
Staphylococcal Phage in Combination with Staphylococcus epidermidis as a Potential Treatment for Staphylococcus aureus-Associated Atopic Dermatitis and Suppressor of Phage-Resistant Mutants

 , , , and
, , , and 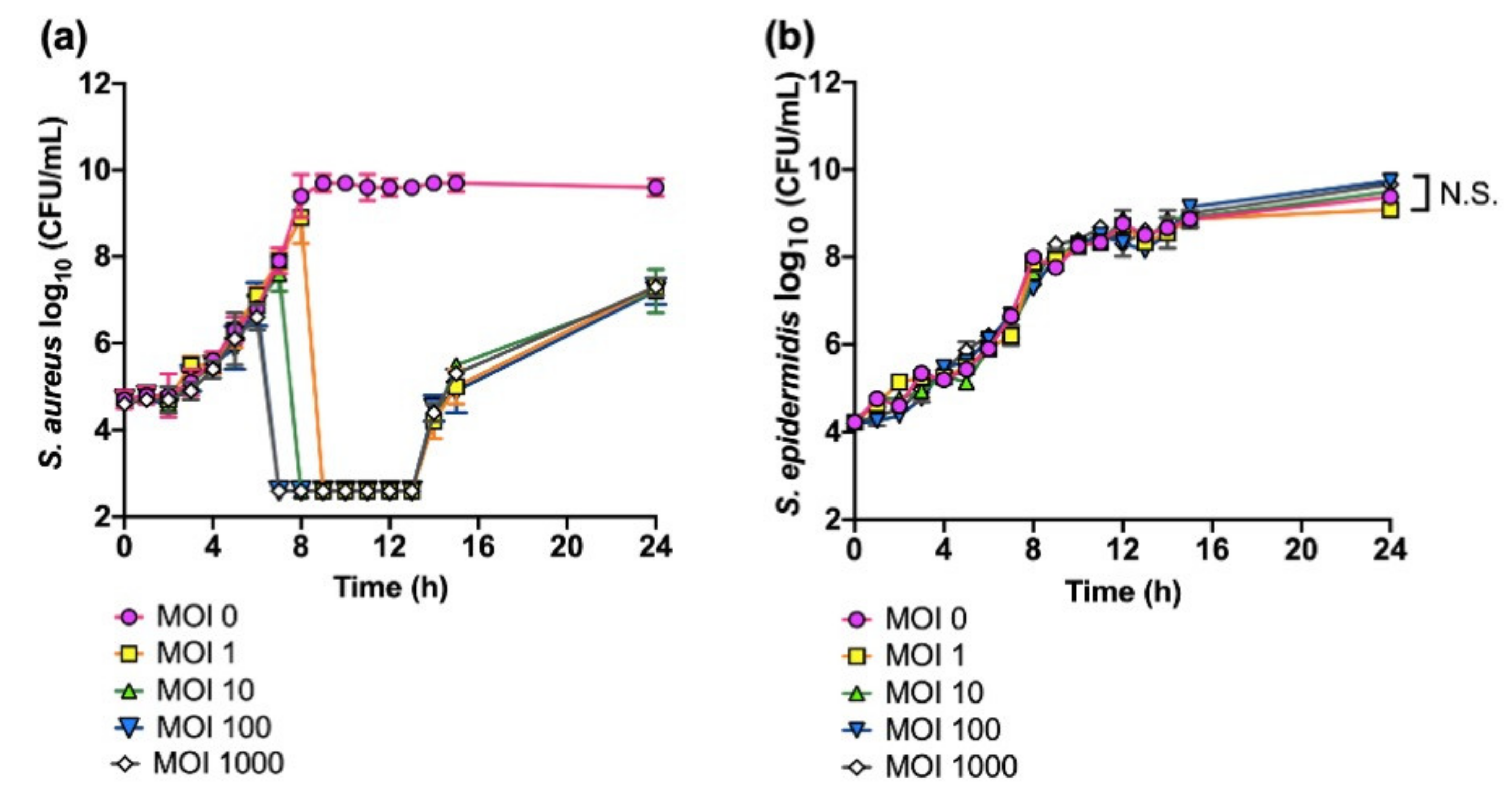{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Bacterial Growth Conditions
2.2. In Vitro Bacterial Killing Assays
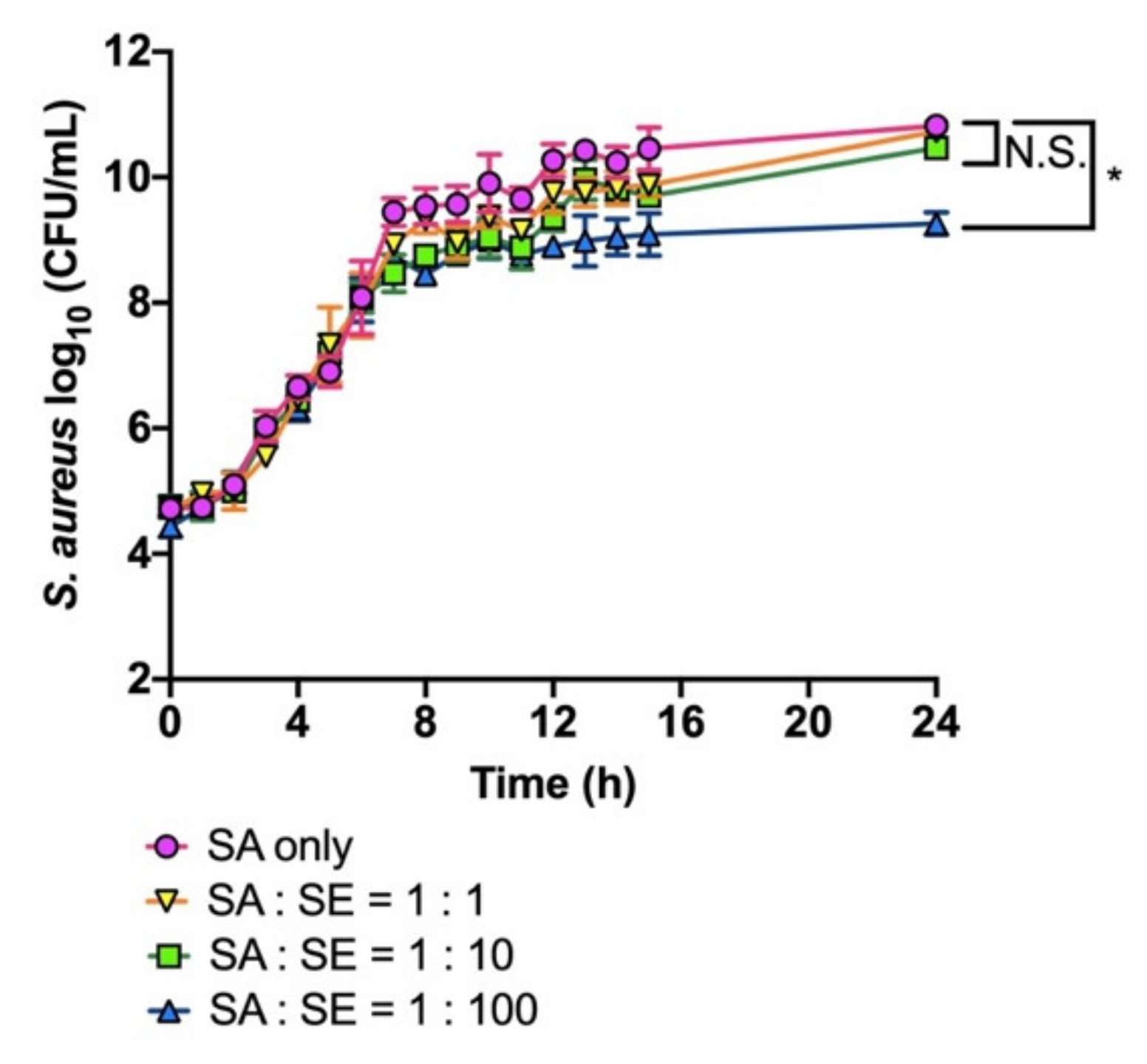
2.3. Co-Cultivation of S. aureus and S. epidermidis
2.4. Plaque Formation Assays
2.5. Adsorption Assays
2.6. Animal Test
2.7. Statistical Analysis
3. Results
3.1. Growth Inhibition of S. aureus
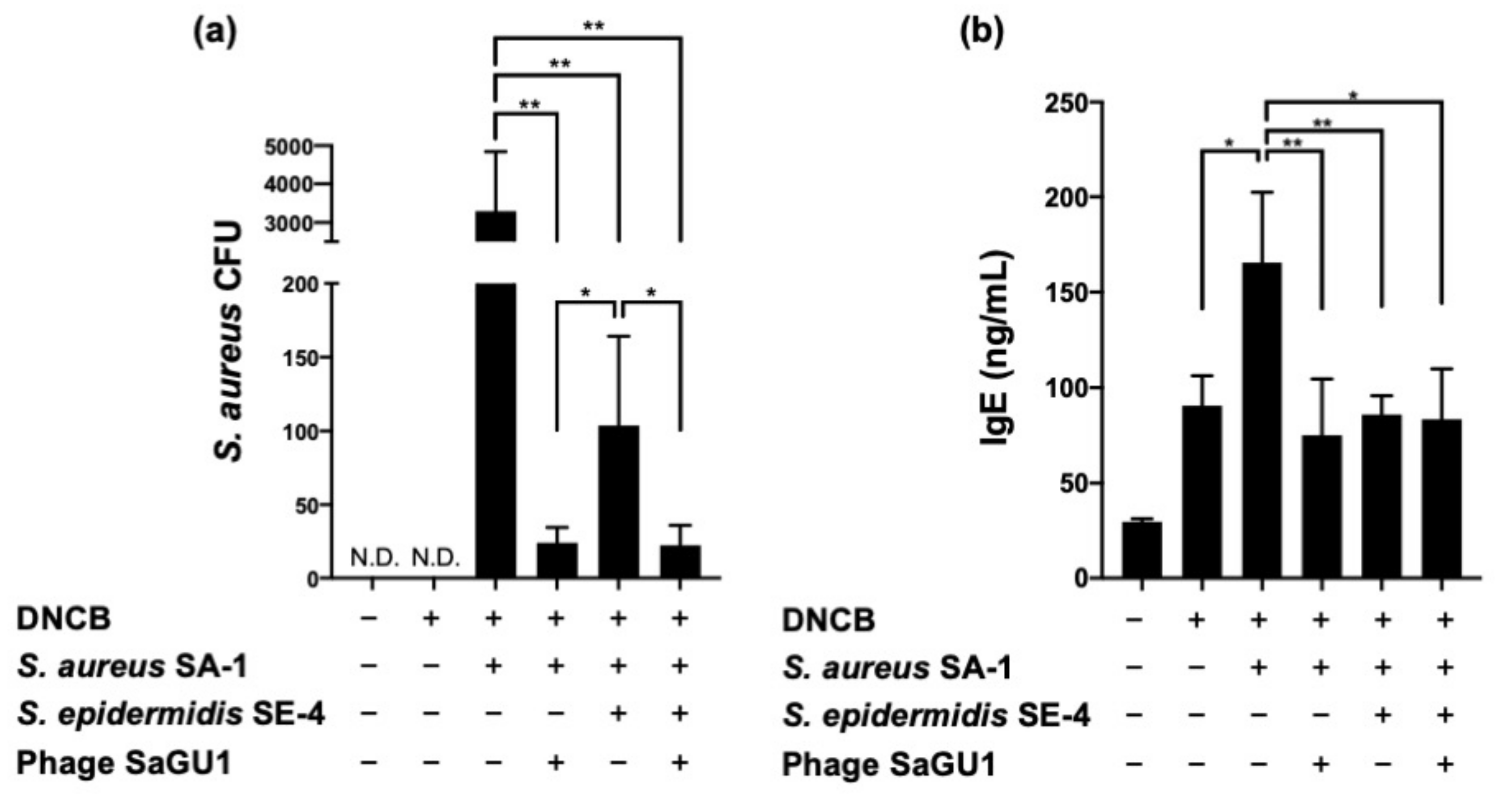
3.2. Effect of Phage SaGU1 in Eliminating S. aureus and on Improving Atopic Symptoms
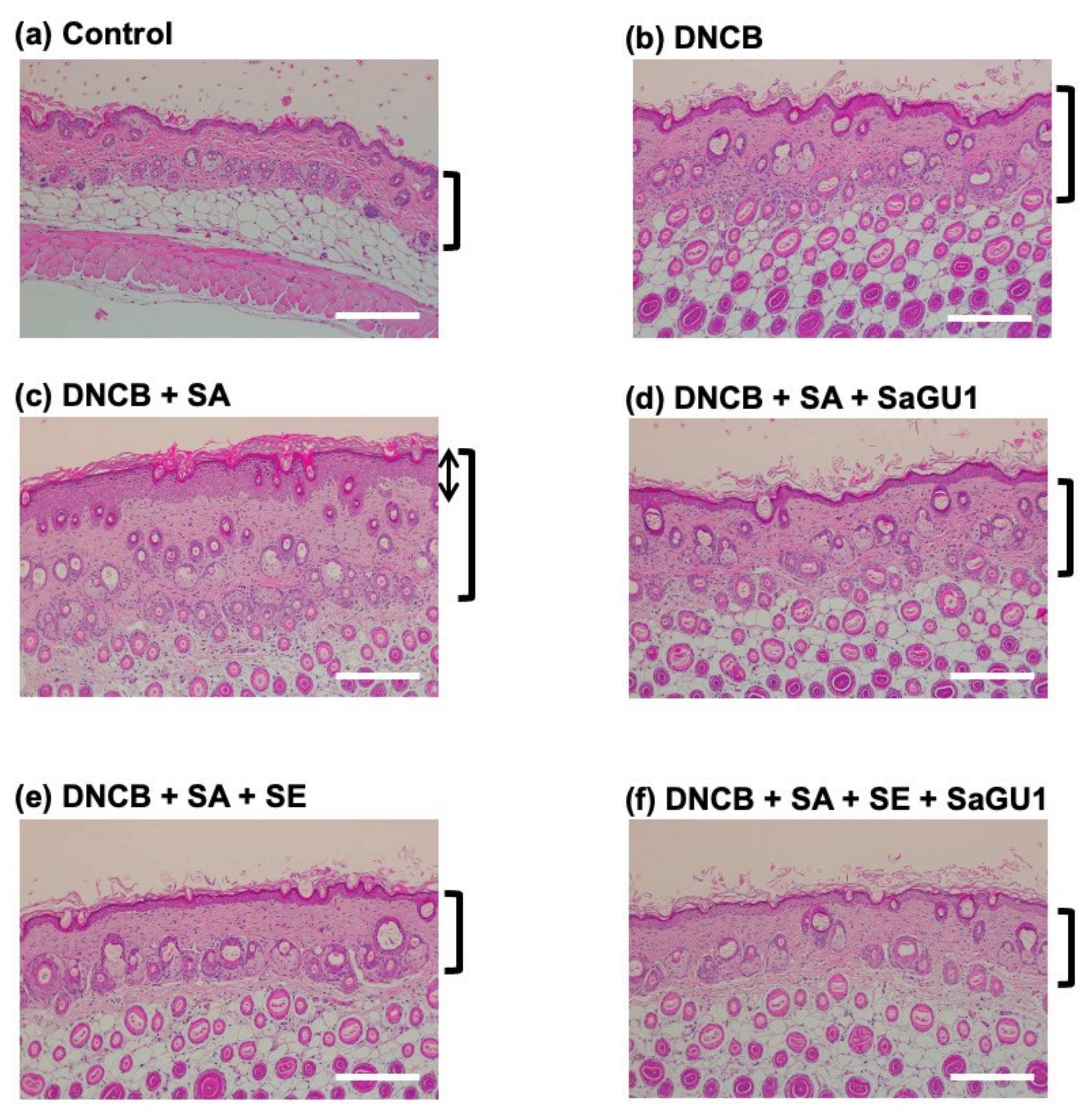
3.3. Improvement of Atopic Dermatitis Symptoms by Phage Therapy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Geoghegan, J.A.; Irvine, A.D.; Foster, T.J. Staphylococcus aureus and atopic dermatitis: A complex and evolving relationship. Trends Microbiol. 2018, 26, 484–497. [Google Scholar] [CrossRef] [PubMed]
- Gordillo Altamirano, F.L.; Barr, J.J. Phage therapy in the postantibiotic era. Clin. Microbiol. Rev. 2019, 32, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Nakatsuji, T.; Chen, T.H.; Narala, S.; Chun, K.A.; Two, A.M.; Yun, T.; Shafiq, F.; Kotol, P.F.; Bouslimani, A.; Melnik, A.V.; et al. Antimicrobials from human skin commensal bacteria protect against Staphylococcus aureus and are deficient in atopic dermatitis. Sci. Transl. Med. 2017, 9, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Koskella, B.; Meaden, S. Understanding bacteriophage specificity in natural microbial communities. Viruses 2013, 5, 806–823. [Google Scholar] [CrossRef] [PubMed]
- Maura, D.; Debarbieux, L. On the interactions between virulent bacteriophages and bacteria in the gut. Bacteriophage 2012, 2, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Paule, A.; Frezza, D.; Edeas, M. Microbiota and phage therapy: Future challenges in medicine. Med. Sci. 2018, 6, 86. [Google Scholar] [CrossRef] [PubMed]
- Reyes, A.; Wu, M.; McNulty, N.P.; Rohwer, F.L.; Gordon, J.I. Gnotobiotic mouse model of phage-bacterial host dynamics in the human gut. Proc. Natl. Acad. Sci. USA 2013, 110, 20236–20241. [Google Scholar] [CrossRef]
- Chadha, P.; Katare, O.P.; Chhibber, S. In vivo efficacy of single phage versus phage cocktail in resolving burn wound infection in BALB/c mice. Microb. Pathog. 2016, 99, 68–77. [Google Scholar] [CrossRef]
- Fish, R.; Kutter, E.; Wheat, G.; Blasdel, B.; Kutateladze, M.; Kuhl, S. Compassionate use of bacteriophage therapy for foot ulcer treatment as an effective step for moving toward clinical trials. Methods Mol. Biol. 2018, 1693, 159–170. [Google Scholar] [CrossRef]
- Wright, A.; Hawkins, C.H.; Änggård, E.E.; Harper, D.R. A controlled clinical trial of a therapeutic bacteriophage preparation in chronic otitis due to antibiotic-resistant Pseudomonas aeruginosa; a preliminary report of efficacy. Clin. Otolaryngol. 2009, 34, 349–357. [Google Scholar] [CrossRef]
- Shimamori, Y.; Pramono, A.K.; Kitao, T.; Suzuki, T.; Aizawa, S.; Kubori, T.; Nagai, H.; Takeda, S.; Ando, H. Isolation and characterization of a novel phage SaGU1 that infects Staphylococcus aureus clinical isolates from patients with atopic dermatitis. bioRxiv 2020. [Google Scholar] [CrossRef]
- Ando, H.; Lemire, S.; Pires, D.; Lu, T.K. Engineering modular viral scaffolds for targeted bacterial population editing. Cell Syst. 2015, 1, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Lee, J.H.; Shin, S.J.; Cho, A.R.; Heo, Y. Molecular mechanism of atopic dermatitis induction following sensitization and challenge with 2,4-dinitrochlorobenzene in mouse skin tissue. Toxicol. Res. 2018, 34, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Baek, S.-J.; Kim, H.-A.; Heo, Y. 2,4-Dinitrochlorobenzene-induced atopic dermatitis like immune alteration in mice. J. Toxicol. Public Health 2006, 22, 357–364. [Google Scholar]
- Hofer, M.F.; Harbeck, R.J.; Schlievert, P.M.; Leung, D.Y. Staphylococcal toxins augment specific IgE responses by atopic patients exposed to allergen. J. Invest. Dermatol. 1999, 112, 171–176. [Google Scholar] [CrossRef][Green Version]
- Tomczak, H.; Wróbel, J.; Jenerowicz, D.; Sadowska-Przytocka, A.; Wachal, M.; Adamski, Z.; Czarnecka-Operacz, M.M. The role of Staphylococcus aureus in atopic dermatitis: Microbiological and immunological implications. Adv. Dermatol. Allergol. 2019, 36, 485–491. [Google Scholar] [CrossRef]
- Abedon, S.T. Phage-antibiotic combination treatments: Antagonistic impacts of antibiotics on the pharmacodynamics of phage therapy? Antibiotics 2019, 8. [Google Scholar] [CrossRef]
- Nakatsuji, T.; Gallo, R.L. The role of the skin microbiome in atopic dermatitis. Ann. Allergy Asthma Immunol. 2019, 122, 263–269. [Google Scholar] [CrossRef]
- Islam, S.U. Clinical uses of probiotics. Medicine 2016, 95, e2658. [Google Scholar] [CrossRef]
- Cremonini, F.; Di Caro, S.; Nista, E.C.; Bartolozzi, F.; Capelli, G.; Gasbarrini, G.; Gasbarrini, A. Meta-analysis: The effect of probiotic administration on antibiotic-associated diarrhoea. Aliment. Pharmacol. Ther. 2002, 16, 1461–1467. [Google Scholar] [CrossRef]
- Wang, Z.; He, Y.; Zheng, Y. Probiotics for the treatment of bacterial vaginosis: A meta-analysis. Int. J. Environ. Res. Public Health 2019, 16, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Goderska, K.; Agudo Pena, S.; Alarcon, T. Helicobacter pylori treatment: Antibiotics or probiotics. Appl. Microbiol. Biotechnol. 2018, 102, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Rautava, S.; Kalliomäki, M.; Isolauri, E. Probiotics during pregnancy and breast-feeding might confer immunomodulatory protection against atopic disease in the infant. J. Allergy Clin. Immunol. 2002, 109, 119–121. [Google Scholar] [CrossRef] [PubMed]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The human skin microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef]
- Lai, Y.; Cogen, A.L.; Radek, K.A.; Park, H.J.; Macleod, D.T.; Leichtle, A.; Ryan, A.F.; Di Nardo, A.; Gallo, R.L. Activation of TLR2 by a small molecule produced by Staphylococcus epidermidis increases antimicrobial defense against bacterial skin infections. J. Investig. Dermatol. 2010, 130, 2211–2221. [Google Scholar] [CrossRef]
- Li, D.; Lei, H.; Li, Z.; Li, H.; Wang, Y.; Lai, Y. A novel lipopeptide from skin commensal activates TLR2/CD36-p38 MAPK signaling to increase antibacterial defense against bacterial infection. PLoS One 2013, 8. [Google Scholar] [CrossRef]
- Abedon, S.T. Phage therapy dosing: The problem(s) with multiplicity of infection (MOI). Bacteriophage 2016, 6, e1220348. [Google Scholar] [CrossRef]
- Raz, A.; Serrano, A.; Hernandez, A.; Euler, C.W.; Fischetti, A. Isolation of phage lysins that effectively kill Pseudomonas aeruginosa in mouse models of lung and skin infection. Antimicrob Agents Chemother. 2019, 63, e00024-19. [Google Scholar] [CrossRef]
- Trigo, G.; Martins, T.G.; Fraga, A.G.; Longatto-Filho, A.; Castro, A.G.; Azeredo, J.; Pedrosa, J. Phage therapy is effective against infection by Mycobacterium ulcerans in a murine footpad model. PLoS Negl. Trop. Dis. 2013, 7. [Google Scholar] [CrossRef]
- D’Accolti, M.; Soffritti, I.; Lanzoni, L.; Bisi, M.; Volta, A.; Mazzacane, S.; Caselli, E. Effective elimination of Staphylococcal contamination from hospital surfaces by a bacteriophage–probiotic sanitation strategy: A monocentric study. Microb. Biotechnol. 2019, 12, 742–751. [Google Scholar] [CrossRef]
- Woo, J.; Ahn, J. Assessment of synergistic combination potential of probiotic and bacteriophage against antibiotic-resistant Staphylococcus aureus exposed to simulated intestinal conditions. Arch. Microbiol. 2014, 196, 719–727. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shimamori, Y.; Mitsunaka, S.; Yamashita, H.; Suzuki, T.; Kitao, T.; Kubori, T.; Nagai, H.; Takeda, S.; Ando, H. Staphylococcal Phage in Combination with Staphylococcus epidermidis as a Potential Treatment for Staphylococcus aureus-Associated Atopic Dermatitis and Suppressor of Phage-Resistant Mutants. Viruses 2021, 13, 7. https://doi.org/10.3390/v13010007
Shimamori Y, Mitsunaka S, Yamashita H, Suzuki T, Kitao T, Kubori T, Nagai H, Takeda S, Ando H. Staphylococcal Phage in Combination with Staphylococcus epidermidis as a Potential Treatment for Staphylococcus aureus-Associated Atopic Dermatitis and Suppressor of Phage-Resistant Mutants. Viruses. 2021; 13(1):7. https://doi.org/10.3390/v13010007
Chicago/Turabian StyleShimamori, Yuzuki, Shoichi Mitsunaka, Hirotaka Yamashita, Tohru Suzuki, Tomoe Kitao, Tomoko Kubori, Hiroki Nagai, Shigeki Takeda, and Hiroki Ando. 2021. "Staphylococcal Phage in Combination with Staphylococcus epidermidis as a Potential Treatment for Staphylococcus aureus-Associated Atopic Dermatitis and Suppressor of Phage-Resistant Mutants" Viruses 13, no. 1: 7. https://doi.org/10.3390/v13010007
APA StyleShimamori, Y., Mitsunaka, S., Yamashita, H., Suzuki, T., Kitao, T., Kubori, T., Nagai, H., Takeda, S., & Ando, H. (2021). Staphylococcal Phage in Combination with Staphylococcus epidermidis as a Potential Treatment for Staphylococcus aureus-Associated Atopic Dermatitis and Suppressor of Phage-Resistant Mutants. Viruses, 13(1), 7. https://doi.org/10.3390/v13010007