
Surveillance and Genetic Characterization of Virulent Newcastle Disease Virus Subgenotype V.3 in Indigenous Chickens from Backyard Poultry Farms and Live Bird Markets in Kenya

 ,
,  ,
,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
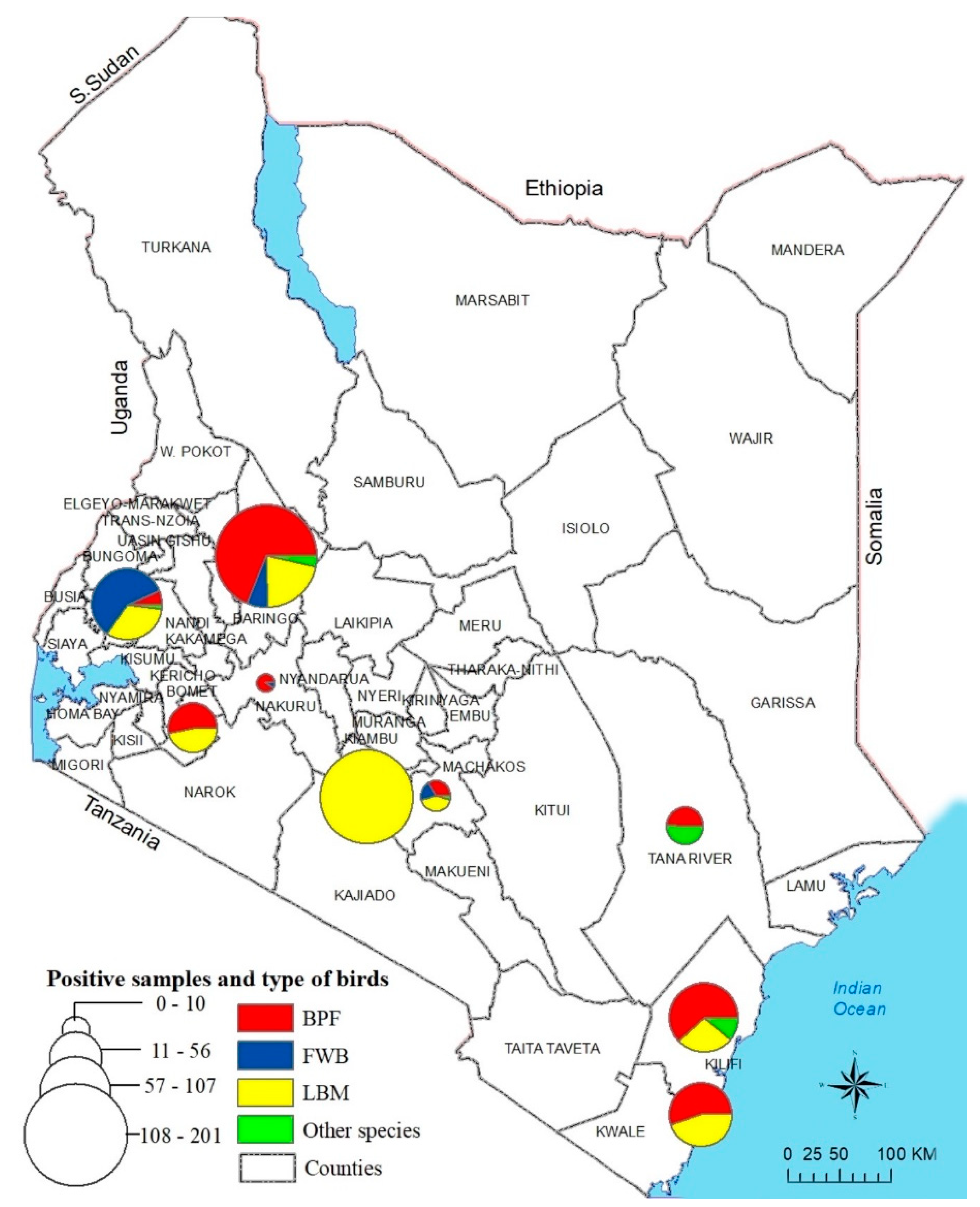
2.1. Sampling Sites
2.2. Sample Collection
2.3. Total RNA Extraction and Virus Detection
2.4. Virus Isolation
2.5. Next-Generation Sequencing
2.6. Genome Sequence Assembly and Characterization
2.6.1. Sequence Assembly and Phylogenetic Analyses
2.6.2. Bayesian Evolutionary and Spatial-Temporal Dispersal Inferences
3. Results
3.1. Sample Analyses
3.1.1. Sample Collection
3.1.2. Vaccination Histories and Clinical Signs Consistent with ND
3.2. Virus Detection
3.3. Characteristics of the Identified Kenyan NDV Isolates
3.3.1. Genomic Characteristics
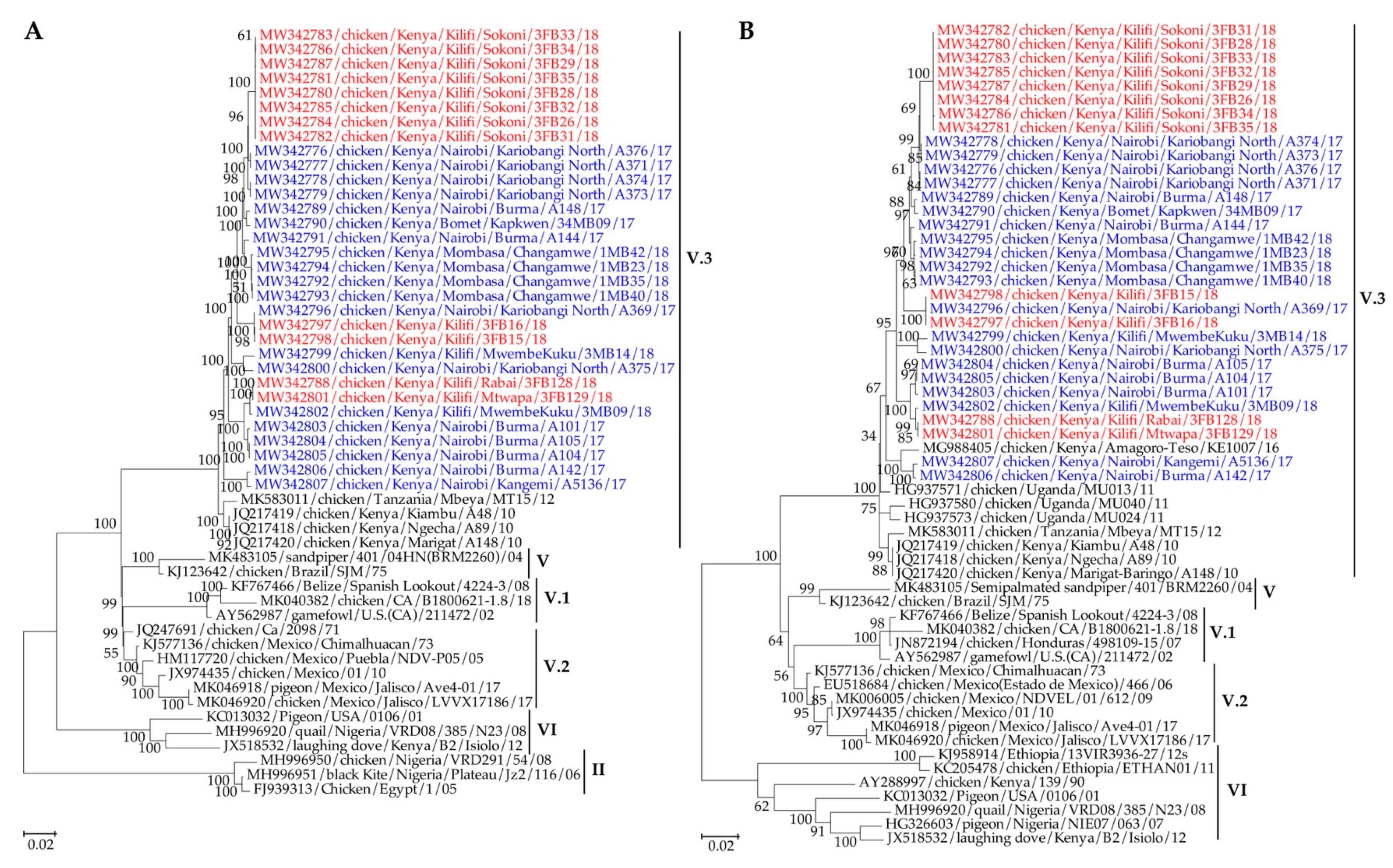
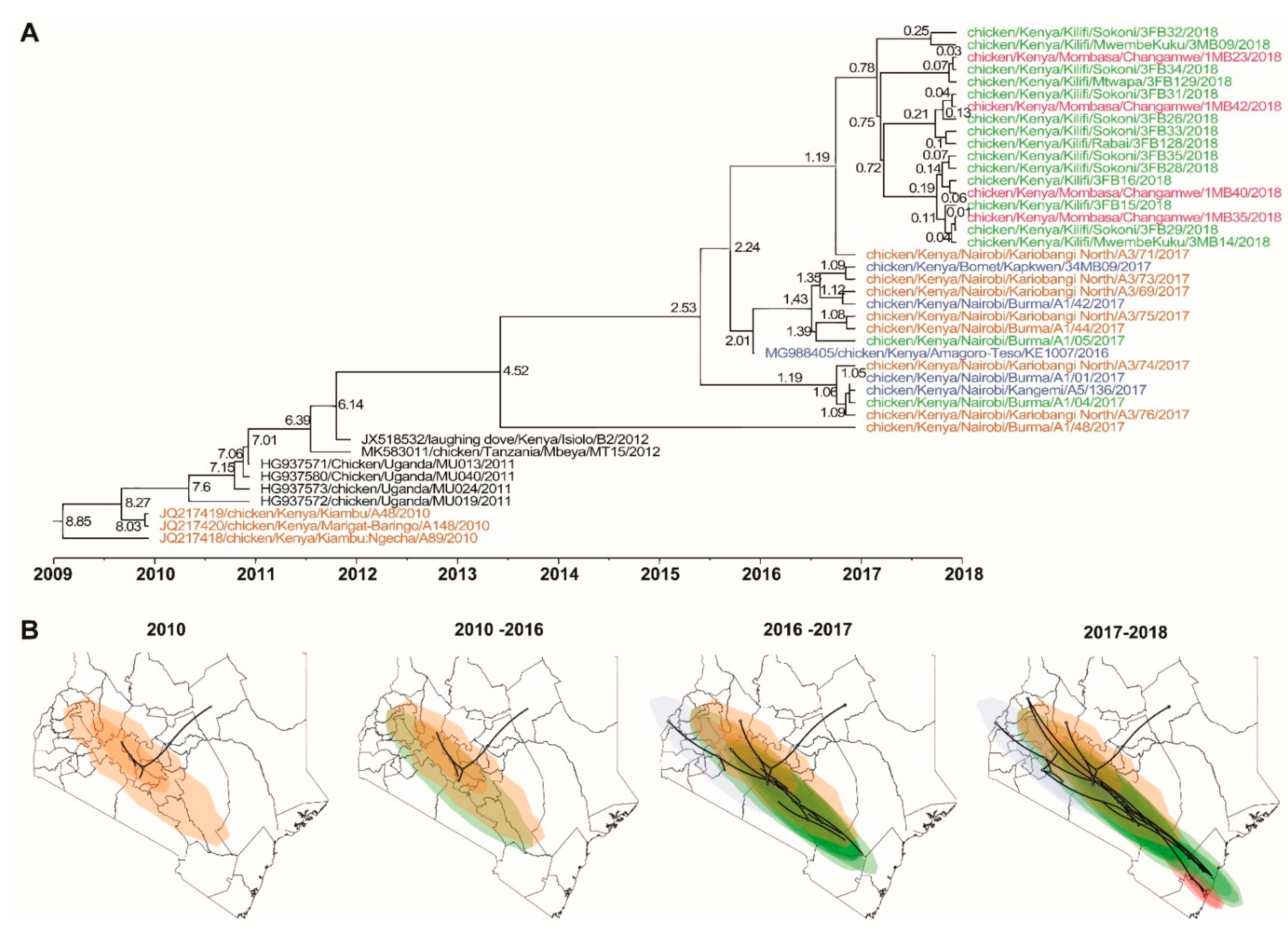
3.3.2. Phylogenetic and Phylogeographic Analyses
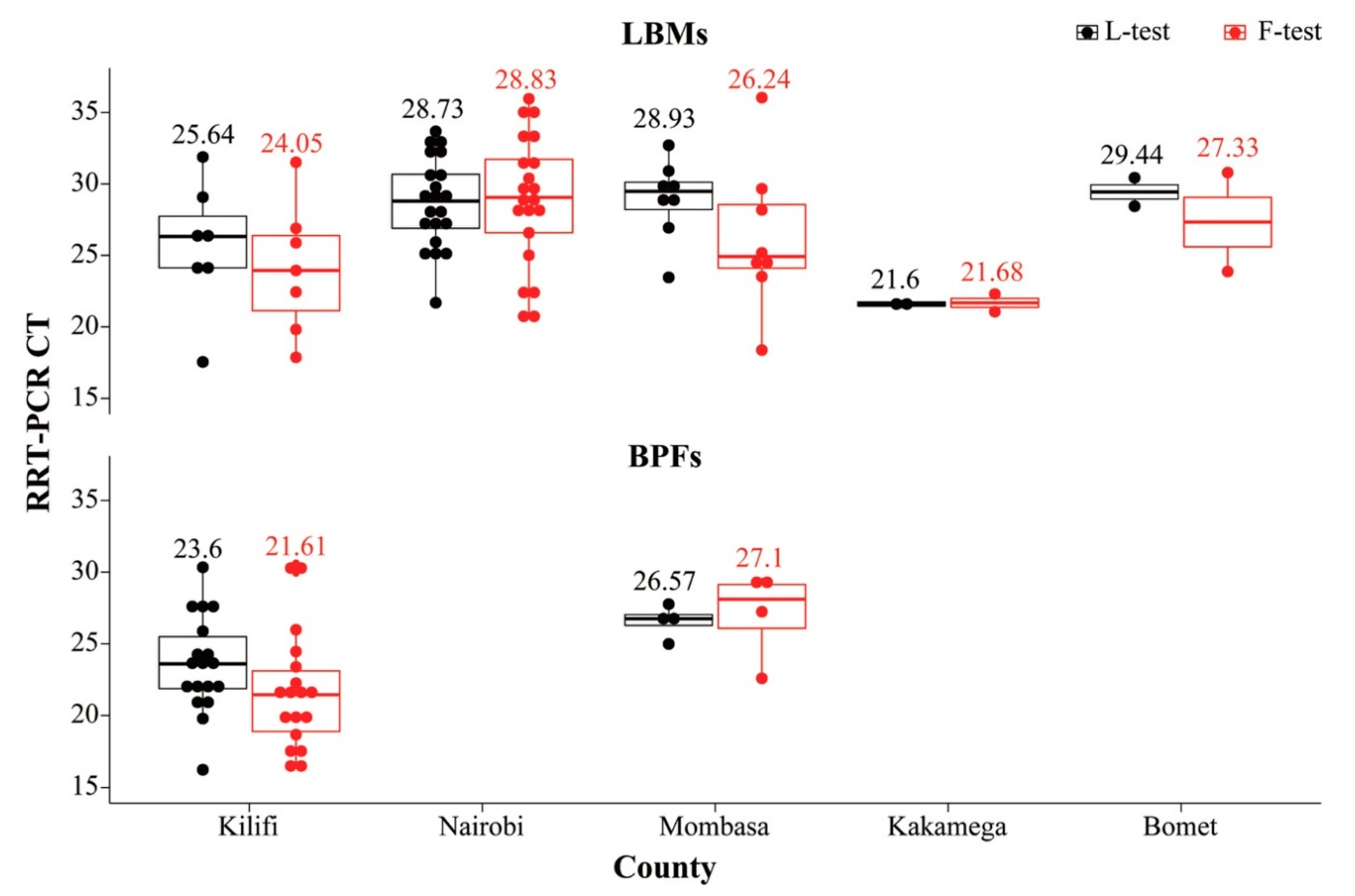
3.3.3. Biological Characteristics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kitalyi, A.J. Village Chicken Production Systems in Rural Africa: Household Food Security and Gender Issues; FAO Animal Production and Health Paper 142; FAO: Rome, Italy, 1998; ISBN 92-5-104160-1. [Google Scholar]
- Kemboi, D.; Chegeh, H.; Bebora, L.; Maingi, N.; Nyaga, P.; Mbuthia, P.; Njagi, L.; Githinji, J. Seasonal Newcastle disease antibody titer dynamics in village chickens of Mbeere District, Eastern Province, Kenya. Livest. Res. Rural Dev. 2013, 25, 181. [Google Scholar]
- Ipara, B.O.; Otieno, D.; Nyikal, R.A.; Makokha, S.N. The role of unregulated chicken marketing practices on the frequency of Newcastle disease outbreaks in Kenya. Poult. Sci. 2019, 98, 6356–6366. [Google Scholar] [CrossRef] [PubMed]
- Munyua, P.M.; Githinji, J.W.; Waiboci, L.W.; Njagi, L.M.; Arunga, G.; Mwasi, L.; Mbabu, R.M.; Macharia, J.M.; Breiman, R.F.; Njenga, M.K.; et al. Detection of influenza A virus in live bird markets in Kenya, 2009–2011. Influenza Other Respir. Viruses 2012, 7, 113–119. [Google Scholar] [CrossRef]
- Martin, P.A.J. The epidemiology of Newcastle disease in village chickens. In Proceedings of the Newcastle Disease in Village Chickens, Control with Thermostable Oral Vaccines; Spradbrow, P.B., Ed.; Australian Centre for International Agricultural Research: Kuala Lumpur, Malaysia, 1991; pp. 40–45. [Google Scholar]
- Njagi, L.; Nyaga, P.; Mbuthia, P.; Bebora, L.; Michieka, J.; Minga, U. A retrospective study of factors associated with Newcastle disease outbreaks in village indigenous chickens. Bull. Anim. Health Prod. Afr. 2010, 58, 22–33. [Google Scholar] [CrossRef]
- Ayala, A.J.; Hernandez, S.M.; Olivier, T.L.; Welch, C.N.; Dimitrov, K.M.; Goraichuk, I.V.; Afonso, C.L.; Miller, P.J. Experimental infection and transmission of Newcastle disease vaccine virus in four wild passerines. Avian Dis. 2019, 63, 389–399. [Google Scholar] [CrossRef]
- Welch, C.N.; Shittu, I.; Abolnik, C.; Solomon, P.; Dimitrov, K.M.; Taylor, T.L.; Williams-Coplin, D.; Goraichuk, I.V.; Meseko, C.A.; Ibu, J.O. Genomic comparison of Newcastle disease viruses isolated in Nigeria between 2002 and 2015 reveals circulation of highly diverse genotypes and spillover into wild birds. Arch. Virol. 2019, 164, 2031–2047. [Google Scholar] [CrossRef]
- Miller, P.J.; Koch, G. Newcastle disease, other avian paramyxoviruses, and avian metapneumovirus infections. In Diseases of Poultry; Swayne, D.E., Glisson, J.R., McDougald, L.R., Nolan, L.K., Suarez, D.L., Venugopal, N., Eds.; Wiley-Blackwell: Ames, IA, USA, 2013; pp. 87–138. ISBN 978-0-470-95899-5. [Google Scholar]
- Oakeley, R.D. The limitations of a feed/water-based heat-stable vaccine delivery system for Newcastle disease-control strategies for backyard poultry flocks in sub-Saharan Africa. Prev. Vet. Med. 2000, 47, 271–279. [Google Scholar] [CrossRef]
- Dimitrov, K.M.; Lee, D.-H.; Williams-Coplin, D.; Olivier, T.L.; Miller, P.J.; Afonso, C.L. Newcastle disease viruses causing recent outbreaks worldwide show unexpectedly high genetic similarity to historical virulent isolates from the 1940s. J. Clin. Microbiol. 2016, 54, 1228–1235. [Google Scholar] [CrossRef]
- OIE World Animal Health Information Database (WAHIS Interface): Detailed Country (Ies) Disease Incidence; WAHIS portal: Animal Health Data; World Organization for Animal Health (OIE): Paris, France, 2020.
- Cattoli, G.; Fusaro, A.; Monne, I.; Molia, S.; Le Menach, A.; Maregeya, B.; Nchare, A.; Bangana, I.; Maina, A.G.; Koffi, J.-N.; et al. Emergence of a new genetic lineage of Newcastle disease virus in West and Central Africa—implications for diagnosis and control. Vet. Microbiol. 2010, 142, 168–176. [Google Scholar] [CrossRef][Green Version]
- Kuhn, J.H.; Wolf, Y.I.; Krupovic, M.; Zhang, Y.-Z.; Maes, P.; Dolja, V.V.; Koonin, E.V. Classify viruses—The gain is worth the pain. Nature 2019, 566, 318–320. [Google Scholar] [CrossRef]
- OIE Chapter 10.9. Newcastle Disease (Article 10.9.1). In Terrestrial Animal Health Code 2014; OIE, Ed.; World Organization for Animal Health (OIE): Paris, France, 2014; Volume II, pp. 515–524. ISBN 978-92-9044-909-6. [Google Scholar]
- Steward, M.; Vipond, I.B.; Millar, N.S.; Emmerson, P.T. RNA editing in Newcastle disease virus. J. Gen. Virol. 1993, 74, 2539–2547. [Google Scholar] [CrossRef] [PubMed]
- Cattoli, G.; Susta, L.; Terregino, C.; Brown, C. Newcastle disease: A review of field recognition and current methods of laboratory detection. J. Vet. Diagn. Investig. 2011, 23, 637–656. [Google Scholar] [CrossRef] [PubMed]
- Alexander, D.J.; Senne, D.A. Newcastle disease and other avian paramyxoviruses. In A Laboratory Manual for the Isolation, Identification and Characterization of Avian Pathogens; Dufour-Zavala, L., Swayne, D.E., Glisson, J.R., Reed, W.M., Jackwood, M.W., Pearson, J.E., Eds.; American Association of Avian Pathologists: Athens, GA, USA, 2008; pp. 135–141. ISBN 978-0-9789163-2-9. [Google Scholar]
- Dimitrov, K.M.; Abolnik, C.; Afonso, C.L.; Albina, E.; Bahl, J.; Berg, M.; Briand, F.-X.; Brown, I.H.; Choi, K.-S.; Chvala, I.; et al. Updated unified phylogenetic classification system and revised nomenclature for Newcastle disease virus. Infect. Genet. Evol. 2019, 74, 103917. [Google Scholar] [CrossRef] [PubMed]
- Ballagi-Pordány, A.; Wehmann, E.; Herczeg, J.; Belák, S.; Lomniczi, B. Identification and grouping of Newcastle disease virus strains by restriction site analysis of a region from the F gene. Arch. Virol. 1996, 141, 243–261. [Google Scholar] [CrossRef]
- Hanson, R.; Spalatin, J.; Jacobson, G. The viscerotropic pathotype of Newcastle disease virus. Avian Dis. 1973, 17, 354–361. [Google Scholar] [CrossRef]
- Fernandes, C.C.; Varani, A.M.; Lemos, E.G.M.; de Miranda, V.F.O.; Silva, K.R.; Fernando, F.S.; Montassier, M.F.S.; Montassier, H.J. Molecular and phylogenetic characterization based on the complete genome of a virulent pathotype of Newcastle disease virus isolated in the 1970s in Brazil. Infect. Genet. Evol. 2014, 26, 160–167. [Google Scholar] [CrossRef]
- Ferreira, H.L.; Taylor, T.; Absalon, A.E.; Dimitrov, K.M.; Cortés-Espinosa, D.; Butt, S.; Marín-Cruz, J.; Goraichuk, I.V.; Volkening, J.; Suarez, D.L.; et al. Presence of Newcastle disease viruses of sub-genotypes Vc and VIn in backyard chickens and in apparently healthy wild birds from Mexico in 2017. Virus Genes 2019, 55, 479–489. [Google Scholar] [CrossRef]
- Byarugaba, D.K.; Mugimba, K.K.; Omony, J.B.; Okitwi, M.; Wanyana, A.; Otim, M.O.; Kirunda, H.; Nakavuma, J.L.; Teillaud, A.; Paul, M.C. High pathogenicity and low genetic evolution of avian paramyxovirus type I (Newcastle disease virus) isolated from live bird markets in Uganda. Virol. J. 2014, 11, 173. [Google Scholar] [CrossRef]
- Msoffe, P.L.; Chiwanga, G.H.; Cardona, C.J.; Miller, P.J.; Suarez, D.L. Isolation and characterization of Newcastle disease virus from live bird markets in Tanzania. Avian Dis. 2019, 63, 634–640. [Google Scholar] [CrossRef]
- Ogali, I.N.; Wamuyu, L.W.; Lichoti, J.K.; Mungube, E.O.; Agwanda, B.; Ommeh, S.C. Molecular characterization of Newcastle disease virus from backyard poultry farms and live bird markets in Kenya. Int. J. Microbiol. 2018, 2018, 2376317. [Google Scholar] [CrossRef]
- WHO Annex 8. Viral transport media (VTM). In Collecting, Preserving and Shipping Specimens for the Diagnosis of Avian Influenza A (H5N1) Virus Infection: Guide for Field Operations; Healing, T., Ed.; World Health Organization: Geneva, Switzerland, 2006; Volume 1, pp. 42–43. ISBN WHO/CDS/EPR/ARO/2006.1. [Google Scholar]
- Ferreira, H.L.; Suarez, D.L. Single-nucleotide polymorphism analysis to select conserved regions for an improved real-time reverse transcription–PCR test specific for Newcastle disease virus. Avian Dis. 2019, 63, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Wise, M.G.; Suarez, D.L.; Seal, B.S.; Pedersen, J.C.; Senne, D.A.; King, D.J.; Kapczynski, D.R.; Spackman, E. Development of a real-time reverse-transcription PCR for detection of Newcastle disease virus RNA in clinical samples. J. Clin. Microbiol. 2004, 42, 329–338. [Google Scholar] [CrossRef] [PubMed]
- OIE Chapter 2.3.14. Newcastle disease. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals (Mammals, Birds and Bees); Biological standards commission World Organisation for Animal Health; OIE: Paris, France, 2012; Volume 1, pp. 555–573. ISBN 978-92-9044-878-5.
- Chevreux, B.; Pfisterer, T.; Drescher, B.; Driesel, A.J.; Müller, W.E.; Wetter, T.; Suhai, S. Using the miraEST assembler for reliable and automated mRNA transcript assembly and SNP detection in sequenced ESTs. Genome Res. 2004, 14, 1147–1159. [Google Scholar] [CrossRef] [PubMed]
- Afgan, E.; Baker, D.; Batut, B.; Van Den Beek, M.; Bouvier, D.; Čech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.A.; et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef]
- Dimitrov, K.M.; Sharma, P.; Volkening, J.D.; Goraichuk, I.V.; Wajid, A.; Rehmani, S.F.; Basharat, A.; Shittu, I.; Joannis, T.M.; Miller, P.J.; et al. A Robust and Cost-Effective Approach to Sequence and Analyze Complete Genomes of Small RNA Viruses. Virol. J. 2017, 14, 72. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Connaris, H.; Takimoto, T.; Russell, R.; Crennell, S.; Moustafa, I.; Portner, A.; Taylor, G. Probing the sialic acid binding site of the hemagglutinin-neuraminidase of Newcastle disease virus: Identification of key amino acids involved in cell binding, catalysis, and fusion. J. Virol. 2002, 76, 1816–1824. [Google Scholar] [CrossRef]
- Suchard, M.A.; Lemey, P.; Baele, G.; Ayres, D.L.; Drummond, A.J.; Rambaut, A. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol. 2018, 4, vey016. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Ho, S.Y.; Phillips, M.J.; Rambaut, A. Relaxed phylogenetics and dating with confidence. PLoS Biol. 2006, 4, e88. [Google Scholar] [CrossRef] [PubMed]
- Gill, M.S.; Lemey, P.; Faria, N.R.; Rambaut, A.; Shapiro, B.; Suchard, M.A. Improving Bayesian population dynamics inference: A coalescent-based model for multiple loci. Mol. Biol. Evol. 2013, 30, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A. FigTree v 1.4.4. Institute of Evolutionary Biology, University of Edinburgh, Edinburgh, UK. [Graphical Viewer Computer Program]. 2018. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 6 December 2020).
- Bielejec, F.; Baele, G.; Vrancken, B.; Suchard, M.A.; Rambaut, A.; Lemey, P. SpreaD3: Interactive visualization of spatiotemporal history and trait evolutionary processes. Mol. Biol. Evol. 2016, 33, 2167–2169. [Google Scholar] [CrossRef]
- Kariithi, H.M.; Welch, C.N.; Ferreira, H.L.; Pusch, E.A.; Ateya, L.O.; Binepal, Y.S.; Apopo, A.A.; Dulu, T.D.; Afonso, C.L.; Suarez, D.L. Genetic characterization and pathogenesis of the first H9N2 low pathogenic avian influenza viruses isolated from chickens in Kenyan live bird markets. Infect. Genet. Evol. 2019, 78, 104074. [Google Scholar] [CrossRef]
- Iorio, R.M.; Borgman, J.B.; Glickman, R.L.; Bratt, M.A. Genetic variation within a neutralizing domain on the haemagglutinin—Neuraminidase glycoprotein of Newcastle disease virus. J. Gen. Virol. 1986, 67, 1393–1403. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef]
- Apopo, A.A.; Kariithi, H.M.; Ateya, L.O.; Binepal, Y.S.; Sirya, J.H.; Dulu, T.D.; Welch, C.N.; Hernandez, S.M.; Afonso, C.L. A retrospective study of Newcastle disease in Kenya. Trop. Anim. Health Prod. 2020, 52, 699–710. [Google Scholar] [CrossRef]
- Peeters, B.; Gruijthuijsen, Y.; De Leeuw, O.; Gielkens, A. Genome replication of Newcastle disease virus: Involvement of the rule-of-six. Arch. Virol. 2000, 145, 1829–1845. [Google Scholar] [CrossRef]
- Goraichuk, I.V.; Msoffe, P.L.; Chiwanga, G.H.; Dimitrov, K.M.; Afonso, C.L.; Suarez, D.L. First complete genome sequence of a subgenotype Vd Newcastle disease virus isolate. Microbiol. Resour. Announc. 2019, 8, e00436-19. [Google Scholar] [CrossRef] [PubMed]
- Iorio, R.M.; Glickman, R.L.; Riel, A.M.; Sheehan, J.P.; Bratt, M.A. Functional and neutralization profile of seven overlapping antigenic sites on the HN glycoprotein of Newcastle disease virus: Monoclonal antibodies to some sites prevent viral attachment. Virus Res. 1989, 13, 245–261. [Google Scholar] [CrossRef]
- Kim, S.-H.; Subbiah, M.; Samuel, A.S.; Collins, P.L.; Samal, S.K. Roles of the fusion and hemagglutinin-neuraminidase proteins in replication, tropism, and pathogenicity of avian paramyxoviruses. J. Virol. 2011, 85, 8582–8596. [Google Scholar] [CrossRef] [PubMed]
- Brown, V.R.; Bevins, S.N. A review of virulent Newcastle disease viruses in the United States and the role of wild birds in viral persistence and spread. Vet. Res. 2017, 48, 68. [Google Scholar] [CrossRef]
- Copland, J.W.; Alders, R.G. The Australian village poultry development programme in Asia and Africa. World Poultry Sci. J. 2005, 61, 31–38. [Google Scholar] [CrossRef][Green Version]
- Bensink, Z.; Spradbrow, P. Newcastle disease virus strain I2—A prospective thermostable vaccine for use in developing countries. Vet. Microbiol. 1999, 68, 131–139. [Google Scholar] [CrossRef]
- Wambura, P.N. Thermostable I-2 Strain of Newcastle Disease Virus as a Rural Vaccine. Ph.D. Thesis, School of Veterinary Science, The University of Queensland, Queensland, Australia, 2003. [Google Scholar]
- Miller, P.J.; Estevez, C.; Yu, Q.; Suarez, D.L.; King, D.J. Comparison of viral shedding following vaccination with inactivated and live Newcastle disease vaccines formulated with wild-type and recombinant viruses. Avian Dis. 2009, 53, 39–49. [Google Scholar] [CrossRef]
- Miller, P.J.; King, D.J.; Afonso, C.L.; Suarez, D.L. Antigenic differences among Newcastle disease virus strains of different genotypes used in vaccine formulation affect viral shedding after a virulent challenge. Vaccine 2007, 25, 7238–7246. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| County | Number of Sampling Locations | BPF Chickens | LBM Chickens | Other Domestic Poultry Species a | WFBs b | Total Birds Tested | PCR-Negative Samples | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total Tested | Putative NDV-Positive | Total Tested | Putative NDV-Positive | Total Tested | Putative NDV-Positive | Total Tested | Putative NDV-Positive | Total Tested | Putative NDV-Positive | |||
| Baringo | 13 | 176 | 129 (73.3%) | 43 | 41 (95.3%) | 7 | 7 (100%) | 43 | 13 (30.2%) | 269 | 190 (70.6%) | 79 (29.4%) |
| Bomet | 4 | 77 | 8 (10.4%) | 50 | 18 (36%) | N/A | N/A | N/A | N/A | 127 | 26 (20.5%) | 101 (79.5%) |
| Tana River | 6 | 89 | 3 (3.4%) | N/A | N/A | 23 | 14 (69.9%) | N/A | N/A | 112 | 17 (15.2%) | 95 (84.8%) |
| Kakamega | 8 | 169 | 61 (36.1%) | 48 | 35 (72.9%) | 32 | 1 (3.1%) | 53 | 3 (5.7%) | 302 | 100 (33.1%) | 202 (66.9%) |
| Kilifi | 20 | 165 | 39 (23.6%) | 38 | 9 (23.7%) | 37 | 5 (13.5%) | N/A | N/A | 240 | 53 (22.1%) | 187 (77.9%) |
| Machakos | 14 | 89 | 5 (5.6%) | 30 | 9 (30%) | 12 | 1 (8.3%) | 50 | 1 (2%) | 181 | 16 (8.8%) | 165 (91.2%) |
| Mombasa | 10 | 74 | 33 (44.6%) | 44 | 26 (59.1%) | N/A | N/A | N/A | N/A | 118 | 59 (50%) | 59 (50%) |
| Nairobi | 8 | N/A | N/A | 178 | 171 (96.1%) | N/A | N/A | N/A | N/A | 178 | 171 (96.1%) | 7 (3.9%) |
| Nakuru | 4 | 75 | 8 (10.7%) | N/A | N/A | N/A | N/A | 19 | 1 (5.3%) | 94 | 9 (9.6%) | 85 (90.4%) |
| Total | 87 | 914 | 286 (31.3%) | 431 | 309 (71.7%) | 111 | 28 (25.2%) | 165 | 18 (10.9%) | 1621 | 641 (39.5%) | 980 (60.5%) |
| Collection Date | Isolate | Sampling County (Location; Type) | Samples Used for NGS a | rRT-PCR (CT) | NGS Assembly Summaries | GenBank Accession No. | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| L-Test | F-Test | Mapped Reads b | Coverage Depth c | Consensus Sequence Length (Bases) | Genome Coverage | GC Content | |||||
| 5/23/2017 | 34MB09 | Bomet (Kapkwen; LBM) | CL (AF) | 28.44 | 23.87 | 306,471 | 0|2512|3381|4164|6149 | 15185 d | 99.9% | 47.7% | MW342790 |
| 3/13/2018 | 3FB129 | Kilifi (Mtwapa; BPF) | CL | 22.23 | 22.26 | 219,858 | 0|1971|2588|3126|4500 | 15,192 | 100.0% | 47.4% | MW342801 |
| 3/8/2018 | 3MB09 | Kilifi (MwembeKuku; LBM) | CL | 31.88 | 31.51 | 107,727 | 0|1037|1277|1477|2271 | 15,192 | 100.0% | 47.3% | MW342802 |
| 3/8/2018 | 3MB14 | Kilifi (MwembeKuku; LBM) | CL | 26.41 | 22.44 | 390,144 | 0|4201|4819|5329|6987 | 15,192 | 100.0% | 46.3% | MW342799 |
| 3/12/2018 | 3FB128 | Kilifi (Rabai; BPF) | CL | 22.19 | 21.83 | 663,414 | 0|6280|7810|9291|13482 | 15,192 | 100.0% | 46.9% | MW342788 |
| 3/9/2018 | 3FB26 | Kilifi (Sokoni; BPF) | CL | 22.03 | 17.77 | 286,241 | 0|2936|3569|4097|5549 | 15,192 | 100.0% | 47.2% | MW342784 |
| 3/9/2018 | 3FB28 | Kilifi (Sokoni; BPF) | OP | 21.81 | 17.26 | 311,051 | 0|2741|3744|4403|6051 | 15,192 | 99.9% | 46.9% | MW342780 |
| 3/9/2018 | 3FB29 | Kilifi (Sokoni; BPF) | CL | 24.17 | 19.73 | 369,615 | 0|3659|4273|4757|6271 | 15,192 | 99.9% | 46.9% | MW342787 |
| 3/9/2018 | 3FB31 | Kilifi (Sokoni; BPF) | OP (AF) | 19.78 | 20.22 | 553,799 | 2|2488|3999|6750|25170 | 15,192 | 100.0% | 47.1% | MW342782 |
| 3/9/2018 | 3FB32 | Kilifi (Sokoni; BPF) | CL | 20.96 | 16.67 | 281,654 | 0|3213|3745|4139|5548 | 15,192 | 100.0% | 46.7% | MW342785 |
| 3/9/2018 | 3FB33 | Kilifi (Sokoni; BPF) | CL | 25.88 | 21.48 | 248,816 | 0|2585|3226|3699|4980 | 15,192 | 100.0% | 47.0% | MW342783 |
| 3/9/2018 | 3FB34 | Kilifi (Sokoni; BPF) | CL | 23.93 | 19.53 | 287,161 | 0|2814|3418|3892|5546 | 15,192 | 100.0% | 47.1% | MW342786 |
| 3/9/2018 | 3FB35 | Kilifi (Sokoni; BPF) | OP | 27.29 | 21.48 | 118,132 | 0|1245|1422|1573|2224 | 15,192 | 100.0% | 47.1% | MW342781 |
| 3/8/2018 | 3FB15 | Kilifi BPF (Kilifi; BPF) | CL | 23.37 | 18.66 | 432,551 | 0|4610|5294|5953|7796 | 15,192 | 100.0% | 46.5% | MW342798 |
| 3/8/2018 | 3FB16 | Kilifi BPF (Kilifi; BPF) | CL | 27.92 | 23.39 | 676,403 | 0|7323|8205|9075|11885 | 15,192 | 100.0% | 46.5% | MW342797 |
| 3/15/2018 | 1MB23 | Mombasa (Changamwe; LBM) | CL (AF) | 32.7 | 28.18 | 933,391 | 1|9109|10694|12216|16029 | 15,192 | 100.0% | 46.6% | MW342794 |
| 3/16/2018 | 1MB35 | Mombasa (Changamwe; LBM) | CL (AF) | 29.12 | 24.31 | 455,182 | 0|4577|5491|6407|9405 | 15,185 e | 100.0% | 46.6% | MW342792 |
| 3/16/2018 | 1MB40 | Mombasa (Changamwe; LBM) | CL (AF) | 29.86 | 25.18 | 380,573 | 0|4080|4664|5252|6662 | 15,192 | 100.0% | 46.7% | MW342793 |
| 3/16/2018 | 1MB42 | Mombasa (Changamwe; LBM) | CL (AF) | 29.83 | 24.66 | 230,927 | 0|2241|2671|3023|4125 | 15,192 | 100.0% | 47.2% | MW342795 |
| 1/27/2017 | A104 | Nairobi (Burma; LBM) | CL (AF) | 32.96 | 33.2 | 516,967 | 0|5247|6245|7073|10094 | 15,192 | 100.0% | 46.6% | MW342805 |
| 1/27/2017 | A142 | Nairobi (Burma; LBM) | OP (AF) | 25.27 | 26.58 | 237,567 | 0|735|1110|2578|14486 | 15,192 f | 99.5% | 46.7% | MW342806 |
| 1/27/2017 | A144 | Nairobi (Burma; LBM) | OP (AF) | 33.67 | 31.2 | 395,414 | 0|1748|2460|4234|26640 | 15,192 | 100.0% | 46.6% | MW342791 |
| 1/27/2017 | A148 | Nairobi (Burma; LBM) | OP (AF) | 21.69 | 28.51 | 338,592 | 0|1368|2271|4082|23996 | 15,192 | 100.0% | 46.6% | MW342789 |
| 1/27/2017 | A101 | Nairobi (Burma; LBM) | OP (AF) | 27.43 | 30.39 | 348,356 | 0|706|1412|3459|26726 | 15,192 | 100.0% | 46.6% | MW342803 |
| 1/27/2017 | A105 | Nairobi (Burma; LBM) | OP (AF) | 28.95 | 33.45 | 307,666 | 1|1433|2306|3486|16181 | 15,192 | 100.0% | 46.1% | MW342804 |
| 1/27/2017 | A5136 | Nairobi (Kangemi; LBM) | CL (AF) | 32.56 | 35.95 | 497,720 | 0|1802|3003|6107|24022 | 15,192 | 100.0% | 46.2% | MW342807 |
| 1/31/2017 | A369 | Nairobi (Kariobangi North; LBM) | OP (AF) | 31.94 | 34.92 | 2,275 | 0|8|13|22|176 | 15,192 | 100.0% | 46.5% | MW342796 |
| 1/31/2017 | A371 | Nairobi (Kariobangi North; LBM) | CL | 32.91 | 28.14 | 369,484 | 0|3604|4275|4889|6842 | 15,192 | 100.0% | 46.5% | MW342777 |
| 1/31/2017 | A373 | Nairobi (Kariobangi North; LBM) | CL | 26.89 | 22.21 | 635,518 | 0|6139|7192|8176|11247 | 15,192 | 100.0% | 46.5% | MW342779 |
| 1/31/2017 | A374 | Nairobi (Kariobangi North; LBM) | CL | 25.47 | 20.62 | 616,616 | 0|5693|6984|8185|11887 | 15,192 | 100.0% | 46.3% | MW342778 |
| 1/31/2017 | A375 | Nairobi (Kariobangi North; LBM) | CL | 24.78 | 20.86 | 378,913 | 0|3629|4389|5030|7902 | 15,192 | 99.9% | 47.5% | MW342800 |
| 1/31/2017 | A376 | Nairobi (Kariobangi North; LBM) | OP | 27.68 | 22.6 | 311,605 | 0|2571|3408|4085|6106 | 15,192 | 100.0% | 47.5% | MW342776 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kariithi, H.M.; Ferreira, H.L.; Welch, C.N.; Ateya, L.O.; Apopo, A.A.; Zoller, R.; Volkening, J.D.; Williams-Coplin, D.; Parris, D.J.; Olivier, T.L.; et al. Surveillance and Genetic Characterization of Virulent Newcastle Disease Virus Subgenotype V.3 in Indigenous Chickens from Backyard Poultry Farms and Live Bird Markets in Kenya. Viruses 2021, 13, 103. https://doi.org/10.3390/v13010103
Kariithi HM, Ferreira HL, Welch CN, Ateya LO, Apopo AA, Zoller R, Volkening JD, Williams-Coplin D, Parris DJ, Olivier TL, et al. Surveillance and Genetic Characterization of Virulent Newcastle Disease Virus Subgenotype V.3 in Indigenous Chickens from Backyard Poultry Farms and Live Bird Markets in Kenya. Viruses. 2021; 13(1):103. https://doi.org/10.3390/v13010103
Chicago/Turabian StyleKariithi, Henry M., Helena L. Ferreira, Catharine N. Welch, Leonard O. Ateya, Auleria A. Apopo, Richard Zoller, Jeremy D. Volkening, Dawn Williams-Coplin, Darren J. Parris, Tim L. Olivier, and et al. 2021. "Surveillance and Genetic Characterization of Virulent Newcastle Disease Virus Subgenotype V.3 in Indigenous Chickens from Backyard Poultry Farms and Live Bird Markets in Kenya" Viruses 13, no. 1: 103. https://doi.org/10.3390/v13010103
APA StyleKariithi, H. M., Ferreira, H. L., Welch, C. N., Ateya, L. O., Apopo, A. A., Zoller, R., Volkening, J. D., Williams-Coplin, D., Parris, D. J., Olivier, T. L., Goldenberg, D., Binepal, Y. S., Hernandez, S. M., Afonso, C. L., & Suarez, D. L. (2021). Surveillance and Genetic Characterization of Virulent Newcastle Disease Virus Subgenotype V.3 in Indigenous Chickens from Backyard Poultry Farms and Live Bird Markets in Kenya. Viruses, 13(1), 103. https://doi.org/10.3390/v13010103