
Monoclonal Antibodies to S and N SARS-CoV-2 Proteins as Probes to Assess Structural and Antigenic Properties of Coronaviruses

 , , , ,
, , , ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmids, Protein Expression and Purification
2.2. Immunization of Mice and Rabbits—Preparation of Sera and Mabs
2.3. Enzyme-Linked Immunosorbent Assays (ELISA)
2.4. Western Blotting
2.5. Immunofluorescence Microscopy and Imaging
2.6. SARS-CoV-2 Microneutralization Assay (VN)
3. Results
3.1. Generation of S-RBD Antibodies with Diverse Binding and Neutralization Properties
3.2. N Protein Antibodies Bind throughout the N Protein and Vary in Their Coronavirus Specificities
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Coronaviridae Study Group of the International Committee on Taxonomy of Viruses. The species Severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef] [Green Version]
- Krause, P.R.; Fleming, T.R.; Longini, I.M.; Peto, R.; Briand, S.; Heymann, D.L.; Beral, V.; Snape, M.D.; Rees, H.; Ropero, A.M.; et al. SARS-CoV-2 Variants and Vaccines. N. Engl. J. Med. 2021, 385, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Lurie, N.; Saville, M.; Hatchett, R.; Halton, J. Developing Covid-19 Vaccines at Pandemic Speed. N. Engl. J. Med. 2020, 382, 1969–1973. [Google Scholar] [CrossRef] [PubMed]
- Baum, A.; Fulton, B.O.; Wloga, E.; Copin, R.; Pascal, K.E.; Russo, V.; Giordano, S.; Lanza, K.; Negron, N.; Ni, M.; et al. Antibody cocktail to SARS-CoV-2 spike protein prevents rapid mutational escape seen with individual antibodies. Science 2020, 369, 1014–1018. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.; Baum, A.; Pascal, K.E.; Russo, V.; Giordano, S.; Wloga, E.; Fulton, B.O.; Yan, Y.; Koon, K.; Patel, K.; et al. Studies in humanized mice and convalescent humans yield a SARS-CoV-2 antibody cocktail. Science 2020, 369, 1010–1014. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.E.; Jang, G.M.; Bouhaddou, M.; Xu, J.; Obernier, K.; White, K.M.; O’Meara, M.J.; Rezelj, V.V.; Guo, J.Z.; Swaney, D.L.; et al. A SARS-CoV-2 protein interaction map reveals targets for drug repurposing. Nature 2020, 583, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Yan, R.; Zhang, Y.; Li, Y.; Xia, L.; Guo, Y.; Zhou, Q. Structural basis for the recognition of SARS-CoV-2 by full-length human ACE2. Science 2020, 367, 1444–1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walls, A.C.; Park, Y.J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292.e6. [Google Scholar] [CrossRef] [PubMed]
- Burbelo, P.D.; Riedo, F.X.; Morishima, C.; Rawlings, S.; Smith, D.; Das, S.; Strich, J.R.; Chertow, D.S.; Davey, R.T.; Cohen, J.I. Sensitivity in Detection of Antibodies to Nucleocapsid and Spike Proteins of Severe Acute Respiratory Syndrome Coronavirus 2 in Patients With Coronavirus Disease 2019. J. Infect. Dis. 2020, 222, 206–213. [Google Scholar] [CrossRef]
- Gibson, D.G.; Young, L.; Chuang, R.Y.; Venter, J.C.; Hutchison, C.A.; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef]
- Stadlbauer, D.; Amanat, F.; Chromikova, V.; Jiang, K.; Strohmeier, S.; Arunkumar, G.A.; Tan, J.; Bhavsar, D.; Capuano, C.; Kirkpatrick, E.; et al. SARS-CoV-2 Seroconversion in Humans: A Detailed Protocol for a Serological Assay, Antigen Production, and Test Setup. Curr. Protoc. Microbiol. 2020, 57, e100. [Google Scholar] [CrossRef] [Green Version]
- Gontu, A.; Srinivasan, S.; Salazar, E.; Nair, M.S.; Nissly, R.H.; Greenawalt, D.; Bird, I.M.; Herzog, C.M.; Ferrari, M.J.; Poojary, I.; et al. Limited window for donation of convalescent plasma with high live-virus neutralizing antibody titers for COVID-19 immunotherapy. Commun. Biol. 2021, 4, 267. [Google Scholar] [CrossRef]
- Studier, F.W. Protein production by auto-induction in high density shaking cultures. Protein Expr. Purif. 2005, 41, 207–234. [Google Scholar] [CrossRef]
- Walker, J.M. The Proteomics Protocols Handbook; Springer: Berlin/Heidelberg, Germany, 2005. [Google Scholar]
- Brendle, S.A.; Culp, T.D.; Broutian, T.R.; Christensen, N.D. Binding and neutralization characteristics of a panel of monoclonal antibodies to human papillomavirus 58. J. Gen. Virol. 2010, 91, 1834–1839. [Google Scholar] [CrossRef] [PubMed]
- Spieker-Polet, H.; Sethupathi, P.; Yam, P.C.; Knight, K.L. Rabbit monoclonal antibodies: Generating a fusion partner to produce rabbit-rabbit hybridomas. Proc. Natl. Acad. Sci. USA 1995, 92, 9348–9352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, N.D.; Kreider, J.W.; Cladel, N.M.; Patrick, S.D.; Welsh, P.A. Monoclonal antibody-mediated neutralization of infectious human papillomavirus type 11. J. Virol. 1990, 64, 5678–5681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, N.D.; Dillner, J.; Eklund, C.; Carter, J.J.; Wipf, G.C.; Reed, C.A.; Cladel, N.M.; Galloway, D.A. Surface conformational and linear epitopes on HPV-16 and HPV-18 L1 virus-like particles as defined by monoclonal antibodies. Virology 1996, 223, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Salazar, E.; Kuchipudi, S.V.; Christensen, P.A.; Eagar, T.; Yi, X.; Zhao, P.; Jin, Z.; Long, S.W.; Olsen, R.J.; Chen, J.; et al. Convalescent plasma anti-SARS-CoV-2 spike protein ectodomain and receptor-binding domain IgG correlate with virus neutralization. J. Clin. Investig. 2020, 130, 6728–6738. [Google Scholar] [CrossRef]
- Christensen, N.D.; Kirnbauer, R.; Schiller, J.T.; Ghim, S.J.; Schlegel, R.; Jenson, A.B.; Kreider, J.W. Human papillomavirus types 6 and 11 have antigenically distinct strongly immunogenic conformationally dependent neutralizing epitopes. Virology 1994, 205, 329–335. [Google Scholar] [CrossRef]
- Gupta, R.; Charron, J.; Stenger, C.L.; Painter, J.; Steward, H.; Cook, T.W.; Faber, W.; Frisch, A.; Lind, E.; Bauss, J.; et al. SARS-CoV-2 (COVID-19) structural and evolutionary dynamicome: Insights into functional evolution and human genomics. J. Biol. Chem. 2020, 295, 11742–11753. [Google Scholar] [CrossRef] [PubMed]
- Opalka, D.; Lachman, C.E.; MacMullen, S.A.; Jansen, K.U.; Smith, J.F.; Chirmule, N.; Esser, M.T. Simultaneous quantitation of antibodies to neutralizing epitopes on virus-like particles for human papillomavirus types 6, 11, 16, and 18 by a multiplexed luminex assay. Clin. Diagn. Lab. Immunol. 2003, 10, 108–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cubuk, J.; Alston, J.J.; Incicco, J.J.; Singh, S.; Stuchell-Brereton, M.D.; Ward, M.D.; Zimmerman, M.I.; Vithani, N.; Griffith, D.; Wagoner, J.A.; et al. The SARS-CoV-2 nucleocapsid protein is dynamic, disordered, and phase separates with RNA. Nat. Commun. 2021, 12, 1936. [Google Scholar] [CrossRef] [PubMed]
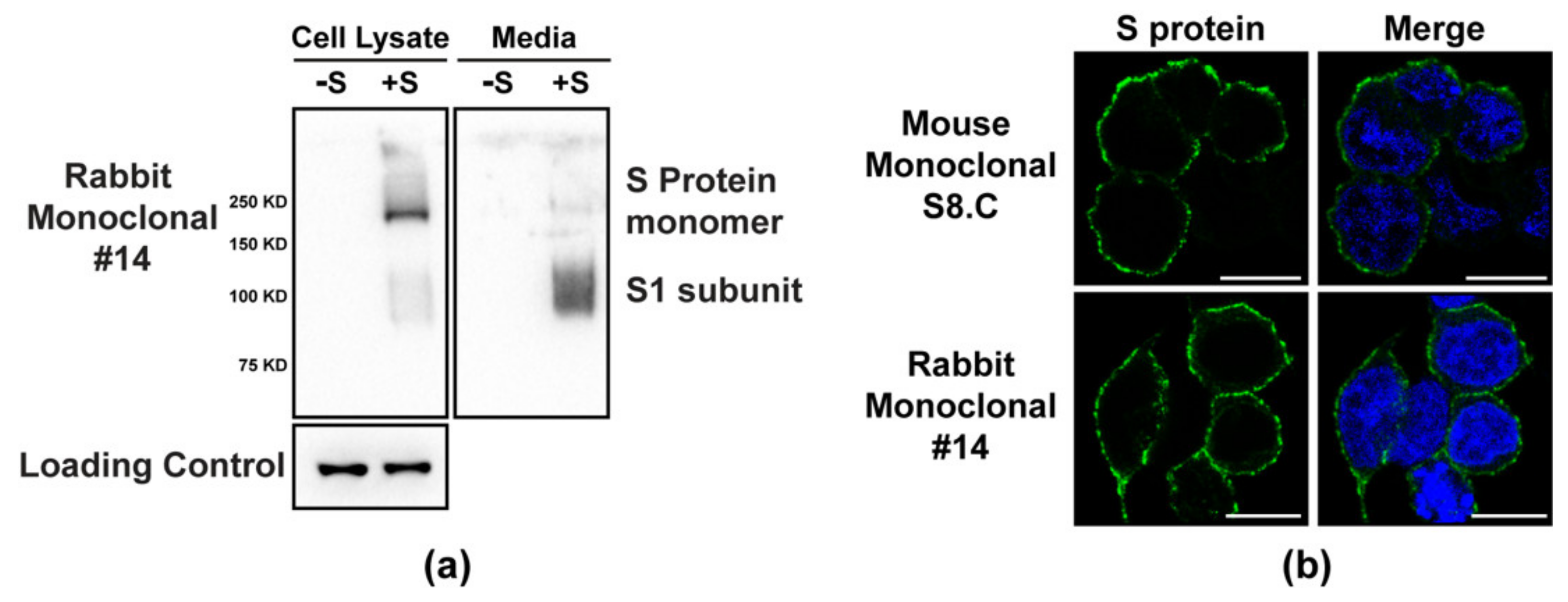



{kind=link}
{kind=link}
{kind=link}
| ELISA a Signals to Various Antigens and Competitive ELISA | ||||||||
|---|---|---|---|---|---|---|---|---|
| Mab (Mouse) | CoV-2 S-RBD (intact) | CoV-2 S-RBD (D and R) | Comp with S3.E b | Comp with #24 c | Comp with #14 d | Comp with #77 e | N (Native) | Isotype |
| S1.D | + | − | − | − | − | − | − | IgG1 |
| S2.D | + | − | + | − | + | + | − | IgG1 |
| S3.E | + | − | N/A | − | + | + | + | IgM |
| S5.F | + | − | + | − | − | + | − | IgG1 |
| S7.A | + | − | + | − | + | + | − | IgG1 |
| S8.C | + | − | + | − | + | + | + | IgG2b |
| S11.A | + | + | − | + | − | − | − | IgG2b |
| S15.B | + | − | + | − | + | + | − | IgG1 |
| S16.C | + | − | + | − | + | + | − | IgG1 |
| S18.C | + | − | − | − | − | − | − | IgG1 |
| S19.K | + | − | − | − | − | − | − | IgG1 |
| S20.I | + | − | + | − | + | + | − | IgG1 |
| S21.D | + | + | − | + | − | − | − | IgG1 |
| S22.D | + | − | + | − | + | + | − | IgG1 |
| S24.H | + | − | + | − | − | − | − | IgM |
| S27.D | + | − | + | − | + | + | + | IgG1 |
| S28.F | + | − | + | − | + | + | + | IgG1 |
| S30.B | + | − | − | − | + | − | − | IgG1 |
| S33.B | + | − | + | − | + | + | + | IgG1 |
| S34.G | + | − | + | − | + | + | − | IgG2b |
| S35.I | + | − | + | − | + | + | − | IgG1 |
| S36.E | + | − | − | − | − | − | − | IgG1 |
| S37.E | + | − | − | − | − | − | − | IgG1 |
| S38.C | + | − | − | − | + | + | − | IgG1 |
| S39.B | + | − | − | − | − | − | − | IgG1 |
| S40.B | + | − | − | − | + | − | − | IgG1 |
| S43.E | + | − | + | − | + | + | + | IgG1 |
| S45.A | + | − | − | − | + | − | − | IgG1 |
| S46.C | + | − | − | − | + | − | − | IgG1 |
| S48.E | + | − | − | − | + | − | − | IgG1 |
| S49.C | + | − | − | − | − | − | − | IgG1 |
| S52.C | + | − | + | − | + | + | − | IgG2b |
| S53.B | + | − | + | − | + | + | − | IgG1 |
| Mab (Rabbit) | CoV-2 S-RBD (intact) | CoV-2 S-RBD (D and R) | Comp with S3.E b | Comp with S8.C f | Comp with S11.A g | Comp with S43.E h | Neut (Native) | Isotype |
| S #14 | + | − | + | + | − | + | + | IgG |
| S #24 | + | + | − | − | + | − | − | IgG |
| S #77 | + | − | + | + | − | + | − | IgG |
| Antibody Designation a | Origin and Isotype | ELISA Titer b | Neutralizing Titer c | Ratio d |
|---|---|---|---|---|
| S3.E (cell culture s/n e) | Mouse Mab (IgM) | 800 | 640 | 1.25 |
| S3.E (CELLine s/n) | Mouse Mab (IgM) | 10,000 | 1280 | 7.8 |
| S8.C (cell culture s/n) | Mouse Mab (IgG2b) | 1000 | 10 | 100 |
| S8.C (CELLine s/n) | Mouse Mab (IgG2b) | 20,000 | 320 | 62.5 |
| S27.D (cell culture s/n) | Mouse Mab (IgG1) | 2000 | 10 | 200 |
| S28.F (cell culture s/n) | Mouse Mab (IgG1) | 2000 | 10 | 200 |
| S33.B (cell culture s/n) | Mouse Mab (IgG1) | 2000 | 10 | 200 |
| S43.E (cell culture s/n) | Mouse Mab (IgG1) | 2000 | 40 | 50 |
| S #14 (CELLine s/n) | Rabbit Mab (IgG) | 1000 | 10 | 100 |
| S #24 (CELLine s/n) | Rabbit Mab (IgG) | 10,000 | 0 | N/A |
| S #77 (CELLine s/n) | Rabbit Mab (IgG) | 800 | 0 | N/A |
| S 5393 (Rabbit serum) | Rabbit Pab | 500 | 80 | 6.25 |
| N 5394 (Rabbit serum) | Rabbit Pab (anti-N) | <10 | 0 | N/A |
| ELISA a Signals for Various Antigens | |||||||
|---|---|---|---|---|---|---|---|
| Mab Clone (Mouse) | CoV-2 b (intact) | CoV-2 c (D and R) | CoV d (Intact) | MERS e (Intact) | Region (aa) f | Western g (CoV-2) | Isotype |
| N1.15C | + | + | − | − | 364–419 | + | IgG1 |
| N1.17A | + | + | + | + | 247–364 | + | IgG2b |
| N2.4K | + | + | + | + | 215–247 | + | IgG1 |
| N2.11K | + | + | + | + | 174–215 | + | IgG1 |
| N2.12A | + | + | + | + | 47–174 | − | IgG1 |
| N2.20A | + | − | − | − | 247–364 | + | IgG1 |
| N3.1B | + | + | + | + | 247–364 | − | IgG1 |
| N3.2A | + | + | + | + | 247–364 | + | IgG1 |
| N3.3B | + | + | + | + | 247–364 | + | IgG2b |
| N3.8C | + | + | + | + | 247–364 | + | IgG1 |
| N3.19A | + | + | + | + | 47–174 | + | IgG1 |
| N4.4F | + | + | + | + | 247–364 | − | IgG1 |
| N4.5D | + | − | − | − | 247–364 | − | IgG1 |
| N4.6A | + | + | + | + | 215–247 | + | IgM |
| N4.10A | + | + | + | + | 247–364 | + | IgG1 |
| N4.12B | + | + | + | − | 247–364 | − | IgG1 |
| N4.16J | + | + | + | + | 247–364 | + | IgG1 |
| N4.17E | + | + | + | − | 247–364 | − | IgG1 |
| N5.6C | + | + | + | + | 247–364 | + | IgG1 |
| N5.9A | + | + | + | − | 364–419 | − | IgG1 |
| N5.13B | + | + | + | + | 247–364 | − | IgG1 |
| N5.16A | + | + | + | − | 247–364 | − | IgG1 |
| N5.17B11 | + | + | + | − | 247–364 | − | IgG1 |
| N5.18D | + | + | + | − | 247–364 | − | IgG1 |
| N6.8A | + | + | + | + | 247–364 | − | IgG1 |
| N6.11A | + | + | + | + | 247–364 | − | IgG1 |
| N6.12B | + | + | + | − | 247–364 | − | IgG1 |
| N6.22A | + | + | + | − | 247–364 | − | IgG1 |
| N7.2A | + | + | + | + | 47–174 | + | IgG1 |
| N7.28C | + | + | + | + | 247–364 | + | IgG2b |
| N7.17B | + | + | + | − | 247–364 | − | IgG1 |
| N7.19A | + | + | + | − | 47–174 | − | IgG1 |
| N7.22D | + | + | + | − | 247–364 | − | IgG2a |
| N7.26A | + | + | + | + | 247–364 | + | IgG2b |
| N8.1E | + | + | + | + | 364–419 | − | IgG2b |
| N8.2B | + | + | + | − | 247–364 | − | IgG1 |
| N8.3B | + | + | − | − | 247–364 | + | IgG2b |
| N8.5H | + | + | + | − | 364–419 | + | IgG1 |
| N8.8E | + | + | + | + | 47–174 | + | IgG1 |
| N8.29B | + | + | + | − | 247–364 | − | IgG1 |
| N9.1B | + | + | + | + | 247–364 | − | IgG1 |
| N9.4B | + | + | + | + | 247–364 | − | IgG1 |
| N9.7A | + | + | + | − | 247–364 | − | IgG1 |
| N9.9B | + | + | + | − | 47–174 | − | IgG1 |
| N9.10B | + | + | + | + | 47–174 | − | IgG2b |
| N9.13A | + | + | + | + | 247–364 | − | IgG1 |
| N9.14C | + | + | + | + | 364–419 | − | IgG1 |
| N9.15A | + | + | + | + | 247–364 | − | IgG1 |
| N9.18B | + | + | + | − | 247–364 | − | IgG1 |
| N9.23I | + | + | + | − | 247–364 | − | IgG1 |
| N9.24G | + | + | + | + | 247–364 | − | IgG1 |
| N9.27C | + | + | + | − | 364–419 | − | IgG1 |
| N10.4I | + | + | + | − | 247–364 | − | IgG1 |
| N10.6B | + | + | + | + | 247–364 | − | IgG1 |
| N10.13C | + | − | − | − | 247–364 | − | IgG1 |
| N10.15H | + | + | − | + | 215–247 | − | IgG1 |
| N10.17D | + | + | + | + | 247–364 | − | IgG1 |
| N10.21E | + | + | + | + | 247–364 | − | IgG1 |
| N10.25A | + | + | + | + | 247–364 | − | IgG1 |
| N10.29B | + | + | − | − | 1–47 | − | IgG1 |
| Mab clone (Rabbit) | CoV-2 b (intact) | CoV-2 c(D and R) | CoV d (intact) | MERS e (intact) | Region (aa) f | Western g (CoV-2) | Isotype |
| N #54 | + | + | + | − | 47–174 | NT | IgG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, R.; Christensen, N.D.; Kaddis Maldonado, R.J.; Bewley, M.C.; Ostman, A.; Sudol, M.; Chen, E.C.; Buchkovich, N.W.; Gontu, A.; Surendran Nair, M.; et al. Monoclonal Antibodies to S and N SARS-CoV-2 Proteins as Probes to Assess Structural and Antigenic Properties of Coronaviruses. Viruses 2021, 13, 1899. https://doi.org/10.3390/v13101899
Kumar R, Christensen ND, Kaddis Maldonado RJ, Bewley MC, Ostman A, Sudol M, Chen EC, Buchkovich NW, Gontu A, Surendran Nair M, et al. Monoclonal Antibodies to S and N SARS-CoV-2 Proteins as Probes to Assess Structural and Antigenic Properties of Coronaviruses. Viruses. 2021; 13(10):1899. https://doi.org/10.3390/v13101899
Chicago/Turabian StyleKumar, Rinki, Neil D. Christensen, Rebecca J. Kaddis Maldonado, Maria C. Bewley, Alexandria Ostman, Malgorzata Sudol, Eunice C. Chen, Natalie W. Buchkovich, Abhinay Gontu, Meera Surendran Nair, and et al. 2021. "Monoclonal Antibodies to S and N SARS-CoV-2 Proteins as Probes to Assess Structural and Antigenic Properties of Coronaviruses" Viruses 13, no. 10: 1899. https://doi.org/10.3390/v13101899
APA StyleKumar, R., Christensen, N. D., Kaddis Maldonado, R. J., Bewley, M. C., Ostman, A., Sudol, M., Chen, E. C., Buchkovich, N. W., Gontu, A., Surendran Nair, M., Nissly, R. H., Minns, A. M., Kapur, V., Rossi, R., Kuchipudi, S. V., Lindner, S. E., Parent, L. J., Flanagan, J. M., & Buchkovich, N. J. (2021). Monoclonal Antibodies to S and N SARS-CoV-2 Proteins as Probes to Assess Structural and Antigenic Properties of Coronaviruses. Viruses, 13(10), 1899. https://doi.org/10.3390/v13101899