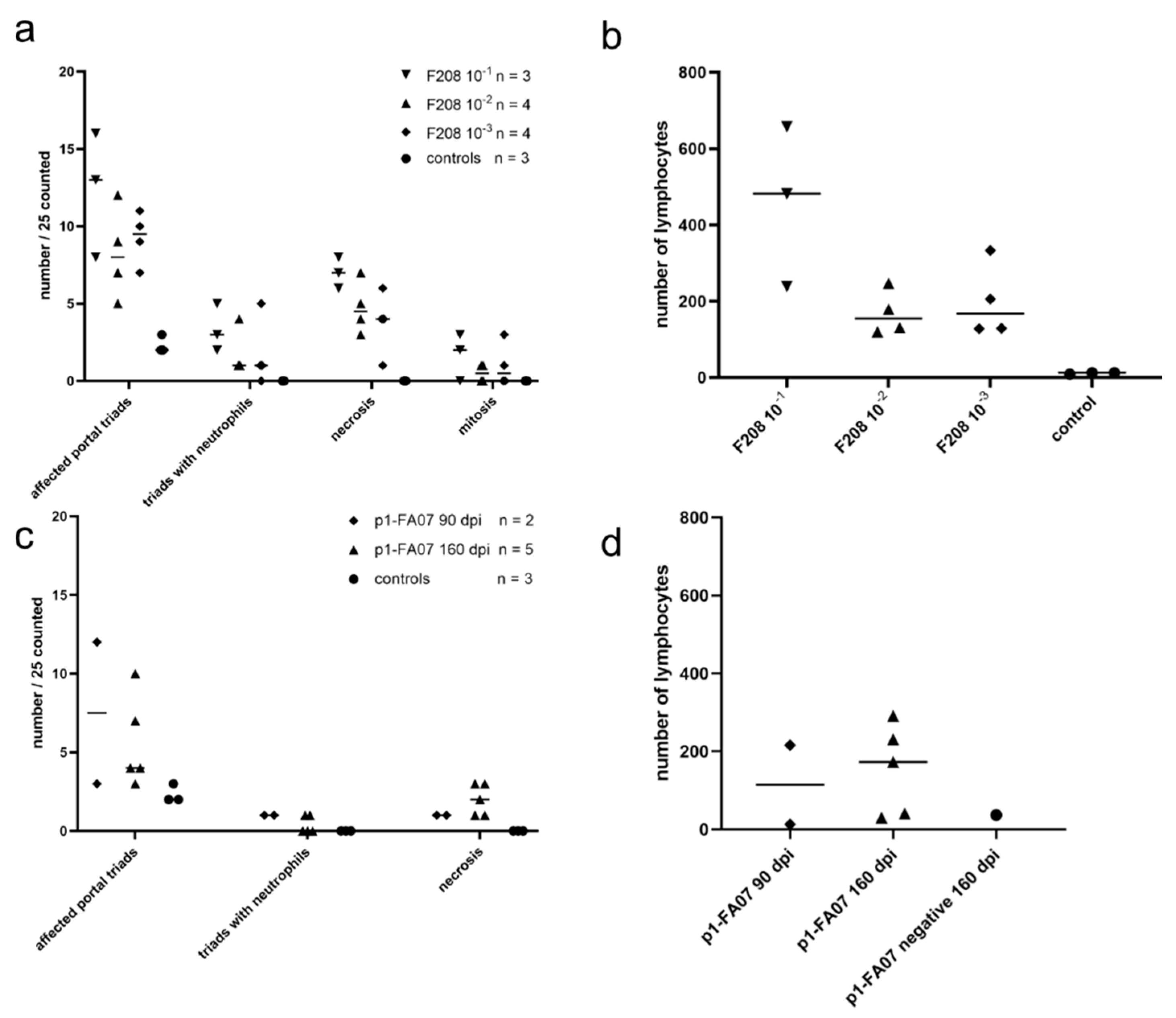
3.4. Bank Vole Infections with Different Dosages of wt and Bank Vole-Passaged BvHV—Inoculation Experiment 3
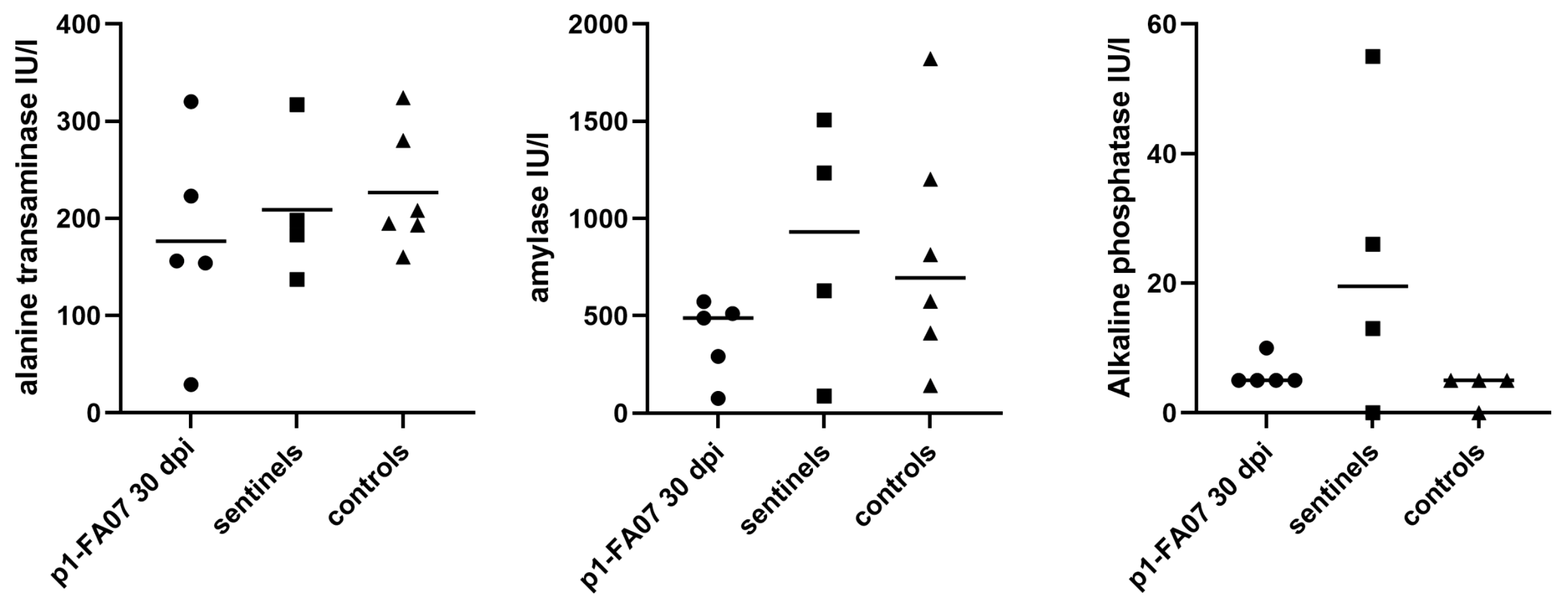
3.5. Long-Term Experimental BvHV Infection in Bank Voles—Inoculation Experiment 4
3.6. Potential Transmission Route of BvHV—Inoculation Experiment 5
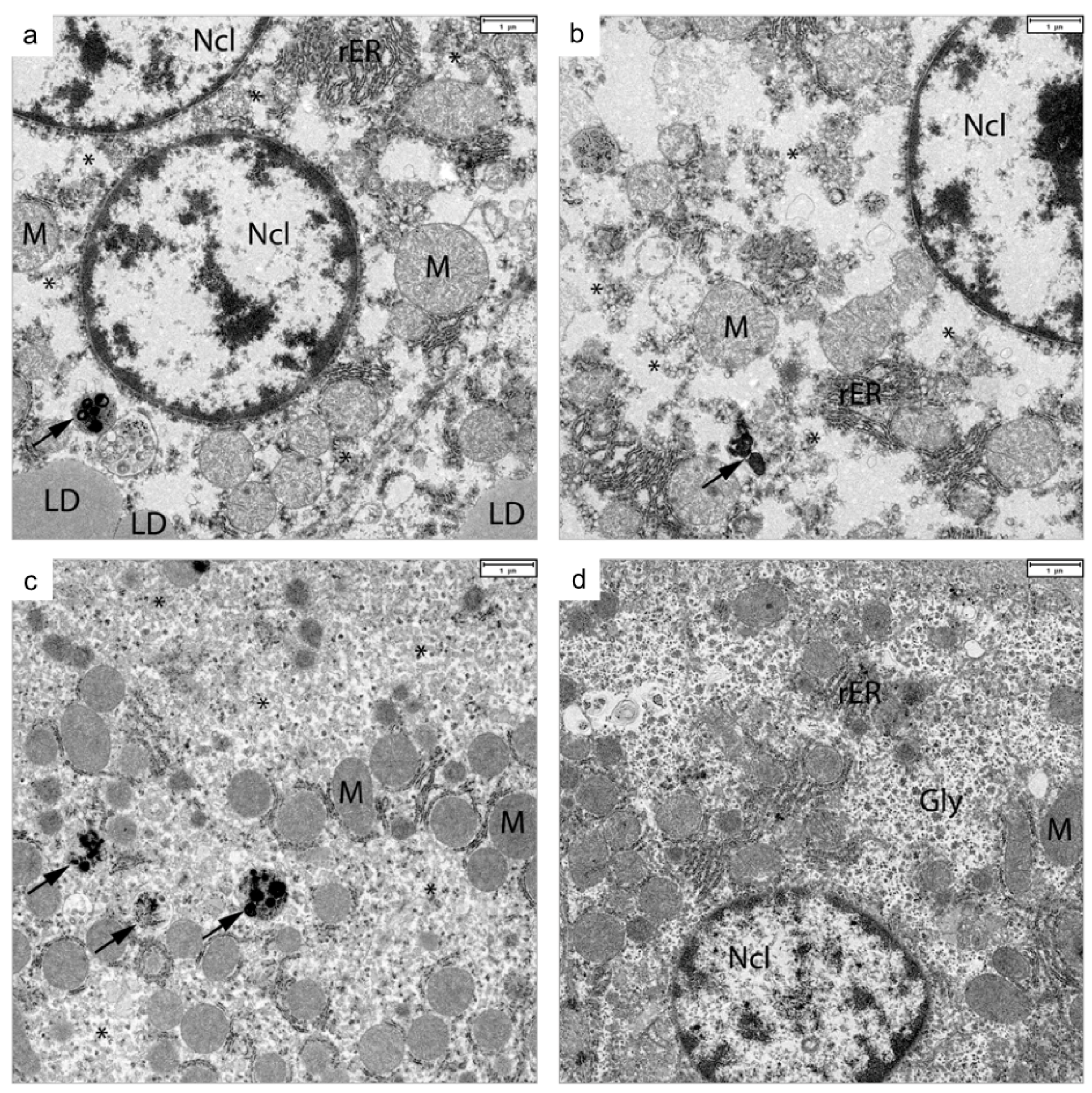
3.7. Electron Microscopy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gower, E.; Estes, C.; Blach, S.; Razavi-Shearer, K.; Razavi, H. Global epidemiology and genotype distribution of the hepatitis C virus infection. J. Hepatol. 2014, 61 (Suppl. 1), S45–S57. [Google Scholar] [CrossRef]
- Chung, R.T.; Baumert, T.F. Curing Chronic Hepatitis C—The Arc of a Medical Triumph. N. Engl. J. Med. 2014, 370, 1576–1578. [Google Scholar] [CrossRef] [PubMed]
- Lawitz, E.; Poordad, F.; Brainard, D.M.; Hyland, R.H.; An, D.; Dvory-Sobol, H.; Symonds, W.T.; McHutchison, J.G.; Membreno, F.E. Sofosbuvir with peginterferon-ribavirin for 12 weeks in previously treated patients with hepatitis C genotype 2 or 3 and cirrhosis. Hepatology 2015, 61, 769–775. [Google Scholar] [CrossRef]
- Najafzadeh, M.; Andersson, K.; Shrank, W.H.; Krumme, A.A.; Matlin, O.S.; Brennan, T.; Avorn, J.; Choudhry, N.K. Cost-effectiveness of novel regimens for the treatment of hepatitis c virus. Ann. Intern. Med. 2015, 162, 407–419. [Google Scholar] [CrossRef] [PubMed]
- Sharafi, H.; Maleki, S.; Alavian, S.M. Prevalence of hepatitis C virus NS5A resistance-associated substitutions in chronic infection with genotype 1: A pooled analysis based on deposited sequences in GenBank. Virus Res. 2018, 259, 54–61. [Google Scholar] [CrossRef]
- Hartlage, A.S.; Murthy, S.; Kumar, A.; Trivedi, S.; Dravid, P.; Sharma, H.; Walker, C.M.; Kapoor, A. Vaccination to prevent T cell subversion can protect against persistent hepacivirus infection. Nat. Commun. 2019, 10, 1113. [Google Scholar] [CrossRef] [PubMed]
- Bukh, J. Animal models for the study of hepatitis c virus infection and related liver disease. Gastroenterology 2012, 142, 1279–1287.e3. [Google Scholar] [CrossRef]
- Amako, Y.; Tsukiyama-Kohara, K.; Katsume, A.; Hirata, Y.; Sekiguchi, S.; Tobita, Y.; Hayashi, Y.; Hishima, T.; Funata, N.; Yonekawa, H.; et al. Pathogenesis of Hepatitis C Virus Infection in Tupaia belangeri. J. Virol. 2010, 84, 303–311. [Google Scholar] [CrossRef]
- Lauck, M.; Sibley, S.D.; Lara, J.; Purdy, M.A.; Khudyakov, Y.; Hyeroba, D.; Tumukunde, A.; Weny, G.; Switzer, W.M.; Chapman, C.A.; et al. A novel hepacivirus with an unusually long and intrinsically disordered ns5a protein in a wild old world primate. J. Virol. 2013, 87, 8971–8981. [Google Scholar] [CrossRef] [PubMed]
- Vercauteren, K.; de Jong, Y.P.; Meuleman, P. HCV animal models and liver disease. J. Hepatol. 2014, 61 (Suppl. 1), S26–S33. [Google Scholar] [CrossRef]
- Kapoor, A.; Simmonds, P.; Gerold, G.; Qaisar, N.; Jain, K.; Henriquez, J.A.; Firth, C.; Hirschberg, D.L.; Rice, C.M.; Shields, S.; et al. Characterization of a canine homolog of hepatitis C virus. Proc. Natl. Acad. Sci. USA 2011, 108, 11608–11613. [Google Scholar] [CrossRef] [PubMed]
- Baechlein, C.; Fischer, N.; Grundhoff, A.; Alawi, M.; Indenbirken, D.; Postel, A.; Baron, A.L.; Offinger, J.; Becker, K.; Beineke, A.; et al. Identification of a novel hepacivirus in domestic cattle from germany. J. Virol. 2015, 89, 7007–7015. [Google Scholar] [CrossRef]
- Corman, V.M.; Grundhoff, A.; Baechlein, C.; Fischer, N.; Gmyl, A.; Wollny, R.; Dei, D.; Ritz, D.; Binger, T.; Adankwah, E.; et al. Highly Divergent Hepaciviruses from African Cattle. J. Virol. 2015, 89, 5876–5882. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Walter, S.; Rasche, A.; Moreira-Soto, A.; Pfaender, S.; Bletsa, M.; Corman, V.M.; Aguilar-Setien, A.; García-Lacy, F.; Hans, A.; Todt, D.; et al. Differential infection patterns and recent evolutionary origins of equine hepaciviruses in donkeys. J. Virol. 2017, 91, e01711-16. [Google Scholar] [CrossRef]
- Pfaender, S.; Cavalleri, J.M.; Walter, S.; Doerrbecker, J.; Campana, B.; Brown, R.J.; Burbelo, P.D.; Postel, A.; Hahn, K.; Riebesehl, N.; et al. Clinical course of infection and viral tissue tropism of hepatitis C virus-like nonprimate hepaciviruses in horses. Hepatology 2015, 61, 447–459. [Google Scholar] [CrossRef]
- Ramsay, J.D.; Evanoff, R.; Wilkinson, T.E., Jr.; Divers, T.J.; Knowles, D.P.; Mealey, R.H. Experimental transmission of equine hepacivirus in horses as a model for hepatitis C virus. Hepatology 2015, 61, 1533–1546. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Jia, K.; Ping, X.; Huang, J.; Luo, A.; Wu, P.; Li, S. Novel bovine hepacivirus in dairy cattle, China. Emerg. Microbes Infect. 2018, 7, 1–3. [Google Scholar] [CrossRef]
- Van Nguyen, D.; Bonsall, D.; Ngo, T.T.; Carrique-Mas, J.; Pham, A.H.; Bryant, J.E.; Thwaites, G.; Baker, S.; Woolhouse, M.; Simmonds, P.; et al. Detection and characterization of homologues of human hepatitis viruses and pegiviruses in rodents and bats in Vietnam. Viruses 2018, 10, 102. [Google Scholar] [CrossRef]
- Bühler, S.; Bartenschlager, R. Promotion of hepatocellular carcinoma by hepatitis C virus. Dig. Dis. 2012, 30, 445–452. [Google Scholar] [CrossRef]
- Billerbeck, E.; Wolfisberg, R.; Fahnøe, U.; Xiao, J.W.; Quirk, C.; Luna, J.M.; Cullen, J.M.; Hartlage, A.S.; Chiriboga, L.; Ghoshal, K.; et al. Mouse models of acute and chronic hepacivirus infection. Science 2017, 357, 204–208. [Google Scholar] [CrossRef]
- Trivedi, S.; Murthy, S.; Sharma, H.; Hartlage, A.S.; Kumar, A.; Gadi, S.V.; Simmonds, P.; Chauhan, L.V.; Scheel, T.K.; Billerbeck, E.; et al. Viral persistence, liver disease, and host response in a hepatitis C–like virus rat model. Hepatology 2018, 68, 435–448. [Google Scholar] [CrossRef]
- Zingaretti, C.; De Francesco, R.; Abrignani, S. Why is it so difficult to develop a hepatitis C virus preventive vaccine? Clin. Microbiol. Infect. 2014, 20 (Suppl. 5), 103–109. [Google Scholar] [CrossRef] [PubMed]
- Atcheson, E.; Li, W.; Bliss, C.M.; Chinnakannan, S.; Heim, K.; Sharpe, H.; Hutchings, C.; Dietrich, I.; Nguyen, D.; Kapoor, A.; et al. Use of an outbred rat hepacivirus challenge model for design and evaluation of efficacy of different immunization strategies for hepatitis C Virus. Hepatology 2020, 71, 794–807. [Google Scholar] [CrossRef] [PubMed]
- Vandegrift, K.J.; Critchlow, J.T.; Kapoor, A.; Friedman, D.A.; Hudson, P.J. Peromyscus as a model system for human hepatitis C: An opportunity to advance our understanding of a complex host parasite system. Semin. Cell. Dev. Biol. 2017, 61, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Drexler, J.F.; Corman, V.M.; Müller, M.A.; Lukashev, A.N.; Gmyl, A.; Coutard, B.; Adam, A.; Ritz, D.; Leijten, L.M.; Van Riel, D.; et al. Evidence for novel hepaciviruses in rodents. PLoS Pathog. 2013, 9, e1003438. [Google Scholar] [CrossRef] [PubMed]
- Firth, C.; Bhat, M.; Firth, M.A.; Williams, S.; Frye, M.; Simmonds, P.; Conte, J.M.; Ng, J.; Garcia, J.; Bhuva, N.P.; et al. Detection of zoonotic pathogens and characterization of novel viruses carried by commensal rattus norvegicus in New York City. mBio 2014, 5, e01933-14. [Google Scholar] [CrossRef]
- Li, L.-L.; Liu, M.-M.; Shen, S.; Zhang, Y.-J.; Xu, Y.-L.; Deng, H.-Y.; Deng, F.; Duan, Z.-J. Detection and characterization of a novel hepacivirus in long-tailed ground squirrels (Spermophilus undulatus) in China. Arch. Virol. 2019, 164, 2401–2410. [Google Scholar] [CrossRef] [PubMed]
- International Committee on Taxonomy of Viruses. ICTV Virus Taxonomy: 2019 Release. Available online: https://talk.ictvonline.org/taxonomy/ (accessed on 3 September 2020).
- Schneider, J.; Hoffmann, B.; Fevola, C.; Schmidt, M.; Imholt, C.; Fischer, S.; Ecke, F.; Hörnfeldt, B.; Magnusson, M.; Olsson, G.; et al. Geographical distribution and genetic diversity of bank vole hepaciviruses in Europe. Viruses 2021, 13, 1258. [Google Scholar] [CrossRef] [PubMed]
- FELASA working group on revision of guidelines for health monitoring of rodents and rabbits; Mähler Convenor, M.; Berard, M.; Feinstein, R.; Gallagher, A.; Illgen-Wilcke, B.; Pritchett-Corning, K.; Raspa, M. FELASA recommendations for the health monitoring of mouse, rat, hamster, guinea pig and rabbit colonies in breeding and experimental units. Lab Anim. 2014, 48, 178–192. [Google Scholar] [PubMed]
- Schlegel, M.; Ali, H.S.; Stieger, N.; Groschup, M.H.; Wolf, R.; Ulrich, R.G. Molecular identification of small mammal species using novel cytochrome b gene-derived degenerated primers. Biochem. Genet. 2011, 50, 440–447. [Google Scholar] [CrossRef]
- Jacob, J.; Ulrich, R.G.; Freise, J.; Schmolz, E. Monitoring populations of rodent reservoirs of zoonotic diseases. Projects, aims and results. Bundesgesundheitsblatt Gesundh. Gesundh. 2014, 57, 511–518. (In German) [Google Scholar] [CrossRef] [PubMed]
- Ulrich, R.G.; Schmidt-Chanasit, J.; Schlegel, M.; Jacob, J.; Pelz, H.-J.; Mertens, M.; Wenk, M.; Büchner, T.; Masur, D.; Sevke, K.; et al. Network “Rodent-borne pathogens” in Germany: Longitudinal studies on the geographical distribution and prevalence of hantavirus infections. Parasitol. Res. 2008, 103 (Suppl. 1), 121–129. [Google Scholar] [CrossRef]
- Nainys, J.; Timinskas, A.; Schneider, J.; Ulrich, R.G.; Gedvilaite, A. Identification of two novel members of the tentative genus wukipolyomavirus in wild rodents. PLoS ONE 2015, 10, e0140916. [Google Scholar] [CrossRef] [PubMed]
- Mertens, M.; Kindler, E.; Emmerich, P.; Esser, J.; Wagner-Wiening, C.; Wölfel, R.; Petraityte-Burneikiene, R.; Schmidt-Chanasit, J.; Zvirbliene, A.; Groschup, M.H.; et al. Phylogenetic analysis of Puumala virus subtype Bavaria, characterization and diagnostic use of its recombinant nucleocapsid protein. Virus Genes 2011, 43, 177–191. [Google Scholar] [CrossRef]
- Höper, D.; Hoffmann, B.; Beer, M. A Comprehensive deep sequencing strategy for full-length genomes of influenza A. PLoS ONE 2011, 6, e19075. [Google Scholar] [CrossRef] [PubMed]
- Juozapaitis, M.; Moreira, É.A.; Mena, I.; Giese, S.; Riegger, D.; Pohlmann, A.; Höper, D.; Zimmer, G.; Beer, M.; García-Sastre, A.; et al. An infectious bat-derived chimeric influenza virus harbouring the entry machinery of an influenza A virus. Nat. Commun. 2014, 5, 4448. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Flanagan, J.; Su, N.; Wang, L.-C.; Bui, S.; Nielson, A.; Wu, X.; Vo, H.-T.; Ma, X.-J.; Luo, Y. RNAscope: A novel in situ RNA analysis platform for formalin-fixed, paraffin-embedded tissues. J. Mol. Diagn. 2012, 14, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Straub, B.K.; Herpel, E.; Singer, S.; Zimbelmann, R.; Breuhahn, K.; Macher-Goeppinger, S.; Warth, A.; Lehmann-Koch, J.; Longerich, T.; Heid, H.; et al. Lipid droplet-associated PAT-proteins show frequent and differential expression in neoplastic steatogenesis. Mod. Pathol. 2010, 23, 480–492. [Google Scholar] [CrossRef] [PubMed]
- Pawella, L.M.; Hashani, M.; Eiteneuer, E.; Renner, M.; Bartenschlager, R.; Schirmacher, P.; Straub, B.K. Perilipin discerns chronic from acute hepatocellular steatosis. J. Hepatol. 2014, 60, 633–642. [Google Scholar] [CrossRef]
- Boisvert, J.; Kunkel, E.J.; Campbell, J.J.; Keeffe, E.B.; Butcher, E.C.; Greenberg, H.B. Liver-infiltrating lymphocytes in end-stage hepatitis C virus: Subsets, activation status, and chemokine receptor phenotypes. J. Hepatol. 2003, 38, 67–75. [Google Scholar] [CrossRef]
- Pybus, O.G.; Markov, P.V.; Wu, A.; Tatem, A.J. Investigating the endemic transmission of the hepatitis C virus. Int. J. Parasitol. 2007, 37, 839–849. [Google Scholar] [CrossRef]
- Brey, I.R.; Merz, A.; Chiramel, A.; Lee, J.-Y.; Chlanda, P.; Haselman, U.; Santarella-Mellwig, R.; Habermann, A.; Hoppe, S.; Kallis, S.; et al. Three-dimensional architecture and biogenesis of membrane structures associated with hepatitis c virus replication. PLoS Pathog. 2012, 8, e1003056. [Google Scholar] [CrossRef]
- Buchalczyk, A. Reproduction, mortality and longevity of the bank vole under laboratory conditions. Acta Theriol. 1970, 13, 153–176. [Google Scholar] [CrossRef]
- Graham, C.S.; Baden, L.R.; Yu, E.; Mrus, J.M.; Carnie, J.; Heeren, T.; Koziel, M.J. Influence of human immunodeficiency virus infection on the course of hepatitis c virus infection: A meta-analysis. Clin. Infect. Dis. 2001, 33, 562–569. [Google Scholar] [CrossRef]
- Salmon-Ceron, D.; Lewden, C.; Morlat, P.; Bévilacqua, S.; Jougla, E.; Bonnet, F.; Héripret, L.; Costagliola, D.; May, T.; Chêne, G. Liver disease as a major cause of death among HIV infected patients: Role of hepatitis C and B viruses and alcohol. J. Hepatol. 2005, 42, 799–805. [Google Scholar] [CrossRef] [PubMed]
- Kage, M.; Shimamatu, K.; Nakashima, E.; Kojiro, M.; Inoue, O.; Yano, M. Long-term evolution of fibrosis from chronic hepatitis to cirrhosis in patients with hepatitis C: Morphometric analysis of repeated biopsies. Hepatology 1997, 25, 1028–1031. [Google Scholar] [CrossRef] [PubMed]
- Wiley, T.E.; McCarthy, M.; Breidi, L.; McCarthy, M.; Layden, T.J. Impact of alcohol on the histological and clinical progression of hepatitis C infection. Hepatology 1998, 28, 805–809. [Google Scholar] [CrossRef]
- Paul, D.; Bartenschlager, R. Hepatitis C virus’s next top models? Nat. Microbiol. 2016, 1, 15018. [Google Scholar] [CrossRef] [PubMed]
- Wolfisberg, R.; Holmbeck, K.; Nielsen, L.; Kapoor, A.; Rice, C.M.; Bukh, J.; Scheel, T.K.H. Replicons of a rodent hepatitis c model virus permit selection of highly permissive cells. J. Virol. 2019, 93, 19. [Google Scholar] [CrossRef]
- Neumann, A.U.; Lam, N.P.; Dahari, H.; Davidian, M.; Wiley, T.E.; Mika, B.P.; Perelson, A.S.; Layden, T.J. Differences in viral dynamics between genotypes 1 and 2 of hepatitis C virus. J. Infect. Dis. 2000, 182, 28–35. [Google Scholar] [CrossRef]
- Zein, N.N. Clinical significance of hepatitis C virus genotypes. Clin. Microbiol. Rev. 2000, 13, 223–235. [Google Scholar] [CrossRef]
- Farci, P.; Purcell, R.H. Clinical significance of hepatitis C virus genotypes and quasispecies. Semin. Liver Dis. 2000, 20, 103–126. [Google Scholar] [PubMed]
- Gerlach, J.; Diepolder, H.M.; Zachoval, R.; Gruener, N.H.; Jung, M.-C.; Ulsenheimer, A.; Schraut, W.W.; Schirren, C.; Waechtler, M.; Backmund, M.; et al. Acute hepatitis C: High rate of both spontaneous and treatment-induced viral clearance. Gastroenterology 2003, 125, 80–88. [Google Scholar] [CrossRef]
- Freeman, A.J.; Marinos, G.; Ffrench, R.A.; Lloyd, A.R. Immunopathogenesis of hepatitis C virus infection. Immunol. Cell Biol. 2001, 79, 515–536. [Google Scholar] [CrossRef] [PubMed]
- Darby, S.C.; Ewart, D.W.; Giangrande, P.L.; Spooner, R.J.; Rizza, C.R.; Dusheiko, G.M.; Lee, C.A.; Ludlam, C.A.; Preston, F.E. Mortality from liver cancer and liver disease in haemophilic men and boys in UK given blood products contaminated with hepatitis C. UK haemophilia centre directors‘ organisation. Lancet 1997, 350, 1425–1431. [Google Scholar] [CrossRef]
- Niederau, C.; Lange, S.; Heintges, T.; Erhardt, A.; Buschkamp, M.; Hürter, D.; Nawrocki, M.; Kruska, L.; Hensel, F.; Petry, W.; et al. Prognosis of chronic hepatitis C: Results of a large, prospective cohort study. Hepatology 1998, 28, 1687–1695. [Google Scholar] [CrossRef] [PubMed]
- Sosa-Jurado, F.; Hernández-Galindo, V.L.; Meléndez-Mena, D.; Mendoza-Torres, M.A.; Martínez-Arroniz, F.J.; Vallejo-Ruiz, V.; Reyes-Leyva, J.; Santos-López, G. Detection of hepatitis C virus RNA in saliva of patients with active infection not associated with periodontal or liver disease severity. BMC Infect. Dis. 2014, 14, 72. [Google Scholar] [CrossRef] [PubMed]
- Babik, J.M.; Cohan, D.; Monto, A.; Hartigan-O’Connor, D.J.; McCune, J.M. The human fetal immune response to hepatitis c virus exposure in utero. J. Infect. Dis. 2010, 203, 196–206. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Designation | Sequence |
|---|---|
| RHV-NS3-lin1_3.2-F | 5′-TGY TGC GAC AGC ACG GCA T |
| RHV-NS3-lin1_3.3-R | 5′-GCG TCC GGR ATT TTR CTC AC |
| RHV-NS3-lin1_3-FAM | 5′-CCG TYG CCT ACT ACC GAG GCG A |
| RHV-NS3-lin4_4.2-F | 5′-ATG ACG GGA TAC ACY GGG AA |
| RHV-NS3-lin4_4.3-R | 5′-CAT KGT SAC TTC ATA TTT GGG CAT |
| RHV-NS3-lin4_4-FAM | 5′-ACT CTG TGT ATG ACA GCT GCY TGA G |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Röhrs, S.; Begeman, L.; Straub, B.K.; Boadella, M.; Hanke, D.; Wernike, K.; Drewes, S.; Hoffmann, B.; Keller, M.; Drexler, J.F.; et al. The Bank Vole (Clethrionomys glareolus)—Small Animal Model for Hepacivirus Infection. Viruses 2021, 13, 2421. https://doi.org/10.3390/v13122421
Röhrs S, Begeman L, Straub BK, Boadella M, Hanke D, Wernike K, Drewes S, Hoffmann B, Keller M, Drexler JF, et al. The Bank Vole (Clethrionomys glareolus)—Small Animal Model for Hepacivirus Infection. Viruses. 2021; 13(12):2421. https://doi.org/10.3390/v13122421
Chicago/Turabian StyleRöhrs, Susanne, Lineke Begeman, Beate K. Straub, Mariana Boadella, Dennis Hanke, Kerstin Wernike, Stephan Drewes, Bernd Hoffmann, Markus Keller, Jan Felix Drexler, and et al. 2021. "The Bank Vole (Clethrionomys glareolus)—Small Animal Model for Hepacivirus Infection" Viruses 13, no. 12: 2421. https://doi.org/10.3390/v13122421
APA StyleRöhrs, S., Begeman, L., Straub, B. K., Boadella, M., Hanke, D., Wernike, K., Drewes, S., Hoffmann, B., Keller, M., Drexler, J. F., Drosten, C., Höper, D., Kuiken, T., Ulrich, R. G., & Beer, M. (2021). The Bank Vole (Clethrionomys glareolus)—Small Animal Model for Hepacivirus Infection. Viruses, 13(12), 2421. https://doi.org/10.3390/v13122421