Screening for the Proteins That Can Interact with Grouper Nervous Necrosis Virus Capsid Protein


 ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Line and Cell Transfection
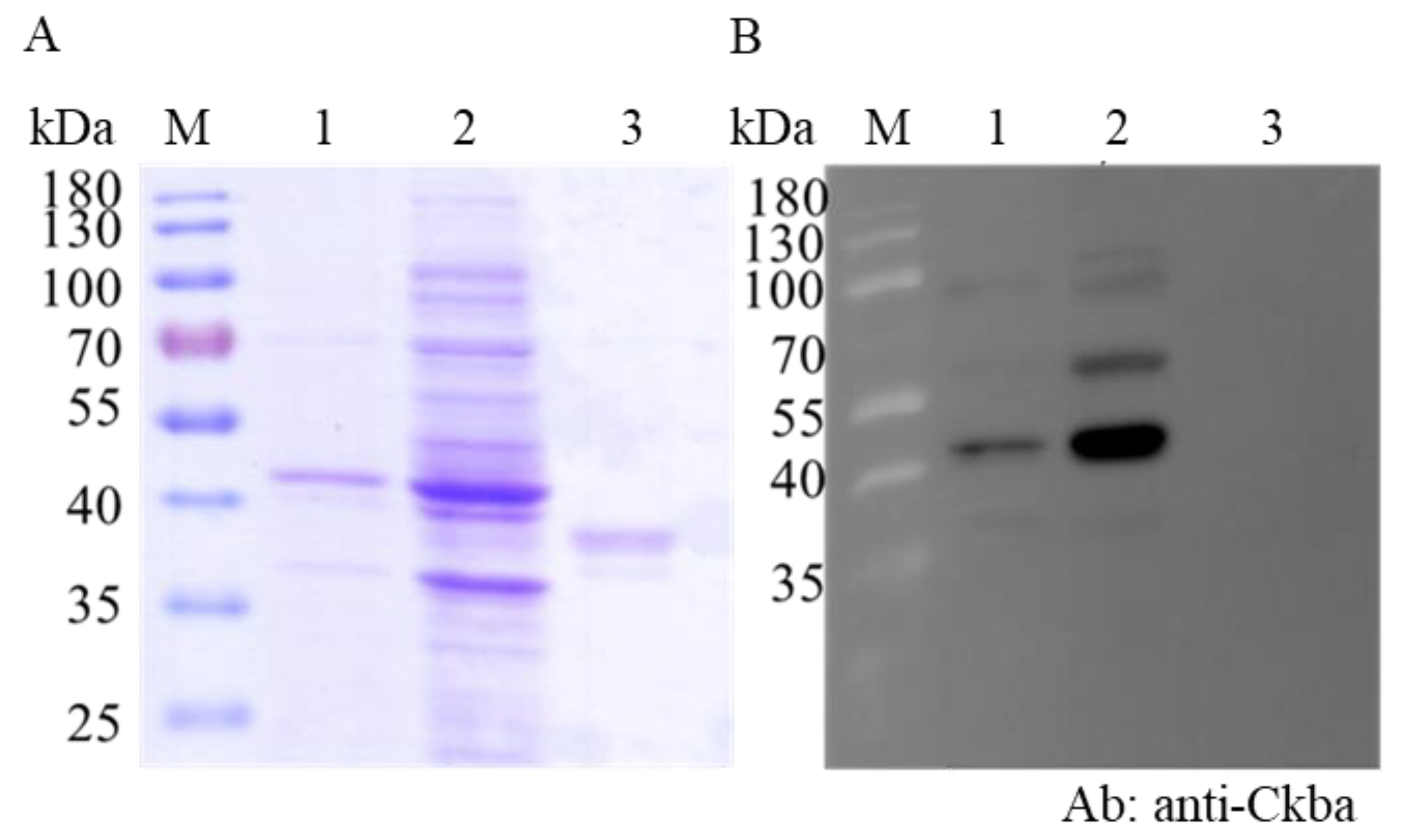
2.2. Expression and Purification of Recombinant NNVCP and CK B-type, and Antibody Preparation
2.3. Virus Infection
2.4. Grouper Tissue Lysates Preparation
2.5. SDS-PAGE
2.6. Western Blot Analyses
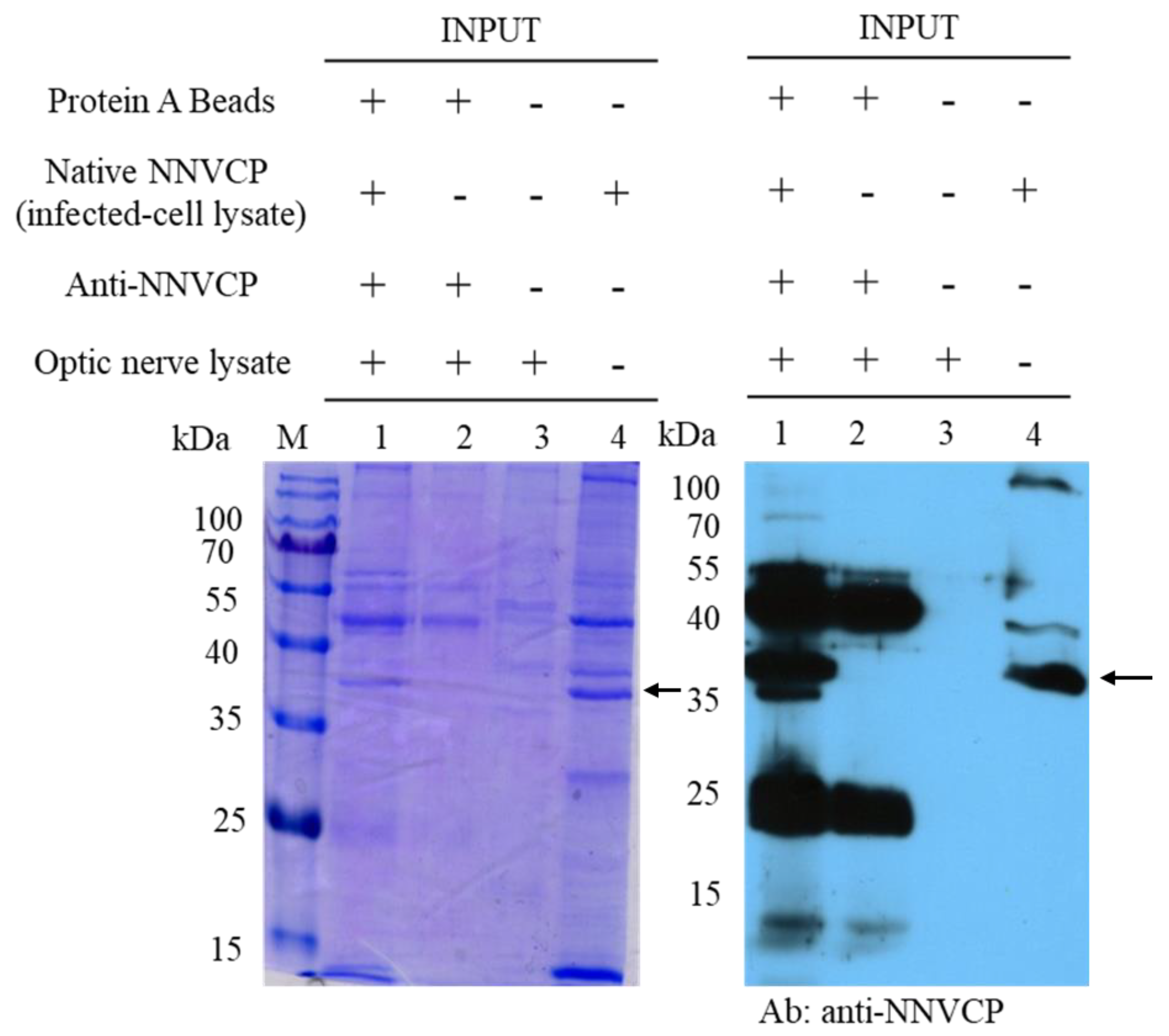
2.7. Immunoprecipitation (IP) Assay
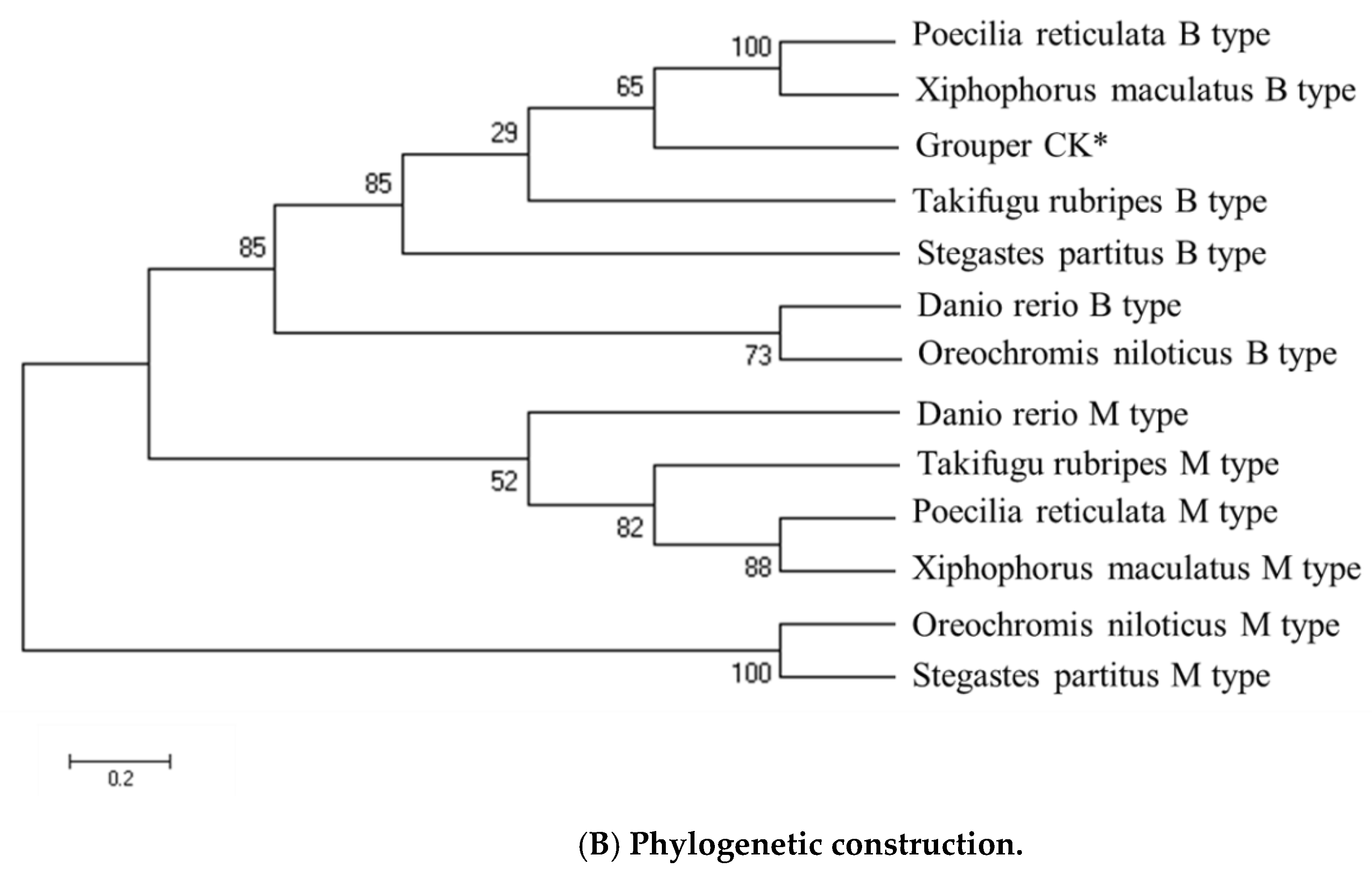
2.8. CK Amino Acid Sequence Analyses and Phylogenetic Construction
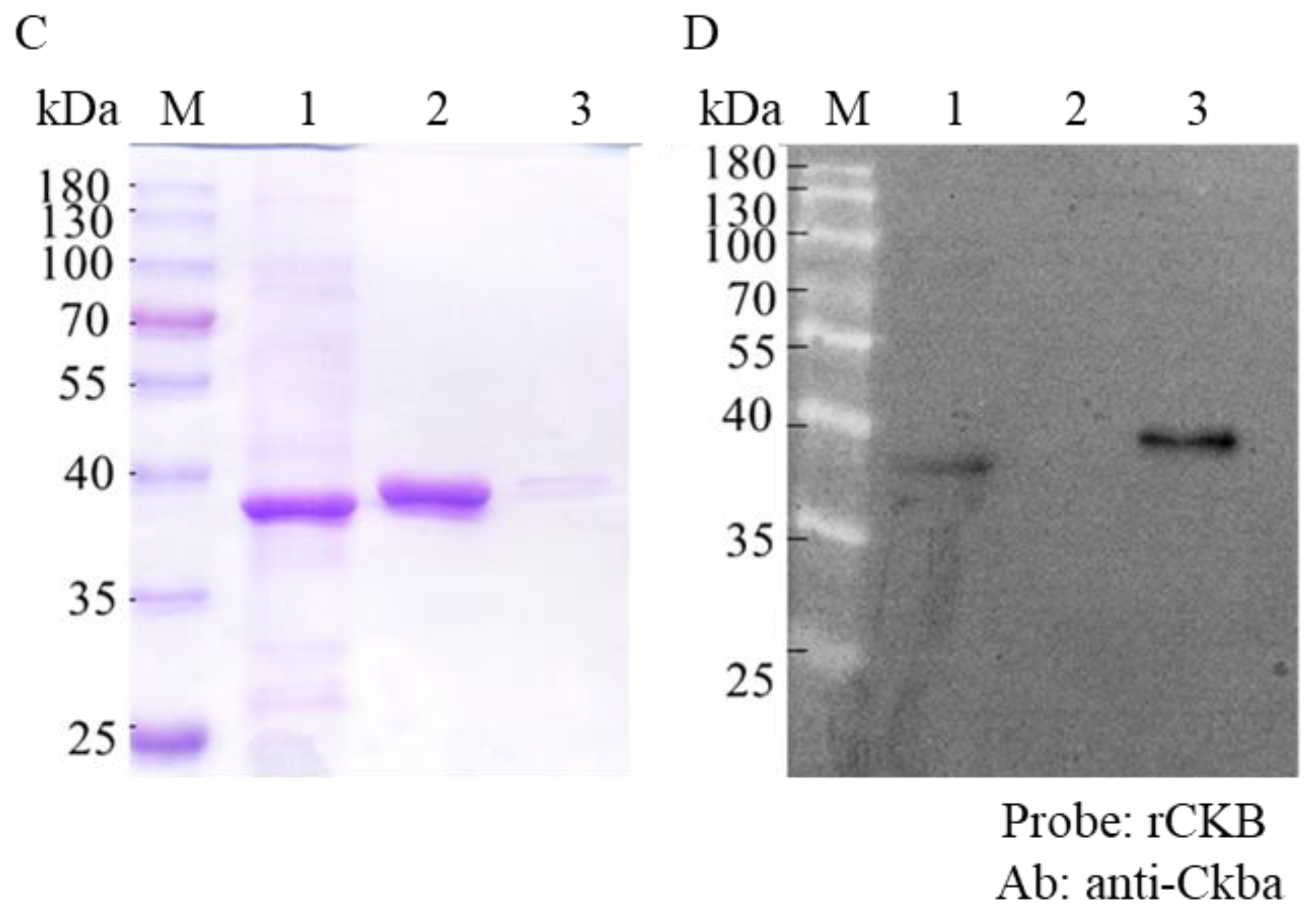
2.9. Far-Western Blot Assay
2.10. Cellular Colocalization of NNVCP and CKB by Indirect Immunofluorescence Assay
2.11. Cell Transfection
2.12. RNA Extraction and qRT-PCR Analysis
3. Results
3.1. Identification of Optic Nerve Proteins Interacting with Native NNV Capsid Protein (NNVCP) by Proteomic Analysis of Immunoprecipitation (IP) Assay
3.2. Sequence Analysis of Grouper Creatine Kinase (CK) Protein and Phylogenetic Construction
3.3. CKB Interacted with NNVCP In Vitro
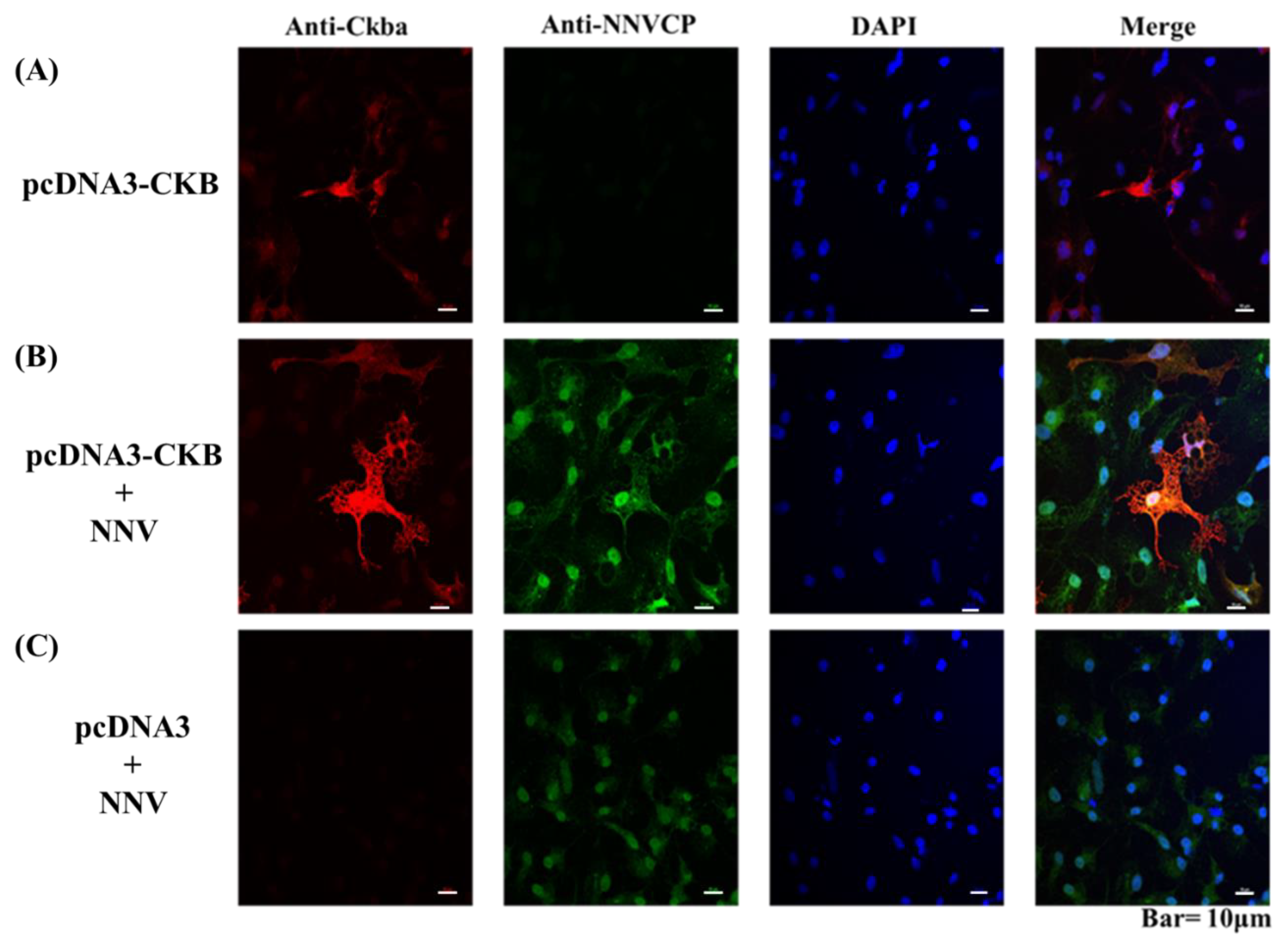
3.4. CKB Colocalized with NNVCP in the GF-1 Cell Line
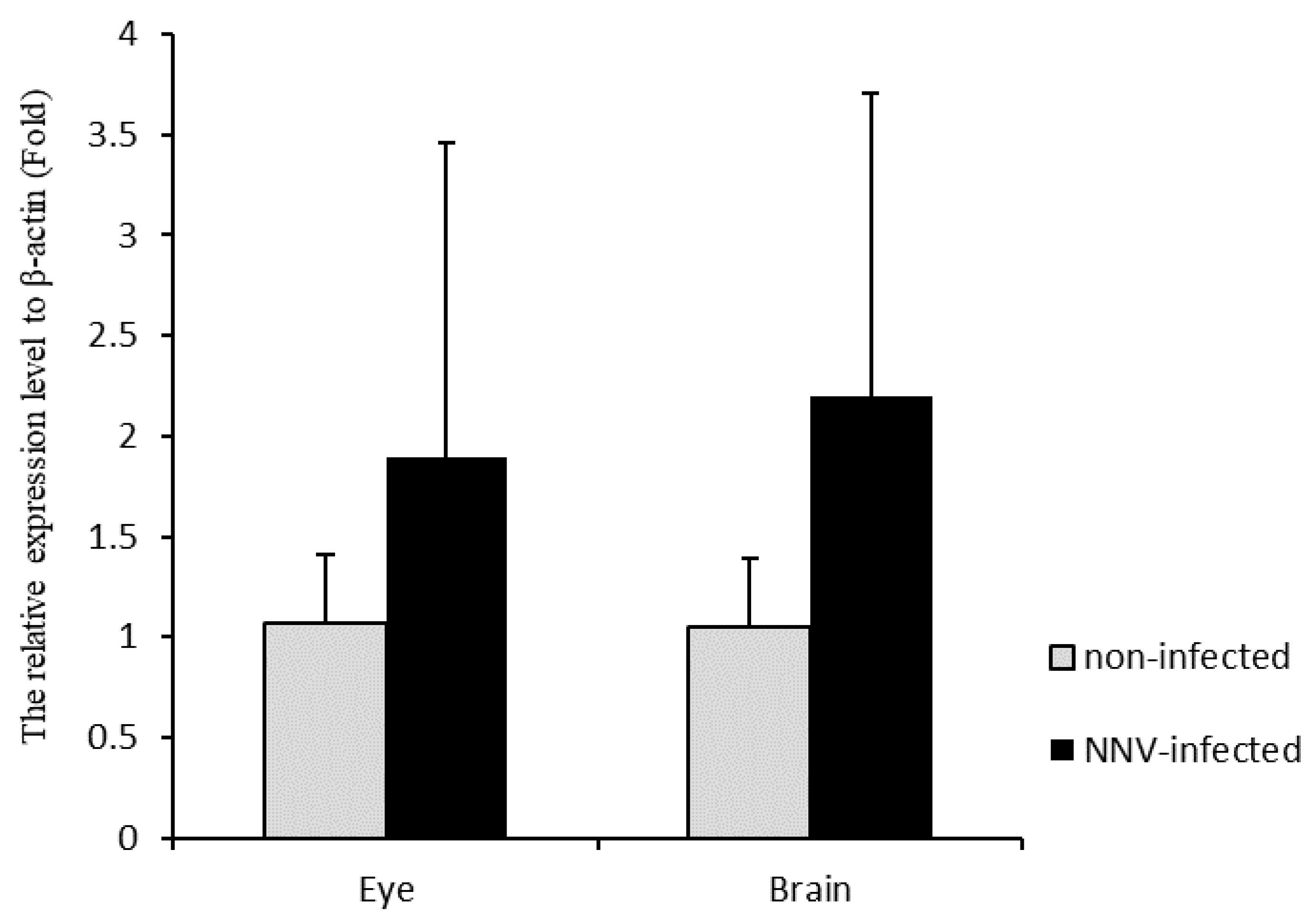
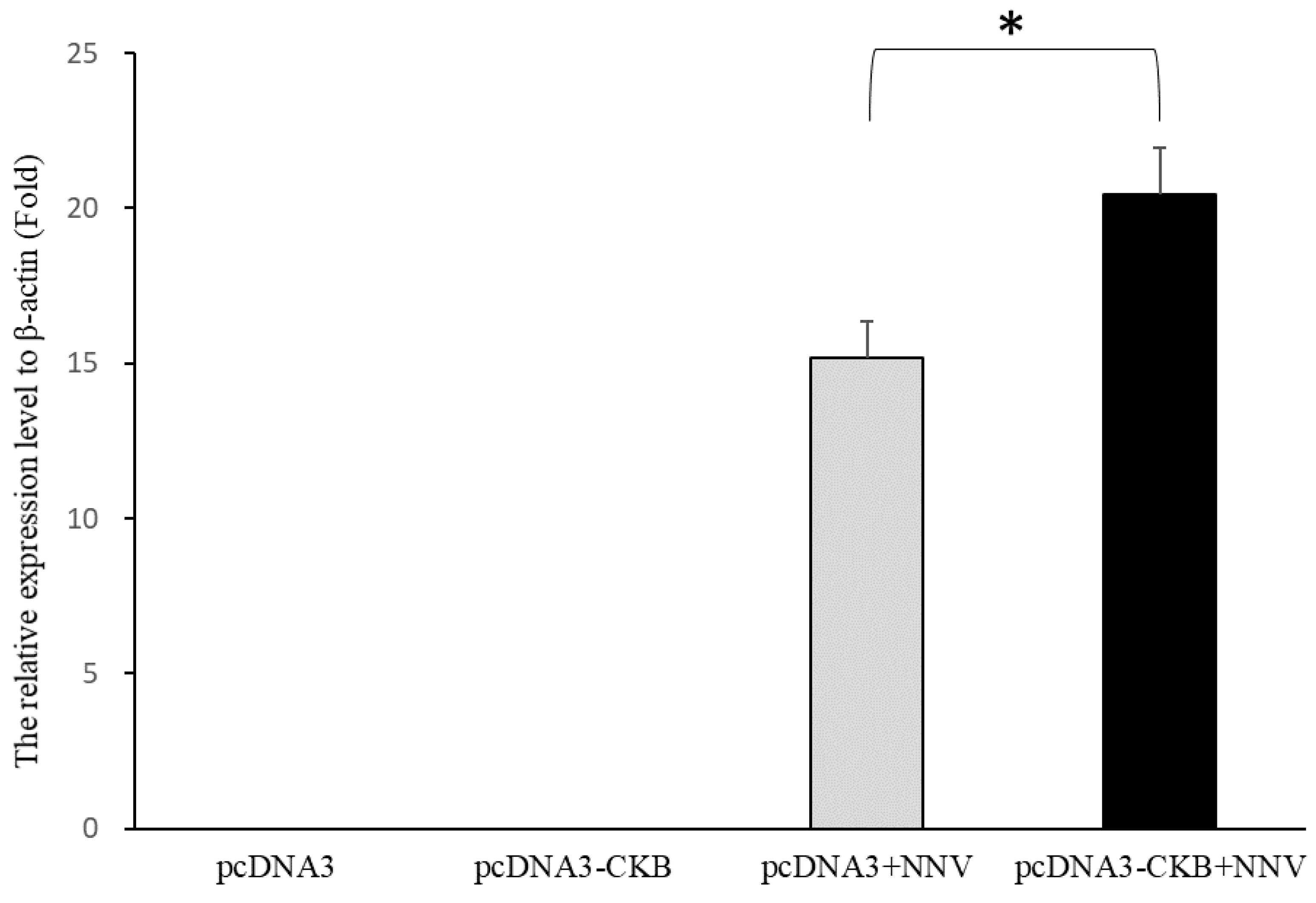
3.5. CKB Participates in NNV Morphogenesis
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hegde, A.; Chen, C.; Qin, Q.; Lam, T.; Sin, Y. Characterization, pathogenicity and neutralization studies of a nervous necrosis virus isolated from grouper, Epinephelus tauvina, in Singapore. Aquaculture 2002, 213, 55–72. [Google Scholar] [CrossRef]
- Qin, Q.; Chang, S.; Ngoh-Lim, G.; Gibson-Kueh, S.; Shi, C.; Lam, T. Characterization of a novel ranavirus isolated from grouper Epinephelus tauvina. Dis. Aquat. Org. 2003, 53, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Huang, Y.; Zhu, W.; Li, C.; Huang, X.; Qin, Q. Isolation and identification of Singapore grouper iridovirus Hainan strain (SGIV-HN) in China. Arch. Virol. 2019, 164, 1869–1872. [Google Scholar] [CrossRef] [PubMed]
- Bandín, I.; Souto, S. Betanodavirus and VER disease: A 30-year research review. Pathogens 2020, 9, 106. [Google Scholar] [CrossRef]
- Arimoto, M.; Maruyama, K.; Furusawa, I. Epizootiology of viral nervous necrosis (VNN) in striped jack. Fish Pathol. 1994, 29, 19–24. [Google Scholar] [CrossRef][Green Version]
- Munday, B.; Kwang, J.; Moody, N. Betanodavirus infections of teleost fish: A review. J. Fish Dis. 2002, 25, 127–142. [Google Scholar] [CrossRef]
- Kara, H.; Chaoui, L.; Derbal, F.; Zaidi, R.; De Boisséson, C.; Baud, M.; Bigarré, L. Betanodavirus-associated mortalities of adult wild groupers E pinephelus marginatus (Lowe) and E pinephelus costae (Steindachner) in A lgeria. J. Fish Dis. 2014, 37, 273–278. [Google Scholar] [CrossRef]
- Moody, N.; Horwood, P.; Reynolds, A.; Mahony, T.; Anderson, I.; Oakey, H. Phylogenetic analysis of betanodavirus isolates from Australian finfish. Dis. Aquat. Org. 2009, 87, 151–160. [Google Scholar] [CrossRef]
- Vendramin, N.; Patarnello, P.; Toffan, A.; Panzarin, V.; Cappellozza, E.; Tedesco, P.; Terlizzi, A.; Terregino, C.; Cattoli, G. Viral Encephalopathy and Retinopathy in groupers (Epinephelus spp.) in southern Italy: A threat for wild endangered species? BMC Vet. Res. 2013, 9, 1–7. [Google Scholar] [CrossRef]
- Costa, J.Z.; Thompson, K.D. Understanding the interaction between Betanodavirus and its host for the development of prophylactic measures for viral encephalopathy and retinopathy. Fish Shellfish Immunol. 2016, 53, 35–49. [Google Scholar] [CrossRef]
- Chi, S.; Lo, C.; Kou, G.; Chang, P.; Peng, S.; Chen, S. Mass mortalities associated with viral nervous necrosis (VNN) disease in two species of hatchery-reared grouper, Epinephelus fuscogutatus and Epinephelus akaara (Temminck & Schlegel). J. Fish Dis. 1997, 20, 185–193. [Google Scholar]
- Mori, K.-I.; Nakai, T.; Muroga, K.; Arimoto, M.; Mushiake, K.; Furusawa, I. Properties of a new virus belonging to Nodaviridae found in larval striped jack (Pseudocaranx dentex) with nervous necrosis. Virology 1992, 187, 368–371. [Google Scholar] [CrossRef]
- Chen, L.-J.; Su, Y.-C.; Hong, J.-R. Betanodavirus non-structural protein B1: A novel anti-necrotic death factor that modulates cell death in early replication cycle in fish cells. Virology 2009, 385, 444–454. [Google Scholar] [CrossRef]
- Fenner, B.J.; Thiagarajan, R.; Chua, H.K.; Kwang, J. Betanodavirus B2 is an RNA interference antagonist that facilitates intracellular viral RNA accumulation. J. Virol. 2006, 80, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, T.; Mise, K.; Takeda, A.; Okinaka, Y.; Mori, K.-I.; Arimoto, M.; Okuno, T.; Nakai, T. Characterization of Striped jack nervous necrosis virus subgenomic RNA3 and biological activities of its encoded protein B2. J. Gen. Virol. 2005, 86, 2807–2816. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Teng, Y.; Zheng, X.; Wu, Y.; Xie, X.; He, J.; Ye, Y.; Wu, Z. Complete sequence of a viral nervous necrosis virus (NNV) isolated from red-spotted grouper (Epinephelus akaara) in China. Arch. Virol. 2012, 157, 777–782. [Google Scholar] [CrossRef]
- Iwamoto, T.; Okinaka, Y.; Mise, K.; Mori, K.-I.; Arimoto, M.; Okuno, T.; Nakai, T. Identification of host-specificity determinants in betanodaviruses by using reassortants between striped jack nervous necrosis virus and sevenband grouper nervous necrosis virus. J. Virol. 2004, 78, 1256–1262. [Google Scholar] [CrossRef]
- Nishizawa, T.; Furuhashi, M.; Nagai, T.; Nakai, T.; Muroga, K. Genomic classification of fish nodaviruses by molecular phylogenetic analysis of the coat protein gene. Appl. Environ. Microbiol. 1997, 63, 1633–1636. [Google Scholar] [CrossRef]
- Kuan, Y.C.; Sheu, F.; Lee, G.C.; Tsai, M.W.; Hung, C.L.; Nan, F.H. Administration of recombinant Reishi immunomodulatory protein (rLZ-8) diet enhances innate immune responses and elicits protection against nervous necrosis virus in grouper Epinephelus coioides. Fish Shellfish Immunol. 2012, 32, 986–993. [Google Scholar] [CrossRef]
- Wang, Y.D.; Rajanbabu, V.; Chen, J.Y. Transcriptome analysis of medaka following epinecidin-1 and TH1-5 treatment of NNV infection. Fish Shellfish Immunol. 2015, 42, 121–131. [Google Scholar] [CrossRef]
- Kuo, H.P.; Chung, C.L.; Hung, Y.F.; Lai, Y.S.; Chiou, P.P.; Lu, M.W.; Kong, Z.L. Comparison of the responses of different recombinant fish type I interferons against betanodavirus infection in grouper. Fish Shellfish Immunol. 2016, 49, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Lomonossoff, G.P.; Marsian, J.; Hurdiss, D.L.; Ranson, N.A.; Ritala, A.; Paley, R.K.; Cano Cejas, I. Plant-made nervous necrosis virus-like particles protect fish against disease. Front. Plant Sci. 2019, 10, 880. [Google Scholar]
- Chang, J.S.; Chi, S.C. GHSC70 is involved in the cellular entry of nervous necrosis virus. J. Virol. 2015, 89, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Jia, K.; Jia, P.; Xiang, Y.; Lu, X.; Liu, W.; Yi, M. Marine medaka heat shock protein 90ab1 is a receptor for red-spotted grouper nervous necrosis virus and promotes virus internalization through clathrin-mediated endocytosis. PLoS Pathog. 2020, 16, e1008668. [Google Scholar] [CrossRef]
- Chang, J.S.; Chi, S.C. Grouper voltage-dependent anion selective channel protein 2 is required for nervous necrosis virus infection. Fish Shellfish Immunol. 2015, 46, 315–322. [Google Scholar] [CrossRef]
- Liu, H.; Sadygov, R.G.; Yates, J.R., 3rd. A model for random sampling and estimation of relative protein abundance in shotgun proteomics. Anal. Chem. 2004, 76, 4193–4201. [Google Scholar] [CrossRef]
- Chen, K.Y.; Hsu, T.C.; Huang, P.Y.; Kang, S.T.; Lo, C.F.; Huang, W.P.; Chen, L.L. Penaeus monodon chitin-binding protein (PmCBP) is involved in white spot syndrome virus (WSSV) infection. Fish Shellfish Immunol. 2009, 27, 460–465. [Google Scholar] [CrossRef]
- Manos, P.; Edmond, J. Immunofluorescent analysis of creatine kinase in cultured astrocytes by conventional and confocal microscopy: A nuclear localization. J. Comp. Neurol. 1992, 326, 273–282. [Google Scholar] [CrossRef]
- Valero, Y.; Arizcun, M.; Esteban, M.Á.; Cuesta, A.; Chaves-Pozo, E. Transcription of histones H1 and H2B is regulated by several immune stimuli in gilthead seabream and European sea bass. Fish Shellfish Immunol. 2016, 57, 107–115. [Google Scholar] [CrossRef]
- Wu, Y.; Chi, S. Cloning and analysis of antiviral activity of a barramundi (Lates calcarifer) Mx gene. Fish Shellfish Immunol. 2007, 23, 97–108. [Google Scholar] [CrossRef]
- Zou, J.; Secombes, C.J. Teleost fish interferons and their role in immunity. Dev. Comp. Immunol. 2011, 35, 1376–1387. [Google Scholar] [CrossRef] [PubMed]
- Leong, J.A.C.; Trobridge, G.D.; Kim, C.H.; Johnson, M.; Simon, B. Interferon-inducible Mx proteins in fish. Immunol. Rev. 1998, 166, 349–363. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-M.; Su, Y.-L.; Shie, P.-S.; Huang, S.-L.; Yang, H.-L.; Chen, T.-Y. Grouper Mx confers resistance to nodavirus and interacts with coat protein. Dev. Comp. Immunol. 2008, 32, 825–836. [Google Scholar] [CrossRef]
- Guerrero, C.A.; Bouyssounade, D.; Zárate, S.; Iša, P.; López, T.; Espinosa, R.; Romero, P.; Méndez, E.; López, S.; Arias, C.F. Heat shock cognate protein 70 is involved in rotavirus cell entry. J. Virol. 2002, 76, 4096–4102. [Google Scholar] [CrossRef]
- Saphire, A.C.; Guan, T.; Schirmer, E.C.; Nemerow, G.R.; Gerace, L. Nuclear import of adenovirus DNA in vitro involves the nuclear protein import pathway and hsc70. J. Biol. Chem. 2000, 275, 4298–4304. [Google Scholar] [CrossRef] [PubMed]
- Agostini, I.; Popov, S.; Li, J.; Dubrovsky, L.; Hao, T.; Bukrinsky, M. Heat-shock protein 70 can replace viral protein R of HIV-1 during nuclear import of the viral preintegration complex. Exp. Cell Res. 2000, 259, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Seeger, C. Hsp90 is required for the activity of a hepatitis B virus reverse transcriptase. Proc. Natl. Acad. Sci. USA 1996, 93, 1060–1064. [Google Scholar] [CrossRef]
- Stahl, M.; Retzlaff, M.; Nassal, M.; Beck, J. Chaperone activation of the hepadnaviral reverse transcriptase for template RNA binding is established by the Hsp70 and stimulated by the Hsp90 system. Nucleic Acids Res. 2007, 35, 6124–6136. [Google Scholar] [CrossRef]
- Park, S.G.; Jung, G. Human hepatitis B virus polymerase interacts with the molecular chaperonin Hsp60. J. Virol. 2001, 75, 6962–6968. [Google Scholar] [CrossRef]
- Cripe, T.P.; Delos, S.E.; Estes, P.A.; Garcea, R.L. In vivo and in vitro association of hsc70 with polyomavirus capsid proteins. J. Virol. 1995, 69, 7807–7813. [Google Scholar] [CrossRef]
- Crouchet, E.; Baumert, T.F.; Schuster, C. Hepatitis C virus-apolipoprotein interactions: Molecular mechanisms and clinical impact. Expert REV Proteomic 2017, 14, 593–606. [Google Scholar] [CrossRef] [PubMed]
- Ku, N.O.; Zhou, X.; Toivola, D.M.; Omary, M.B. The cytoskeleton of digestive epithelia in health and disease. Am. J. Physiol. 1999, 277, G1108–G1137. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, E.; Weber, K. Intermediate filaments: Structure, dynamics, function, and disease. Annu. Rev. Biochem. 1994, 63, 345–382. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Q.; An, X.; Yang, Y.X.; Hu, H.D.; Ren, H.; Hu, P. Keratin 8 is involved in hepatitis B virus replication. J. Med. Virol. 2014, 86, 687–694. [Google Scholar] [CrossRef]
- Bar, H.; Fischer, D.; Goudeau, B.; Kley, R.A.; Clemen, C.S.; Vicart, P.; Herrmann, H.; Vorgerd, M.; Schroder, R. Pathogenic effects of a novel heterozygous R350P desmin mutation on the assembly of desmin intermediate filaments in vivo and in vitro. Hum. Mol. Genet. 2005, 14, 1251–1260. [Google Scholar] [CrossRef]
- Vassar, R.; Coulombe, P.A.; Degenstein, L.; Albers, K.; Fuchs, E. Mutant keratin expression in transgenic mice causes marked abnormalities resembling a human genetic skin disease. Cell 1991, 64, 365–380. [Google Scholar] [CrossRef]
- Richens, J.L.; Spencer, H.L.; Butler, M.; Cantlay, F.; Vere, K.A.; Bajaj, N.; Morgan, K.; O’Shea, P. Rationalising the role of Keratin 9 as a biomarker for Alzheimer’s disease. Sci. Rep. 2016, 6, 22962. [Google Scholar] [CrossRef]
- Neilson, D.E.; Adams, M.D.; Orr, C.M.; Schelling, D.K.; Eiben, R.M.; Kerr, D.S.; Anderson, J.; Bassuk, A.G.; Bye, A.M.; Childs, A.-M. Infection-triggered familial or recurrent cases of acute necrotizing encephalopathy caused by mutations in a component of the nuclear pore, RANBP2. Am. J. Hum. Genet. 2009, 84, 44–51. [Google Scholar] [CrossRef]
- Woo, J.; Jung, A.; Lakshmana, M.; Bedrossian, A.; Lim, Y.; Bu, J.; Park, S.; Koo, E.; Mook-Jung, I.; Kang, D. Pivotal role of the RanBP9-cofilin pathway in Aβ-induced apoptosis and neurodegeneration. Cell Death Differ. 2012, 19, 1413. [Google Scholar] [CrossRef]
- Liu, T.; Roh, S.; Woo, J.; Ryu, H.; Kang, D. Cooperative role of RanBP9 and P73 in mitochondria-mediated apoptosis. Cell Death Dis. 2013, 4, e476. [Google Scholar] [CrossRef]
- Grünblatt, E.; Riederer, P. Aldehyde dehydrogenase (ALDH) in Alzheimer’s and Parkinson’s disease. J. Neural. Transm. 2016, 123, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Dong, H.; Li, S.; Munir, M.; Chen, J.; Luo, Y.; Sun, Y.; Liu, L.; Qiu, H.J. Hemoglobin subunit beta interacts with the capsid protein and antagonizes the growth of classical swine fever virus. J. Virol. 2013, 87, 5707–5717. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, D.; Zangerle, R.; Artner-Dworzak, E.; Weiss, G.; Fritsch, P.; Tilz, G.P.; Dierich, M.P.; Wachter, H. Association between immune activation, changes of iron metabolism and anaemia in patients with HIV infection. Eur. J. Haematol. 1993, 50, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, T.; Takano, R.; Muroga, K. Mapping a neutralizing epitope on the coat protein of striped jack nervous necrosis virus. J. Gen. Virol. 1999, 80 Pt 11, 3023–3027. [Google Scholar] [CrossRef]
- Bravo, R.; Parra, V.; Gatica, D.; Rodriguez, A.E.; Torrealba, N.; Paredes, F.; Wang, Z.V.; Zorzano, A.; Hill, J.A.; Jaimovich, E.; et al. Endoplasmic reticulum and the unfolded protein response: Dynamics and metabolic integration. Int. Rev. Cell Mol. Biol. 2013, 301, 215–290. [Google Scholar] [CrossRef]
- Wu, Y.M.; Hsu, C.H.; Wang, C.H.; Liu, W.; Chang, W.H.; Lin, C.S. Role of the DxxDxD motif in the assembly and stability of betanodavirus particles. Arch. Virol. 2008, 153, 1633–1642. [Google Scholar] [CrossRef]
- Chin, D.; Means, A.R. Calmodulin: A prototypical calcium sensor. Trends Cell Biol. 2000, 10, 322–328. [Google Scholar] [CrossRef]
- Yamniuk, A.P.; Vogel, H.J. Calmodulin’s flexibility allows for promiscuity in its interactions with target proteins and peptides. Mol. Biotechnol. 2004, 27, 33–57. [Google Scholar] [CrossRef]
- Dreux, M.; Chisari, F.V. Viruses and the autophagy machinery. Cell Cycle Georget. Tex. 2010, 9, 1295–1307. [Google Scholar] [CrossRef]
- Vlach, J.; Samal, A.B.; Saad, J.S. Solution structure of calmodulin bound to the binding domain of the HIV-1 matrix protein. J. Biol. Chem. 2014, 289, 8697–8705. [Google Scholar] [CrossRef]
- Jin, Y.; Zhang, R.; Wu, W.; Duan, G. Antiviral and Inflammatory Cellular Signaling Associated with Enterovirus 71 Infection. Viruses 2018, 10, 155. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, M.J.; Sherer, N.M.; Marks, C.B.; Pypaert, M.; Mothes, W. Actin- and myosin-driven movement of viruses along filopodia precedes their entry into cells. J. Cell Biol. 2005, 170, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, H.; Nakamura, M.; Ohno, T.; Matsuda, Y.; Yuda, Y.; Nonomura, Y. Myosin-actin interaction plays an important role in human immunodeficiency virus type 1 release from host cells. Proc. Natl. Acad. Sci. USA 1995, 92, 2026–2030. [Google Scholar] [CrossRef] [PubMed]
- Ye, T.; Zong, R.; Zhang, X. Involvement of interaction between viral VP466 and host tropomyosin proteins in virus infection in shrimp. Gene 2012, 505, 254–258. [Google Scholar] [CrossRef]
- Dolder, M.; Wendt, S.; Wallimann, T. Mitochondrial creatine kinase in contact sites: Interaction with porin and adenine nucleotide translocase, role in permeability transition and sensitivity to oxidative damage. Biol. Signals Recept. 2001, 10, 93–111. [Google Scholar] [CrossRef]
- Wallimann, T.; Wyss, M.; Brdiczka, D.; Nicolay, K.; Eppenberger, H. Intracellular compartmentation, structure and function of creatine kinase isoenzymes in tissues with high and fluctuating energy demands: The ‘phosphocreatine circuit’for cellular energy homeostasis. Biochem. J. 1992, 281, 21–40. [Google Scholar] [CrossRef]
- Megarajan, S.; Ranjan, R.; Xavier, B.; Ghosh, S.; Sadu, N.; Bathina, C.; Gopalakrishnan, A. Molecular detection of betanodavirus in orange-spotted grouper (Epinephelus coioides) broodstock maintained in recirculating aquaculture systems and sea cages. Aquac. Int. 2020, 349–362. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Functional Classification and Description | |
|---|---|
| Ribosomal protein | |
|
|
|
|
|
|
|
|
|
|
|
|
| Immunity | |
|
|
|
|
|
|
| Glucose metabolisms and ATP generation | |
|
|
|
|
|
|
| |
| Cytoskeleton proteins | |
|
|
|
|
|
|
| |
| Ca2+ binding protein | |
|
|
|
|
| |
| Ion regulation protein | |
|
|
| Lipid metabolism proteins | |
|
|
| |
| Apoptosis protein | |
|
|
| Protein hydrolysis | |
|
|
| Other | |
|
|
| GenBank Accession No. | Description | Score * |
|---|---|---|
| AAR97600.2 | beta actin (Epinephelus coioides) | 2492.00 |
| 3JBM | C Chain C, Virus-like Particle of Orange-spotted Grouper Nervous Necrosis Virus | 1706.91 |
| AER42656.1 | keratin 8, partial (Epinephelus coioides) | 1457.49 |
| AAX78203.1 | immunoglobulin mu heavy chain (Epinephelus coioides) | 1263.15 |
| AAX78206.1 | immunoglobulin mu heavy chain (Epinephelus coioides) | 999.30 |
| AGG55392.1 | voltage-dependent anion selective channel protein 2 (Epinephelus coioides) | 944.17 |
| AAX78208.1 | immunoglobulin mu heavy chain (Epinephelus coioides) | 914.30 |
| AEG78351.1 | keratin type II E3, partial (Epinephelus coioides) | 790.40 |
| ACM48181.1 | apolipoprotein AI, partial (Epinephelus coioides) | 745.80 |
| AGG55391.1 | heat shock cognate protein 70 (Epinephelus coioides) | 745.79 |
| ACH73075.1 | keratin 8, partial (Epinephelus coioides) | 740.04 |
| AIS72878.1 | heat shock protein 60 (Epinephelus coioides) | 679.74 |
| AEW43726.1 | transferrin (Epinephelus coioides) | 645.54 |
| AER42652.1 | hemoglobin beta chain (Epinephelus coioides) | 623.08 |
| ADG29138.1 | tropomyosin (Epinephelus coioides) | 539.59 |
| ABW74631.1 | immunoglobulin light chain (Epinephelus coioides) | 504.86 |
| AHB51756.1 | calmodulin (Epinephelus coioides) | 491.05 |
| ABW04131.1 | glyceraldehyde-3-phosphate dehydrogenase, partial (Epinephelus coioides) | 484.23 |
| AAS55942.1 | immunoglobulin light chain variable region (Epinephelus coioides) | 476.38 |
| ABW04145.1 | smooth muscle cell-specific protein SM22 alpha (Epinephelus coioides) | 430.16 |
| AEO89322.1 | voltage dependent anion channel protein 1 (Epinephelus coioides) | 421.60 |
| AER42688.1 | tropomyosin alpha-4 chain, partial (Epinephelus coioides) | 418.42 |
| ABW04135.1 | natural killer cell enhancement factor (Epinephelus coioides) | 361.05 |
| AEG78406.1 | hemoglobin alpha chain, partial (Epinephelus coioides) | 347.41 |
| ADG29126.1 | fructose-bisphosphate aldolase A (Epinephelus coioides) | 340.73 |
| AEG78409.1 | 60S ribosomal protein L30 (Epinephelus coioides) | 325.27 |
| ADG29180.1 | triosephosphate isomerase B (Epinephelus coioides) | 310.40 |
| ACL98136.1 | type I keratin, partial (Epinephelus coioides) | 305.82 |
| ABW04143.1 | S100-like calcium binding protein (Epinephelus coioides) | 298.82 |
| ABW04124.1 | ADP-ATP translocase (Epinephelus coioides) | 288.91 |
| ACH73065.1 | ribosomal protein S7 (Epinephelus coioides) | 283.43 |
| ADZ76534.1 | myosin light chain 3 (Epinephelus coioides) | 279.24 |
| ABW04132.1 | Hnrpa01 protein, partial (Epinephelus coioides) | 269.70 |
| ACV04938.1 | heat shock protein 90 (Epinephelus coioides) | 269.27 |
| ADG29156.1 | histone H2B (Epinephelus coioides) | 265.26 |
| AOW69105.1 | elongation factor 1-alpha (Epinephelus coioides) | 264.68 |
| ADG29136.1 | beta-enolase (Epinephelus coioides) | 247.11 |
| AEG78428.1 | 60S ribosomal protein L10a (Epinephelus coioides) | 246.47 |
| AAW29021.1 | lactate dehydrogenase-A, partial (Epinephelus coioides) | 212.87 |
| ABW04139.1 | ribosomal protein LP0 (Epinephelus coioides) | 212.25 |
| ABW04136.1 | nucleoside-diphosphate kinase (Epinephelus coioides) | 199.15 |
| ADG29150.1 | 60S ribosomal protein L19, partial (Epinephelus coioides) | 186.15 |
| ADG29176.1 | muscle phosphoglycerate mutase 2 (Epinephelus coioides) | 184.01 |
| ACH73061.1 | ribosomal protein L7, partial (Epinephelus coioides) | 157.36 |
| ABW04127.1 | brain-type fatty acid binding protein (Epinephelus coioides) | 156.32 |
| ABW04123.1 | 60S ribosomal protein L27 (Epinephelus coioides) | 152.44 |
| ADZ99127.1 | immunoglobulin heavy chain variable region, partial (Epinephelus coioides) | 146.61 |
| AEG78395.1 | aldehyde dehydrogenase family 9 member A1-A, partial (Epinephelus coioides) | 136.08 |
| ADG29178.1 | tryptase-2 precursor, partial (Epinephelus coioides) | 135.49 |
| ACL98142.1 | desmin, partial (Epinephelus coioides) | 120.60 |
| ABW04138.1 | ribosomal protein L23, partial (Epinephelus coioides) | 110.93 |
| ACH73060.1 | ictacalcin (Epinephelus coioides) | 104.23 |
| ABW04130.1 | galactoside binding lectin (Epinephelus coioides) | 100.05 |
| AER42692.1 | 60S ribosomal protein LP1 (Epinephelus coioides) | 99.41 |
| AEG78402.1 | 40S ribosomal protein S18 (Epinephelus coioides) | 87.32 |
| ADG29169.1 | 60S ribosomal protein L13, partial (Epinephelus coioides) | 86.49 |
| ADG29143.1 | 60S ribosomal protein L31 (Epinephelus coioides) | 67.76 |
| ACM41841.1 | creatine kinase, partial (Epinephelus coioides) | 58.09 |
| AEG78365.1 | 40S ribosomal protein S2 (Epinephelus coioides) | 57.41 |
| AEG78426.1 | eukaryotic translation elongation factor 2, partial (Epinephelus coioides) | 55.83 |
| ABW74647.1 | immunoglobulin light chain (Epinephelus coioides) | 51.54 |
| ACM41842.1 | 14 kDa apolipoprotein, partial (Epinephelus coioides) | 50.87 |
| ACL98134.1 | ran protein, partial (Epinephelus coioides) | 48.98 |
| AHA43788.1 | calreticulin (Epinephelus coioides) | 38.35 |
| ABW04141.1 | ribosomal protein S13, partial (Epinephelus coioides) | 32.62 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, P.-Y.; Hsiao, H.-C.; Wang, S.-W.; Lo, S.-F.; Lu, M.-W.; Chen, L.-L. Screening for the Proteins That Can Interact with Grouper Nervous Necrosis Virus Capsid Protein. Viruses 2020, 12, 985. https://doi.org/10.3390/v12090985
Huang P-Y, Hsiao H-C, Wang S-W, Lo S-F, Lu M-W, Chen L-L. Screening for the Proteins That Can Interact with Grouper Nervous Necrosis Virus Capsid Protein. Viruses. 2020; 12(9):985. https://doi.org/10.3390/v12090985
Chicago/Turabian StyleHuang, Po-Yu, Han-Chia Hsiao, Szu-Wen Wang, Shao-Fu Lo, Ming-Wei Lu, and Li-Li Chen. 2020. "Screening for the Proteins That Can Interact with Grouper Nervous Necrosis Virus Capsid Protein" Viruses 12, no. 9: 985. https://doi.org/10.3390/v12090985
APA StyleHuang, P.-Y., Hsiao, H.-C., Wang, S.-W., Lo, S.-F., Lu, M.-W., & Chen, L.-L. (2020). Screening for the Proteins That Can Interact with Grouper Nervous Necrosis Virus Capsid Protein. Viruses, 12(9), 985. https://doi.org/10.3390/v12090985