Differential Detection of Encapsidated versus Unencapsidated Enterovirus RNA in Samples Containing Pancreatic Enzymes—Relevance for Diabetes Studies

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Participating Virology Laboratories
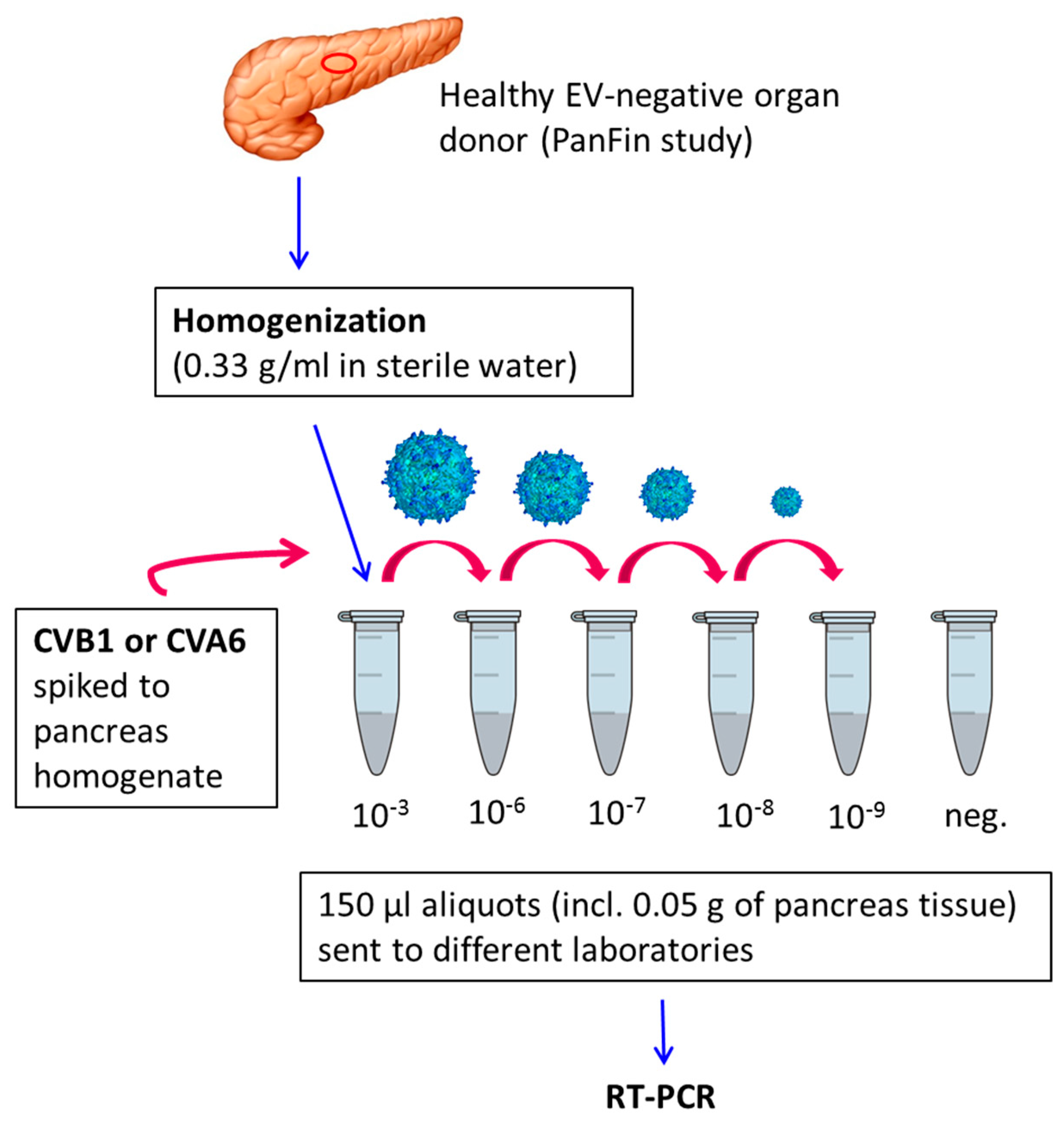
2.2. Preparation of Virus-Spiked Human Pancreas Homogenate (University of Tampere, Finland)
2.3. Virus Detection by RT-PCR
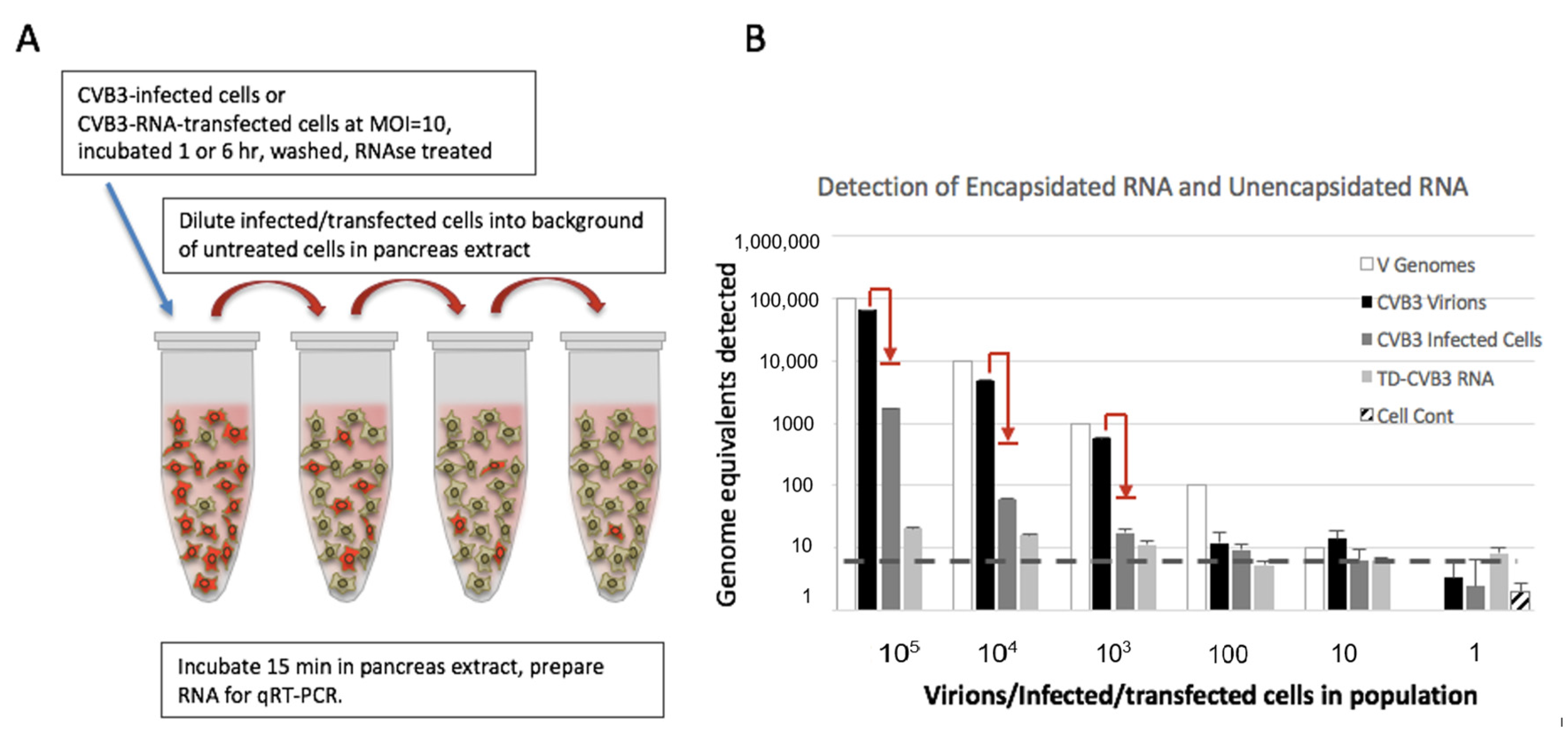
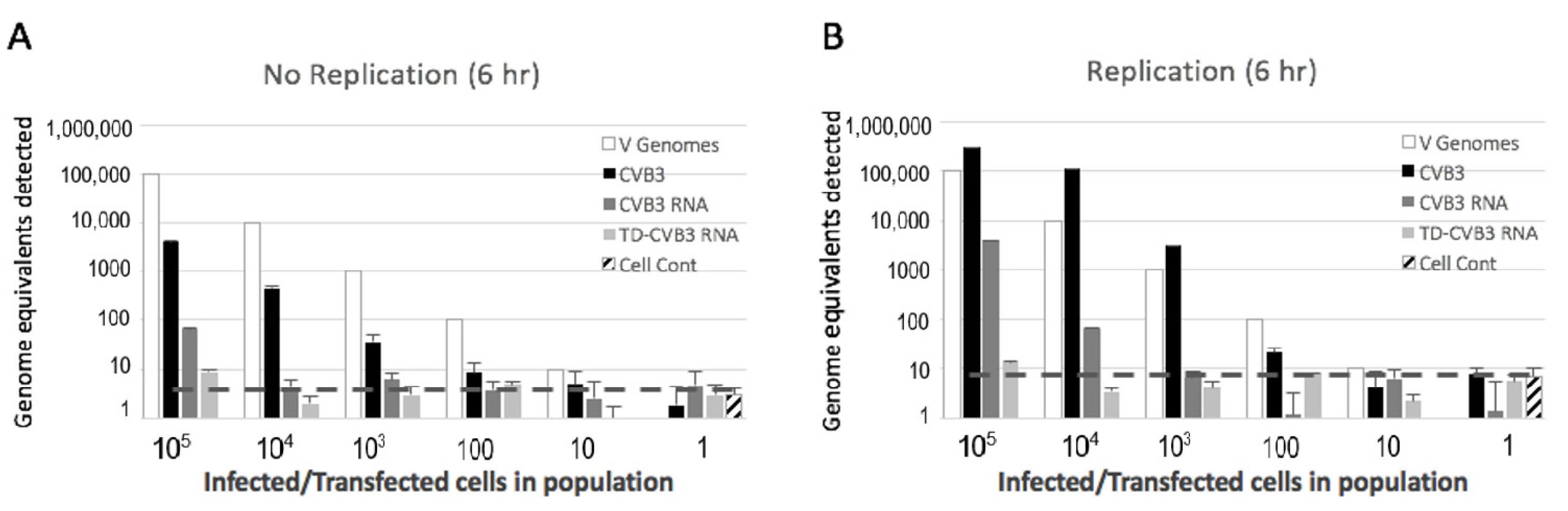
2.4. Preparation and Detection of Full-Length and Terminally Deleted Coxsackievirus B3 (Baylor College of Medicine, Houston, Texas)
3. Results
3.1. Sensitivity of Different RT-PCR Methods to Detect Enterovirus RNA
3.2. Detection of Encapsidated and Unencapsidated Enterovirus RNA
3.3. Detection of Replicating and Non-Replicating Enterovirus
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Morgan, N.G.; Richardson, S.J. Enteroviruses as causative agents in type 1 diabetes: loose ends or lost cause? Trends Endocrinol. Metab. 2014, 25, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Krogvold, L.; Edwin, B.; Buanes, T.; Frisk, G.; Skog, O.; Anagandula, M.; Korsgren, O.; Undlien, D.; Eike, M.C.; Richardson, S.J.; et al. Detection of a low-grade enteroviral infection in the islets of Langerhans of living patients newly diagnosed with type 1 diabetes. Diabetes 2015, 64, 1682–1687. [Google Scholar] [CrossRef]
- Dotta, F.; Censini, S.; van Halteren, A.G.; Marselli, L.; Masini, M.; Dionisi, S.; Mosca, F.; Boggi, U.; Muda, A.O.; Del Prato, S.; et al. Coxsackie B4 virus infection of beta cells and natural killer cell insulitis in recent-onset type 1 diabetic patients. Proc. Natl. Acad. Sci. USA 2007, 104, 5115–5120. [Google Scholar] [CrossRef] [PubMed]
- Richardson, S.J.; Leete, P.; Dhayal, S.; Russell, M.A.; Oikarinen, M.; Laiho, J.E.; Svedin, E.; Lind, K.; Rosenling, T.; Chapman, N.; et al. Detection of enterovirus in the islet cells of patients with type 1 diabetes: what do we learn from immunohistochemistry? Reply to Hansson SF, Korsgren S, Ponten F et al [letter]. Diabetologia 2014, 57, 647–649. [Google Scholar] [CrossRef] [PubMed]
- Ylipaasto, P.; Klingel, K.; Lindberg, A.M.; Otonkoski, T.; Kandolf, R.; Hovi, T.; Roivainen, M. Enterovirus infection in human pancreatic islet cells, islet tropism in vivo and receptor involvement in cultured islet beta cells. Diabetologia 2004, 47, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Richardson, S.J.; Leete, P.; Bone, A.J.; Foulis, A.K.; Morgan, N.G. Expression of the enteroviral capsid protein VP1 in the islet cells of patients with type 1 diabetes is associated with induction of protein kinase R and downregulation of Mcl-1. Diabetologia 2013, 56, 185–193. [Google Scholar] [CrossRef]
- Chapman, N.M.; Kim, K.S.; Drescher, K.M.; Oka, K.; Tracy, S. 5' terminal deletions in the genome of a coxsackievirus B2 strain occurred naturally in human heart. Virology 2008, 375, 480–491. [Google Scholar] [CrossRef]
- Tracy, S.; Smithee, S.; Alhazmi, A.; Chapman, N. Coxsackievirus can persist in murine pancreas by deletion of 5' terminal genomic sequences. J. Med. Virol. 2015, 87, 240–247. [Google Scholar] [CrossRef]
- Kim, K.S.; Tracy, S.; Tapprich, W.; Bailey, J.; Lee, C.K.; Kim, K.; Barry, W.H.; Chapman, N.M. 5'-Terminal deletions occur in coxsackievirus B3 during replication in murine hearts and cardiac myocyte cultures and correlate with encapsidation of negative-strand viral RNA. J. Virol. 2005, 79, 7024–7041. [Google Scholar] [CrossRef]
- Sharma, N.; O'Donnell, B.J.; Flanegan, J.B. 3'-Terminal sequence in poliovirus negative-strand templates is the primary cis-acting element required for VPgpUpU-primed positive-strand initiation. J. Virol. 2005, 79, 3565–3577. [Google Scholar] [CrossRef]
- Vogt, D.A.; Andino, R. An RNA element at the 5'-end of the poliovirus genome functions as a general promoter for RNA synthesis. PLoS Pathog. 2010, 6, e1000936. [Google Scholar] [CrossRef] [PubMed]
- Ertel, K.J.; Brunner, J.E.; Semler, B.L. Mechanistic consequences of hnRNP C binding to both RNA termini of poliovirus negative-strand RNA intermediates. J. Virol. 2010, 84, 4229–4242. [Google Scholar] [CrossRef] [PubMed]
- Andino, R.; Rieckhof, G.E.; Achacoso, P.L.; Baltimore, D. Poliovirus RNA synthesis utilizes an RNP complex formed around the 5'-end of viral RNA. EMBO J. 1993, 12, 3587–3598. [Google Scholar] [CrossRef] [PubMed]
- Nugent, C.I.; Johnson, K.L.; Sarnow, P.; Kirekegaard, K. Functional coupling between replication and packaging of poliovirus replicon RNA. J. Virol. 1999, 73, 427–435. [Google Scholar] [CrossRef]
- Augereau, C.; Lemaigre, F.P.; Jacquemin, P. Extraction of High-Quality RNA From Pancreatic Tissues for Gene Expression Studies. Anal. Biochem. 2016, 500, 60–62. [Google Scholar] [CrossRef]
- Kaddis, J.S.; Pugliese, A.; Atkinson, M.A. A Run on the Biobank: What Have We Learned About Type 1 Diabetes From the nPOD Tissue Repository? Curr. Opin. Endocrinol. Diabetes Obes. 2015, 22, 290–295. [Google Scholar] [CrossRef]
- Laiho, J.E.; Oikarinen, M.; Richardson, S.J.; Frisk, J.; Nyalwidhe, J.; Burch, T.C.; Morris, M.A.; Oikarinen, S.; Pugliese, A.; Dotta, F.; et al. Relative sensitivity of immunohistochemistry, multiple reaction monitoring mass spectrometry, in situ hybridization and PCR to detect Coxsackievirus B1 in A549 cells. J. Clin. Virol. 2016, 77, 21–28. [Google Scholar] [CrossRef]
- Tauriainen, S.; Salmela, K.; Rantala, I.; Knip, M.; Hyöty, H. Collecting high-quality pancreatic tissue for experimental study from organ donors with signs of beta-cell autoimmunity. Diabetes Metab. Res. Rev. 2010, 26, 585–592. [Google Scholar] [CrossRef]
- Lonnrot, M.; Sjoroos, M.; Salminen, K.; Maaronen, M.; Hyypiä, T.; Hyöty, H. Diagnosis of enterovirus and rhinovirus infections by RT-PCR and time-resolved fluorometry with lanthanide chelate labeled probes. J. Med. Virol. 1999, 59, 378–384. [Google Scholar] [CrossRef]
- Honkanen, H.; Oikarinen, S.; Pakkanen, O.; Ruokoranta, T.; Pulkki, M.M.; Laitinen, O.H.; Tauriainen, S.; Korpela, S.; Lappalainen, M.; Vuorinen, T.; et al. Human enterovirus 71 strains in the background population and in hospital patients in Finland. J. Clin. Virol. 2013, 56, 348–353. [Google Scholar] [CrossRef]
- Genoni, A.; Canducci, F.; Rossi, A.; Broccolo, B.; Chumakov, K.; Bono, G.; Salerno-Uriarte, J.; Salvatoni, A.; Pugliese, A.; Toniolo, A. Revealing Enterovirus Infection in Chronic Human Disorders: An Integrated Diagnostic Approach. Sci. Rep. 2017, 7, 5013. [Google Scholar] [CrossRef] [PubMed]
- Smithee, S.; Tracy, S.; Chapman, N.M. Mutational Disruption of cis-Acting Replication Element 2C in Coxsackievirus B3 Leads to 5′-Terminal Genomic Deletions. J. Virol. 2015, 89, 11761–11772. [Google Scholar] [CrossRef] [PubMed]
- Kempf, B.J.; Barton, D.J. Poliovirus 2A(Pro) increases viral mRNA and polysome stability coordinately in time with cleavage of eIF4G. J. Virol. 2008, 82, 5847–5859. [Google Scholar] [CrossRef] [PubMed]
- Vehik, K.; Fiske, S.W.; Logan, C.A.; Agardh, D.; Cilio, C.M.; Hagopian, W.; Simell, O.; Roivainen, M.; She, J.X.; Brieseet, T.; et al. Methods, Quality Control and Specimen Management in an International Multicentre Investigation of Type 1 Diabetes: TEDDY. Diabetes Metab. Res. Rev. 2013, 29, 557–567. [Google Scholar] [CrossRef] [PubMed]
- Feuer, R.; Ruller, C.M.; An, N.; Tabor-Godwin, J.M.; Rhoades, R.E.; Maciejewski, S.; Pagarigan, R.R.; Cornell, C.T.; Crocker, S.J.; Kiosseset, W.B.; et al. Viral persistence and chronic immunopathology in the adult central nervous system following Coxsackievirus infection during the neonatal period. J. Virol. 2009, 83, 9356–9369. [Google Scholar] [CrossRef]
- N'Guyen, Y.; Lesaffre, F.; Metz, D.; Tassan, S.; Saade, Y.; Boulagnon, C.; Fornes, P.; Renois, F.; Andreoletti, L. Enterovirus but not Parvovirus B19 is associated with idiopathic dilated cardiomyopathy and endomyocardial CD3, CD68, or HLA-DR expression. J. Med. Virol. 2017, 89, 55–63. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Laboratory | Method | EV Group | Forward Primer | Reverse Primer | Probe | Location in the Genome * |
|---|---|---|---|---|---|---|
| Houston | PCR 1 | 5′UTR-A-D | CGGCCCCTGAATGCGGCTAA | GAAACACGGACACCCAAAGTA | 449–563 | |
| Tampere | PCR 1 | 5′UTR-A-D | CGGCCCCTGAATGCGGCTAA | GAAACACGGACACCCAAAGTA | TAITCGGTTCCGCTGC | 449–563 |
| PCR 2 | 5′UTR-A-D | CGGCCCCTGAATGCGGCTAA | GAAACACGGACACCCAAAGTA | FAM-TCTGTGGCG GAA CCGACTA-TAMRA FAM-TCTGCAGCGGAA CCGACTA-TAMRA | 449–563 | |
| Varese | PCR 1 and 2 | 5′UTR-A | GTGTAGATCAGGTCGATGAGTCAC | ATTGTCACCATAAGCAGCCA | 306–597 | |
| 5′UTR-B | GACCAAGCACTTCTGTTACCC | GTCACCATAAGCAGCCAATATA | 161–594 | |||
| 5′UTR-C | GGTGTGAAGAGCCTATTGAGC | GATTGTCACCATAAGCAGCCA | 413–598 | |||
| 5′UTR-D | TGGTCCAGGCTGCGTT | AACACGGACACCCAAAGTAGT | 351–561 |
| Houston | Tampere | Varese | |||
|---|---|---|---|---|---|
| Sample | PCR 1 | PCR 1 | PCR 2 | PCR 1 | PCR 2 |
| CVB1 in water | 10−7 | 10−9 | 10−9 | 10−6 | n.a. |
| CVB1 in pancreas extract | 10−8 | 10−9 | 10−9 | 10−9 | 10−9 |
| CVB1 in pancreas extract, 20 h on ice | 10−8 | 10−8 | 10−8 | 10−9 | 10−9 |
| CVA6 in pancreas extract | 10−7 | 10−7 | 10−7 | 10−8 | 10−9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oikarinen, M.; Bertolet, L.; Toniolo, A.; Oikarinen, S.; Laiho, J.E.; Pugliese, A.; Lloyd, R.E.; Hyöty, H.; the nPOD-V Study Group. Differential Detection of Encapsidated versus Unencapsidated Enterovirus RNA in Samples Containing Pancreatic Enzymes—Relevance for Diabetes Studies. Viruses 2020, 12, 747. https://doi.org/10.3390/v12070747
Oikarinen M, Bertolet L, Toniolo A, Oikarinen S, Laiho JE, Pugliese A, Lloyd RE, Hyöty H, the nPOD-V Study Group. Differential Detection of Encapsidated versus Unencapsidated Enterovirus RNA in Samples Containing Pancreatic Enzymes—Relevance for Diabetes Studies. Viruses. 2020; 12(7):747. https://doi.org/10.3390/v12070747
Chicago/Turabian StyleOikarinen, Maarit, Lori Bertolet, Antonio Toniolo, Sami Oikarinen, Jutta E. Laiho, Alberto Pugliese, Richard E. Lloyd, Heikki Hyöty, and the nPOD-V Study Group. 2020. "Differential Detection of Encapsidated versus Unencapsidated Enterovirus RNA in Samples Containing Pancreatic Enzymes—Relevance for Diabetes Studies" Viruses 12, no. 7: 747. https://doi.org/10.3390/v12070747
APA StyleOikarinen, M., Bertolet, L., Toniolo, A., Oikarinen, S., Laiho, J. E., Pugliese, A., Lloyd, R. E., Hyöty, H., & the nPOD-V Study Group. (2020). Differential Detection of Encapsidated versus Unencapsidated Enterovirus RNA in Samples Containing Pancreatic Enzymes—Relevance for Diabetes Studies. Viruses, 12(7), 747. https://doi.org/10.3390/v12070747