
Top Three Strategies of ss(+)RNA Plant Viruses: Great Opportunists and Ecosystem Tuners with a Small Genome

 ,
, 


{kind=link}
Abstract
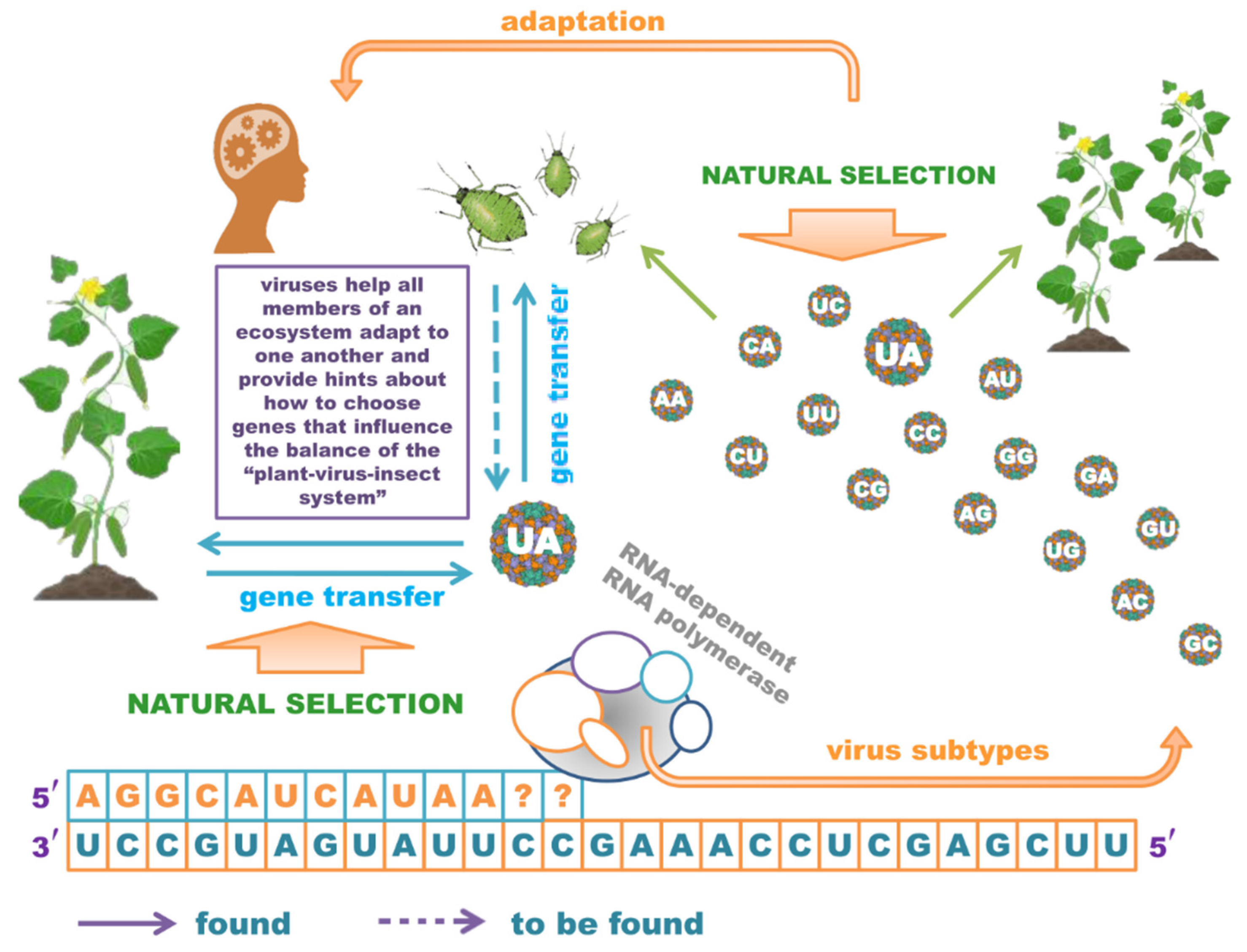
1. Introduction
2. Micro Boy Scouts: Always Ready to Translate
3. Never-Ending Treasures of Mutations
4. Tiny Ecosystem Tuners
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ledford, H. First known gene transfer from plant to insect identified. Nat. Cell Biol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Misof, B.; Liu, S.; Meusemann, K.; Peters, R.S.; Donath, A.; Mayer, C.; Frandsen, P.B.; Ware, J.; Flouri, T.; Beutel, R.G.; et al. Phylogenomics resolves the timing and pattern of insect evolution. Science 2014, 346, 763–767. [Google Scholar] [CrossRef]
- International Committee on Taxonomy of Viruses (ICTV). Available online: https://talk.ictvonline.org/ (accessed on 9 July 2020).
- Rampersad, S.; Tennant, P. Replication and Expression Strategies of Viruses. Viruses 2018, 55–82. [Google Scholar]
- Hofmann, C.; Sambade, A.; Heinlein, M. Plasmodesmata and intercellular transport of viral RNA. Biochem. Soc. Trans. 2007, 35, 142–145. [Google Scholar] [CrossRef]
- Miras, M.; Miller, W.A.; Truniger, V.; Aranda, M.A. Non-canonical Translation in Plant RNA Viruses. Front. Plant Sci. 2017, 8, 494. [Google Scholar] [CrossRef]
- Aiewsakun, P.; Katzourakis, A. Time-Dependent Rate Phenomenon in Viruses. J. Virol. 2016, 90, 7184–7195. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.A.; Giedroc, D.P. Ribosomal Frameshifting in Decoding Plant Viral RNAs. In Recoding: Expansion of Decoding Rules Enriches Gene Expression; Springer: New York, NY, USA, 2009; Volume 24, pp. 193–220. [Google Scholar]
- Belshaw, R.; Pybus, O.G.; Rambaut, A. The evolution of genome compression and genomic novelty in RNA viruses. Genome Res. 2007, 17, 1496–1504. [Google Scholar] [CrossRef]
- Wu, G.-W.; Cui, X.; Dai, Z.; He, R.; Li, Y.; Yu, K.; Bernards, M.; Chen, X.; Wang, A. A plant RNA virus hijacks endocytic proteins to establish its infection in plants. Plant J. 2019, 101, 384–400. [Google Scholar] [CrossRef] [PubMed]
- Vignuzzi, M.; Andino, R. Closing the gap: The challenges in converging theoretical, computational, experimental and real-life studies in virus evolution. Curr. Opin. Virol. 2012, 2, 515–518. [Google Scholar] [CrossRef]
- Pellett, P.E.; Mitra, S.; Holland, T.C. Basics of virology. Neurol. Rehabil. 2014, 123, 45–66. [Google Scholar] [CrossRef]
- Venkataraman, S.; Prasad, B.V.L.S.; Selvarajan, R. RNA Dependent RNA Polymerases: Insights from Structure, Function and Evolution. Viruses 2018, 10, 76. [Google Scholar] [CrossRef] [PubMed]
- Sanjuán, R.; Domingo-Calap, P. Mechanisms of viral mutation. Cell. Mol. Life Sci. 2016, 73, 4433–4448. [Google Scholar] [CrossRef] [PubMed]
- Elena, S.F.; Carrasco, P.; Daròs, J.-A.; Sanjuán, R. Mechanisms of genetic robustness in RNA viruses. EMBO Rep. 2006, 7, 168–173. [Google Scholar] [CrossRef]
- Mattenberger, F.; Latorre, V.; Tirosh, O.; Stern, A.; Geller, R. Globally defining the effects of mutations in a picornavirus capsid. eLife 2021, 10, e64256. [Google Scholar] [CrossRef]
- Shu, B.; Gong, P. Structural basis of viral RNA-dependent RNA polymerase catalysis and translocation. Proc. Natl. Acad. Sci. USA 2016, 113, E4005–E4014. [Google Scholar] [CrossRef]
- Roossinck, M.J. Plant RNA virus evolution. Curr. Opin. Microbiol. 2003, 6, 406–409. [Google Scholar] [CrossRef]
- Ahlquist, P.; Noueiry, A.O.; Lee, W.-M.; Kushner, D.B.; Dye, B.T. Host Factors in Positive-Strand RNA Virus Genome Replication. J. Virol. 2003, 77, 8181–8186. [Google Scholar] [CrossRef]
- Bao, X.; Roossinck, M.J. A life history view of mutualistic viral symbioses: Quantity or quality for cooperation? Curr. Opin. Microbiol. 2013, 16, 514–518. [Google Scholar] [CrossRef]
- Xu, P.; Chen, F.; Mannas, J.P.; Feldman, T.; Sumner, L.W.; Roossinck, M.J. Virus infection improves drought tolerance. New Phytol. 2008, 180, 911–921. [Google Scholar] [CrossRef]
- Van Molken, T.; De Caluwe, H.; Hordijk, C.A.; Leon-Reyes, A.; Snoeren, T.A.L.; van Dam, N.; Stuefer, J. Virus infection decreases the attractiveness of white clover plants for a non-vectoring herbivore. Oecologia 2012, 170, 433–444. [Google Scholar] [CrossRef]
- Lacroix, C.; Seabloom, E.; Borer, E. Environmental nutrient supply alters prevalence and weakens competitive interactions among coinfecting viruses. New Phytol. 2014, 204, 424–433. [Google Scholar] [CrossRef]
- Cui, J.; Holmes, E.C. Endogenous RNA viruses of plants in insect genomes. Virology 2012, 427, 77–79. [Google Scholar] [CrossRef] [PubMed]
- Marcon, H.S.; Costa-Silva, J.; Lorenzetti, A.P.R.; Marino, C.L.; Domingues, U.S. Genome-wide analysis of EgEVE_1, a transcriptionally active endogenous viral element associated to small RNAs in Eucalyptus genomes. Genet. Mol. Biol. 2017, 40, 217–225. [Google Scholar] [CrossRef]
- Simchuk, A.; Ivashov, A. Influence of Genetic Variation in Oak Leaf Roller Pupae and Their Host Plants on Body Sizes of Their Parasitoids, Itoplectis maculator (Fabricius, 1775). Psyche A J. Èntomol. 2011, 2011, 682572. [Google Scholar] [CrossRef][Green Version]
- Roossinck, M.J.; García-Arenal, F. Ecosystem simplification, biodiversity loss and plant virus emergence. Curr. Opin. Virol. 2015, 10, 56–62. [Google Scholar] [CrossRef]
- Oberemok, V.V. Method of Elimination of Phyllophagous Insects from Order Lepidoptera. Ukraine Patent UA 36445, 27 October 2008. [Google Scholar]
- Oberemok, V.V.; Laikova, K.V.; Repetskaya, A.I.; Kenyo, I.M.; Gorlov, M.V.; Kasich, I.N.; Krasnodubets, A.M.; Gal’Chinsky, N.V.; Fomochkina, I.I.; Zaitsev, A.S.; et al. A Half-Century History of Applications of Antisense Oligonucleotides in Medicine, Agriculture and Forestry: We Should Continue the Journey. Molecules 2018, 23, 1302. [Google Scholar] [CrossRef] [PubMed]
- Gal’chinsky, N.; Useinov, R.; Yatskova, E.; Laikova, K.; Novikov, I.; Gorlov, M.; Trikoz, N.; Sharmagiy, A.; Plugatar, Y.; Oberemok, V. A breakthrough in the efficiency of contact DNA insecticides: Rapid high mortality rates in the sap-sucking insects Dynaspidiotus britannicus Comstock and Unaspis euonymi Newstead. J. Plant Prot. Res. 2020, 60, 220–223. [Google Scholar] [CrossRef]
- Useinov, R.Z.; Gal’chinsky, N.; Yatskova, E.; Novikov, I.; Puzanova, Y.; Trikoz, N.; Sharmagiy, A.; Plugatar, Y.; Laikova, K.; Oberemok, V. To bee or not to bee: Creating DNA insecticides to replace non-selective organophosphate insecticides for use against the soft scale insect Ceroplastes japonicus Green. J. Plant Prot. Res. 2020, 60, 406–409. [Google Scholar] [CrossRef]
- Hughes, A.L. Evolution of inhibitors of apoptosis in baculoviruses and their insect hosts infection. Infect. Genet. Evol. 2002, 2, 3–10. [Google Scholar] [CrossRef]
- Clem, R.J.; Passarelli, A.L. Baculoviruses: Sophisticated Pathogens of Insects. PLoS Pathog. 2013, 9, e1003729. [Google Scholar] [CrossRef] [PubMed]
- Cerio, R.J.; Vandergaast, R.; Friesen, P.D. Host Insect Inhibitor-of-Apoptosis SfIAP Functionally Replaces Baculovirus IAP but Is Differentially Regulated by Its N-Terminal Leader. J. Virol. 2010, 84, 11448–11460. [Google Scholar] [CrossRef] [PubMed]
- Oberemok, V.V.; Laikova, K.V.; Zaitsev, A.S.; Shumskykh, M.N.; Kasich, I.N.; Gal’Chinsky, N.V.; Bekirova, V.V.; Makarov, V.V.; Agranovsky, A.A.; Gushchin, V.A.; et al. Molecular Alliance of Lymantria dispar Multiple Nucleopolyhedrovirus and a Short Unmodified Antisense Oligonucleotide of Its Anti-Apoptotic IAP-3 Gene: A Novel Approach for Gypsy Moth Control. Int. J. Mol. Sci. 2017, 18, 2446. [Google Scholar] [CrossRef] [PubMed]
- Oberemok, V.V.; Laikova, K.V.; Zaitsev, A.S.; Nyadar, P.M.; Gninenko, Y.I.; Gushchin, V.A.; Makarov, V.V.; Agranovsky, A.A. Original Article. Topical treatment of LdMNPV-infected gypsy moth caterpillars with 18 nucleotides long antisense fragment from LdMNPV IAP3 gene triggers higher levels of apoptosis in infected cells and mortality of the pest. J. Plant Prot. Res. 2016, 57, 18–24. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oberemok, V.V.; Puzanova, Y.V.; Kubyshkin, A.V.; Kamenetsky-Goldstein, R. Top Three Strategies of ss(+)RNA Plant Viruses: Great Opportunists and Ecosystem Tuners with a Small Genome. Viruses 2021, 13, 2304. https://doi.org/10.3390/v13112304
Oberemok VV, Puzanova YV, Kubyshkin AV, Kamenetsky-Goldstein R. Top Three Strategies of ss(+)RNA Plant Viruses: Great Opportunists and Ecosystem Tuners with a Small Genome. Viruses. 2021; 13(11):2304. https://doi.org/10.3390/v13112304
Chicago/Turabian StyleOberemok, Volodymyr V., Yelizaveta V. Puzanova, Anatoly V. Kubyshkin, and Rina Kamenetsky-Goldstein. 2021. "Top Three Strategies of ss(+)RNA Plant Viruses: Great Opportunists and Ecosystem Tuners with a Small Genome" Viruses 13, no. 11: 2304. https://doi.org/10.3390/v13112304
APA StyleOberemok, V. V., Puzanova, Y. V., Kubyshkin, A. V., & Kamenetsky-Goldstein, R. (2021). Top Three Strategies of ss(+)RNA Plant Viruses: Great Opportunists and Ecosystem Tuners with a Small Genome. Viruses, 13(11), 2304. https://doi.org/10.3390/v13112304