In Vitro Systems for Studying Different Genotypes/Sub-Genotypes of Hepatitis B Virus: Strengths and Limitations



Abstract
1. Introduction
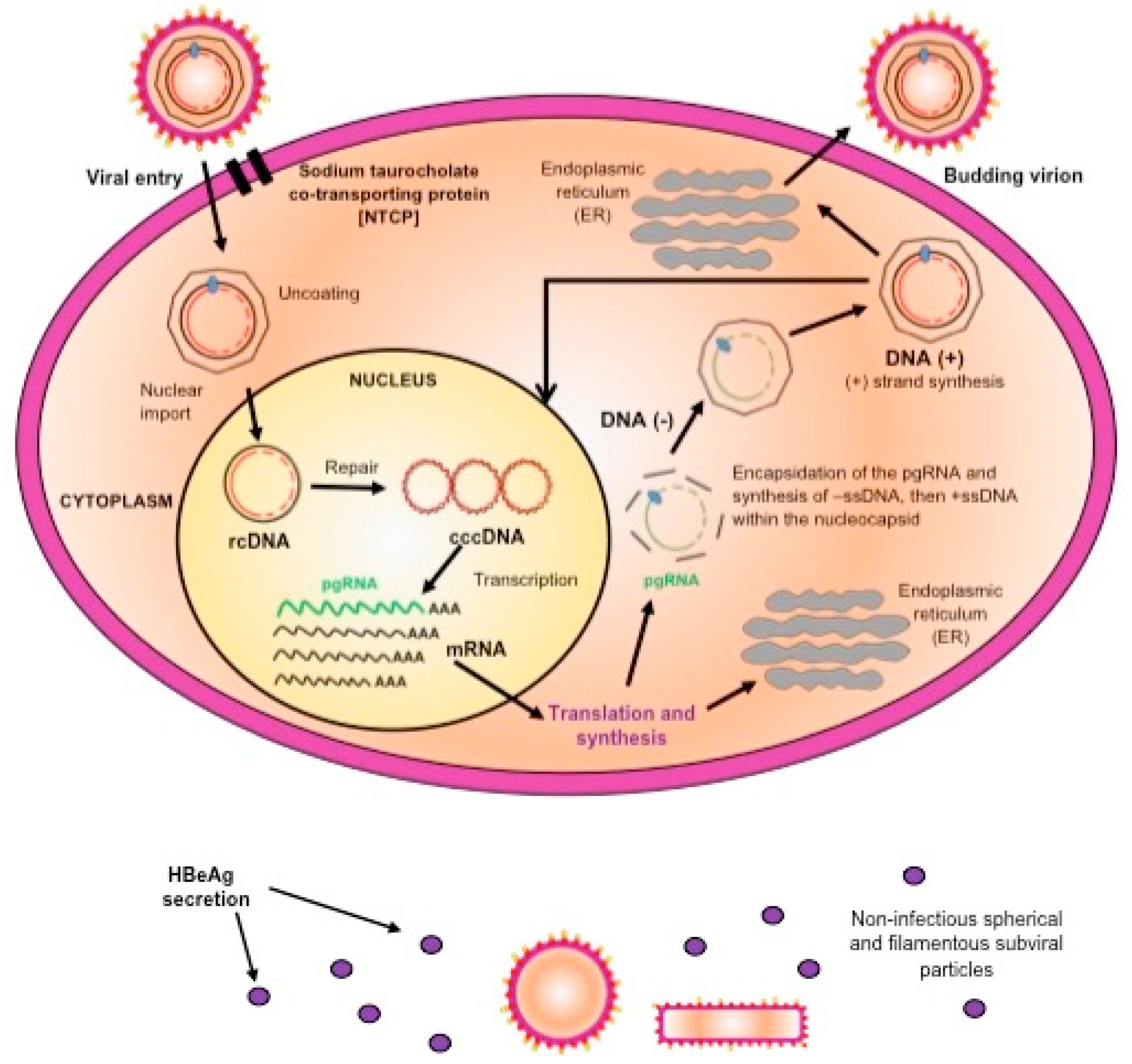
2. Molecular Biology of Hepatitis B Virus
3. Genotypes/Sub-Genotypes of HBV
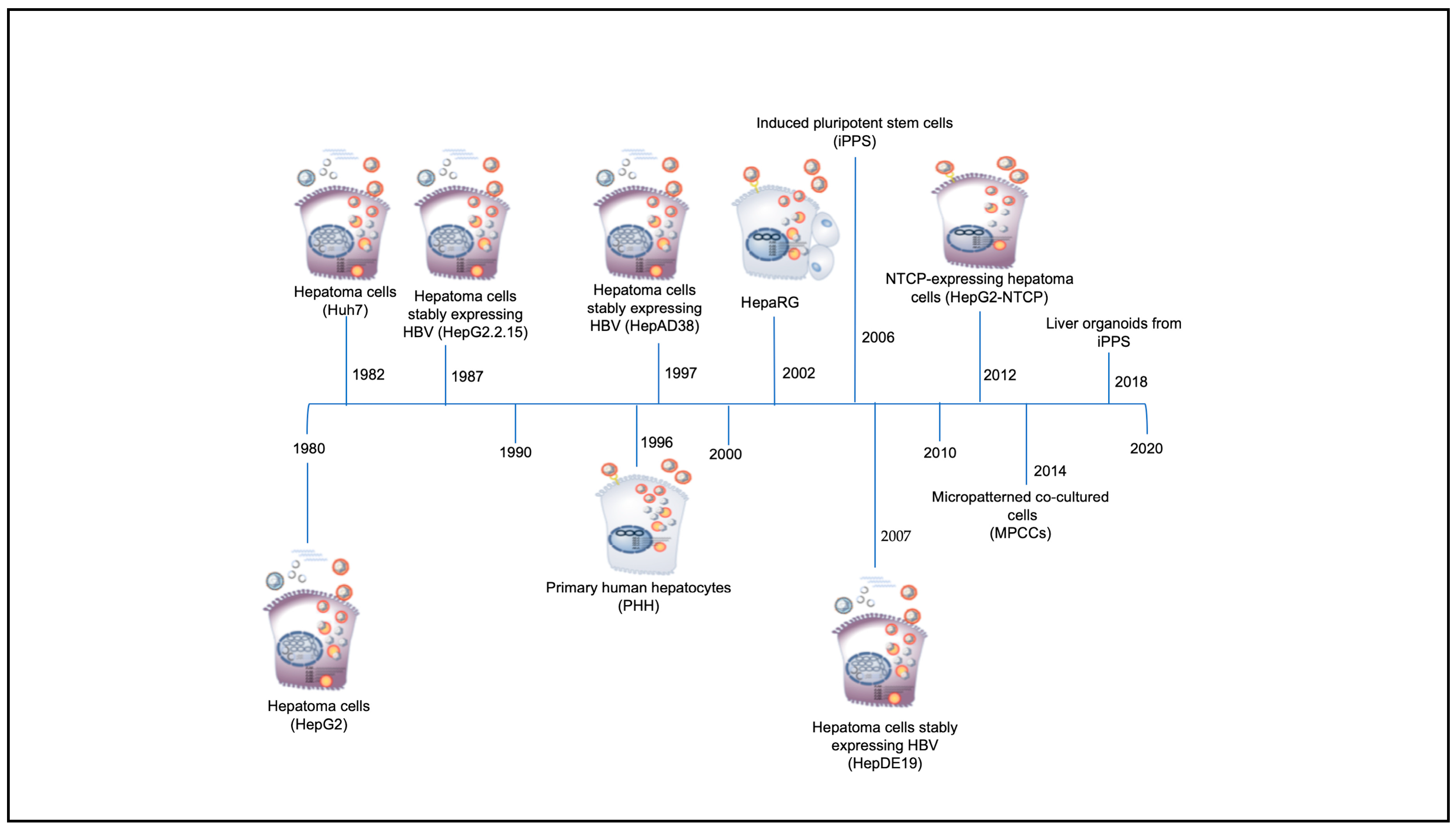
4. In Vitro Systems for the Study of HBV
4.1. In Vitro Model Systems Based on Hepatoma Cells (HepG2, Huh7, HepG2.2.15, and HepAD38)
4.2. In Vitro Model Systems Based on Primary Human Hepatocytes (PHH)
4.3. In Vitro Model Systems Based on Differentiated Hepatoma Cell Lines (HepaRG)
4.4. In Vitro Model Systems Based on NTCP Expressing Cell Lines
4.5. In Vitro Model Systems Based on Inducible Pluripotent Stem Cell (iPSCs)
4.6. In Vitro Model Systems Based on Micropatterned Co-Cultured Cells (MPPCs)
4.7. In Vitro Model Systems Based on Liver Organoids
- Express NTCP
- Maintain hepatocyte function and susceptibility to HBV infection indefinitely
- Not require the addition of DMSO to maintain hepatocyte function or PEG to promote infection
- Be capable of being infected with high efficiency with multiple HBV genotypes/sub-genotypes and variants.
- Express the host factors necessary to support HBV infection
- Have high longevity to support the complete viral life cycle
- Have an intact innate immune response
- Have functional pathways
- Be genetically homogeneous
- Recapitulate HBV infection seen in patients or in vivo systems
- Be renewable
- Be of unlimited supply
- Allow for the testing of a wide range of antiviral and immunomodulatory agents
- Be low cost
- Allow for miniaturization
- Be ethically acceptable
5. The Use of In Vitro Systems to Study Genotypes/Sub-Genotypes of HBV
6. Knowledge Gaps and Future Prospective
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Prince, A.M. Relation of Australia and SH antigens. Lancet 1968, 2, 462–463. [Google Scholar] [CrossRef]
- Prince, A.M. An antigen detected in the blood during the incubation period of serum hepatitis. Proc. Natl. Acad. Sci. USA 1968, 60, 814–821. [Google Scholar] [CrossRef]
- Blumberg, B.S.; Alter, H.J.; Visnich, S. A “New” Antigen in Leukemia Sera. JAMA 1965, 191, 541–546. [Google Scholar] [CrossRef]
- Blumberg, B.S.; Gerstley, B.J.; Hungerford, D.A.; London, W.T.; Sutnick, A.I. A serum antigen (Australia antigen) in Down’s syndrome, leukemia, and hepatitis. Ann. Intern. Med. 1967, 66, 924–931. [Google Scholar] [CrossRef]
- Alter, H.J.; Blumberg, B.S. Further studies on a “new” human isoprecipitin system (Australia antigen). Blood 1966, 27, 297–309. [Google Scholar] [CrossRef]
- World Health Organization. Global Hepatitis Report, 2017; WHO: Geneva, Switzerland, 2017; ISBN 978-92-4-156545-5. [Google Scholar]
- Flink, H.J.; van Zonneveld, M.; Hansen, B.E.; de Man, R.A.; Schalm, S.W.; Janssen, H.L.A.; Grp, H.-S. Treatment with Peg-interferon alpha-2b for HBeAg-positive chronic hepatitis B: HBsAg loss is associated with HBV genotype. Am. J. Gastroenterol. 2006, 101, 297–303. [Google Scholar] [CrossRef]
- Sonneveld, M.J.; Rijckborst, V.; Boucher, C.A.B.; Hansen, B.E.; Janssen, H.L.A. Prediction of Sustained Response to Peginterferon Alfa-2b for Hepatitis B e Antigen Positive Chronic Hepatitis B Using On-Treatment Hepatitis B Surface Antigen Decline. Hepatology 2010, 52, 1251–1257. [Google Scholar] [CrossRef] [PubMed]
- Tseng, T.C.; Yu, M.L.; Liu, C.J.; Lin, C.L.; Huang, Y.W.; Hsu, C.S.; Liu, C.H.; Kuo, S.F.T.; Pan, C.J.H.; Yang, S.S.; et al. Effect of host and viral factors on hepatitis B e antigen-positive chronic hepatitis B patients receiving pegylated interferon-alpha-2a therapy. Antivir. Ther. 2011, 16, 629–637. [Google Scholar] [CrossRef]
- Arends, P.; Rijckborst, V.; Zondervan, P.E.; Buster, E.; Cakaloglu, Y.; Ferenci, P.; Tabak, F.; Akarca, U.S.; Simon, K.; Sonneveld, M.J.; et al. Loss of intrahepatic HBsAg expression predicts sustained response to peginterferon and is reflected by pronounced serum HBsAg decline. J. Viral Hepat. 2014, 21, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Revill, P.A.; Chisari, F.V.; Block, J.M.; Dandri, M.; Gehring, A.J.; Guo, H.; Hu, J.; Kramvis, A.; Lampertico, P.; Janssen, H.L.A.; et al. A global scientific strategy to cure hepatitis B. Lancet Gastroenterol. Hepatol. 2019, 4, 545–558. [Google Scholar] [CrossRef]
- Yan, H.; Zhong, G.; Xu, G.; He, W.; Jing, Z.; Gao, Z.; Huang, Y.; Qi, Y.; Peng, B.; Wang, H.; et al. Sodium taurocholate cotransporting polypeptide is a functional receptor for human hepatitis B and D virus. Elife 2012, 1, e00049. [Google Scholar] [CrossRef] [PubMed]
- Kramvis, A. Genotypes and genetic variability of hepatitis B virus. Intervirology 2014, 57, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Kramvis, A.; Kew, M.; Francois, G. Hepatitis B virus genotypes. Vaccine 2005, 23, 2409–2423. [Google Scholar] [CrossRef] [PubMed]
- Velkov, S.; Ott, J.J.; Protzer, U.; Michler, T. The Global Hepatitis B Virus Genotype Distribution Approximated from Available Genotyping Data. Genes 2018, 9, 495. [Google Scholar] [CrossRef] [PubMed]
- Kramvis, A.; Kew, M.C. Epidemiology of hepatitis B virus in Africa, its genotypes and clinical associations of genotypes. Hepatol. Res. 2007, 37, S9–S19. [Google Scholar] [CrossRef] [PubMed]
- Tiollais, P.; Pourcel, C.; Dejean, A. The hepatitis B virus. Nature 1985, 317, 489–495. [Google Scholar] [CrossRef]
- Beck, J.; Nassal, M. Hepatitis B virus replication. World J. Gastroenterol. 2007, 13, 48–64. [Google Scholar] [CrossRef]
- Yu, H.; Yuan, Q.; Ge, S.X.; Wang, H.Y.; Zhang, Y.L.; Chen, Q.R.; Zhang, J.; Chen, P.J.; Xia, N.S. Molecular and phylogenetic analyses suggest an additional hepatitis B virus genotype “I”. PLoS ONE 2010, 5, e9297. [Google Scholar] [CrossRef]
- Norder, H.; Courouce, A.M.; Coursaget, P.; Echevarria, J.M.; Lee, S.D.; Mushahwar, I.K.; Robertson, B.H.; Locarnini, S.; Magnius, L.O. Genetic diversity of hepatitis B virus strains derived worldwide: Genotypes, subgenotypes, and HBsAg subtypes. Intervirology 2004, 47, 289–309. [Google Scholar] [CrossRef]
- Tatematsu, K.; Tanaka, Y.; Kurbanov, F.; Sugauchi, F.; Mano, S.; Maeshiro, T.; Nakayoshi, T.; Wakuta, M.; Miyakawa, Y.; Mizokami, M. A genetic variant of hepatitis B virus divergent from known human and ape genotypes isolated from a Japanese patient and provisionally assigned to new genotype J. J. Virol. 2009, 83, 10538–10547. [Google Scholar] [CrossRef]
- Locarnini, S.; Littlejohn, M.; Aziz, M.N.; Yuen, L. Possible origins and evolution of the hepatitis B virus (HBV). Semin. Cancer Biol. 2013, 23, 561–575. [Google Scholar] [CrossRef] [PubMed]
- Littlejohn, M.; Davies, J.; Yuen, L.; Edwards, R.; Sozzi, T.; Jackson, K.; Cowie, B.; Tong, S.; Davis, J.; Locarnini, S. Molecular virology of hepatitis B virus, sub-genotype C4 in northern Australian Indigenous populations. J. Med. Virol. 2014, 86, 695–706. [Google Scholar] [CrossRef] [PubMed]
- Kramvis, A.; Kew, M.C. Relationship of genotypes of hepatitis B virus to mutations, disease progression and response to antiviral therapy. J. Viral Hepat. 2005, 12, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Kramvis, A.; Arakawa, K.; Yu, M.C.; Nogueira, R.; Stram, D.O.; Kew, M.C. Relationship of serological subtype, basic core promoter and precore mutations to genotypes/subgenotypes of hepatitis B virus. J. Med. Virol. 2008, 80, 27–46. [Google Scholar] [CrossRef] [PubMed]
- Kramvis, A.; Kostaki, E.G.; Hatzakis, A.; Paraskevis, D. Immunomodulatory Function of HBeAg Related to Short-Sighted Evolution, Transmissibility, and Clinical Manifestation of Hepatitis B Virus. Front. Microbiol. 2018, 9, 2521. [Google Scholar] [CrossRef]
- Kao, J.H. Hepatitis B viral genotypes: Clinical relevance and molecular characteristics. J. Gastroenterol. Hepatol. 2002, 17, 643–650. [Google Scholar] [CrossRef]
- McMahon, B.J. The influence of hepatitis B virus genotype and subgenotype on the natural history of chronic hepatitis B. Hepatol. Int. 2009, 3, 334–342. [Google Scholar] [CrossRef]
- Gerlich, W.H. Prophylactic vaccination against hepatitis B: Achievements, challenges and perspectives. Med. Microbiol. Immunol. 2015, 204, 39–55. [Google Scholar] [CrossRef]
- Gerlich, W.H. Do we need better hepatitis B vaccines? Indian J. Med. Res. 2017, 145, 414–419. [Google Scholar] [CrossRef]
- De Meyer, S.; Gong, Z.J.; Suwandhi, W.; van Pelt, J.; Soumillion, A.; Yap, S.H. Organ and species specificity of hepatitis B virus (HBV) infection: A review of literature with a special reference to preferential attachment of HBV to human hepatocytes. J. Viral Hepat. 1997, 4, 145–153. [Google Scholar] [CrossRef]
- Chen, J.; Wu, M.; Liu, K.; Zhang, W.; Li, Y.; Zhou, X.; Bai, L.; Yuan, Z. New insights into hepatitis B virus biology and implications for novel antiviral strategies. Natl. Sci. Rev. 2015, 2, 296–313. [Google Scholar] [CrossRef]
- Lamontagne, J.; Mell, J.C.; Bouchard, M.J. Transcriptome-Wide Analysis of Hepatitis B Virus-Mediated Changes to Normal Hepatocyte Gene Expression. PLoS Pathog. 2016, 12, e1005438. [Google Scholar] [CrossRef] [PubMed]
- Dupinay, T.; Gheit, T.; Roques, P.; Cova, L.; Chevallier-Queyron, P.; Tasahsu, S.I.; Le Grand, R.; Simon, F.; Cordier, G.; Wakrim, L.; et al. Discovery of naturally occurring transmissible chronic hepatitis B virus infection among Macaca fascicularis from Mauritius Island. Hepatology 2013, 58, 1610–1620. [Google Scholar] [CrossRef] [PubMed]
- Purcell, R.H.; London, W.T.; McAuliffe, V.J.; Palmer, A.E.; Kaplan, P.M.; Gerin, J.L.; Wagner, J.; Popper, H.; Lvovsky, E.; Wong, D.C.; et al. Modification of chronic hepatitis-B virus infection in chimpanzees by administration of an interferon inducer. Lancet 1976, 2, 757–761. [Google Scholar] [CrossRef]
- Burwitz, B.J.; Wettengel, J.M.; Muck-Hausl, M.A.; Ringelhan, M.; Ko, C.; Festag, M.M.; Hammond, K.B.; Northrup, M.; Bimber, B.N.; Jacob, T.; et al. Hepatocytic expression of human sodium-taurocholate cotransporting polypeptide enables hepatitis B virus infection of macaques. Nat. Commun. 2017, 8, 2146. [Google Scholar] [CrossRef] [PubMed]
- von Weizsacker, F.; Kock, J.; MacNelly, S.; Ren, S.; Blum, H.E.; Nassal, M. The tupaia model for the study of hepatitis B virus: Direct infection and HBV genome transduction of primary tupaia hepatocytes. Methods Mol. Med. 2004, 96, 153–161. [Google Scholar] [CrossRef]
- Guha, C.; Mohan, S.; Roy-Chowdhury, N.; Roy-Chowdhury, J. Cell culture and animal models of viral hepatitis. Part I: Hepatitis B. Lab. Anim. 2004, 33, 37–46. [Google Scholar] [CrossRef]
- Lampertico, P.; Malter, J.S.; Gerber, M.A. Development and application of an in vitro model for screening anti-hepatitis B virus therapeutics. Hepatology 1991, 13, 422–426. [Google Scholar]
- Witt-Kehati, D.; Bitton Alaluf, M.; Shlomai, A. Advances and Challenges in Studying Hepatitis B Virus In Vitro. Viruses 2016, 8, 21. [Google Scholar] [CrossRef]
- Hayes, C.N.; Chayama, K. HBV culture and infectious systems. Hepatol. Int. 2016, 10, 559–566. [Google Scholar] [CrossRef]
- Tuttleman, J.S.; Pugh, J.C.; Summers, J.W. In vitro experimental infection of primary duck hepatocyte cultures with duck hepatitis B virus. J. Virol. 1986, 58, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Tyler, G.V.; Snyder, R.L.; Summers, J. Experimental infection of the woodchuck (Marmota monax monax) with woodchuck hepatitis virus. Lab. Investig. 1986, 55, 51–55. [Google Scholar] [PubMed]
- Marion, P.L.; Oshiro, L.S.; Regnery, D.C.; Scullard, G.H.; Robinson, W.S. A virus in Beechey ground squirrels that is related to hepatitis B virus of humans. Proc. Natl. Acad. Sci. USA 1980, 77, 2941–2945. [Google Scholar] [CrossRef] [PubMed]
- Allweiss, L.; Dandri, M. Experimental in vitro and in vivo models for the study of human hepatitis B virus infection. J. Hepatol. 2016, 64, S17–S31. [Google Scholar] [CrossRef]
- Sandhu, P.; Haque, M.; Humphries-Bickley, T.; Ravi, S.; Song, J. Hepatitis B Virus Immunopathology, Model Systems, and Current Therapies. Front. Immunol. 2017, 8, 436. [Google Scholar] [CrossRef]
- Kock, J.; Nassal, M.; MacNelly, S.; Baumert, T.F.; Blum, H.E.; von Weizsacker, F. Efficient infection of primary tupaia hepatocytes with purified human and woolly monkey hepatitis B virus. J. Virol. 2001, 75, 5084–5089. [Google Scholar] [CrossRef]
- Meier, A.; Mehrle, S.; Weiss, T.S.; Mier, W.; Urban, S. Myristoylated PreS1-domain of the hepatitis B virus L-protein mediates specific binding to differentiated hepatocytes. Hepatology 2013, 58, 31–42. [Google Scholar] [CrossRef]
- Bouchard, M.J.; Navas-Martin, S. Hepatitis B and C virus hepatocarcinogenesis: Lessons learned and future challenges. Cancer Lett. 2011, 305, 123–143. [Google Scholar] [CrossRef]
- Gearhart, T.L.; Bouchard, M.J. The hepatitis B virus HBx protein modulates cell cycle regulatory proteins in cultured primary human hepatocytes. Virus Res. 2011, 155, 363–367. [Google Scholar] [CrossRef]
- Rawat, S.; Bouchard, M.J. The hepatitis B virus (HBV) HBx protein activates AKT to simultaneously regulate HBV replication and hepatocyte survival. J. Virol. 2015, 89, 999–1012. [Google Scholar] [CrossRef]
- Seeger, M.C.; Zoulim, F.; Mason, W.S. Hepadnaviruses. In Fields Virology; Knipe, D.M., Howley, P.M., Eds.; Wolters Kluwer/Lippincott Williams & Wilkins Health: Philadelphia, PA, USA, 2013; pp. 3376–3436. [Google Scholar]
- Galle, P.R.; Hagelstein, J.; Kommerell, B.; Volkmann, M.; Schranz, P.; Zentgraf, H. In vitro experimental infection of primary human hepatocytes with hepatitis B virus. Gastroenterology 1994, 106, 664–673. [Google Scholar] [CrossRef]
- Gripon, P.; Diot, C.; Theze, N.; Fourel, I.; Loreal, O.; Brechot, C.; Guguen-Guillouzo, C. Hepatitis B virus infection of adult human hepatocytes cultured in the presence of dimethyl sulfoxide. J. Virol. 1988, 62, 4136–4143. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.; Chen, Q.; Yang, L.Y.; Li, W.Y.; Cao, X.B.; Wu, J.R.; Peng, Y.P.; Chen, M.R. Hepatitis B virus infection and replication in primarily cultured human fetal hepatocytes. World J. Gastroenterol. 2007, 13, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Ochiya, T.; Tsurimoto, T.; Ueda, K.; Okubo, K.; Shiozawa, M.; Matsubara, K. An in vitro system for infection with hepatitis B virus that uses primary human fetal hepatocytes. Proc. Natl. Acad. Sci. USA 1989, 86, 1875–1879. [Google Scholar] [CrossRef]
- Tyakht, A.V.; Ilina, E.N.; Alexeev, D.G.; Ischenko, D.S.; Gorbachev, A.Y.; Semashko, T.A.; Larin, A.K.; Selezneva, O.V.; Kostryukova, E.S.; Karalkin, P.A.; et al. RNA-Seq gene expression profiling of HepG2 cells: The influence of experimental factors and comparison with liver tissue. BMC Genom. 2014, 15, 1108. [Google Scholar] [CrossRef]
- Belloni, L.; Allweiss, L.; Guerrieri, F.; Pediconi, N.; Volz, T.; Pollicino, T.; Petersen, J.; Raimondo, G.; Dandri, M.; Levrero, M. IFN-alpha inhibits HBV transcription and replication in cell culture and in humanized mice by targeting the epigenetic regulation of the nuclear cccDNA minichromosome. J. Clin. Investig. 2012, 122, 529–537. [Google Scholar] [CrossRef]
- Chang, C.M.; Jeng, K.S.; Hu, C.P.; Lo, S.J.; Su, T.S.; Ting, L.P.; Chou, C.K.; Han, S.H.; Pfaff, E.; Salfeld, J.; et al. Production of hepatitis B virus in vitro by transient expression of cloned HBV DNA in a hepatoma cell line. EMBO J. 1987, 6, 675–680. [Google Scholar] [CrossRef]
- Chen, R.Y.; Edwards, R.; Shaw, T.; Colledge, D.; Delaney, W.E.t.; Isom, H.; Bowden, S.; Desmond, P.; Locarnini, S.A. Effect of the G1896A precore mutation on drug sensitivity and replication yield of lamivudine-resistant HBV in vitro. Hepatology 2003, 37, 27–35. [Google Scholar] [CrossRef]
- Delaney, W.E.t.; Miller, T.G.; Isom, H.C. Use of the hepatitis B virus recombinant baculovirus-HepG2 system to study the effects of (-)-beta-2’,3’-dideoxy-3’-thiacytidine on replication of hepatitis B virus and accumulation of covalently closed circular DNA. Antimicrob. Agents Chemother. 1999, 43, 2017–2026. [Google Scholar] [CrossRef]
- Knowles, B.B.; Howe, C.C.; Aden, D.P. Human hepatocellular carcinoma cell lines secrete the major plasma proteins and hepatitis B surface antigen. Science 1980, 209, 497–499. [Google Scholar] [CrossRef]
- Levrero, M.; Pollicino, T.; Petersen, J.; Belloni, L.; Raimondo, G.; Dandri, M. Control of cccDNA function in hepatitis B virus infection. J. Hepatol. 2009, 51, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Nakabayashi, H.; Taketa, K.; Miyano, K.; Yamane, T.; Sato, J. Growth of human hepatoma cells lines with differentiated functions in chemically defined medium. Cancer Res. 1982, 42, 3858–3863. [Google Scholar] [PubMed]
- Pollicino, T.; Belloni, L.; Raffa, G.; Pediconi, N.; Squadrito, G.; Raimondo, G.; Levrero, M. Hepatitis B virus replication is regulated by the acetylation status of hepatitis B virus cccDNA-bound H3 and H4 histones. Gastroenterology 2006, 130, 823–837. [Google Scholar] [CrossRef] [PubMed]
- Seeger, C.; Mason, W.S. Sodium-dependent taurocholic cotransporting polypeptide: A candidate receptor for human hepatitis B virus. Gut 2013, 62, 1093–1095. [Google Scholar] [CrossRef]
- Sureau, C.; Eichberg, J.W.; Hubbard, G.B.; Romet-Lemonne, J.L.; Essex, M. A molecularly cloned hepatitis B virus produced in vitro is infectious in a chimpanzee. J. Virol. 1988, 62, 3064–3067. [Google Scholar] [CrossRef]
- Yaginuma, K.; Shirakata, Y.; Kobayashi, M.; Koike, K. Hepatitis B virus (HBV) particles are produced in a cell culture system by transient expression of transfected HBV DNA. Proc. Natl. Acad. Sci. USA 1987, 84, 2678–2682. [Google Scholar] [CrossRef]
- Ding, X.R.; Yang, J.; Sun, D.C.; Lou, S.K.; Wang, S.Q. Whole genome expression profiling of hepatitis B virus-transfected cell line reveals the potential targets of anti-HBV drugs. Pharm. J. 2008, 8, 61–70. [Google Scholar] [CrossRef][Green Version]
- Gripon, P.; Cannie, I.; Urban, S. Efficient inhibition of hepatitis B virus infection by acylated peptides derived from the large viral surface protein. J. Virol. 2005, 79, 1613–1622. [Google Scholar] [CrossRef]
- Gripon, P.; Rumin, S.; Urban, S.; Le Seyec, J.; Glaise, D.; Cannie, I.; Guyomard, C.; Lucas, J.; Trepo, C.; Guguen-Guillouzo, C. Infection of a human hepatoma cell line by hepatitis B virus. Proc. Natl. Acad. Sci. USA 2002, 99, 15655–15660. [Google Scholar] [CrossRef]
- Guo, H.; Jiang, D.; Zhou, T.; Cuconati, A.; Block, T.M.; Guo, J.T. Characterization of the intracellular deproteinized relaxed circular DNA of hepatitis B virus: An intermediate of covalently closed circular DNA formation. J. Virol. 2007, 81, 12472–12484. [Google Scholar] [CrossRef]
- Hantz, O.; Parent, R.; Durantel, D.; Gripon, P.; Guguen-Guillouzo, C.; Zoulim, F. Persistence of the hepatitis B virus covalently closed circular DNA in HepaRG human hepatocyte-like cells. J. Gen. Virol. 2009, 90, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Ladner, S.K.; Otto, M.J.; Barker, C.S.; Zaifert, K.; Wang, G.H.; Guo, J.T.; Seeger, C.; King, R.W. Inducible expression of human hepatitis B virus (HBV) in stably transfected hepatoblastoma cells: A novel system for screening potential inhibitors of HBV replication. Antimicrob. Agents Chemother. 1997, 41, 1715–1720. [Google Scholar] [CrossRef] [PubMed]
- Li, G.Q.; Xu, W.Z.; Wang, J.X.; Deng, W.W.; Li, D.; Gu, H.X. Combination of small interfering RNA and lamivudine on inhibition of human B virus replication in HepG2.2.15 cells. World J. Gastroenterol. 2007, 13, 2324–2327. [Google Scholar] [CrossRef] [PubMed]
- Luangsay, S.; Gruffaz, M.; Isorce, N.; Testoni, B.; Michelet, M.; Faure-Dupuy, S.; Maadadi, S.; Ait-Goughoulte, M.; Parent, R.; Rivoire, M.; et al. Early inhibition of hepatocyte innate responses by hepatitis B virus. J. Hepatol. 2015, 63, 1314–1322. [Google Scholar] [CrossRef] [PubMed]
- Lucifora, J.; Durantel, D.; Testoni, B.; Hantz, O.; Levrero, M.; Zoulim, F. Control of hepatitis B virus replication by innate response of HepaRG cells. Hepatology 2010, 51, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Lucifora, J.; Xia, Y.; Reisinger, F.; Zhang, K.; Stadler, D.; Cheng, X.; Sprinzl, M.F.; Koppensteiner, H.; Makowska, Z.; Volz, T.; et al. Specific and nonhepatotoxic degradation of nuclear hepatitis B virus cccDNA. Science 2014, 343, 1221–1228. [Google Scholar] [CrossRef]
- Sun, D.; Nassal, M. Stable HepG2- and Huh7-based human hepatoma cell lines for efficient regulated expression of infectious hepatitis B virus. J. Hepatol. 2006, 45, 636–645. [Google Scholar] [CrossRef]
- Tsurimoto, T.; Fujiyama, A.; Matsubara, K. Stable expression and replication of hepatitis B virus genome in an integrated state in a human hepatoma cell line transfected with the cloned viral DNA. Proc. Natl. Acad. Sci. USA 1987, 84, 444–448. [Google Scholar] [CrossRef]
- Bouezzedine, F.; Fardel, O.; Gripon, P. Interleukin 6 inhibits HBV entry through NTCP down regulation. Virology 2015, 481, 34–42. [Google Scholar] [CrossRef]
- Liu, Y.; Li, J.; Chen, J.; Li, Y.; Wang, W.; Du, X.; Song, W.; Zhang, W.; Lin, L.; Yuan, Z. Hepatitis B virus polymerase disrupts K63-linked ubiquitination of STING to block innate cytosolic DNA-sensing pathways. J. Virol. 2015, 89, 2287–2300. [Google Scholar] [CrossRef]
- Hoh, A.; Heeg, M.; Ni, Y.; Schuch, A.; Binder, B.; Hennecke, N.; Blum, H.E.; Nassal, M.; Protzer, U.; Hofmann, M.; et al. Hepatitis B Virus-Infected HepG2hNTCP Cells Serve as a Novel Immunological Tool To Analyze the Antiviral Efficacy of CD8+ T Cells In Vitro. J. Virol. 2015, 89, 7433–7438. [Google Scholar] [CrossRef] [PubMed]
- Yan, R.; Zhang, Y.; Cai, D.; Liu, Y.; Cuconati, A.; Guo, H. Spinoculation Enhances HBV Infection in NTCP-Reconstituted Hepatocytes. PLoS ONE 2015, 10, e0129889. [Google Scholar] [CrossRef] [PubMed]
- Thomas, E.; Liang, T.J. Experimental models of hepatitis B and C—New insights and progress. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 362–374. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Carpentier, A.; Cheng, X.; Block, P.D.; Zhao, Y.; Zhang, Z.; Protzer, U.; Liang, T.J. Human stem cell-derived hepatocytes as a model for hepatitis B virus infection, spreading and virus-host interactions. J. Hepatol. 2017, 66, 494–503. [Google Scholar] [CrossRef]
- Sakurai, F.; Mitani, S.; Yamamoto, T.; Takayama, K.; Tachibana, M.; Watashi, K.; Wakita, T.; Iijima, S.; Tanaka, Y.; Mizuguchi, H. Human induced-pluripotent stem cell-derived hepatocyte-like cells as an in vitro model of human hepatitis B virus infection. Sci. Rep. 2017, 7, 45698. [Google Scholar] [CrossRef]
- Schwartz, R.E.; Fleming, H.E.; Khetani, S.R.; Bhatia, S.N. Pluripotent stem cell-derived hepatocyte-like cells. Biotechnol. Adv. 2014, 32, 504–513. [Google Scholar] [CrossRef]
- Shlomai, A.; Schwartz, R.E.; Ramanan, V.; Bhatta, A.; de Jong, Y.P.; Bhatia, S.N.; Rice, C.M. Modeling host interactions with hepatitis B virus using primary and induced pluripotent stem cell-derived hepatocellular systems. Proc. Natl. Acad. Sci. USA 2014, 111, 12193–12198. [Google Scholar] [CrossRef]
- Nie, Y.Z.; Zheng, Y.W.; Miyakawa, K.; Murata, S.; Zhang, R.R.; Sekine, K.; Ueno, Y.; Takebe, T.; Wakita, T.; Ryo, A.; et al. Recapitulation of hepatitis B virus-host interactions in liver organoids from human induced pluripotent stem cells. EBioMedicine 2018, 35, 114–123. [Google Scholar] [CrossRef]
- Sells, M.A.; Chen, M.L.; Acs, G. Production of hepatitis B virus particles in Hep G2 cells transfected with cloned hepatitis B virus DNA. Proc. Natl. Acad. Sci. USA 1987, 84, 1005–1009. [Google Scholar] [CrossRef]
- Brandon, E.F.; Raap, C.D.; Meijerman, I.; Beijnen, J.H.; Schellens, J.H. An update on in vitro test methods in human hepatic drug biotransformation research: Pros and cons. Toxicol. Appl. Pharmacol. 2003, 189, 233–246. [Google Scholar] [CrossRef]
- Choi, S.; Sainz, B., Jr.; Corcoran, P.; Uprichard, S.; Jeong, H. Characterization of increased drug metabolism activity in dimethyl sulfoxide (DMSO)-treated Huh7 hepatoma cells. Xenobiotica 2009, 39, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Clayton, R.F.; Rinaldi, A.; Kandyba, E.E.; Edward, M.; Willberg, C.; Klenerman, P.; Patel, A.H. Liver cell lines for the study of hepatocyte functions and immunological response. Liver Int. 2005, 25, 389–402. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Chen, P.; Hou, X.; Xu, W.; Wang, D.; Wang, T.Y.; Zhang, L.; Zheng, G.; Gao, Z.L.; He, C.Y.; et al. The recombined cccDNA produced using minicircle technology mimicked HBV genome in structure and function closely. Sci. Rep. 2016, 6, 25552. [Google Scholar] [CrossRef] [PubMed]
- Delaney, W.E.t.; Isom, H.C. Hepatitis B virus replication in human HepG2 cells mediated by hepatitis B virus recombinant baculovirus. Hepatology 1998, 28, 1134–1146. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Li, J.; Yue, L.; Bai, L.; Li, Y.; Chen, J.; Zhang, X.; Yuan, Z. Establishment of Cre-mediated HBV recombinant cccDNA (rcccDNA) cell line for cccDNA biology and antiviral screening assays. Antivir. Res. 2018, 152, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Araki, K.; Miyazaki, J.; Hino, O.; Tomita, N.; Chisaka, O.; Matsubara, K.; Yamamura, K. Expression and replication of hepatitis B virus genome in transgenic mice. Proc. Natl. Acad. Sci. USA 1989, 86, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Wang, A.H.; Shao, L.H.; Wang, G.; Yao, Y.Y.; Sai, L.T.; Chen, F.Z.; Zheng, F.; Li, Y.; Ma, L.X. A new cell culture system for infection with hepatitis B virus that fuses HepG2 cells with primary human hepatocytes. J. Int. Med. Res. 2009, 37, 650–661. [Google Scholar] [CrossRef]
- Li, G.; Zhu, Y.; Shao, D.; Chang, H.; Zhang, X.; Zhou, D.; Gao, Y.; Lan, K.; Deng, Q. Recombinant covalently closed circular DNA of hepatitis B virus induces long-term viral persistence with chronic hepatitis in a mouse model. Hepatology 2018, 67, 56–70. [Google Scholar] [CrossRef]
- Cai, D.; Mills, C.; Yu, W.; Yan, R.; Aldrich, C.E.; Saputelli, J.R.; Mason, W.S.; Xu, X.; Guo, J.T.; Block, T.M.; et al. Identification of disubstituted sulfonamide compounds as specific inhibitors of hepatitis B virus covalently closed circular DNA formation. Antimicrob. Agents Chemother. 2012, 56, 4277–4288. [Google Scholar] [CrossRef]
- Cai, D.; Wang, X.; Yan, R.; Mao, R.; Liu, Y.; Ji, C.; Cuconati, A.; Guo, H. Establishment of an inducible HBV stable cell line that expresses cccDNA-dependent epitope-tagged HBeAg for screening of cccDNA modulators. Antivir. Res. 2016, 132, 26–37. [Google Scholar] [CrossRef]
- Gripon, P.; Diot, C.; Guguen-Guillouzo, C. Reproducible high level infection of cultured adult human hepatocytes by hepatitis B virus: Effect of polyethylene glycol on adsorption and penetration. Virology 1993, 192, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Runge, D.; Runge, D.M.; Jager, D.; Lubecki, K.A.; Beer Stolz, D.; Karathanasis, S.; Kietzmann, T.; Strom, S.C.; Jungermann, K.; Fleig, W.E.; et al. Serum-free, long-term cultures of human hepatocytes: Maintenance of cell morphology, transcription factors, and liver-specific functions. Biochem. Biophys. Res. Commun. 2000, 269, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Runge, D.M.; Runge, D.; Dorko, K.; Pisarov, L.A.; Leckel, K.; Kostrubsky, V.E.; Thomas, D.; Strom, S.C.; Michalopoulos, G.K. Epidermal growth factor- and hepatocyte growth factor-receptor activity in serum-free cultures of human hepatocytes. J. Hepatol. 1999, 30, 265–274. [Google Scholar] [CrossRef]
- Guillouzo, A.; Corlu, A.; Aninat, C.; Glaise, D.; Morel, F.; Guguen-Guillouzo, C. The human hepatoma HepaRG cells: A highly differentiated model for studies of liver metabolism and toxicity of xenobiotics. Chem. Biol. Interact. 2007, 168, 66–73. [Google Scholar] [CrossRef]
- Kanebratt, K.P.; Andersson, T.B. Evaluation of HepaRG cells as an in vitro model for human drug metabolism studies. Drug Metab. Dispos. 2008, 36, 1444–1452. [Google Scholar] [CrossRef]
- Kanebratt, K.P.; Andersson, T.B. HepaRG cells as an in vitro model for evaluation of cytochrome P450 induction in humans. Drug Metab. Dispos. 2008, 36, 137–145. [Google Scholar] [CrossRef]
- Krepstakies, M.; Lucifora, J.; Nagel, C.H.; Zeisel, M.B.; Holstermann, B.; Hohenberg, H.; Kowalski, I.; Gutsmann, T.; Baumert, T.F.; Brandenburg, K.; et al. A new class of synthetic peptide inhibitors blocks attachment and entry of human pathogenic viruses. J. Infect. Dis. 2012, 205, 1654–1664. [Google Scholar] [CrossRef]
- Ni, Y.; Lempp, F.A.; Mehrle, S.; Nkongolo, S.; Kaufman, C.; Falth, M.; Stindt, J.; Koniger, C.; Nassal, M.; Kubitz, R.; et al. Hepatitis B and D Viruses Exploit Sodium Taurocholate Co-transporting Polypeptide for Species-Specific Entry into Hepatocytes. Gastroenterology 2014, 146, 1070–1083. [Google Scholar] [CrossRef]
- Schulze, A.; Gripon, P.; Urban, S. Hepatitis B virus infection initiates with a large surface protein-dependent binding to heparan sulfate proteoglycans. Hepatology 2007, 46, 1759–1768. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Liu, Y.; Sui, J.; Li, W. NTCP opens the door for hepatitis B virus infection. Antivir. Res. 2015, 121, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Michailidis, E.; Pabon, J.; Xiang, K.; Park, P.; Ramanan, V.; Hoffmann, H.H.; Schneider, W.M.; Bhatia, S.N.; de Jong, Y.P.; Shlomai, A.; et al. A robust cell culture system supporting the complete life cycle of hepatitis B virus. Sci. Rep. 2017, 7, 16616. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Si-Tayeb, K.; Noto, F.K.; Nagaoka, M.; Li, J.; Battle, M.A.; Duris, C.; North, P.E.; Dalton, S.; Duncan, S.A. Highly efficient generation of human hepatocyte-like cells from induced pluripotent stem cells. Hepatology 2010, 51, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Khetani, S.R.; Bhatia, S.N. Microscale culture of human liver cells for drug development. Nat. Biotechnol. 2008, 26, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Khetani, S.R. Micropatterned Co-Cultures of Human Hepatocytes and Stromal Cells for the Assessment of Drug Clearance and Drug-Drug Interactions. Curr. Protoc. Toxicol. 2017, 72, 14–17. [Google Scholar] [CrossRef] [PubMed]
- Gural, N.; Mancio-Silva, L.; He, J.; Bhatia, S.N. Engineered Livers for Infectious Diseases. Cell. Mol. Gastroenter. 2018, 5, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Prior, N.; Inacio, P.; Huch, M. Liver organoids: From basic research to therapeutic applications. Gut 2019, 68, 2228–2237. [Google Scholar] [CrossRef]
- Gunther, S.; Li, B.C.; Miska, S.; Kruger, D.H.; Meisel, H.; Will, H. A novel method for efficient amplification of whole hepatitis B virus genomes permits rapid functional analysis and reveals deletion mutants in immunosuppressed patients. J. Virol. 1995, 69, 5437–5444. [Google Scholar] [CrossRef]
- Ren, S.; Nassal, M. Hepatitis B virus (HBV) virion and covalently closed circular DNA formation in primary tupaia hepatocytes and human hepatoma cell lines upon HBV genome transduction with replication-defective adenovirus vectors. J. Virol. 2001, 75, 1104–1116. [Google Scholar] [CrossRef]
- Sprinzl, M.F.; Oberwinkler, H.; Schaller, H.; Protzer, U. Transfer of hepatitis B virus genome by adenovirus vectors into cultured cells and mice: Crossing the species barrier. J. Virol. 2001, 75, 5108–5118. [Google Scholar] [CrossRef]
- Sugiyama, M.; Tanaka, Y.; Kato, T.; Orito, E.; Ito, K.; Acharya, S.K.; Gish, R.G.; Kramvis, A.; Shimada, T.; Izumi, N.; et al. Influence of hepatitis B virus genotypes on the intra- and extracellular expression of viral DNA and antigens. Hepatology 2006, 44, 915–924. [Google Scholar] [CrossRef]
- Lucifora, J.; Durantel, D.; Belloni, L.; Barraud, L.; Villet, S.; Vincent, I.E.; Margeridon-Thermet, S.; Hantz, O.; Kay, A.; Levrero, M.; et al. Initiation of hepatitis B virus genome replication and production of infectious virus following delivery in HepG2 cells by novel recombinant baculovirus vector. J. Gen. Virol 2008, 89, 1819–1828. [Google Scholar] [CrossRef] [PubMed]
- Clippinger, A.J.; Gearhart, T.L.; Bouchard, M.J. Hepatitis B virus X protein modulates apoptosis in primary rat hepatocytes by regulating both NF-κB and the mitochondrial permeability transition pore. J. Virol. 2009, 83, 4718–4731. [Google Scholar] [CrossRef] [PubMed]
- Inoue, J.; Ueno, Y.; Nagasaki, F.; Wakui, Y.; Kondo, Y.; Fukushima, K.; Niitsuma, H.; Shimosegawa, T. Enhanced intracellular retention of a hepatitis B virus strain associated with fulminant hepatitis. Virology 2009, 395, 202–209. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, Y.; Ye, S.; Wang, T.; Zhao, R.; Chen, F.; Abe, K.; Jin, X. The response to interferon is influenced by hepatitis B virus genotype in vitro and in vivo. Virus Res. 2013, 171, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Bhoola, N.H.; Reumann, K.; Kew, M.C.; Will, H.; Kramvis, A. Construction of replication competent plasmids of hepatitis B virus subgenotypes A1, A2 and D3 with authentic endogenous promoters. J. Virol. Methods 2014, 203, 54–64. [Google Scholar] [CrossRef]
- Zhang, Z.; Xia, J.; Sun, B.; Dai, Y.; Li, X.; Schlaak, J.F.; Lu, M. In vitro and in vivo replication of a chemically synthesized consensus genome of hepatitis B virus genotype B. J. Virol. Methods 2015, 213, 57–64. [Google Scholar] [CrossRef]
- Sozzi, V.; Walsh, R.; Littlejohn, M.; Colledge, D.; Jackson, K.; Warner, N.; Yuen, L.; Locarnini, S.A.; Revill, P.A. In Vitro Studies Show that Sequence Variability Contributes to Marked Variation in Hepatitis B Virus Replication, Protein Expression, and Function Observed across Genotypes. J. Virol. 2016, 90, 10054–10064. [Google Scholar] [CrossRef]
- Berke, J.M.; Tan, Y.; Verbinnen, T.; Dehertogh, P.; Vergauwen, K.; Vos, A.; Lenz, O.; Pauwels, F. Antiviral profiling of the capsid assembly modulator BAY41-4109 on full-length HBV genotype A-H clinical isolates and core site-directed mutants in vitro. Antivir. Res. 2017, 144, 205–215. [Google Scholar] [CrossRef]
- Jia, H.; Qin, Y.; Chen, C.; Zhang, F.; Li, C.; Zong, L.; Wang, Y.; Zhang, J.; Li, J.; Wen, Y.; et al. The Envelope Gene of Hepatitis B Virus Is Implicated in Both Differential Virion Secretion and Genome Replication Capacities between Genotype B and Genotype C Isolates. Viruses 2017, 9, 62. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, M.; Gong, M.; Xu, Y.; Xie, C.; Deng, H.; Li, X.; Wu, H.; Wang, Z. Inhibition of hepatitis B virus replication via HBV DNA cleavage by Cas9 from Staphylococcus aureus. Antivir. Res. 2018, 152, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Bannister, E.; Sozzi, V.; Mason, H.; Locarnini, S.A.; Hardiker, W.; Revill, P.A. Analysis of the in vitro replication phenotype of African hepatitis B virus (HBV) genotypes and subgenotypes present in Australia identifies marked differences in DNA and protein expression. Virology 2019, 54, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Kostyushev, D.; Brezgin, S.; Kostyusheva, A.; Zarifyan, D.; Goptar, I.; Chulanov, V. Orthologous CRISPR/Cas9 systems for specific and efficient degradation of covalently closed circular DNA of hepatitis B virus. Cell Mol. Life Sci. 2019, 76, 1779–1794. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, E.M.; Kornepati, A.V.; Cullen, B.R. Targeting hepatitis B virus cccDNA using CRISPR/Cas9. Antivir. Res. 2015, 123, 188–192. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Hasegawa, I.; Kato, T.; Orito, E.; Hirashima, N.; Acharya, S.K.; Gish, R.G.; Kramvis, A.; Kew, M.C.; Yoshihara, N.; et al. A case-control study for differences among hepatitis B virus infections of genotypes A (subtypes Aa and Ae) and D. Hepatology 2004, 40, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Kramvis, A.; Kew, M.C. Molecular characterization of subgenotype A1 (subgroup Aa) of hepatitis B virus. Hepatol. Res. 2007, 37, S27–S32. [Google Scholar] [CrossRef]
- Kramvis, A. Molecular characteristics and clinical relevance of African genotypes and subgenotypes of hepatitis B virus. S. Afr. Med. J. 2018, 108, 17–21. [Google Scholar] [CrossRef]
- Kramvis, A.; Paraskevis, D. Subgenotype A1 of HBV—Tracing human migrations in and out of Africa. Antivir. Ther. 2013, 18, 513–521. [Google Scholar] [CrossRef]
- The NIH Hepatitis B Cure Strategic Plan Working Group. Strategic Plan for Trans-NIH Research to Cure Hepatitis B; Department of Health and Human Service—USA: Washington, DC, USA, 2019. [Google Scholar]
- ICE-HBV. HBV Research Protocols. Available online: https://ice-hbv.org/protocol/ (accessed on 2 March 2020).



{kind=link}
{kind=link}
| Model System | Advantages | Disadvantages | Studies | Reference | |
|---|---|---|---|---|---|
| Animal Model Systems | Primary Tupaia hepatocytes (PTH) | Can be infected with HBV | Low HBV infection efficiency; Lack of genetically uniform Tupaia belangeri strains High cost | Identification of the sodium taurocholate co-transporting polypeptide (NTCP) as the receptor for HBV infection; Studying cccDNA formation | [12,46,47,48] |
| Primary hepatocytes derived from small animal models such as rats and mice | Can support HBV replication; Intact innate immune response | Cannot be infected with HBV and requires bypassing the initial receptor-mediated infection of the cell by direct transfection or transduction of the HBV DNA genome; cccDNA is not formed in mouse cells. | Studying HBV replication from the post-entry stages; Studying the effects of HBV replication and HBV proteins on cellular physiology | [33,49,50,51,52] | |
| Human Model Systems | Primary hepatocytes derived from macaques transduced with NTCP | Can support HBV infection | Need to be transduced with NTCP | Macaque primary hepatocytes transduced with NTCP were susceptible to HBV, whereas untransduced cells could not be infected | [36] |
| Primary human hepatocytes (PHH) | Ideal and gold standard in vitro model system that can be infected with HBV | Limited availability and lifespan; Loss of hepatocyte function and susceptibility to HBV infection within days of isolation and culture; Unpredictable variability between hepatocyte donors | Studying HBV infection; Studying innate immune response to HBV infection; Studying metabolism and drug toxicity | [32,45,46,49,52,53,54] | |
| Human fetal hepatocytes | Can be infected with HBV | Limited availability; Limited infection efficiency and apparent absence of viral spreading; Unpredictable variability between hepatocyte donors; Ethical considerations | Studying HBV infection | [55,56] | |
| Human Model Systems | Transiently transfected or transduced immortalized tumor-derived or transformed liver cell lines (e.g., Huh7, HepG2) | Can support HBV replication and transcription; Convenient system; Minimal variability; Relatively cheap | Cannot be infected with HBV and requires bypassing the initial receptor-mediated infection of the cell by direct transfection or transduction of the HBV DNA genome; Cellular signaling pathways are significantly altered and therefore do not recapitulate the physiology of normal hepatocytes; Low to minimal cell-to-cell spread; Gene expression in tumor-derived or transformed cell lines differs from that in normal hepatocytes [57] | Molecular characterization of HBV; Studying HBV replication and regulation and comparing replication of (sub)genotypes; Testing efficiency of novel anti-HBV drugs; Drug resistance studies | [33,40,58,59,60,61,62,63,64,65,66,67,68] |
| Stably transfected immortalized tumor-derived or transformed liver cell lines, (e.g., HepaRG, HepAD38, HepDE19, HepG2.2.15) | Can support HBV replication and transcription; Good source of virions for infection; HepaRG cell line has morphological and functional features similar to that of PHHs | Cannot be infected with HBV and requires bypassing the initial receptor-mediated infection of the cell by direct transfection or transduction of the HBV DNA genome; HBV expressed from integrated HBV DNA genome and not cccDNA in the case of HepG2.2.15 and AD38 cells; Requires the addition of dimethyl sulfoxide (DMSO) to promote differentiation (HepaRG) and supplementation of PEG to promote viral entry in the case of HepaRG | Molecular characterization of HBV; Studying virus–host interactions Studying HBV replication and regulation; Testing efficiency of novel anti-HBV drugs; Studies addressing the role of the innate immune response in counteracting HBV infection; Testing efficiency of novel anti-HBV drugs | [40,69,70,71,72,73,74,75,76,77,78,79,80,81,82] | |
| NTCP-expressing hepatoma cell lines | Can be infected with HBV allowing initial stages of infection to be studied; 50% of cells can express HBV versus only 7% in HepaRG; Easy to handle | Requires high multiplicity of infection and addition of PEG for successful infection; Infection is short-lived; Reduced viral spread and cccDNA levels; Majority of studies using cell culture derived HBV have been restricted to genotype D. | Studying the initial stages of HBV infection; Testing antiviral drug efficacy, especially entry inhibitors | [40,52,83,84] | |
| Human Model Systems | Hepatocyte-like cells (HLCs/iHeps) derived from pluripotent stem cells (iPSCs) | Reliable source that can be differentiated into mature hepatocytes; Supply unlimited and renewable; Less variable than PHHs; Can be established from different donors with and without HBV infection or liver disease | Expensive system to set up and high degree of expertise is required; Low hepatic function; Unpredictable variability between donors; Certain signaling pathways may be impaired; Inhibition of the innate immune response is required for HBV infection | Studying the host factors essential for HBV infection and replication; Comparison of infection to other in vitro models; Testing of antiviral agents | [12,40,85,86,87,88,89] |
| Micropatterned co-cultured cells | Maintains hepatocytic function over weeks after plating; Supports HBV infection; Active innate immune response; Renewable; Minitiarized | Unpredictable variability between hepatocyte donors; Low infection efficiency and apparent absence of viral spreading; Inhibition of the innate immune response is required for infection to occur; HBV viral particles collected from the model system is not infectious; Logistically and technically challenging | Comparison of infection to other in vitro models; Studying drug toxicity and drug interactions; Testing of antiviral agents | [40,89] | |
| Liver organoids from human induced pluripotent stem cells (iPSCs) | Cells differentiate with strong hepatic function; More susceptible to HBV infection when compared to HLCs derived iPSCs; Prolonged propagation of HBV for up to 20 days; Generation of infectious virions; Recapitulates virus-induced liver dysfunction | Highly sophisticated and labor-intensive system to establish; Some hepatic characteristics may differ from adult hepatocytes | Studying virus–host interactions; Has the potential to be used to study personalized hepatitis treatment | [90] | |
| (Sub)Genotype/Serotype of HBV | Model System Based on Different Cell Lines | Source of Viral Particles | Studies | Reference/ Year |
|---|---|---|---|---|
| Serotype ayw | Huh6 Huh7 HepG2 | 2.1 mer HBV in psV08 | Comparison of HBV transfection into different cell lines | [59]/1987 |
| Serotype adr | Huh7 Huh2.2 Primary human lens epithelial cells (HLEC1) | 2.0 mer HBV in pBR322 1.3 mer HBV in pBR322 | Comparison of HBV transfection into different cell lines | [68]/1987 |
| Seroype ayw | Huh7 | 1.0 mer HBV without a vector 1.0 mer HBV in pro-melanin concentrating hormone (pMCH) vector 2.0 mer HBV in pSM2 vector | Functional characterization of HBV | [120]/1995 |
| Sub-genotype D3 | HepG2 | 1.3 mer HBV in a baculovirus vector (Bac-HBV) pBlueBac4.5 | Molecular characterization of HBV | [96]/1998 |
| Serotype ayw | Primary tupaia hepatocytes (PTH) Huh7 HepG2 | 1.3 mer in an adenovirus vector (Ad-HBV) pTG9530 | Comparison of HBV transduction/infection into different in vitro model systems | [121]/2001 |
| Serotype ayw | PTH Rat Chicken Duck Primary human hepatocytes (PHH) HepG2 HEK293 | 1.3 mer HBV in an adenovirus vector (Ad-HBV) pAdTrack | Comparison of infection efficiency of HBV between different in vitro models | [122]/2001 |
| Genotype A (Serotype adw) Genotype D (Serotype awy) | HepG2 | 1.0 mer HBV in pUC19 | Regulation of HBV minichromosome | [65]/2006 |
| Huh7 | ||||
| Sub-genotype A1, A2, B1, B2 Genotypes C and D | Huh7 | 1.24 mer HBV in pGEM-T Easy | Functional characterization of HBV genotypes | [123]/2006 |
| Genotype D (Serotype ayw) | HepG2 | 1.1 mer Bac-HBV pTriEx 1.3 mer Bac-HBV pTriEx | Functional characterization of HBV | [124]/2008 |
| Genotype D (Serotype ayw) | HepaRG | Supernatant of 1.1 mer Bac-HBV Supernatant of HepG2.2.15 | Functional characterization of HBV | [124]/2008 |
| Genotype D (Serotype ayw3) | Rat | 1.2 mer HBV + 0.1 mer HBx under the control of simian virus 40 early promoter | Studying the effects of HBx on cellular physiology | [125]/2009 |
| Sub-genotype B2 | HepG2 | 1.3 mer HBV in pUC118 vector (Endogenous promoter) | Molecular characterization of HBV mutations | [126]/2009 |
| Genotypes B and D | PTH PHH HEK293 HEK293T Hela HepG2 Huh7 SMCC-7721 BEL-7404 | Plasma from a chronic HBV carrier Supernatant of 1.05 mer Ad-HBV and under the control of the cytomegalovirus (CMV) promoter transfected in Huh7 cells (Exogenous promoter) | Identification of NTCP as the receptor for HBV infection | [12]/2012 |
| Sub-genotype A2, Sub-genotype B1 Sub-genotype C2 Sub-genotype D2 Sub-genotype I1 | HepG2 | 1.1 mer Hybrid HBV DNA (1.0 mer sub-genotype A2 HBV isolate + 0.1 mer Serotype adw2 HBV) in pUC19 vector 1.1 mer Hybrid HBV DNA (1.0 mer sub-genotype B1 + 0.1 mer serotype adw2 HBV) in pUC19 vector 1.1 mer Hybrid HBV DNA (1.0 mer sub-genotype C2 + 0.1 mer serotype adw2 HBV) in pUC19 vector 1.1 mer Hybrid HBV DNA (1.0 mer sub-genotype D2 + 0.1 mer serotype adw2 HBV) in pUC19 vector 1.1 mer Hybrid HBV DNA (1.0 mer sub-genotype I1 + 0.1 mer serotype adw2 HBV) in pUC19 vector | Testing of drug efficacy for various genotypes of HBV | [127]/2013 |
| Huh7 | ||||
| Sub-genotype A1, A2, D3 | Huh7 | 1.28 mer HBV DNA in pCDNA vector with cytomegalovirus (CMV) promoter removed (endogenous promoter) | Molecular characterization of HBV (sub)genotypes | [128]/2014 |
| Genotype A Genotype D | Micropatterned coculture (MPCC) iPSCs-iHeps | Plasma from patients | Comparison of infection efficiencies with different in vitro model systems | [89]/2014 |
| Genotype B | Huh7 | 1.3 mer HBV DNA in pBluescript KS (+) vector (pHBV1.3B) | Molecular characterization of genotype B | [129]/2015 |
| Sub-genotype A2, B2, C2, D3 Genotype J | Huh7 | 1.3 mer HBV DNA in pUC57 vector (Endogenous promoter) | Molecular characterization of HBV (sub)genotypes | [130]/2016 |
| HepG2 | ||||
| Genotypes A, B, C, D, E, F, G, H | HepG2 | 1.1 mer in pCDNA-3.1 vector (Exogenous promoter) 1.3 mer in pCDNA-3.1 vector (-CMV) (Endogenous promoter) | Testing of drug efficacy for various genotypes of HBV | [131]/2017 |
| HepG2-TA2-7 | ||||
| HepG2.117 | ||||
| Genotypes B and C | Huh7 | 1.1 mer HBV DNA in pCDNA3.1 zeo (−) vector (Exogenous promoter) | Functional characterization of HBV proteins | [132]/2017 |
| Genotypes B, C and D | Huh7 transfected with replication-competent plasmids or cccDNA HepG.2.15 (genotype D) HepAD38 (genotype D) | 1.2 mer HBV DNA in pUC19 (pHBV-1.3B, pHBV-1.3C) [128], genotype D and pAAV/HBV1.2 rcccDNA system, including prcccDNA and pCMV-Cre | Examination of Staphylococcus aureus clustered regularly interspaced short palindromic repeats (CRISPR)-associated (Cas) system (SaCas9) on HBV replication in transfected and stably transfected cell lines | [133]/2018 * |
| Sub-genotype A1 (Serotype adw2) Sub-genotype D2 (Serotype ayw3) Sub-genotype D6 (Serotype ayw2) Genotype E (Serotype ayw4) | HepG2 | 1.3 mer HBV DNA in pUC57 (Endogenous promoter) | Molecular characterization of HBV (sub)genotypes | [134]/2019 |
| Huh7 | ||||
| Genotype D | HepG2 HepG2-1.5merHBV HeptG2-1.1merHBV | 1.1 mer HBV DNA rcccDNA (+/− methylation) | Comparison of anti-HBV activity of 4 orthologous CRISPR/Cas9 systems | [135]/2019 * |
| Genotypes A and D | iPSC derived HLCs and MPCCs | Infection with three stocks of plasma derived from three different donors. Two stocks were genotype D, the other genotype A | Modelling of HBV-host interactions and anti-HBV drug testing of entecavir and interferon-β (IFN-β) | [89]/2014 |
| Genotypes C and D | iPPSC derived HLCs | Fiber-modified adenovirus (Ad) vector containing genotype C (Ad-HBV: AdK7-gLuc-HBV) | Transduction of iPS-HLCs with HBV and comparison to expression in PHHs and HepG2-NTCP-C4 cells. Testing of antiviral agents entecavir and myrcludex | [87]/2017 |
| Genotypes C and D | iPPSC derived HLCs | Genotype D derived from the culture supernatant of HepG2.2.15.7 cells | Infection of iPS-HLCs with HBV and comparison to expression in PHHs and HepG2-NTCP-C4 cells. Testing of antiviral agents entecavir and myrcludex | [87]/2017 |
| Genotype D | Liver organoids | Infection with genotype D derived from HepG2.2.15 | Comparison with infection of iPSC-HLCs, HepG2-TET-NTCP organoids, PHH | [90]/2018 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wose Kinge, C.N.; Bhoola, N.H.; Kramvis, A. In Vitro Systems for Studying Different Genotypes/Sub-Genotypes of Hepatitis B Virus: Strengths and Limitations. Viruses 2020, 12, 353. https://doi.org/10.3390/v12030353
Wose Kinge CN, Bhoola NH, Kramvis A. In Vitro Systems for Studying Different Genotypes/Sub-Genotypes of Hepatitis B Virus: Strengths and Limitations. Viruses. 2020; 12(3):353. https://doi.org/10.3390/v12030353
Chicago/Turabian StyleWose Kinge, Constance N., Nimisha H. Bhoola, and Anna Kramvis. 2020. "In Vitro Systems for Studying Different Genotypes/Sub-Genotypes of Hepatitis B Virus: Strengths and Limitations" Viruses 12, no. 3: 353. https://doi.org/10.3390/v12030353
APA StyleWose Kinge, C. N., Bhoola, N. H., & Kramvis, A. (2020). In Vitro Systems for Studying Different Genotypes/Sub-Genotypes of Hepatitis B Virus: Strengths and Limitations. Viruses, 12(3), 353. https://doi.org/10.3390/v12030353