
The Past, the Present, and the Future of the Size Exclusion Chromatography in Extracellular Vesicles Separation



Abstract
1. Introduction
2. History of Size Exclusion Chromatography (SEC) and Commercially Available SEC Beads
3. Current SEC Methods for EV Separation
3.1. SEC in Separating Cell Culture Medium
3.2. Blood
3.3. Semen
3.4. Urine
3.5. Other Bodily Fluids
4. Discussion of the SEC Techniques Used for EV Separation
5. Future Directions: Towards a Particle Purification Liquid Chromatography System
5.1. Column Chemistry
5.2. Online Characterization
5.3. Fraction Collection
5.4. Additional Factors for Consideration
5.4.1. Pre and Post Column Concentrating Steps and Mobile Phase
5.4.2. Scalability
5.4.3. Biohazard Containment
6. Conclusions
7. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gage, S.H.; Fish, P.A. Fat digestion, absorption, and assimilation in man and animals as determined by the dark-field microscope, and a fat-soluble dye. Am. J. Anat. 1924, 34, 1–85. [Google Scholar] [CrossRef]
- Edmunds, J. Note on a New Paraboloid Illuminator for use beneath the Microscope Stage. Also Note on the Resolution of Podura Scale by means of the New Paraboloid. Mon. Microsc. J. 1877, 18, 78–85. [Google Scholar] [CrossRef][Green Version]
- Müller, F. Über einen bisher nicht beobachteten Formbestandteil des Blutes. Cent. f. Pathol. Anat 1896, 7, 529–539. [Google Scholar]
- Frazer, A.C.; Stewart, H.C. Ultramicroscopic particles in normal human blood. J. Physiol. 1937, 90, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Elkes, J.J.; Frazer, A.C.; Stewart, H.C. The composition of particles seen in normal human blood under dark-ground illumination. J. Physiol. 1939, 95, 68–82. [Google Scholar] [CrossRef] [PubMed]
- Chargaff, E.; West, R. The biological significance of the thromboplastic protein of blood. J. Biol. Chem. 1946, 166, 189–197. [Google Scholar] [CrossRef]
- Frailing, J.M.; Owen, C.I. Chylomicrons of Blood: A Resumé of Literature and a Recommended Counting Procedure. Am. J. Clin. Pathol. 1951, 21, 508–514. [Google Scholar] [CrossRef]
- Wolf, P. The Nature and Significance of Platelet Products in Human Plasma. Br. J. Haematol. 1967, 13, 269–288. [Google Scholar] [CrossRef]
- Abels, E.R.; Breakefield, X.O. Introduction to Extracellular Vesicles: Biogenesis, RNA Cargo Selection, Content, Release, and Uptake. Cell. Mol. Neurobiol. 2016, 36, 301–312. [Google Scholar] [CrossRef]
- Kalra, H.; Drummen, G.P.; Mathivanan, S. Focus on extracellular vesicles: Introducing the next small big thing. Int. J. Mol. Sci. 2016, 17, 170. [Google Scholar] [CrossRef]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Araldi, E.; Krämer-Albers, E.-M.; Hoen, E.N.-t.; Peinado, H.; Psonka-Antonczyk, K.M.; Rao, P.; van Niel, G.; Yáñez-Mó, M.; Nazarenko, I. International Society for Extracellular Vesicles: First annual meeting, April 17–21, 2012: ISEV-2012. J. Extracell. Vesicles 2012, 1, 19995. [Google Scholar] [CrossRef]
- Lötvall, J.; Rajendran, L.; Gho, Y.-S.; Thery, C.; Wauben, M.; Raposo, G.; Sjöstrand, M.; Taylor, D.; Telemo, E.; Breakefield, X.O. The launch of Journal of Extracellular Vesicles (JEV), the official journal of the International Society for Extracellular Vesicles-about microvesicles, exosomes, ectosomes and other extracellular vesicles. J. Extracell. Vesicles 2012, 1. [Google Scholar] [CrossRef] [PubMed]
- Ainsztein, A.M.; Brooks, P.J.; Dugan, V.G.; Ganguly, A.; Guo, M.; Howcroft, T.K.; Kelley, C.A.; Kuo, L.S.; Labosky, P.A.; Lenzi, R. The NIH extracellular RNA communication consortium. J. Extracell. Vesicles 2015, 4, 27493. [Google Scholar] [CrossRef]
- Laurent, L.C.; Abdel-Mageed, A.B.; Adelson, P.D.; Arango, J.; Balaj, L.; Breakefield, X.; Carlson, E.; Carter, B.S.; Majem, B.; Chen, C.C.; et al. Meeting report: Discussions and preliminary findings on extracellular RNA measurement methods from laboratories in the NIH Extracellular RNA Communication Consortium. J. Extracell. Vesicles 2015, 4, 26533. [Google Scholar] [CrossRef]
- Witwer, K.W.; Buzás, E.I.; Bemis, L.T.; Bora, A.; Lässer, C.; Lötvall, J.; Nolte-‘t Hoen, E.N.; Piper, M.G.; Sivaraman, S.; Skog, J.; et al. Standardization of sample collection, isolation and analysis methods in extracellular vesicle research. J. Extracell. Vesicles 2013, 2, 20360. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.F.; Pegtel, D.M.; Lambertz, U.; Leonardi, T.; O’driscoll, L.; Pluchino, S.; Ter-Ovanesyan, D.; Nolte-‘t Hoen, E.N. ISEV position paper: Extracellular vesicle RNA analysis and bioinformatics. J. Extracell. Vesicles 2013, 2, 22859. [Google Scholar] [CrossRef]
- Lener, T.; Gimona, M.; Aigner, L.; Börger, V.; Buzas, E.; Camussi, G.; Chaput, N.; Chatterjee, D.; Court, F.A.; Portillo, H.A.d.; et al. Applying extracellular vesicles based therapeutics in clinical trials–an ISEV position paper. J. Extracell. Vesicles 2015, 4, 30087. [Google Scholar] [CrossRef]
- Mateescu, B.; Kowal, E.J.; van Balkom, B.W.; Bartel, S.; Bhattacharyya, S.N.; Buzás, E.I.; Buck, A.H.; de Candia, P.; Chow, F.W.; Das, S. Obstacles and opportunities in the functional analysis of extracellular vesicle RNA–an ISEV position paper. J. Extracell. Vesicles 2017, 6, 1286095. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Coumans, F.A.; Brisson, A.R.; Buzas, E.I.; Dignat-George, F.; Drees, E.E.; El-Andaloussi, S.; Emanueli, C.; Gasecka, A.; Hendrix, A.; Hill, A.F. Methodological guidelines to study extracellular vesicles. Circ. Res. 2017, 120, 1632–1648. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, C.; Vizio, D.D.; Sahoo, S.; Théry, C.; Witwer, K.W.; Wauben, M.; Hill, A.F. Techniques used for the isolation and characterization of extracellular vesicles: Results of a worldwide survey. J. Extracell. Vesicles 2016, 5, 32945. [Google Scholar] [CrossRef] [PubMed]
- Linares, R.; Tan, S.; Gounou, C.; Arraud, N.; Brisson, A.R. High-speed centrifugation induces aggregation of extracellular vesicles. J. Extracell. Vesicles 2015, 4, 29509. [Google Scholar] [CrossRef] [PubMed]
- Shao, H.; Im, H.; Castro, C.M.; Breakefield, X.; Weissleder, R.; Lee, H. New Technologies for Analysis of Extracellular Vesicles. Chem. Rev. 2018, 118, 1917–1950. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Freitas, D.; Kim, H.S.; Fabijanic, K.; Li, Z.; Chen, H.; Mark, M.T.; Molina, H.; Martin, A.B.; Bojmar, L. Identification of distinct nanoparticles and subsets of extracellular vesicles by asymmetric flow field-flow fractionation. Nat. Cell Biol. 2018, 20, 332–343. [Google Scholar] [CrossRef] [PubMed]
- Shao, H.; Chung, J.; Balaj, L.; Charest, A.; Bigner, D.D.; Carter, B.S.; Hochberg, F.H.; Breakefield, X.O.; Weissleder, R.; Lee, H. Protein typing of circulating microvesicles allows real-time monitoring of glioblastoma therapy. Nat. Med. 2012, 18, 1835. [Google Scholar] [CrossRef]
- Arkesteijn, G.; Nauwelaers, F.; van den Engh, G.; Wauben, M. Prerequisites for the analysis and sorting of extracellular vesicle subpopulations by high-resolution flow cytometry. Cytometry. Part A J. Int. Soc. Anal. Cytol. 2015, 89, 135–147. [Google Scholar]
- Kormelink, T.G.; Arkesteijn, G.J.; Nauwelaers, F.A.; van den Engh, G.; Nolte-’t Hoen, E.N.; Wauben, M.H. Prerequisites for the analysis and sorting of extracellular vesicle subpopulations by high-resolution flow cytometry. Cytom. Part A 2016, 89, 135–147. [Google Scholar] [CrossRef]
- Tian, Y.; Ma, L.; Gong, M.; Su, G.; Zhu, S.; Zhang, W.; Wang, S.; Li, Z.; Chen, C.; Li, L. Protein profiling and sizing of extracellular vesicles from colorectal cancer patients via flow cytometry. ACS Nano 2018, 12, 671–680. [Google Scholar] [CrossRef]
- Choi, D.; Montermini, L.; Jeong, H.; Sharma, S.; Meehan, B.; Rak, J. Mapping subpopulations of cancer cell-derived extracellular vesicles and particles by nano-flow cytometry. ACS Nano 2019, 13, 10499–10511. [Google Scholar] [CrossRef]
- van der Pol, E.; de Rond, L.; Coumans, F.A.; Gool, E.L.; Böing, A.N.; Sturk, A.; Nieuwland, R.; van Leeuwen, T.G. Absolute sizing and label-free identification of extracellular vesicles by flow cytometry. Nanomed. Nanotechnol. Biol. Med. 2018, 14, 801–810. [Google Scholar] [CrossRef]
- Clayton, A.; Court, J.; Navabi, H.; Adams, M.; Mason, M.D.; Hobot, J.A.; Newman, G.R.; Jasani, B. Analysis of antigen presenting cell derived exosomes, based on immuno-magnetic isolation and flow cytometry. J. Immunol. Methods 2001, 247, 163–174. [Google Scholar] [CrossRef]
- Monguió-Tortajada, M.; Gálvez-Montón, C.; Bayes-Genis, A.; Roura, S.; Borràs, F.E. Extracellular vesicle isolation methods: Rising impact of size-exclusion chromatography. Cell. Mol. Life Sci. 2019, 76, 2369–2382. [Google Scholar] [CrossRef]
- Gholizadeh, S.; Draz, M.S.; Zarghooni, M.; Sanati-Nezhad, A.; Ghavami, S.; Shafiee, H.; Akbari, M. Microfluidic approaches for isolation, detection, and characterization of extracellular vesicles: Current status and future directions. Biosens. Bioelectron. 2017, 91, 588–605. [Google Scholar] [CrossRef]
- Théry, C.; Amigorena, S.; Raposo, G.; Clayton, A. Isolation and Characterization of Exosomes from Cell Culture Supernatants and Biological Fluids. Curr. Protoc. Cell Biol. 2006, 30, 3–22. [Google Scholar] [CrossRef]
- Zhang, H.; Lyden, D. Asymmetric-flow field-flow fractionation technology for exomere and small extracellular vesicle separation and characterization. Nat. Protoc. 2019, 14, 1027–1053. [Google Scholar] [CrossRef] [PubMed]
- Gordon, A.H.; Martin, A.J.; Synge, R.L. Partition chromatography in the study of protein constituents. Biochem. J. 1943, 37, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Gordon, A.; Martin, A.; Synge, R. Technical notes on the partition chromatography of acetamino-acids with silica gel. Biochem. J. 1944, 38, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; Synge, R. Separation of the higher monoamino-acids by counter-current liquid-liquid extraction: The amino-acid composition of wool. Biochem. J. 1941, 35, 91–121. [Google Scholar] [CrossRef]
- Ettre, L.S. The birth of partition chromatography. LCGC N. Am. 2001, 19, 506–512. [Google Scholar]
- Lindqvist, B.; StorgÅRds, T. Molecular-sieving Properties of Starch. Nature 1955, 175, 511–512. [Google Scholar] [CrossRef]
- Porath, J.; Flodin, P.E.R. Gel Filtration: A Method for Desalting and Group Separation. Nature 1959, 183, 1657–1659. [Google Scholar] [CrossRef]
- Belew, M.; Porath, J.; Fohlman, J.; Janson, J.-C. Adsorption phenomena on sephacryl® S-200 superfine. J. Chromatogr. A 1978, 147, 205–212. [Google Scholar] [CrossRef]
- Morgan-Capner, P.; Davies, E.; Pattison, J. Rubella-specific IgM detection using Sephacryl S-300 gel filtration. J. Clin. Pathol. 1980, 33, 1082–1085. [Google Scholar] [CrossRef]
- Morgan, G.; Ramsden, D.B. Comparison of the gel chromatographic properties of Sephacryl S-200 Superfine and Sephadex G-150. J. Chromatogr. A 1978, 161, 319–323. [Google Scholar] [CrossRef]
- Iverius, P.-H. Coupling of glycosaminoglycans to agarose beads (Sepharose 4B). Biochem. J. 1971, 124, 677–683. [Google Scholar] [CrossRef]
- Thompson, S.T.; Cass, K.H.; Stellwagen, E. Blue dextran-sepharose: An affinity column for the dinucleotide fold in proteins. Proc. Natl. Acad. Sci. USA 1975, 72, 669–672. [Google Scholar] [CrossRef] [PubMed]
- Ogata, S.-I.; Muramatsu, T.; Kobata, A. Fractionation of Glycopeptides by Affinity Column Chromatography on Concanavalin A-Sepharose1. J. Biochem. 1975, 78, 687–696. [Google Scholar] [CrossRef]
- Okada, N.; Shindo-Okada, N.; Nishimura, S. Isolation of mammalian tRNAAsp and tRNATyr by lectin-Sepharose affinity column chromatography. Nucleic Acids Res. 1977, 4, 415–423. [Google Scholar] [CrossRef]
- Weisgraber, K.H.; Mahley, R.W. Subfractionation of human high density lipoproteins by heparin-Sepharose affinity chromatography. J. Lipid Res. 1980, 21, 316–325. [Google Scholar] [CrossRef]
- Hauschka, P.; Mavrakos, A.; Iafrati, M.; Doleman, S.; Klagsbrun, M. Growth factors in bone matrix. Isolation of multiple types by affinity chromatography on heparin-Sepharose. J. Biol. Chem. 1986, 261, 12665–12674. [Google Scholar] [CrossRef]
- Hjertén, S. A new method for preparation of agarose for gel electrophoresis. Biochim. Biophys. Acta 1962, 62, 445–449. [Google Scholar] [CrossRef]
- Hjertén, S.; Mosbach, R. “Molecular-sieve” chromatography of proteins on columns of cross-linked polyacrylamide. Anal. Biochem. 1962, 3, 109–118. [Google Scholar] [CrossRef]
- Draus, F. Separation of a cat submaxillary mucoid on Biogel. Arch. Oral Biol. 1965, 10, 719–721. [Google Scholar] [CrossRef]
- Jos, J.; Charbonnier, L.; Mosse, J.; Olives, J.; De Tand, M.-F.; Rey, J. The toxic fraction of gliadin digests in coeliac disease. Isolation by chromatography on Biogel P-10. Clin. Chim. Acta 1982, 119, 263–274. [Google Scholar] [CrossRef]
- Van der Westhuyzen, D.; Böhm, E.; Von Holt, C. Fractionation of chicken erythrocyte whole histone into the six main components by gel exclusion chromatography. Biochim. Et Biophys. Acta (BBA)-Protein Struct. 1974, 359, 341–345. [Google Scholar] [CrossRef]
- Huang, H.-C.; Cole, R.D. Fractionation of chromatin fragments on columns of Biogel A50-m at different salt concentrations. FEBS Lett. 1985, 183, 270–274. [Google Scholar] [CrossRef]
- Takemoto, L.J.; Azari, P. Isolation and characterization of covalently linked, high molecular weight proteins from human cataractous lens. Exp. Eye Res. 1977, 24, 63–70. [Google Scholar] [CrossRef]
- Meehan, B.; Rak, J.; Di Vizio, D. Oncosomes–large and small: What are they, where they came from? J. Extracell. Vesicles 2016, 5, 33109. [Google Scholar] [CrossRef] [PubMed]
- Witwer, K.W.; Théry, C. Extracellular vesicles or exosomes? On primacy, precision, and popularity influencing a choice of nomenclature. J. Extracell. Vesicles 2019, 8, 1648167. [Google Scholar] [CrossRef]
- Foster, B.; Balassa, T.; Benen, T.; Dominovic, M.; Elmadjian, G.; Florova, V.; Fransolet, M.; Kestlerova, A.; Kmiecik, G.; Kostadinova, I. Extracellular vesicles in blood, milk and body fluids of the female and male urogenital tract and with special regard to reproduction. Crit. Rev. Clin. Lab. Sci. 2016, 53, 379–395. [Google Scholar] [CrossRef]
- Kaddour, H.; Lyu, Y.; Welch, J.L.; Paromov, V.; Mandape, S.N.; Sakhare, S.S.; Pandhare, J.; Stapleton, J.T.; Pratap, S.; Dash, C. Proteomics profiling of autologous blood and semen exosomes from HIV-infected and uninfected individuals reveals compositional and functional variabilities. Mol. Cell. Proteom. 2020, 19, 78–100. [Google Scholar] [CrossRef]
- Dixon, C.L.; Sheller-Miller, S.; Saade, G.R.; Fortunato, S.J.; Lai, A.; Palma, C.; Guanzon, D.; Salomon, C.; Menon, R. Amniotic Fluid Exosome Proteomic Profile Exhibits Unique Pathways of Term and Preterm Labor. Endocrinology 2018, 159, 2229–2240. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Mitsialis, S.A.; Aslam, M.; Vitali Sally, H.; Vergadi, E.; Konstantinou, G.; Sdrimas, K.; Fernandez-Gonzalez, A.; Kourembanas, S. Exosomes Mediate the Cytoprotective Action of Mesenchymal Stromal Cells on Hypoxia-Induced Pulmonary Hypertension. Circulation 2012, 126, 2601–2611. [Google Scholar] [CrossRef] [PubMed]
- Hajj, G.N.M.; Arantes, C.P.; Dias, M.V.S.; Roffé, M.; Costa-Silva, B.; Lopes, M.H.; Porto-Carreiro, I.; Rabachini, T.; Lima, F.R.; Beraldo, F.H.; et al. The unconventional secretion of stress-inducible protein 1 by a heterogeneous population of extracellular vesicles. Cell. Mol. Life Sci. 2013, 70, 3211–3227. [Google Scholar] [CrossRef]
- Redzic, J.S.; Kendrick, A.A.; Bahmed, K.; Dahl, K.D.; Pearson, C.G.; Robinson, W.A.; Robinson, S.E.; Graner, M.W.; Eisenmesser, E.Z. Extracellular Vesicles Secreted from Cancer Cell Lines Stimulate Secretion of MMP-9, IL-6, TGF-β1 and EMMPRIN. PLoS ONE 2013, 8, e71225. [Google Scholar] [CrossRef]
- Nordin, J.Z.; Lee, Y.; Vader, P.; Mäger, I.; Johansson, H.J.; Heusermann, W.; Wiklander, O.P.B.; Hällbrink, M.; Seow, Y.; Bultema, J.J.; et al. Ultrafiltration with size-exclusion liquid chromatography for high yield isolation of extracellular vesicles preserving intact biophysical and functional properties. Nanomed. Nanotechnol. Biol. Med. 2015, 11, 879–883. [Google Scholar] [CrossRef]
- Willms, E.; Johansson, H.J.; Mäger, I.; Lee, Y.; Blomberg, K.E.M.; Sadik, M.; Alaarg, A.; Smith, C.I.E.; Lehtiö, J.; El Andaloussi, S.; et al. Cells release subpopulations of exosomes with distinct molecular and biological properties. Sci. Rep. 2016, 6, 22519. [Google Scholar] [CrossRef]
- Mol, E.A.; Goumans, M.-J.; Doevendans, P.A.; Sluijter, J.P.G.; Vader, P. Higher functionality of extracellular vesicles isolated using size-exclusion chromatography compared to ultracentrifugation. Nanomed. Nanotechnol. Biol. Med. 2017, 13, 2061–2065. [Google Scholar] [CrossRef]
- Benedikter, B.J.; Bouwman, F.G.; Vajen, T.; Heinzmann, A.C.A.; Grauls, G.; Mariman, E.C.; Wouters, E.F.M.; Savelkoul, P.H.; Lopez-Iglesias, C.; Koenen, R.R.; et al. Ultrafiltration combined with size exclusion chromatography efficiently isolates extracellular vesicles from cell culture media for compositional and functional studies. Sci. Rep. 2017, 7, 15297. [Google Scholar] [CrossRef] [PubMed]
- Corso, G.; Mäger, I.; Lee, Y.; Görgens, A.; Bultema, J.; Giebel, B.; Wood, M.J.A.; Nordin, J.Z.; Andaloussi, S.E.L. Reproducible and scalable purification of extracellular vesicles using combined bind-elute and size exclusion chromatography. Sci. Rep. 2017, 7, 11561. [Google Scholar] [CrossRef] [PubMed]
- Lobb, R.; Möller, A. Size exclusion chromatography: A simple and reliable method for exosome purification. In Extracellular Vesicles; Springer: Berlin/Heidelberg, Germany, 2017; pp. 105–110. [Google Scholar]
- Taylor, D.D.; Gerçel-Taylor, Ç.; Lyons, K.S.; Stanson, J.; Whiteside, T.L. T-Cell Apoptosis and Suppression of T-Cell Receptor/CD3-ζ by Fas Ligand-Containing Membrane Vesicles Shed from Ovarian Tumors. Clin. Cancer Res. 2003, 9, 5113–5119. [Google Scholar] [PubMed]
- Kim, J.W.; Wieckowski, E.; Taylor, D.D.; Reichert, T.E.; Watkins, S.; Whiteside, T.L. Fas ligand–positive membranous vesicles isolated from sera of patients with oral cancer induce apoptosis of activated T lymphocytes. Clin. Cancer Res. 2005, 11, 1010–1020. [Google Scholar] [PubMed]
- Taylor, D.D.; Zacharias, W.; Gercel-Taylor, C. Exosome Isolation for Proteomic Analyses and RNA Profiling. In Serum/Plasma Proteomics: Methods and Protocols; Simpson, R.J., Greening, D.W., Eds.; Humana Press: Totowa, NJ, USA, 2011; pp. 235–246. [Google Scholar]
- Shu, S.L.; Yang, Y.; Allen, C.L.; Hurley, E.; Tung, K.H.; Minderman, H.; Wu, Y.; Ernstoff, M.S. Purity and yield of melanoma exosomes are dependent on isolation method. J. Extracell. Vesicles 2020, 9, 1692401. [Google Scholar] [CrossRef]
- Li, M.; Lou, D.; Chen, J.; Shi, K.; Wang, Y.; Zhu, Q.; Liu, F.; Zhang, Y. Deep dive on the proteome of salivary extracellular vesicles: Comparison between ultracentrifugation and polymer-based precipitation isolation. Anal. Bioanal. Chem. 2021, 413, 365–375. [Google Scholar] [CrossRef]
- Van Deun, J.; Mestdagh, P.; Sormunen, R.; Cocquyt, V.; Vermaelen, K.; Vandesompele, J.; Bracke, M.; De Wever, O.; Hendrix, A. The impact of disparate isolation methods for extracellular vesicles on downstream RNA profiling. J. Extracell. Vesicles 2014, 3, 24858. [Google Scholar] [CrossRef]
- Karttunen, J.; Heiskanen, M.; Navarro-Ferrandis, V.; Das Gupta, S.; Lipponen, A.; Puhakka, N.; Rilla, K.; Koistinen, A.; Pitkänen, A. Precipitation-based extracellular vesicle isolation from rat plasma co-precipitate vesicle-free microRNAs. J. Extracell. Vesicles 2019, 8, 1555410. [Google Scholar] [CrossRef]
- Wang, T.; Anderson, K.W.; Turko, I.V. Assessment of Extracellular Vesicles Purity Using Proteomic Standards. Anal. Chem. 2017, 89, 11070–11075. [Google Scholar] [CrossRef]
- DDong, L.; Zieren, R.C.; Horie, K.; Kim, C.; Mallick, E.; Jing, Y.; Feng, M.; Kuczler, M.D.; Green, J.; Amend, S.R.; et al. Comprehensive evaluation of methods for small extracellular vesicles separation from human plasma, urine and cell culture medium. J. Extracell. Vesicles 2020, 10, e12044. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, J.D.; Chevillet, J.R.; Kroh, E.M.; Ruf, I.K.; Pritchard, C.C.; Gibson, D.F.; Mitchell, P.S.; Bennett, C.F.; Pogosova-Agadjanyan, E.L.; Stirewalt, D.L.; et al. Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proc. Natl. Acad. Sci. USA 2011, 108, 5003. [Google Scholar] [CrossRef]
- Vickers, K.C.; Palmisano, B.T.; Shoucri, B.M.; Shamburek, R.D.; Remaley, A.T. MicroRNAs are transported in plasma and delivered to recipient cells by high-density lipoproteins. Nat. Cell Biol. 2011, 13, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Zeöld, A.; Sándor, G.O.; Kiss, A.; Soós, A.Á.; Tölgyes, T.; Bursics, A.; Szűcs, Á.; Harsányi, L.; Kittel, Á.; Gézsi, A.; et al. Shared extracellular vesicle miRNA profiles of matched ductal pancreatic adenocarcinoma organoids and blood plasma samples show the power of organoid technology. Cell. Mol. Life Sci. 2021, 78, 3005–3020. [Google Scholar] [CrossRef] [PubMed]
- Just, J.; Yan, Y.; Farup, J.; Sieljacks, P.; Sloth, M.; Venø, M.; Gu, T.; de Paoli, F.V.; Nyengaard, J.R.; Bæk, R.; et al. Blood flow-restricted resistance exercise alters the surface profile, miRNA cargo and functional impact of circulating extracellular vesicles. Sci. Rep. 2020, 10, 5835. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Quek, C.; Li, X.; Bellingham, S.A.; Ellett, L.J.; Shambrook, M.; Zafar, S.; Zerr, I.; Lawson, V.A.; Hill, A.F. Distribution of microRNA profiles in pre-clinical and clinical forms of murine and human prion disease. Commun. Biol. 2021, 4, 411. [Google Scholar] [CrossRef] [PubMed]
- Böing, A.N.; van der Pol, E.; Grootemaat, A.E.; Coumans, F.A.W.; Sturk, A.; Nieuwland, R. Single-step isolation of extracellular vesicles by size-exclusion chromatography. J. Extracell. Vesicles 2014, 3, 23430. [Google Scholar] [CrossRef]
- de Menezes-Neto, A.; Sáez, M.J.F.; Lozano-Ramos, I.; Segui-Barber, J.; Martin-Jaular, L.; Ullate, J.M.E.; Fernandez-Becerra, C.; Borrás, F.E.; Del Portillo, H.A. Size-exclusion chromatography as a stand-alone methodology identifies novel markers in mass spectrometry analyses of plasma-derived vesicles from healthy individuals. J. Extracell. Vesicles 2015, 4, 27378. [Google Scholar] [CrossRef]
- Welton, J.L.; Webber, J.P.; Botos, L.-A.; Jones, M.; Clayton, A. Ready-made chromatography columns for extracellular vesicle isolation from plasma. J. Extracell. Vesicles 2015, 4, 27269. [Google Scholar] [CrossRef]
- Baranyai, T.; Herczeg, K.; Onódi, Z.; Voszka, I.; Módos, K.; Marton, N.; Nagy, G.; Mäger, I.; Wood, M.J.; El Andaloussi, S. Isolation of exosomes from blood plasma: Qualitative and quantitative comparison of ultracentrifugation and size exclusion chromatography methods. PLoS ONE 2015, 10, e0145686. [Google Scholar] [CrossRef]
- Brown, B.A.; Zeng, X.; Todd, A.R.; Barnes, L.F.; Winstone, J.M.A.; Trinidad, J.C.; Novotny, M.V.; Jarrold, M.F.; Clemmer, D.E. Charge Detection Mass Spectrometry Measurements of Exosomes and other Extracellular Particles Enriched from Bovine Milk. Anal. Chem. 2020, 92, 3285–3292. [Google Scholar] [CrossRef]
- Hong, C.-S.; Funk, S.; Muller, L.; Boyiadzis, M.; Whiteside, T.L. Isolation of biologically active and morphologically intact exosomes from plasma of patients with cancer. J. Extracell. Vesicles 2016, 5, 29289. [Google Scholar] [CrossRef]
- Gámez-Valero, A.; Monguió-Tortajada, M.; Carreras-Planella, L.; Franquesa, M.l.; Beyer, K.; Borràs, F.E. Size-Exclusion Chromatography-based isolation minimally alters Extracellular Vesicles’ characteristics compared to precipitating agents. Sci. Rep. 2016, 6, 33641. [Google Scholar] [CrossRef] [PubMed]
- Simonsen, J.B. What are we looking at? Extracellular vesicles, lipoproteins, or both? Circ. Res. 2017, 121, 920–922. [Google Scholar] [CrossRef] [PubMed]
- Sluijter, J.P.G.; Davidson, S.M.; Boulanger, C.M.; Buzas, E.I.; De Kleijn, D.P.V.; Engel, F.B.; Giricz, Z.; Hausenloy, D.J.; Kishore, R.; Lecour, S. Extracellular vesicles in diagnostics and therapy of the ischaemic heart: Position Paper from the Working Group on Cellular Biology of the Heart of the European Society of Cardiology. Cardiovasc. Res. 2018, 114, 19–34. [Google Scholar] [CrossRef]
- Ridger, V.C.; Boulanger, C.M.; Angelillo-Scherrer, A.; Badimon, L.; Blanc-Brude, O.; Bochaton-Piallat, M.-L.; Boilard, E.; Buzas, E.I.; Caporali, A.; Dignat-George, F. Microvesicles in vascular homeostasis and diseases. Position paper of the European Society of Cardiology (ESC) working group on atherosclerosis and vascular biology. Thromb. Haemost. 2017, 117, 1296–1316. [Google Scholar] [PubMed]
- Karimi, N.; Cvjetkovic, A.; Jang, S.C.; Crescitelli, R.; Hosseinpour Feizi, M.A.; Nieuwland, R.; Lötvall, J.; Lässer, C. Detailed analysis of the plasma extracellular vesicle proteome after separation from lipoproteins. Cell. Mol. Life Sci. 2018, 75, 2873–2886. [Google Scholar] [CrossRef]
- Buschmann, D.; Kirchner, B.; Hermann, S.; Märte, M.; Wurmser, C.; Brandes, F.; Kotschote, S.; Bonin, M.; Steinlein, O.K.; Pfaffl, M.W.; et al. Evaluation of serum extracellular vesicle isolation methods for profiling miRNAs by next-generation sequencing. J. Extracell. Vesicles 2018, 7, 1481321. [Google Scholar] [CrossRef]
- Takov, K.; Yellon, D.M.; Davidson, S.M. Comparison of small extracellular vesicles isolated from plasma by ultracentrifugation or size-exclusion chromatography: Yield, purity and functional potential. J. Extracell. Vesicles 2019, 8, 1560809. [Google Scholar] [CrossRef]
- Ronquist, G. Prostasomes: Their Characterisation: Implications for Human Reproduction. In The Male Role in Pregnancy Loss and Embryo Implantation Failure; Bronson, R., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 191–209. [Google Scholar]
- Ronquist, G. Prostasomes are mediators of intercellular communication: From basic research to clinical implications. J. Intern. Med. 2012, 271, 400–413. [Google Scholar] [CrossRef]
- Ronquist, G.; Brody, I. The prostasome: Its secretion and function in man. Biochim. Biophys. Acta 1985, 822, 203–218. [Google Scholar] [CrossRef]
- Peeker, R.; Abramsson, L.; Marklund, S.L. Superoxide dismutase isoenzymes in human seminal plasma and spermatozoa. Mol. Hum. Reprod. 1997, 3, 1061–1066. [Google Scholar] [CrossRef]
- Saez, F.; Motta, C.; Boucher, D.; Grizard, G. Antioxidant capacity of prostasomes in human semen. Mol. Hum. Reprod. 1998, 4, 667–672. [Google Scholar] [CrossRef]
- Jones, J.L.; Saraswati, S.; Block, A.S.; Lichti, C.F.; Mahadevan, M.; Diekman, A.B. Galectin-3 is associated with prostasomes in human semen. Glycoconj. J. 2010, 27, 227–236. [Google Scholar] [CrossRef]
- Aalberts, M.; van Dissel-Emiliani, F.M.F.; van Adrichem, N.P.H.; van Wijnen, M.; Wauben, M.H.M.; Stout, T.A.E.; Stoorvogel, W. Identification of Distinct Populations of Prostasomes That Differentially Express Prostate Stem Cell Antigen, Annexin A1, and GLIPR2 in Humans1. Biol. Reprod. 2012, 86, 82. [Google Scholar] [CrossRef] [PubMed]
- Brouwers, J.F.; Aalberts, M.; Jansen, J.W.A.; van Niel, G.; Wauben, M.H.; Stout, T.A.E.; Helms, J.B.; Stoorvogel, W. Distinct lipid compositions of two types of human prostasomes. Proteomics 2013, 13, 1660–1666. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Y.; Kaddour, H.; Kopcho, S.; Panzner, T.D.; Shouman, N.; Kim, E.-Y.; Martinson, J.; McKay, H.; Martinez-Maza, O.; Margolick, J.B.; et al. Human Immunodeficiency Virus (HIV) Infection and Use of Illicit Substances Promote Secretion of Semen Exosomes that Enhance Monocyte Adhesion and Induce Actin Reorganization and Chemotactic Migration. Cells 2019, 8, 1027. [Google Scholar] [CrossRef]
- Kaddour, H.; Lyu, Y.; Shouman, N.; Mohan, M.; Okeoma, C.M. Development of Novel High-Resolution Size-Guided Turbidimetry-Enabled Particle Purification Liquid Chromatography (PPLC): Extracellular Vesicles and Membraneless Condensates in Focus. Int. J. Mol. Sci. 2020, 21, 5361. [Google Scholar] [CrossRef] [PubMed]
- Welch, J.L.; Kaddour, H.; Winchester, L.; Fletcher, C.V.; Stapleton, J.T.; Okeoma, C.M. Semen Extracellular Vesicles From HIV-1–Infected Individuals Inhibit HIV-1 Replication In Vitro, and Extracellular Vesicles Carry Antiretroviral Drugs In Vivo. JAIDS J. Acquir. Immune Defic. Syndr. 2020, 83, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Kaddour, H.; Panzner, T.D.; Welch, J.L.; Shouman, N.; Mohan, M.; Stapleton, J.T.; Okeoma, C.M. Electrostatic Surface Properties of Blood and Semen Extracellular Vesicles: Implications of Sialylation and HIV-Induced Changes on EV Internalization. Viruses 2020, 12, 1117. [Google Scholar] [CrossRef]
- Welch, J.L.; Stapleton, J.T.; Okeoma, C.M. Vehicles of intercellular communication: Exosomes and HIV-1. J. Gen. Virol. 2019, 100, 350–366. [Google Scholar] [CrossRef]
- Welch, J.L.; Kaddour, H.; Schlievert, P.M.; Stapleton, J.T.; Okeoma, C.M. Semen exosomes promote transcriptional silencing of HIV-1 by disrupting NF-κB/Sp1/Tat circuitry. J. Virol. 2018, 92, e00731-18. [Google Scholar] [CrossRef] [PubMed]
- Sun, I.O.; Lerman, L.O. Urinary Extracellular Vesicles as Biomarkers of Kidney Disease: From Diagnostics to Therapeutics. Diagnostics 2020, 10, 311. [Google Scholar] [CrossRef] [PubMed]
- Erdbrügger, U.; Le, T.H. Extracellular vesicles in renal diseases: More than novel biomarkers? J. Am. Soc. Nephrol. 2016, 27, 12–26. [Google Scholar] [CrossRef] [PubMed]
- Lindblom, H.; Söderberg, L.; Cooper, E.H.; Turner, R. Urinary protein isolation by high-performance ion exchange chromatography. J. Chromatogr. A 1983, 266, 187–196. [Google Scholar] [CrossRef]
- Rood, I.M.; Deegens, J.K.; Merchant, M.L.; Tamboer, W.P.; Wilkey, D.W.; Wetzels, J.F.; Klein, J.B. Comparison of three methods for isolation of urinary microvesicles to identify biomarkers of nephrotic syndrome. Kidney Int. 2010, 78, 810–816. [Google Scholar] [CrossRef]
- Lozano-Ramos, I.; Bancu, I.; Oliveira-Tercero, A.; Armengol, M.P.; Menezes-Neto, A.; Portillo, H.A.D.; Lauzurica-Valdemoros, R.; Borràs, F.E. Size-exclusion chromatography-based enrichment of extracellular vesicles from urine samples. J. Extracell. Vesicles 2015, 4, 27369. [Google Scholar] [CrossRef]
- Lozano-Ramos, S.I.; Bancu, I.; Carreras-Planella, L.; Monguió-Tortajada, M.; Cañas, L.; Juega, J.; Bonet, J.; Armengol, M.P.; Lauzurica, R.; Borràs, F.E. Molecular profile of urine extracellular vesicles from normo-functional kidneys reveal minimal differences between living and deceased donors. BMC Nephrol. 2018, 19, 189. [Google Scholar] [CrossRef]
- Ogawa, Y.; Kanai-Azuma, M.; Akimoto, Y.; Kawakami, H.; Yanoshita, R. Exosome-like vesicles with dipeptidyl peptidase IV in human saliva. Biol. Pharm. Bull. 2008, 31, 1059–1062. [Google Scholar] [CrossRef]
- Ogawa, Y.; Miura, Y.; Harazono, A.; Kanai-Azuma, M.; Akimoto, Y.; Kawakami, H.; Yamaguchi, T.; Toda, T.; Endo, T.; Tsubuki, M. Proteomic analysis of two types of exosomes in human whole saliva. Biol. Pharm. Bull. 2011, 34, 13–23. [Google Scholar] [CrossRef]
- Aqrawi, L.A.; Galtung, H.K.; Vestad, B.; Øvstebø, R.; Thiede, B.; Rusthen, S.; Young, A.; Guerreiro, E.M.; Utheim, T.P.; Chen, X. Identification of potential saliva and tear biomarkers in primary Sjögren’s syndrome, utilising the extraction of extracellular vesicles and proteomics analysis. Arthritis Res. Ther. 2017, 19, 14. [Google Scholar] [CrossRef]
- Blans, K.; Hansen, M.S.; Sørensen, L.V.; Hvam, M.L.; Howard, K.A.; Möller, A.; Wiking, L.; Larsen, L.B.; Rasmussen, J.T. Pellet-free isolation of human and bovine milk extracellular vesicles by size-exclusion chromatography. J. Extracell. Vesicles 2017, 6, 1294340. [Google Scholar] [CrossRef] [PubMed]
- Tietje, A.; Maron, K.N.; Wei, Y.; Feliciano, D.M. Cerebrospinal Fluid Extracellular Vesicles Undergo Age Dependent Declines and Contain Known and Novel Non-coding RNAs. PLoS ONE 2014, 9, e113116. [Google Scholar] [CrossRef]
- Carreras-Planella, L.; Soler-Majoral, J.; Rubio-Esteve, C.; Lozano-Ramos, S.I.; Franquesa, M.; Bonet, J.; Troya-Saborido, M.I.; Borràs, F.E. Characterization and proteomic profile of extracellular vesicles from peritoneal dialysis efflux. PLoS ONE 2017, 12, e0176987. [Google Scholar] [CrossRef] [PubMed]
- Foers, A.D.; Chatfield, S.; Dagley, L.F.; Scicluna, B.J.; Webb, A.I.; Cheng, L.; Hill, A.F.; Wicks, I.P.; Pang, K.C. Enrichment of extracellular vesicles from human synovial fluid using size exclusion chromatography. J. Extracell. Vesicles 2018, 7, 1490145. [Google Scholar] [CrossRef] [PubMed]
- Sidhom, K.; Obi, P.O.; Saleem, A. A Review of Exosomal Isolation Methods: Is Size Exclusion Chromatography the Best Option? Int. J. Mol. Sci. 2020, 21, 6466. [Google Scholar] [CrossRef]
- Van Deun, J.; Jo, A.; Li, H.; Lin, H.Y.; Weissleder, R.; Im, H.; Lee, H. Integrated Dual-Mode Chromatography to Enrich Extracellular Vesicles from Plasma. Adv. Biosyst. 2020, 1900310. [Google Scholar] [CrossRef]
- Audhya, T.; Reddy, J.; Zaneveld, L.J.D. Purification and Partial Chemical Characterization of a Glycoprotein with Antifertility Activity from Human Seminal Plasma1. Biol. Reprod. 1987, 36, 511–521. [Google Scholar] [CrossRef]
- Winzor, D.J. Analytical exclusion chromatography. J. Biochem. Biophys. Methods 2003, 56, 15–52. [Google Scholar] [CrossRef]
- Elsayed, M.M.; Cevc, G. Turbidity spectroscopy for characterization of submicroscopic drug carriers, such as nanoparticles and lipid vesicles: Size determination. Pharm. Res. 2011, 28, 2204–2222. [Google Scholar] [CrossRef]
- Wang, A.; Miller, C.C.; Szostak, J.W. Core-Shell Modeling of Light Scattering by Vesicles: Effect of Size, Contents, and Lamellarity. Biophys. J. 2019, 116, 659–669. [Google Scholar] [CrossRef]
- Wang, A.; Garmann, R.F.; Manoharan, V.N. Tracking E. coli runs and tumbles with scattering solutions and digital holographic microscopy. Opt. Express 2016, 24, 23719–23725. [Google Scholar] [CrossRef]
- Varga, Z.; Yuana, Y.; Grootemaat, A.E.; Van der Pol, E.; Gollwitzer, C.; Krumrey, M.; Nieuwland, R. Towards traceable size determination of extracellular vesicles. J. Extracell. Vesicles 2014, 3, 23298. [Google Scholar] [CrossRef] [PubMed]
- Wyss, R.; Grasso, L.; Wolf, C.; Grosse, W.; Demurtas, D.; Vogel, H. Molecular and dimensional profiling of highly purified extracellular vesicles by fluorescence fluctuation spectroscopy. Anal. Chem. 2014, 86, 7229–7233. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Gong, M.; Hu, Y.; Liu, H.; Zhang, W.; Zhang, M.; Hu, X.; Aubert, D.; Zhu, S.; Wu, L.; et al. Quality and efficiency assessment of six extracellular vesicle isolation methods by nano-flow cytometry. J. Extracell. Vesicles 2020, 9, 1697028. [Google Scholar] [CrossRef] [PubMed]
- Sultana, H.; Sultana, H.; Neelakanta, G.; Neelakanta, G.; Sultana, H.; Sultana, H.; Neelakanta, G.; Neelakanta, G. Arthropod exosomes as bubbles with message(s) to transmit vector-borne diseases. Curr. Opin. Insect Sci. 2020, 40, 39–47. [Google Scholar] [CrossRef] [PubMed]



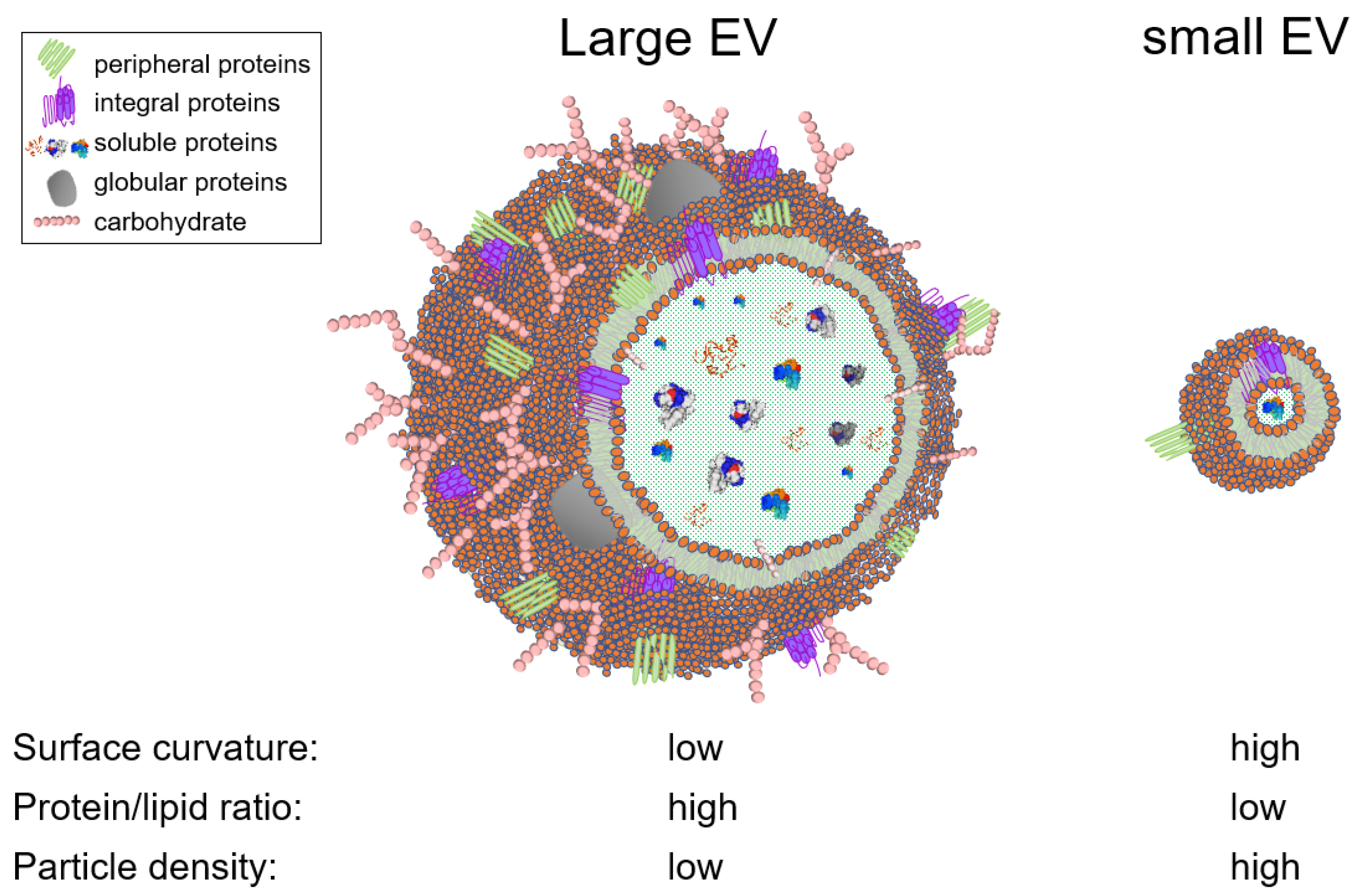{kind=link}
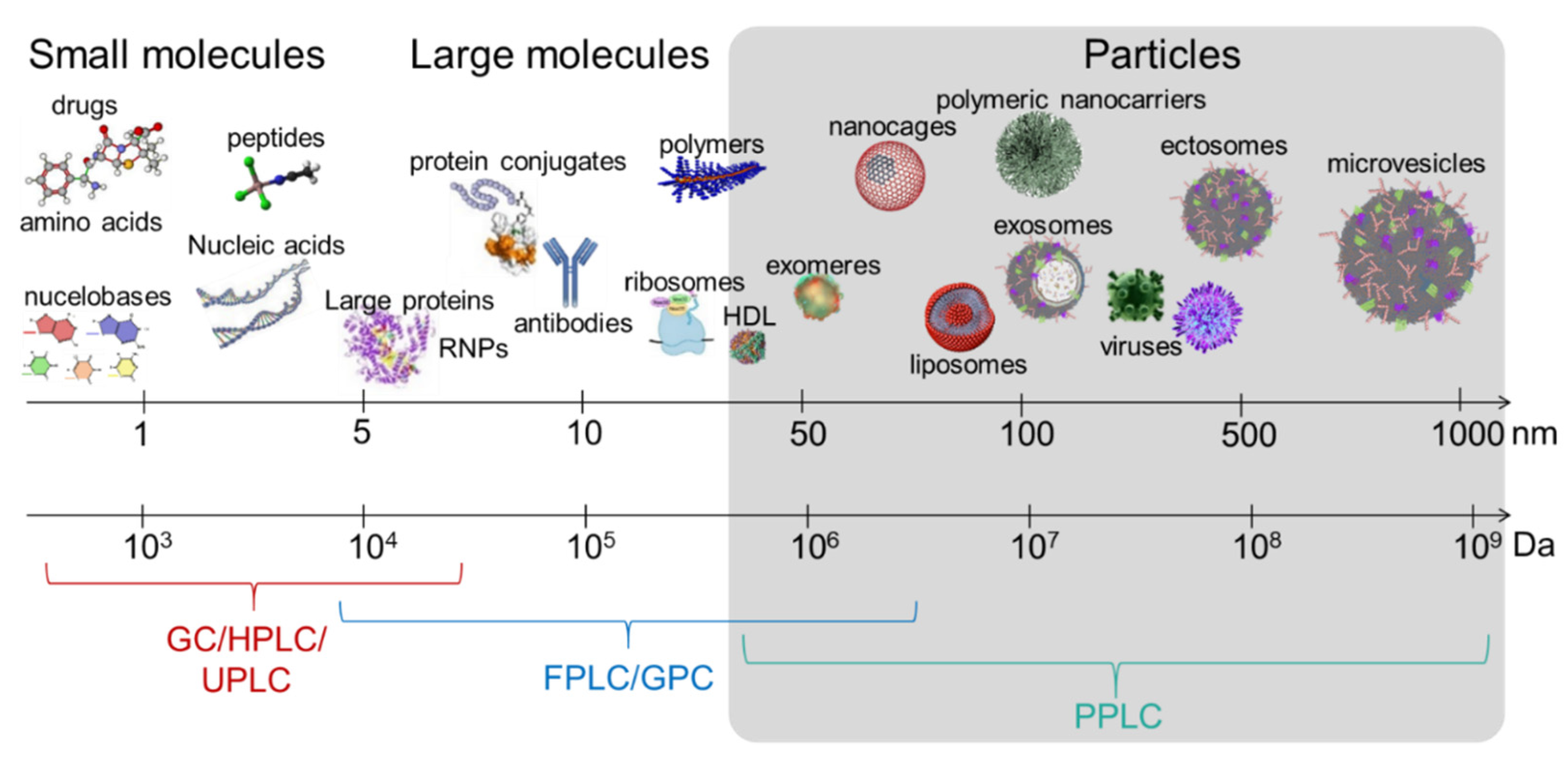{kind=link}
| Company | Trade Name | Matrix | Product Number | Fractionation Range for Globular Protein (Da) |
|---|---|---|---|---|
| Cytvia ¥ | Sephadex | Crosslinked dextran | G10 | <700 |
| G15 | <1500 | |||
| G25 | 1000–5000 | |||
| G50 | 1000–30,000 | |||
| G75 | 3000–80,000 | |||
| G100 | 4000–150,000 | |||
| G200 * | 1000–200,000 | |||
| Sephacryl | Crosslinked allyl dextran and N,N’-methylene bisacrylamide | S-100 HR | 1000–100,000 | |
| S-200 HR | 5000–250,000 | |||
| S-300 HR | 10,000–1,500,000 | |||
| S-400 HR | 20,000–8,000,000 | |||
| S-500 HR | 40,000–20,000,000 | |||
| Sepharose | Crosslinked agarose | CL-2B | 70,000–40,000,000 | |
| CL-4B | 70,000–20,000,000 | |||
| CL-6B | 10,000–4,000,000 | |||
| Superose | 6 | 5000–5,000,000 | ||
| 12 | 1000–300,000 | |||
| Superdex | Composite dextran/crosslinked agarose | 30 | <10,000 | |
| 75 | 3000–70,000 | |||
| 200 | 10,000–600,000 | |||
| BioRad | Bio-Gel P | Crosslinked polyacrylamide and N,N’-methylene bisacrylamide | P-2 | 100–1800 |
| P-4 | 800–4000 | |||
| P-6 | 1000–6000 | |||
| P-10 | 1500–20,000 | |||
| P-30 | 2500–40,000 | |||
| P-60 | 3000–60,000 | |||
| P-100 | 50,000–100,000 | |||
| Bio-Gel A | Crosslinked agarose | A-0.5 m * | <10,000–500,000 | |
| A-1.5 m * | <10,000–1,500,000 | |||
| A-5 m * | 10,000–5,000,000 | |||
| A-15 m | 40,000–15,000,000 | |||
| A-50 m * | 100,000–50,000,000 | |||
| A-150 m * | 100,000–150,000,000 | |||
| Tosoh | ToyoPEARL | polymethacrylate | HW-40 | >10,000 £ |
| HW-50 | >80,000 £ | |||
| HW-55 | >700,000 £ | |||
| HW-65 | >5,000,000 £ | |||
| HW-75 | >50,000,000 £ |
| Study | Tissue/Cell Line/Biofluid | Column Size | Beads | Commercial/Lab Made | Fractions | System Used | Separation Profile Shown |
|---|---|---|---|---|---|---|---|
| Lee et al. [65] | Mouse bone marrow-derived MSCs, MSCs from human umbilical cord Wharton’s jelly, and human dermal fibroblasts | 1.6 × 60 cm | Hiprep Sephacryl S-400 HR | prepacked (GE healthcare) | 140 of 1 mL each | ÄKTA purifier liquid chromatography system | yes |
| Hajj et al. [66] | Astrocytes | 50 cm | Superose 12 prep grade 3000-kDa exclusion | prepacked (GE healthcare) | 25 of 3.5 mL each | NR | no |
| Redzic et al. [67] | cell lines: MCF-7, MDA-MB-231, L3.6pL and Hek293Fpl, U937 | NR | preparatory Superose 6 | prepacked (GE healthcare) | NR | NR | yes |
| Nordin et al. [68] | Cell lines: NSC-34, N2a, B16F10, HEK293T, iPSCs | NR | Sephacryl S-400 column | prepacked (GE healthcare) | 2 mL | ÄKTA prime (GE Healthcare) | yes |
| Willms et al. [69] | Cell lines: B16F10, A431, N2a, H5V, hTERT | 1.6 × 60 cm | HiPrep Sephacryl S-400 HR | prepacked (GE healthcare) | 2 mL | ÄKTA prime (GE Healthcare) | yes |
| Mol et al. [70] | cell lines: CPCs and HMECs | NR | S400 highprep column | NR | NR | ÄKTAStart (GE Healthcare) | no |
| Benedikter et al. [71] | cell line: BEAS-2B | 10 mL | Sepharose CL-4B | in-house | 0.5 mL | gravity | no |
| Corso et al. [72] | N2a and myoblast C2C12 | HiScreen Capto Core 700 column | GE Healthcare Life Sciences | NR | ÄKTA prime plus or ÄKTA Pure 25 | yes | |
| Taylor et al. [74] | human blood serum | 1.5 × 45 cm | Bio-Gel A50 m | in-house | NR | NR | no |
| Kim et al. [75] | human blood serum, FasL-transduced PCI-13 cell lines and of normal human fibroblasts | 1 × 35 cm | Sepharose 2B | in-house | 1 mL | NR | no |
| Taylor et al. [76] | ascites specimens from women diagnosed with stage III serous adenocarcinoma of the ovary | 2.5 × 16 cm | 2% agarose (Agarose Bead Technologies, Tampa, FL) | 2 mL | |||
| Shu et al. [77] | 2183-Her4 and 888-mel melanoma cell lines | N/A | N/A | Exo-spin columns from cell Guidance systems | N/A | N/A | N/A |
| Wang et al. [81] | human blood serum | 1 × 30 cm | Superose 6 Increase 10/300 GL column | prepacked (GE healthcare) | 0.5 mL | AKTA FPLC | yes |
| Arroyo et al. [83] | Human blood serum and plasma | 0.9 × 30 cm | Sephacryl S-500 | in-house | 25 of 1 mL | NR | yes |
| Vickers et al. [84] | Human blood plasma | 1 × 30 cm or 3 × 10 cm | Superose 6 HR 10/30 or Sephacryl S-200 | prepacked (GE healthcare) or in-house | NR | NR | yes |
| Böing et al. [88] | Human plasma | 1.6 × 6.2 cm | Sepharose CL-2B | in-house | 26 of 0.5 mL | gravity | yes |
| De Menezes-Neto et al. [89]. | Human plasma | 10 mL | Sepharose CL-2B | in-house | 30 of 0.5 mL | gravity | yes |
| Welton et al. [90] | Du145 cell line and human plasma | NR | Exo-Spin™ Midi Columns | Prepacked (CellGS) | 30 of 0.5 mL | gravity | yes |
| Baranyai et al. [91] | Human blood plasma | 10 mL | Sepharose 2B, CL-4B, or Sephacryl S-400 | in-house | 10 of 1 mL | gravity | no |
| Rat plasma | 120 mL | Sephacryl S-400 (GE Healthcare) | NR | 2 mL–5 mL | ÄKTA pure 25 L | yes | |
| Hong et al. [93] | Human plasma | 1.5 ×12 cm (10 mL) | Sepharose 2B | in-house | 5 of 1 mL | gravity | No |
| Gámez-Valero et al. [94] | Human blood plasma | 12 mL | Sepharose CL-2B | in-house | 20 of 0.5 mL | gravity | yes |
| Jones et al. [106] | Human seminal plasma | 35 mL | Sephacryl S300 | NR | 28 NR volume | NR | No |
| Aalberts et al. [107] | Sucrose density gradient isolated prostasomes | 70 × 2.6 cm | Sephacryl S-1000 | prepacked (GE healthcare) | 2 mL | NR | No |
| Brouwers et al. [108] | Human seminal plasma | 70 × 2.6 cm | Sephacryl S-1000 | prepacked (GE healthcare) | 2 mL | NR | No |
| Lyu et al. [109] | Human seminal plasma | 22 × 1 cm | Sephadex G-50 fine | in-house | 60 of 0.2 mL | gravity | yes |
| Kaddour et al. [110] | Human seminal plasma | 100 × 1 cm | Gradient of 6 Sephadex beads | in-house | ~800 of 0.2 mL | PPLC | yes |
| Rood et al. [118] | Human urine | 30 × 0.78 cm | BioSep-SEC-S4000 | prepacked (Phenomenex) | 1 mL | UltimateTM 3000 HPLC | yes |
| Lozano-Ramos et al. [119] | Human urine | 10 mL | Sepharose-CL2B | in-house | 20 of 0.5 mL | gravity | yes |
| Lozano-Ramos et al. [120] | Human urine | 10 mL | Sepharose-CL2B | in-house | 20 of 0.5 mL | gravity | yes |
| Ogawa et al. [121] | Human saliva | 1.5 × 50 cm | Sepharose-CL4B | in-house | 80 NR volume | NR | yes |
| Ogawa et al. [122] | Human saliva | 1.5 × 50 cm | Sephacryl S-500 | in-house | 100 NR volume | NR | yes |
| Aqrawi et al. [123] | Human saliva and tears | NR | qEV | prepacked (Izon) | 16 of 0.5 mL | gravity | no |
| Blans et al. [124] | Human and bovine milk | 2.5 × 88 cm (432 mL) | Sephacryl S-500 | NR | 193 of 4.4 mL | FPLC | yes |
| 238 mL | Sephacryl S-500 | NR | 119 of 2.65 mL | FPLC | yes | ||
| Tietje et al. [125] | Human CSF | N/A | Sephacryl S-400 HR | prepacked spin columns (GE healthcare) | N/A | N/A | N/A |
| Carreras-Planella et al. [126] | peritoneal dialysis efflux | 12 mL | Sepharose-CL2B | in-house | 20 of 0.5 mL | gravity | yes |
| Foers et al. [127] | synovial fluid | 2.6 × 60 cm (120 mL) | HiPrep 26/60 Sephacryl S-500 HR | prepacked (GE Healthcare) | NR | NR | yes |
| Current EV Purification Challenges | PPLC Solutions | |
|---|---|---|
| Subpopulation separation requires antibodies whose elution compromises EV integrity |  | Dye-free separation of intact EV subpopulations |
| Low reproducibility between different laboratories |  | A fully automated fractionation and characterization instrument to reduce handling error |
| Sample variability (different cell lines, different body fluids, different donors, disease states, etc.) |  | A separation method that is tailored to and valid for each sample type |
| Characterization performed post-purification introducing handling errors (storage time, temperature, freeze-thaw cycles, etc.) |  | in situ monitoring for real-time complete biophysical characterization |
| small number of isolated fractions in density gradient purification decreases the power of subpopulation separation |  | Up to a few thousands of fractions for near single-vesicle resolution |
| Techniques are either laborious, expensive, and/or require special skills |  | User-friendly, cost-effective system that requires minimum handling effort |
| Lack of significant room for improvement and scalability in available techniques (density gradient, AF4, immunocapture, etc.) |  | Ample room for improvement by optional addition of extra detectors (light scattering and/or fluorescence) and extra column sizes/chemistries. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaddour, H.; Tranquille, M.; Okeoma, C.M. The Past, the Present, and the Future of the Size Exclusion Chromatography in Extracellular Vesicles Separation. Viruses 2021, 13, 2272. https://doi.org/10.3390/v13112272
Kaddour H, Tranquille M, Okeoma CM. The Past, the Present, and the Future of the Size Exclusion Chromatography in Extracellular Vesicles Separation. Viruses. 2021; 13(11):2272. https://doi.org/10.3390/v13112272
Chicago/Turabian StyleKaddour, Hussein, Malik Tranquille, and Chioma M. Okeoma. 2021. "The Past, the Present, and the Future of the Size Exclusion Chromatography in Extracellular Vesicles Separation" Viruses 13, no. 11: 2272. https://doi.org/10.3390/v13112272
APA StyleKaddour, H., Tranquille, M., & Okeoma, C. M. (2021). The Past, the Present, and the Future of the Size Exclusion Chromatography in Extracellular Vesicles Separation. Viruses, 13(11), 2272. https://doi.org/10.3390/v13112272