Progress in the Development of Universal Influenza Vaccines


Abstract
1. Introduction
2. Development of Universal Vaccines Targeting Influenza Virus Surface Proteins
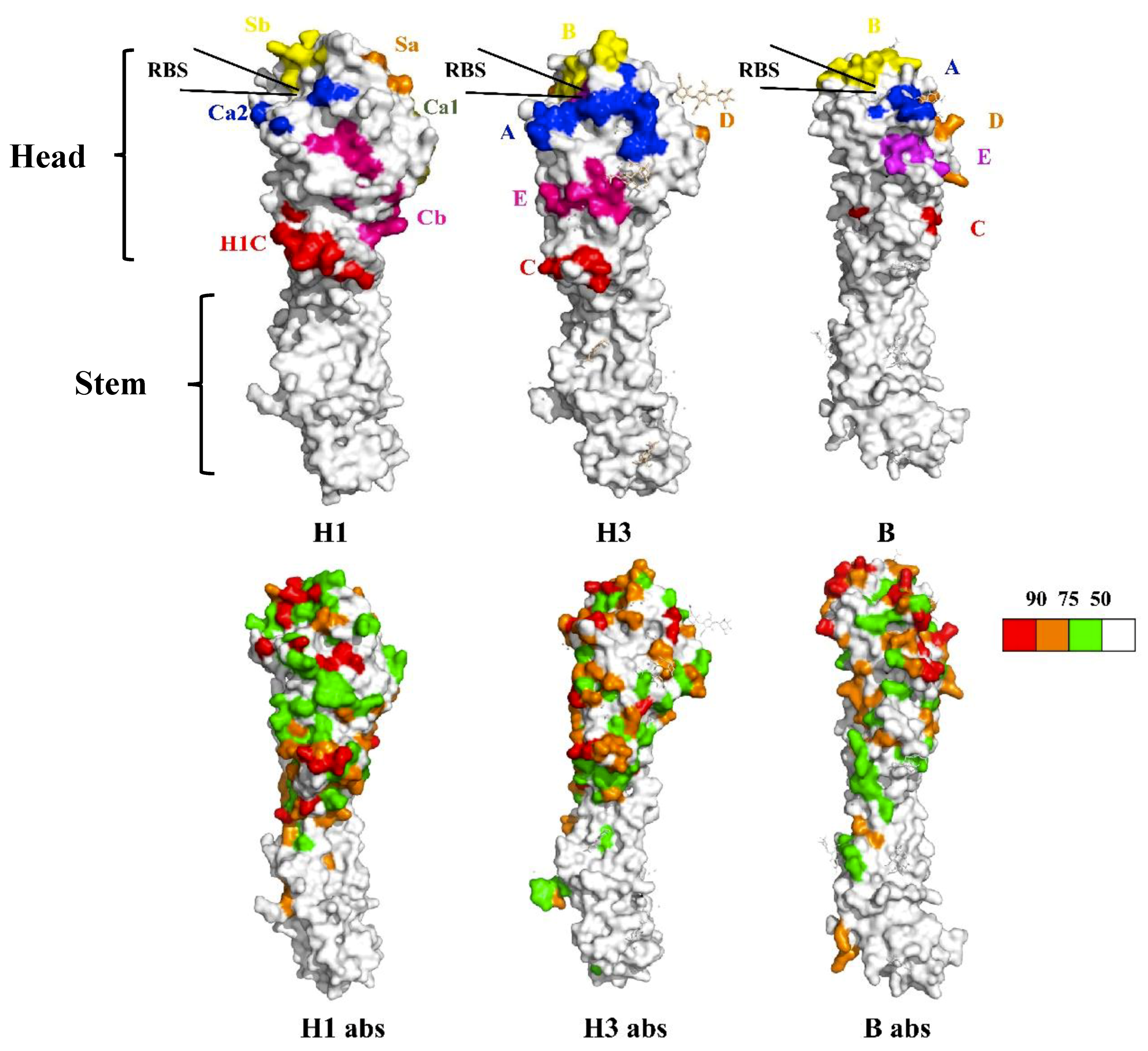
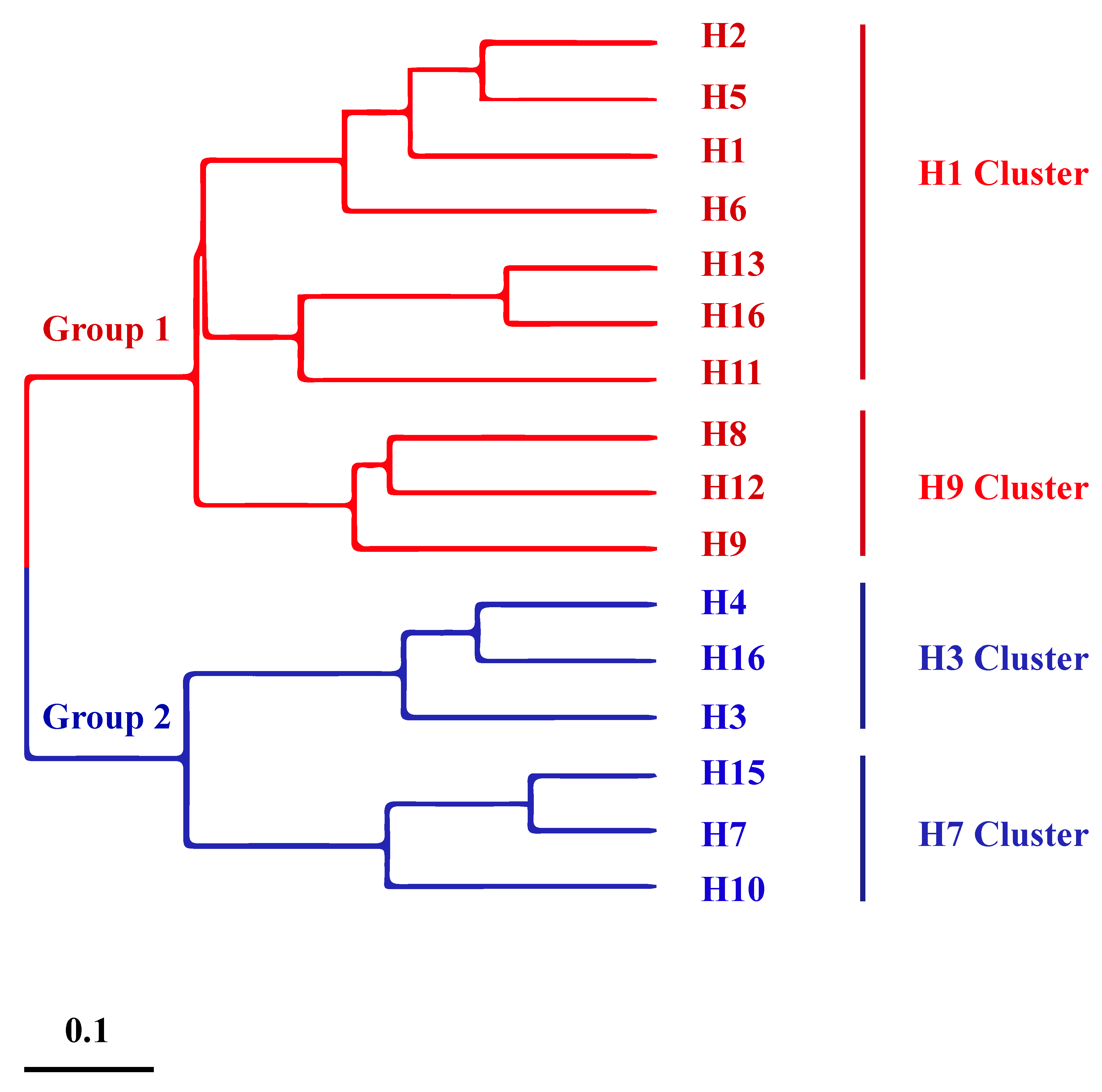
2.1. Universal Vaccines Targeting the Stem Region of HA
2.2. Chimeric HA Universal Vaccines
2.3. NA Universal Vaccines
3. Universal Influenza Vaccines Targeting Other Structural Proteins of Influenza Viruses
4. Novel Universal Influenza Vaccines
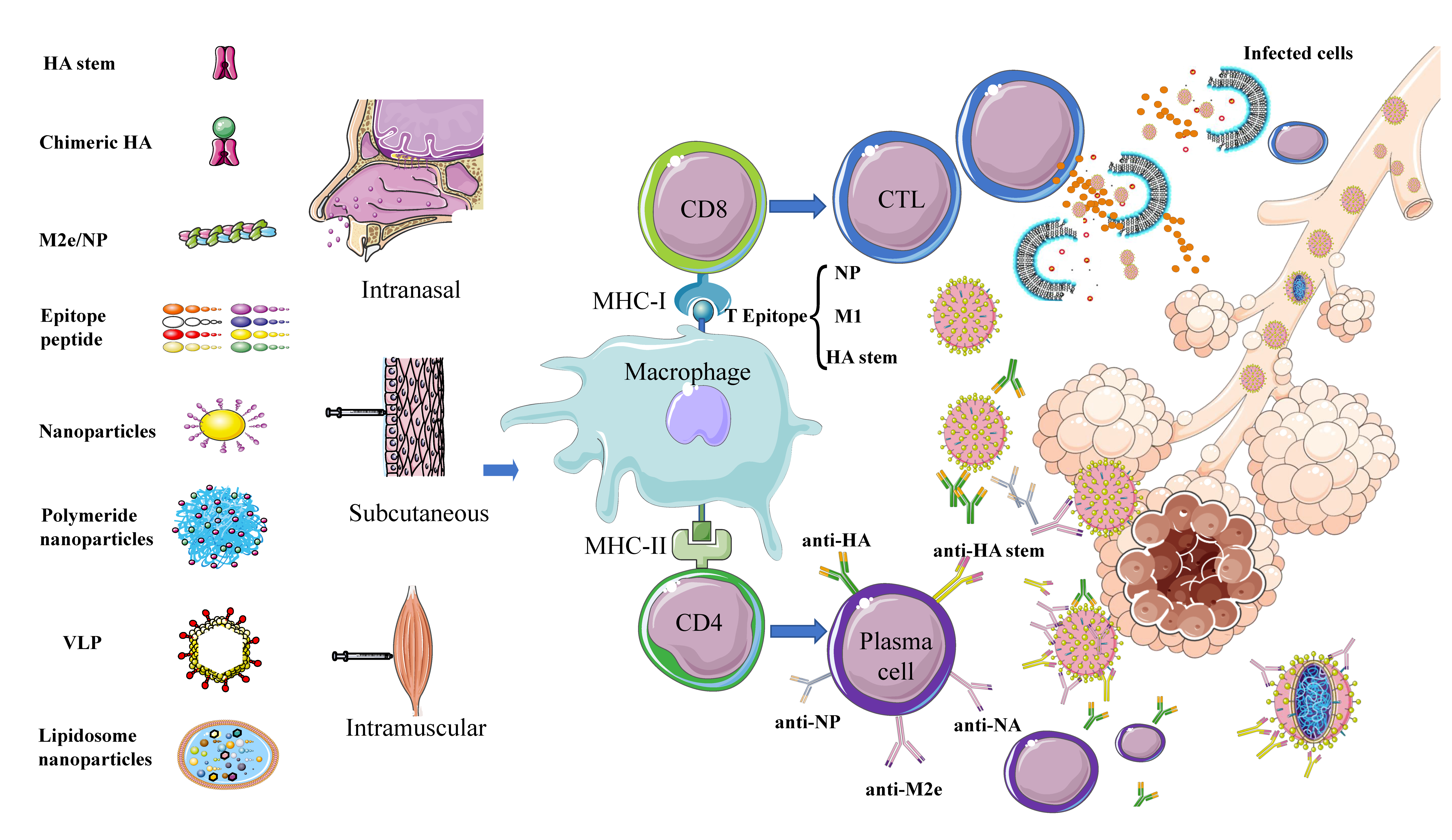
4.1. Universal Influenza Vaccines Targeting T/B Cell Epitopes
4.2. Universal Mosaic Influenza Vaccines
4.3. Nanoparticle Universal Influenza Vaccines
5. Adjuvants
6. Perspective
Author Contributions
Funding
Conflicts of Interest
References
- Iv, M.L.; Dunagan, M.; Kurebayashi, Y.; Takimoto, T. Key role of the influenza a virus pa gene segment in the emergence of pandemic viruses. Viruses 2020, 12, 365. [Google Scholar] [CrossRef]
- Sun, Y.; Hu, Z.; Zhang, X.; Chen, M.; Wang, Z.; Xu, G.; Bi, Y.; Tong, Q.; Wang, M.; Sun, H.; et al. R195K mutation in the PA-X protein increases the virulence and transmission of influenza A virus in mammalian hosts. J. Virol. 2020, 94. [Google Scholar] [CrossRef] [PubMed]
- Cheung, P.; Ye, Z.; Lee, T.; Chen, H.; Chan, C.; Jin, D. PB1-F2 protein of highly pathogenic influenza A (H7N9) virus selectively suppresses RNA-induced NLRP3 inflammasome activation through inhibition of MAVS-NLRP3 interaction. J. Leukoc. Biol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, R.; Li, Q.; Wang, F.; Zhu, B.; Zheng, M.; Cui, H.; Wen, J.; Zhao, G. Host cell interactome of PB1 N40 protein of H5N1 influenza A virus in chicken cells. J. Proteom. 2019, 197, 34–41. [Google Scholar] [CrossRef]
- Vasin, A.; Temkina, O.; Egorov, V.; Klotchenko, S.; Plotnikova, M.; Kiselev, O. Molecular mechanisms enhancing the proteome of influenza a viruses: An overview of recently discovered proteins. Virus Res. 2014, 185, 53–63. [Google Scholar] [CrossRef]
- Klein, N.; Fireman, B.; Goddard, K.; Zerbo, O.; Asher, J.; Zhou, J.; King, J.; Lewis, N. Vaccine effectiveness of cell-culture relative to egg-based inactivated influenza vaccine during the 2017-18 influenza season. PLoS ONE 2020, 15, e0229279. [Google Scholar] [CrossRef]
- Sarsenbayeva, G.; Issagulov, T.; Kassenov, M.; Abitay, R.; Orynbayev, M.; Stukova, M.; Pisareva, M.; Davlyatshin, T.; Lespek, K.; Khairullin, B. Safety and immunogenicity of trivalent inactivated influenza vaccine in adults 60 years of age and older: A phase II, a randomized, comparative trial in Kazakhstan. Hum. Vaccines Immunother. 2020. [Google Scholar] [CrossRef]
- Avalos, L.; Ferber, J.; Zerbo, O.; Naleway, A.; Bulkley, J.; Thompson, M.; Cragan, J.; Williams, J.; Odouli, R.; Kauffman, T.; et al. Trivalent inactivated influenza vaccine (IIV3) during pregnancy and six-month infant development. Vaccine 2020, 38, 2326–2332. [Google Scholar] [CrossRef]
- Trombetta, C.M.; Gianchecchi, E.; Montomoli, E. Influenza vaccines: Evaluation of the safety profile. Hum. Vaccin Immunother. 2018, 14, 657–670. [Google Scholar] [CrossRef]
- Lee, G.; Chu, K.; Inn, K.; Moon, E.; Quan, F. Vaccine Efficacy Induced by 2009 Pandemic H1N1 Virus-Like Particles Differs from that Induced by Split Influenza Virus. Immunol. Investig. 2019. [Google Scholar] [CrossRef]
- Kim, K.; Lee, Y.; Park, S.; Jung, Y.; Lee, Y.; Ko, E.; Kim, Y.; Li, X.; Kang, S. Neuraminidase expressing virus-like particle vaccine provides effective cross protection against influenza virus. Virology 2019, 535, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Subbarao, K. Live Attenuated Cold-Adapted Influenza Vaccines. Cold Spring Harbor Perspect. Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- Holzer, B.; Morgan, S.; Martini, V.; Sharma, R.; Clark, B.; Chiu, C.; Salguero, F.; Tchilia, N.E. Immunogenicity and protective efficacy of seasonal human live attenuated cold-adapted influenza virus vaccine in pigs. Front. Immunol. 2019, 10, 2625. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, L.; Blanco-Lobo, P.; Reilly, E.; Maehigashi, T.; Nogales, A.; Smith, A.; Topham, D.; Dewhurst, S.; Kim, B.; Martínez-Sobrido, L. Comparative study of the temperature sensitive, cold adapted and attenuated mutations present in the master donor viruses of the two commercial human live attenuated influenza vaccines. Viruses 2019, 11, 928. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.; Kim, J.; Byun, Y.; Son, A.; Lee, J.; Lee, Y.; Chang, J.; Seong, B. Pan-influenza a protection by prime-boost vaccination with cold-adapted live-attenuated influenza vaccine in a mouse model. Front. Immunol. 2018, 9, 116. [Google Scholar] [CrossRef]
- Li, H.; Li, Q.; Li, B.; Guo, Y.; Xing, J.; Xu, Q.; Liu, L.; Zhang, J.; Qi, W.; Jia, W.; et al. Continuous reassortment of clade 2.3.4.4 H5N6 Highly pathogenetic avian influenza viruses demonstrating high risk to public health. Pathogens 2020, 9, 670. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, C.; Zhang, H.; Liu, G.D.; Xue, C.; Cao, Y. Targeting hemagglutinin: Approaches for broad protection against the influenza a virus. Viruses 2019, 11, 405. [Google Scholar] [CrossRef]
- Saha, P.; Biswas, M.; Gupta, R.; Majumdar, A.; Mitra, S.; Banerjee, A.; Mukherjee, A.; Dutta, S.; Chawla-Sarkar, M. Molecular characterization of Influenza A pandemic H1N1 viruses circulating in eastern India during 2017–2019: Antigenic diversity in comparison to the vaccine strains. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2020, 81, 104270. [Google Scholar] [CrossRef]
- Xue, K.; Bloom, J. Linking influenza virus evolution within and between human hosts. Virus Evol. 2020, 6, veaa010. [Google Scholar] [CrossRef]
- Wille, M.; Holmes, E. The ecology and evolution of influenza viruses. Cold Spring Harb. Perspect. Med. 2019. [Google Scholar] [CrossRef]
- Zolotarova, O.; Budzanivska, I.; Leibenko, L.; Radchenko, L.; Mironenko, A. Antigenic site variation in the hemagglutinin of pandemic influenza A(H1N1)pdm09 viruses between 2009–2017 in Ukraine. Pathogens 2019, 8, 194. [Google Scholar] [CrossRef] [PubMed]
- Stray, S.; Pittman, L. Subtype- and antigenic site-specific differences in biophysical influences on evolution of influenza virus hemagglutinin. Virol. J. 2012, 9, 91. [Google Scholar] [CrossRef] [PubMed]
- Chromikova, V.; Tan, J.; Aslam, S.; Rajabhathor, A.; Bermudez-Gonzalez, M.; Ayllon, J.; Simon, V.; García-Sastre, A.; Salaun, B.; Nachbagauer, R.; et al. Activity of human serum antibodies in an influenza virus hemagglutinin stalk-based ADCC reporter assay correlates with activity in a CD107a degranulation assay. Vaccine 2020, 38. [Google Scholar] [CrossRef]
- Krammer, F.; Palese, P. Universal influenza virus vaccines that target the conserved hemagglutinin stalk and conserved sites in the head domain. J. Infect. Dis. 2019, 219, S62–S67. [Google Scholar] [CrossRef]
- Steel, J.; Lowen, A.C.; Wang, T.T.; Yondola, M.; Gao, Q.; Haye, K.; Garcia-Sastre, A.; Palese, P. Influenza virus vaccine based on the conserved hemagglutinin stalk domain. mBio 2010, 1. [Google Scholar] [CrossRef]
- Wohlbold, T.; Nachbagauer, R.; Margine, I.; Tan, G.; Hirsh, A.; Krammer, F. Vaccination with soluble headless hemagglutinin protects mice from challenge with divergent influenza viruses. Vaccine 2015, 33, 3314–3321. [Google Scholar] [CrossRef]
- Yassine, H.; Boyington, J.; McTamney, P.; Wei, C.; Kanekiyo, M.; Kon, G.W.; Gallagher, J.; Wang, L.; Zhang, Y.; Joyce, M.; et al. Hemagglutinin-stem nanoparticles generate heterosubtypic influenza protection. Nat. Med. 2015, 21, 1065–1070. [Google Scholar] [CrossRef]
- Impagliazzo, A.; Milder, F.; Kuipers, H.; Wagner, M.; Zhu, X.; Hoffman, R.; van Meersbergen, R.; Huizingh, J.; Wanningen, P.; Verspui, J.J.; et al. A stable trimeric influenza hemagglutinin stem as a broadly protective immunogen. Science 2015, 349, 1301–1306. [Google Scholar] [CrossRef]
- Golchin, M.; Moghadaszadeh, M.; Tavakkoli, H.; Ghanbarpour, R.; Dastmalchi, S. Recombinant M2e-HA2 fusion protein induced immunity responses against intranasally administered H9N2 influenza virus. Microb. Pathog. 2018, 115, 183–188. [Google Scholar] [CrossRef]
- Trucchi, C.; Paganino, C.; Amicizia, D.; Orsi, A.; Tisa, V.; Piazza, M.; Icardi, G.; Ansaldi, F. Universal influenza virus vaccines: What needs to happen next? Expert Opin. Biol. Ther. 2019, 19, 671–683. [Google Scholar] [CrossRef]
- Estrada, L.D.; Schultz-Cherry, S. Development of a universal influenza vaccine. J. Immunol. 2019, 202, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Boyoglu-Barnum, S.; Hutchinson, G.B.; Boyington, J.C.; Moin, S.M.; Gillespie, R.A.; Tsybovsky, Y.; Stephens, T.; Vaile, J.R.; Lederhofer, J.; Corbett, K.S.; et al. Glycan repositioning of influenza hemagglutinin stem facilitates the elicitation of protective cross-group antibody responses. Nat. Commun. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Bajic, G.; Maron, M.; Adachi, Y.; Onodera, T.; McCarthy, K.; McGee, C.; Sempowski, G.; Takahashi, Y.; Kelsoe, G.; Kuraoka, M.; et al. Influenza antigen engineering focuses immune responses to a subdominant but broadly protective viral Epitope. Cell Host Microbe 2019, 25, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Broecker, F.; Liu, S.; Suntronwong, N.; Sun, W.; Bailey, M.; Nachbagauer, R.; Krammer, F.; Palese, P. A mosaic hemagglutinin-based influenza virus vaccine candidate protects mice from challenge with divergent H3N2 strains. NPJ Vaccines 2019, 4, 31. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, D.; Guptill, J.; Naficy, A.; Nachbagauer, R.; Berlanda-Scorza, F.; Feser, J.; Wilson, P.; Solórzano, A.; van der Wielen, M.; Walter, E.; et al. Immunogenicity of chimeric haemagglutinin-based, universal influenza virus vaccine candidates: Interim results of a randomised, placebo-controlled, phase 1 clinical trial. Lancet Infect. Dis. 2020, 20, 80–91. [Google Scholar] [CrossRef]
- Asthagiri Arunkumar, G.; McMahon, M.; Pavot, V.; Aramouni, M.; Ioannou, A.; Lambe, T.; Gilbert, S.; Krammer, F. Vaccination with viral vectors expressing NP, M1 and chimeric hemagglutinin induces broad protection against influenza virus challenge in mice. Vaccine 2019, 37, 5567–5577. [Google Scholar] [CrossRef]
- Liu, W.; Nachbagauer, R.; Stadlbauer, D.; Solórzano, A.; Berlanda-Scorza, F.; García-Sastre, A.; Palese, P.; Krammer, F.; Albrecht, R. Sequential immunization with live-attenuated chimeric hemagglutinin-based vaccines confers heterosubtypic immunity against influenza a viruses in a preclinical ferret model. Front. Immunol. 2019, 10, 756. [Google Scholar] [CrossRef]
- McMahon, M.; Asthagiri Arunkumar, G.; Liu, W.; Stadlbauer, D.; Albrecht, R.; Pavot, V.; Aramouni, M.; Lambe, T.; Gilbert, S.; Krammer, F. Vaccination with viral vectors expressing chimeric hemagglutinin, NP and M1 antigens protects ferrets against influenza virus challenge. Front. Immunol. 2019, 10, 2005. [Google Scholar] [CrossRef]
- Sun, W.; Zheng, A.; Miller, R.; Krammer, F.; Palese, P. An inactivated influenza virus vaccine approach to targeting the conserved hemagglutinin stalk and M2e domains. Vaccines 2019, 7, 117. [Google Scholar] [CrossRef]
- Gubareva, L.; Mohan, T. Antivirals Targeting the Neuraminidase. Cold Spring Harb. Perspect. Med. 2020. [Google Scholar] [CrossRef]
- Abed, Y.; Schibler, M.; Checkmahomed, L.; Carbonneau, J.; Venable, M.; Fage, C.; Giannotti, F.; Goncalves, A.; Kaiser, L.; Boivin, G. Molecular pathway of influenza pan-neuraminidase inhibitor resistance in an immunocompromised patient. Antivir. Ther. 2020. [Google Scholar] [CrossRef] [PubMed]
- Lampejo, T. Influenza and antiviral resistance: An overview. Eur. J. Clin. Microbiol. Infect. Dis. 2020. [Google Scholar] [CrossRef] [PubMed]
- Wohlbold, T.; Nachbagauer, R.; Xu, H.; Tan, G.; Hirsh, A.; Brokstad, K.; Cox, R.; Palese, P.; Krammer, F. Vaccination with adjuvanted recombinant neuraminidase induces broad heterologous, but not heterosubtypic, cross-protection against influenza virus infection in mice. mBio 2015, 6, e02556. [Google Scholar] [CrossRef]
- Stadlbauer, D.; Zhu, X.; McMahon, M.; Turner, J.; Wohlbold, T.; Schmitz, A.; Strohmeier, S.; Yu, W.; Nachbagauer, R.; Mudd, P.; et al. Broadly protective human antibodies that target the active site of influenza virus neuraminidase. Science 2019, 366, 499–504. [Google Scholar] [CrossRef]
- Piepenbrink, M.; Nogales, A.; Basu, M.; Fucile, C.; Liesveld, J.; Keefer, M.; Rosenberg, A.; Martinez-Sobrido, L.; Kobie, J. Broad and protective influenza B virus neuraminidase antibodies in humans after vaccination and their clonal persistence as plasma cells. mBio 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Ong, H.; Yong, C.; Tan, W.; Yeap, S.; Omar, A.; Razak, M.; Ho, K. An influenza a vaccine based on the extracellular domain of matrix 2 protein protects BALB/C mice against H1N1 and H3N2. Vaccines 2019, 7, 91. [Google Scholar] [CrossRef]
- Antrobus, R.D.; Berthoud, T.K.; Mullarkey, C.E.; Hoschler, K.; Coughlan, L.; Zambon, M.; Hill, A.V.; Gilbert, S.C. Coadministration of seasonal influenza vaccine and MVA-NP+M1 simultaneously achieves potent humoral and cell-mediated responses. Mol. Ther. 2014, 22, 233–238. [Google Scholar] [CrossRef]
- Goodman, A.; Heinen, P.; Guerra, S.; Vijayan, A.; Sorzano, C.; Gomez, C.; Esteban, M. A human multi-epitope recombinant vaccinia virus as a universal T cell vaccine candidate against influenza virus. PLoS ONE 2011, 6, e25938. [Google Scholar] [CrossRef]
- Kim, J.; Cheong, S.; Lee, M. Evaluation of protective immunity of peptide vaccines composed of a 15-mer N-terminal matrix protein 2 and a helper t-cell epitope derived from influenza a virus. Immune Netw. 2019, 19, e29. [Google Scholar] [CrossRef]
- Blokhina, E.; Mardanova, E.; Stepanova, L.; Tsybalova, L.; Ravin, N. Plant-produced recombinant influenza a virus candidate vaccine based on flagellin linked to conservative fragments of M2 protein and hemagglutintin. Plants 2020, 9, 162. [Google Scholar] [CrossRef]
- Bimler, L.; Song, A.; Le, D.; Murphy, S.A.; Paust, S. AuNP-M2e+sCpG vaccination of juvenile mice generates lifelong protective immunity to influenza a virus infection. Immun. Ageing 2019, 16, 23. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Ren, Z.; Wang, Q.; Ge, S.; Liu, Y.; Liu, C.; Liu, F.; Hu, Y.; Li, J.; Bao, J.; et al. Infection of African swine fever in wild boar, China, 2018. Transbound Emerg. Dis. 2019, 66, 1395–1398. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kang, J.; Chang, J. Nucleoprotein vaccine induces cross-protective cytotoxic T lymphocytes against both lineages of influenza B virus. Clin. Exp. Vaccine Res. 2019, 8, 54–63. [Google Scholar] [CrossRef]
- Kim, M.; Kang, J.; Kim, J.; Jung, H.; Lee, H.; Chang, J. Single mucosal vaccination targeting nucleoprotein provides broad protection against two lineages of influenza B virus. Antivir. Res. 2019, 163, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Antrobus, R.D.; Coughlan, L.; Berthoud, T.K.; Dicks, M.D.; Hill, A.V.; Lambe, T.; Gilbert, S.C. Clinical assessment of a novel recombinant simian adenovirus ChAdOx1 as a vectored vaccine expressing conserved influenza A antigens. Mol. Ther. 2014, 22, 668–674. [Google Scholar] [CrossRef]
- Coughlan, L.; Sridhar, S.; Payne, R.; Edmans, M.; Milicic, A.; Venkatraman, N.; Lugonja, B.; Clifton, L.; Qi, C.; Folegatti, P.M.; et al. Heterologous two-dose vaccination with simian adenovirus and poxvirus vectors elicits long-lasting cellular immunity to influenza virus a in healthy adults. EBioMedicine 2018, 29, 146–154. [Google Scholar] [CrossRef]
- Del Campo, J.; Pizzorno, A.; Djebali, S.; Bouley, J.; Haller, M.; Perez-Vargas, J.; Lina, B.; Boivin, G.; Hamelin, M.E.; Nicolas, F.; et al. OVX836 a recombinant nucleoprotein vaccine inducing cellular responses and protective efficacy against multiple influenza A subtypes. NPJ Vaccines 2019, 4, 4. [Google Scholar] [CrossRef]
- Zhao, D.; Liu, R.; Zhang, X.; Li, F.; Wang, J.; Zhang, J.; Liu, X.; Wang, L.; Zhang, J.; Wu, X.; et al. Replication and virulence in pigs of the first African swine fever virus isolated in China. Emerg. Microbes Infect. 2019, 8, 438–447. [Google Scholar] [CrossRef]
- Rowell, J.; Lo, C.; Price, G.; Misplon, J.; Crim, R.; Jayanti, P.; Beeler, J.; Epstein, S. The effect of respiratory viruses on immunogenicity and protection induced by a candidate universal influenza vaccine in mice. PLoS ONE 2019, 14, e0215321. [Google Scholar] [CrossRef]
- Folegatti, P.M.; Bellamy, D.; Flaxman, A.; Mair, C.; Ellis, C.; Ramon, R.L.; Ramos Lopez, F.; Mitton, C.; Baker, M.; Poulton, I.; et al. Safety and Immunogenicity of the heterosubtypic influenza a vaccine MVA-NP+M1 manufactured on the AGE1.CR.pIX avian cell line. Vaccines 2019, 7, 33. [Google Scholar] [CrossRef]
- Swayze, H.; Allen, J.; Folegatti, P.; Yu, L.M.; Gilbert, S.; Hill, A.; Ellis, C.; Butler, C.C. A phase IIb study to determine the safety and efficacy of candidate INfluenza Vaccine MVA-NP+M1 in combination with licensed Ina CTivated infl Uenza vaccine in adult S aged 65 years and above (INVICTUS): A study protocol. F1000Res 2019, 8, 719. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Lu, H.; Zhang, D.; Tian, M.; Hu, B.; Wang, Z.; Jin, N. Protection against H1N1 influenza challenge by a DNA vaccine expressing H3/H1 subtype hemagglutinin combined with MHC class II-restricted epitopes. Virology J. 2010, 7, 363. [Google Scholar] [CrossRef]
- Eickhoff, C.; Terry, F.; Peng, L.; Meza, K.; Sakala, I.; Van, A.D.; Moise, L.; Martin, W.; Schriewer, J.; Buller, R.; et al. Highly conserved influenza T cell epitopes induce broadly protective immunity. Vaccine 2019, 37, 5371–5381. [Google Scholar] [CrossRef]
- Hassan, A.; Amen, O.; Sayedahmed, E.; Vemula, S.; Amoah, S.; York, I.; Gangappa, S.; Sambhara, S.; Mittal, S. Adenovirus vector-based multi-epitope vaccine provides partial protection against H5, H7, and H9 avian influenza viruses. PLoS ONE 2017, 12, e0186244. [Google Scholar] [CrossRef] [PubMed]
- Atsmon, J.; Caraco, Y.; Ziv-Sefer, S.; Shaikevich, D.; Abramov, E.; Volokhov, I.; Bruzil, S.; Haima, K.Y.; Gottlieb, T.; Ben-Yedidia, T. Priming by a novel universal influenza vaccine (Multimeric-001)-a gateway for improving immune response in the elderly population. Vaccine 2014, 32, 5816–5823. [Google Scholar] [CrossRef]
- Atsmon, J.; Kate-Ilovitz, E.; Shaikevich, D.; Singer, Y.; Volokhov, I.; Haim, K.Y.; Ben-Yedidia, T. Safety and immunogenicity of multimeric-001--a novel universal influenza vaccine. J. Clin. Immunol. 2012, 32, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Lowell, G.H.; Ziv, S.; Bruzil, S.; Babecoff, R.; Ben-Yedidia, T. Back to the future: Immunization with M-001 prior to trivalent influenza vaccine in 2011/12 enhanced protective immune responses against 2014/15 epidemic strain. Vaccine 2017, 35, 713–715. [Google Scholar] [CrossRef]
- Van Doorn, E.; Liu, H.; Ben-Yedidia, T.; Hassin, S.; Visontai, I.; Norley, S.; Frijlink, H.W.; Hak, E. Evaluating the immunogenicity and safety of a BiondVax-developed universal influenza vaccine (Multimeric-001) either as a standalone vaccine or as a primer to H5N1 influenza vaccine: Phase IIb study protocol. Medicine 2017, 96, e6339. [Google Scholar] [CrossRef]
- Pleguezuelos, O.; Dille, J.; de Groen, S.; Oftung, F.; Niesters, H.G.M.; Islam, M.A.; Næss, L.M.; Hungnes, O.; Aldarij, N.; Idema, D.L.; et al. Immunogenicity, safety, and efficacy of a standalone universal influenza vaccine, FLU-v, in healthy adults: A randomized clinical trial. Ann. Intern. Med. 2020, 172, 453–462. [Google Scholar] [CrossRef]
- Pleguezuelos, O.; James, E.; Fernandez, A.; Lopes, V.; Rosas, L.A.; Cervantes-Medina, A.; Cleath, J.; Edwards, K.; Neitzey, D.; Gu, W.; et al. Efficacy of FLU-v, a broad-spectrum influenza vaccine, in a randomized phase IIb human influenza challenge study. NPJ Vaccines 2020, 5, 22. [Google Scholar] [CrossRef]
- Van Doorn, E.; Pleguezuelos, O.; Liu, H.; Fernandez, A.; Bannister, R.; Stoloff, G.; Oftung, F.; Norley, S.; Huckriede, A.; Frijlink, H.W.; et al. Evaluation of the immunogenicity and safety of different doses and formulations of a broad spectrum influenza vaccine (FLU-v) developed by SEEK: Study protocol for a single-center, randomized, double-blind and placebo-controlled clinical phase IIb trial. BMC Infect. Dis. 2017, 17, 241. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, K.; Wegmann, F.; Tomaka, F.; Walsh, S.; Tan, C.; Lavreys, L.; Ansel, J.; Kanjilal, D.; Jaegle, K.; Nkolola, J.; et al. Comparison of shortened mosaic HIV-1 vaccine schedules: A randomised, double-blind, placebo-controlled phase 1 trial (IPCAVD010/HPX1002) and a preclinical study in rhesus monkeys (NHP 17-22). Lancet HIV 2020. [Google Scholar] [CrossRef]
- Zou, C.; Murakoshi, H.; Kuse, N.; Akahoshi, T.; Chikata, T.; Gatanaga, H.; Oka, S.; Hanke, T.; Takiguchi, M. Effective suppression of HIV-1 replication by cytotoxic T lymphocytes specific for pol epitopes in conserved mosaic vaccine immunogens. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [PubMed]
- Mega, E. Mosaic’ HIV vaccine to be tested in thousands of people across the world. Nature 2019, 572, 165–166. [Google Scholar] [CrossRef]
- Kamlangdee, A.; Kingstad-Bakke, B.; Osorio, J. Mosaic H5 hemagglutinin provides broad humoral and cellular immune responses against influenza viruses. J. Virol. 2016, 90, 6771–6783. [Google Scholar] [CrossRef]
- Florek, N.; Kamlangdee, A.; Mutschler, J.; Kingstad-Bakke, B.; Schultz-Darken, N.; Broman, K.; Osorio, J.; Friedrich, T. A modified vaccinia Ankara vaccine vector expressing a mosaic H5 hemagglutinin reduces viral shedding in rhesus macaques. PLoS ONE 2017, 12, e0181738. [Google Scholar] [CrossRef]
- Kamlangdee, A.; Kingstad-Bakke, B.; Anderson, T.K.; Goldberg, T.L.; Osorio, J.E. Broad protection against avian influenza virus by using a modified vaccinia Ankara virus expressing a mosaic hemagglutinin gene. J. Virol. 2014, 88, 13300–13309. [Google Scholar] [CrossRef]
- Corder, B.; Bullard, B.; DeBeauchamp, J.; Ilyushina, N.; Webby, R.; Weaver, E. Influenza H1 mosaic hemagglutinin vaccine induces broad immunity and protection in mice. Vaccines 2019, 7, 195. [Google Scholar] [CrossRef]
- Sun, W.; Kirkpatrick, E.; Ermler, M.; Nachbagauer, R.; Broecker, F.; Krammer, F.; Palese, P. Development of Influenza b universal vaccine candidates using the "Mosaic" hemagglutinin approach. J. Virol. 2019, 93. [Google Scholar] [CrossRef]
- Ross, T.; DiNapoli, J.; Giel-Moloney, M.; Bloom, C.; Bertran, K.; Balzli, C.; Strugnell, T.; Sá E Silva, M.; Mebatsion, T.; Bublot, M.; et al. A computationally designed H5 antigen shows immunological breadth of coverage and protects against drifting avian strains. Vaccine 2019, 37, 2369–2376. [Google Scholar] [CrossRef]
- Qi, M.; Zhang, X.; Sun, X.; Zhang, X.; Yao, Y.; Liu, S.; Chen, Z.; Li, W.; Zhang, Z.; Chen, J.; et al. Intranasal nanovaccine confers homo- and hetero-subtypic influenza protection. Small 2018, 14, e1703207. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Guo, J.; Turner, D.; Tizard, I. Development of a novel dual-domain nanoparticle antigen construct for universal influenza vaccine. Vaccine 2017, 35, 7026–7032. [Google Scholar] [CrossRef] [PubMed]
- Biswas, A.; Chakrabarti, A.K.; Dutta, S. Current challenges: From the path of "original antigenic sin" towards the development of universal flu vaccines. Int. Rev. Immunol. 2020, 39, 21–36. [Google Scholar] [CrossRef] [PubMed]
- Sulczewski, F.; Liszbinski, R.; Romão, P.; Rodrigues Junior, L. Nanoparticle vaccines against viral infections. Arch. Virol. 2018, 163, 2313–2325. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhu, W.; Luo, Y.; Wang, B. Gold nanoparticles conjugating recombinant influenza hemagglutinin trimers and flagellin enhanced mucosal cellular immunity. Nanomed. Nanotechnol. Biol. Med. 2018, 14, 1349–1360. [Google Scholar] [CrossRef]
- Portnoff, A.; Patel, N.; Massare, M.; Zhou, H.; Tian, J.; Zhou, B.; Shinde, V.; Glenn, G.; Smith, G. Influenza hemagglutinin nanoparticle vaccine elicits broadly neutralizing antibodies against structurally distinct domains of H3N2 HA. Vaccines 2020, 8, 99. [Google Scholar] [CrossRef]
- Wang, Y.; Deng, L.; Gonzalez, G.X.; Luthra, L.; Dong, C.; Ma, Y.; Zou, J.; Kang, S.-M.; Wang, B.-Z. Double-layered M2e-NA protein nanoparticle immunization induces broad cross-protection against different influenza viruses in mice. Adv. Healthc. Mater. 2020, 9, e1901176. [Google Scholar] [CrossRef]
- Georgiev, I.; Joyce, M.; Chen, R.; Leung, K.; McKee, K.; Druz, A.; Van Galen, J.; Kanekiyo, M.; Tsybovsky, Y.; Yang, E.; et al. Two-Component ferritin nanoparticles for multimerization of diverse trimeric antigens. ACS Infect. Dis. 2018, 4, 788–796. [Google Scholar] [CrossRef]
- Deng, L.; Wang, B. A perspective on nanoparticle universal influenza vaccines. ACS Infect. Dis. 2018, 4, 1656–1665. [Google Scholar] [CrossRef]
- Mezhenskaya, D.; Isakova-Sivak, I.; Rudenko, L. M2e-based universal influenza vaccines: A historical overview and new approaches to development. J. Biomed. Sci. 2019, 26, 76. [Google Scholar] [CrossRef]
- Kanekiyo, M.; Joyce, M.; Gillespie, R.; Gallagher, J.; Andrews, S.; Yassine, H.; Wheatley, A.; Fisher, B.; Ambrozak, D.; Creanga, A.; et al. Mosaic nanoparticle display of diverse influenza virus hemagglutinins elicits broad B cell responses. Nat. Immunol. 2019, 20, 362–372. [Google Scholar] [CrossRef] [PubMed]
- Bernasconi, V.; Bernocchi, B.; Ye, L.; Lê, M.; Omokanye, A.; Carpentier, R.; Schön, K.; Saelens, X.; Staeheli, P.; Betbeder, D.; et al. Porous nanoparticles with self-adjuvanting M2e-fusion protein and recombinant hemagglutinin provide strong and broadly protective immunity against influenza virus infections. Front. Immunol. 2018, 9, 2060. [Google Scholar] [CrossRef] [PubMed]
- Adler-Moore, J.; Ernst, W.; Kim, H.; Ward, N.; Chiang, S.; Do, T.; Fujii, G. Monomeric M2e antigen in VesiVax liposomes stimulates protection against type a strains of influenza comparable to liposomes with multimeric forms of M2e. J. Liposome Res. 2017, 27, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Dhakal, S.; Cheng, X.; Salcido, J.; Renu, S.; Bondra, K.; Lakshmanappa, Y.; Misch, C.; Ghimire, S.; Feliciano-Ruiz, N.; Hogshead, B.; et al. Liposomal nanoparticle-based conserved peptide influenza vaccine and monosodium urate crystal adjuvant elicit protective immune response in pigs. Int. J. Nanomed. 2018, 13, 6699–6715. [Google Scholar] [CrossRef]
- Wang, J.; Li, P.; Yu, Y.; Fu, Y.; Jiang, H.; Lu, M.; Sun, Z.; Jiang, S.; Lu, L.; Wu, M.X. Pulmonary surfactant–biomimetic nanoparticles potentiate heterosubtypic influenza immunity. Science 2020, 367. [Google Scholar] [CrossRef]
- Awate, S.; Babiuk, L.A.; Mutwiri, G. Mechanisms of action of adjuvants. Front. Immunol. 2013, 4, 114. [Google Scholar] [CrossRef]
- Uddowla, S.; Freytag, L.C.; Clements, J.D. Effect of adjuvants and route of immunizations on the immune response to recombinant plague antigens. Vaccine 2007, 25, 7984–7993. [Google Scholar] [CrossRef]
- Renu, S.; Feliciano-Ruiz, N.; Ghimire, S.; Han, Y.; Schrock, J.; Dhakal, S.; Patil, V.; Krakowka, S.; Renukaradhya, G. Poly(I:C) augments inactivated influenza virus-chitosan nanovaccine induced cell mediated immune response in pigs vaccinated intranasally. Vet. Microbiol. 2020, 242, 108611. [Google Scholar] [CrossRef]
- Schussek, S.; Bernasconi, V.; Mattsson, J.; Wenzel, U.; Strömberg, A.; Gribonika, I.; Schön, K.; Lycke, N. The CTA1-DD adjuvant strongly potentiates follicular dendritic cell function and germinal center formation, which results in improved neonatal immunization. Mucosal Immunol. 2020. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, S.; Bi, S.; Li, N.; Bi, Y.; Liu, W.; Wang, B. Long-lasting protective immunity against H7N9 infection is induced by intramuscular or CpG-adjuvanted intranasal immunization with the split H7N9 vaccine. Int. Immunopharmacol. 2020, 78, 106013. [Google Scholar] [CrossRef]
- Mathew, M.; Virmani, N.; Bera, B.; Anand, T.; Kumar, R.; Balena, V.; Sansanwal, R.; Pavulraj, S.; Sundaram, K.; Virmani, M.; et al. Protective efficacy of inactivated reverse genetics based equine influenza vaccine candidate adjuvanted with montanide pet gel in murine model. J. Vet. Med Sci. 2019, 81, 1753–1762. [Google Scholar] [CrossRef] [PubMed]
- Short, K.; Miller, S.; Walsh, L.; Cybulski, V.; Bazin, H.; Evans, J.; Burkhart, D. Co-encapsulation of synthetic lipidated TLR4 and TLR7/8 agonists in the liposomal bilayer results in a rapid, synergistic enhancement of vaccine-mediated humoral immunity. J. Control. Release 2019, 315, 186–196. [Google Scholar] [CrossRef] [PubMed]
- Saelens, X. The role of matrix protein 2 ectodomain in the development of universal influenza vaccines. J. Infect. Dis. 2019, 219, S68–S74. [Google Scholar] [CrossRef] [PubMed]
- Rudicell, R.; Garinot, M.; Kanekiyo, M.; Kamp, H.; Swanson, K.; Chou, T.; Dai, S.; Bedel, O.; Simard, D.; Gillespie, R.; et al. Comparison of adjuvants to optimize influenza neutralizing antibody responses. Vaccine 2019, 37, 6208–6220. [Google Scholar] [CrossRef]
- Reed, S.G.; Orr, M.T.; Fox, C.B. Key roles of adjuvants in modern vaccines. Nat. Med. 2013, 19, 1597–1608. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Vaccine Type | Protein Expression System | Protection Ratio | Advantage | Disadvantage | References | |
|---|---|---|---|---|---|---|
| Homologous | Heterologous | |||||
| HA stem | Eukaryotic expression | Complete protection | Partial protection; Poor protection for different HA groups | Single HA stem can produce extensive protection for the same HA group | Protection is limited by group differences | [22,23,24,25,26,27,28,29,30,31,32,33] |
| Chimeric HA | Eukaryotic expression or rescue chimeric attenuated virus vaccine | Complete protection | Partial protection; Poor protection for different HA groups | Easy production of chimeric attenuated vaccine | Protection is limited by the type of chimeric subtype and requires multiple immunizations with different chimeric vaccines | [24,33,34,35,36,37,38,39] |
| NA | Eukaryotic expression | Complete protection | Partially protected or unprotected | Strong ability to protect different HA subtype strains of the same NA type | Poor protection against different NA types | [40,41,42,43,44,45] |
| M1, M2e, NP | Viral vectors, plasmids | Partial protection | Generates different protection according to M1, M2e and NP sequence differences | Wide range of protection without being limited by HA group differences | Not fully protected; Poor immunogenicity requires tandem or combined expression with other proteins | [39,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61] |
| Epitope peptide | Chemical synthesis, prokaryotic expression, viral vectors, plasmids | Partial protection | Different protection depending on how much dominant epitope of certain flu covered | Simple, stable, easy to synthesize, non-toxic; not restricted by HA group differences | Difficult to screen for co-conserved epitopes in large influenza databases; Limited by population MHC diversity; Poor immunogenicity | [62,63,64,65,66,67,68,69,70,71] |
| Mosaic | Viral vector, eukaryotic expression | Complete protection | Different protection based on the subtype on which the Mosaic design is based | Strong protection and extensive protection for different clades of the same subtype | Difficult to find a sequence that covers all epitopes in the large influenza database | [72,73,74,75,76,77,78,79,80] |
| Nanoparticles | Gold nanoparticles, polymers, VLPs, liposomes | Different protection according to the type of antigen loaded | Efficiently improve immunogenicity; Load multiple antigens | Complex preparation process | [81,82,83,84,85,86,87,88,89,90,91,92,93,94,95] | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, W.; Luo, T.; Liu, W.; Li, J. Progress in the Development of Universal Influenza Vaccines. Viruses 2020, 12, 1033. https://doi.org/10.3390/v12091033
Sun W, Luo T, Liu W, Li J. Progress in the Development of Universal Influenza Vaccines. Viruses. 2020; 12(9):1033. https://doi.org/10.3390/v12091033
Chicago/Turabian StyleSun, Wenqiang, Tingrong Luo, Wenjun Liu, and Jing Li. 2020. "Progress in the Development of Universal Influenza Vaccines" Viruses 12, no. 9: 1033. https://doi.org/10.3390/v12091033
APA StyleSun, W., Luo, T., Liu, W., & Li, J. (2020). Progress in the Development of Universal Influenza Vaccines. Viruses, 12(9), 1033. https://doi.org/10.3390/v12091033