A Broad and Potent H1-Specific Human Monoclonal Antibody Produced in Plants Prevents Influenza Virus Infection and Transmission in Guinea Pigs


Abstract
1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Production of KPF1 in HEK293T Cells
2.3. Production of KPF1 in Plants
2.4. Coomassie Blue Staining and Western Blot
2.5. Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. Immunofluorescence Assay (IFA)
2.7. Microneutralization Assays (MNAs)
2.8. Hemagglutination Inhibition (HAI) Assays
2.9. In Vivo Experiments
2.10. Statistical Analysis
3. Results
3.1. Production of the Human Monoclonal Antibody KPF1 in Tobacco Plants
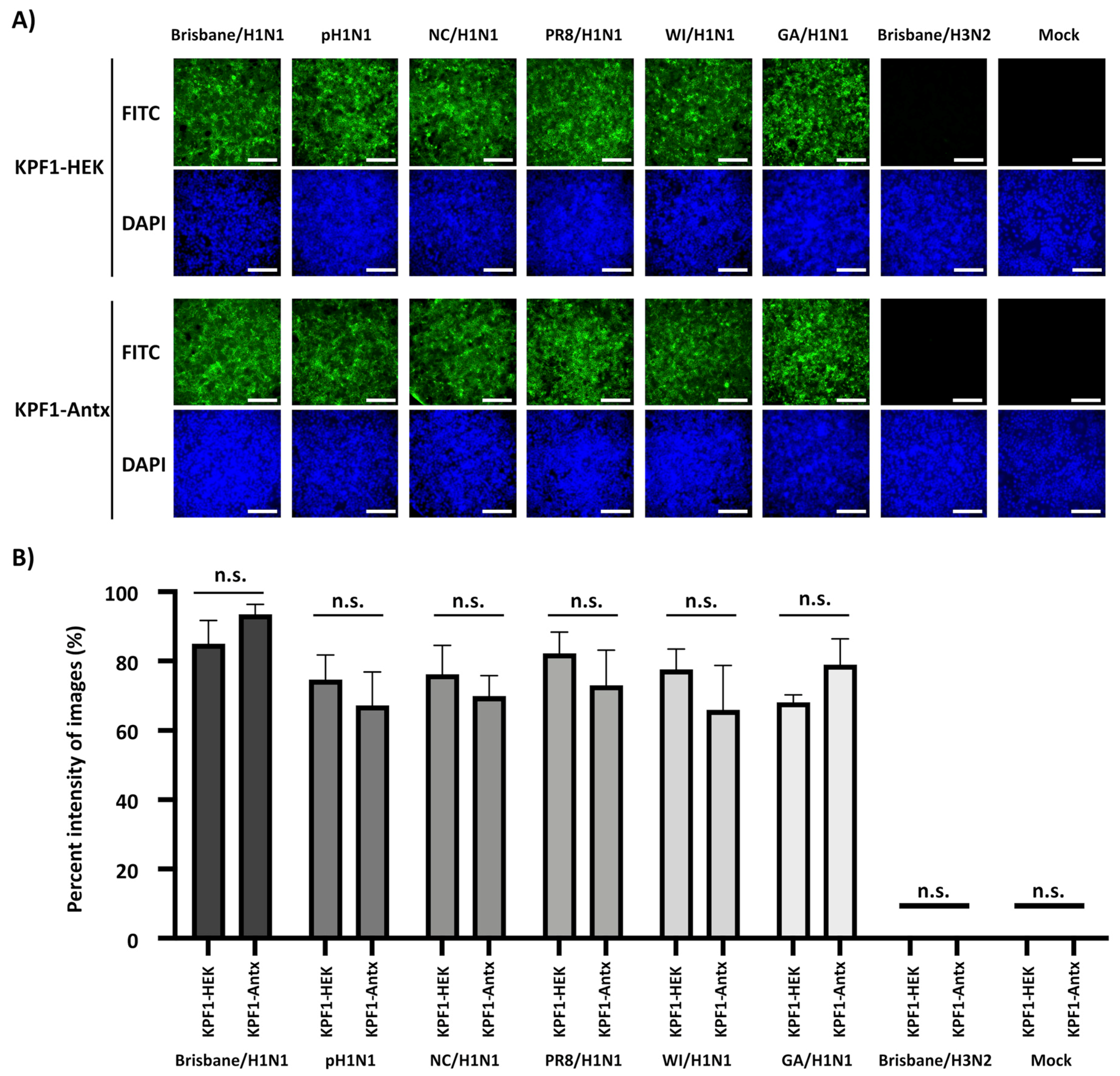
3.2. Reactivity of KPF1-Antx and KPF1-HEK hMAbs In Vitro
3.3. Cross-Reactivity of KPF1-Antx hMAb to H1N1 IAV
3.4. Broad Neutralization and Hemagglutination Inhibition Activity by KPF1-Antx hMAb
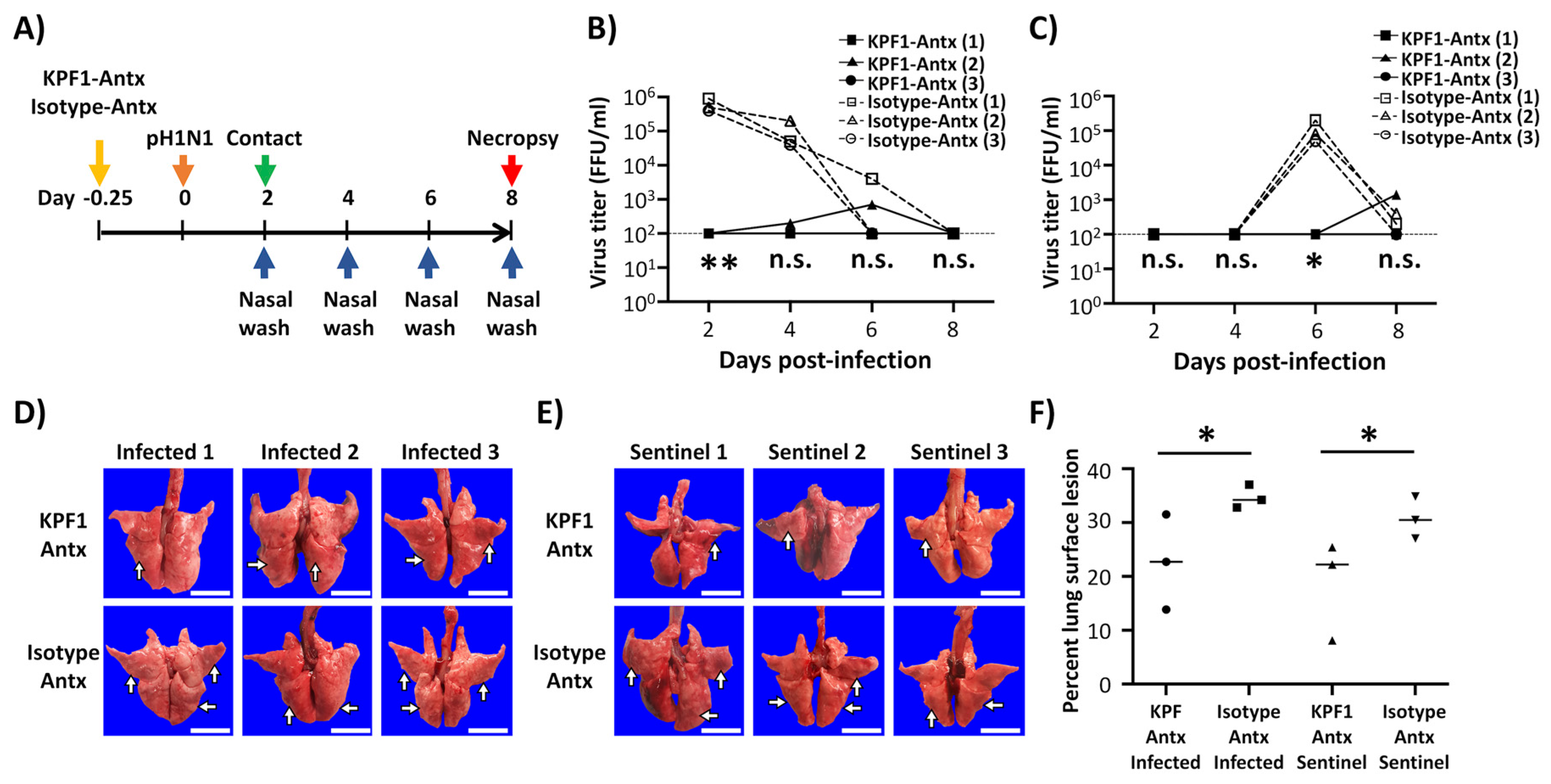
3.5. Prophylactic Activity of KPF1-Antx hMAb In Vivo
3.6. Therapeutic Activity of KPF1-Antx hMAb In Vivo
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Knipe, D.M.; Howley, P.M.; Cohen, J.I.; Griffin, D.E.; Lamb, R.A.; Martin, M.A.; Racaniello, V.R.; Roizman, B. Fields Virology, 6th ed.; Wolters Kluwer Health: Philadelphia, PA, USA, 2013. [Google Scholar]
- Li, K.S.; Guan, Y.; Wang, J.; Smith, G.J.; Xu, K.M.; Duan, L.; Rahardjo, A.P.; Puthavathana, P.; Buranathai, C.; Nguyen, T.D.; et al. Genesis of a highly pathogenic and potentially pandemic H5N1 influenza virus in eastern Asia. Nature 2004, 430, 209–213. [Google Scholar] [CrossRef]
- Barr, I.G.; McCauley, J.; Cox, N.; Daniels, R.; Engelhardt, O.G.; Fukuda, K.; Grohmann, G.; Hay, A.; Kelso, A.; Klimov, A.; et al. Writing Committee of the World Health Organization Consultation on Northern Hemisphere Influenza Vaccine Composition, f. Epidemiological, antigenic and genetic characteristics of seasonal influenza A(H1N1), A(H3N2) and B influenza viruses: Basis for the WHO recommendation on the composition of influenza vaccines for use in the 2009–2010 northern hemisphere season. Vaccine 2010, 28, 1156–1167. [Google Scholar] [PubMed]
- Clark, A.M.; DeDiego, M.L.; Anderson, C.S.; Wang, J.; Yang, H.; Nogales, A.; Martinez-Sobrido, L.; Zand, M.S.; Sangster, M.Y.; Topham, D.J. Antigenicity of the 2015–2016 seasonal H1N1 human influenza virus HA and NA proteins. PLoS ONE 2017, 12, e0188267. [Google Scholar] [CrossRef] [PubMed]
- Federici, C.; Cavazza, M.; Costa, F.; Jommi, C. Health care costs of influenza-related episodes in high income countries: A systematic review. PLoS ONE 2018, 13, e0202787. [Google Scholar] [CrossRef] [PubMed]
- Molinari, N.A.; Ortega-Sanchez, I.R.; Messonnier, M.L.; Thompson, W.W.; Wortley, P.M.; Weintraub, E.; Bridges, C.B. The annual impact of seasonal influenza in the US: Measuring disease burden and costs. Vaccine 2007, 25, 5086–5096. [Google Scholar] [CrossRef]
- Neumann, G.; Kawaoka, Y. Transmission of influenza A viruses. Virology 2015, 479–480, 234–246. [Google Scholar] [CrossRef]
- Herfst, S.; Imai, M.; Kawaoka, Y.; Fouchier, R.A. Avian influenza virus transmission to mammals. Curr. Top. Microbiol. Immunol. 2014, 385, 137–155. [Google Scholar]
- De Clercq, E. Antiviral agents active against influenza A viruses. Nat. Rev. Drug Discov. 2006, 5, 1015–1025. [Google Scholar] [CrossRef]
- Mifsud, E.J.; Hayden, F.G.; Hurt, A.C. Antivirals targeting the polymerase complex of influenza viruses. Antiviral Res. 2019, 169, 104545. [Google Scholar] [CrossRef]
- O’Hanlon, R.; Shaw, M.L. Baloxavir marboxil: The new influenza drug on the market. Curr. Opin. Virol. 2019, 35, 14–18. [Google Scholar] [CrossRef]
- Su, S.; Fu, X.; Li, G.; Kerlin, F.; Veit, M. Novel Influenza D virus: Epidemiology, pathology, evolution and biological characteristics. Virulence 2017, 8, 1580–1591. [Google Scholar] [CrossRef] [PubMed]
- Baker, S.F.; Nogales, A.; Finch, C.; Tuffy, K.M.; Domm, W.; Perez, D.R.; Topham, D.J.; Martinez-Sobrido, L. Influenza A and B virus intertypic reassortment through compatible viral packaging signals. J. Virol. 2014, 88, 10778–10791. [Google Scholar] [CrossRef] [PubMed]
- Influenza Research Database. Available online: https://www.fludb.org/brc/home.spg?decorator=influenza (accessed on 23 December 2019).
- Parrish, C.R.; Kawaoka, Y. The origins of new pandemic viruses: The acquisition of new host ranges by canine parvovirus and influenza A viruses. Annu Rev. Microbiol. 2005, 59, 553–586. [Google Scholar] [CrossRef] [PubMed]
- Parrish, C.R.; Murcia, P.R.; Holmes, E.C. Influenza virus reservoirs and intermediate hosts: Dogs, horses, and new possibilities for influenza virus exposure of humans. J. Virol. 2015, 89, 2990–2994. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.; Zhu, X.; Li, Y.; Shi, M.; Zhang, J.; Bourgeois, M.; Yang, H.; Chen, X.; Recuenco, S.; Gomez, J.; et al. New world bats harbor diverse influenza A viruses. PLoS Pathog. 2013, 9, e1003657. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.; Li, Y.; Rivailler, P.; Conrardy, C.; Castillo, D.A.; Chen, L.M.; Recuenco, S.; Ellison, J.A.; Davis, C.T.; York, I.A.; et al. A distinct lineage of influenza A virus from bats. Proc. Natl. Acad. Sci. USA 2012, 109, 4269–4274. [Google Scholar] [CrossRef]
- Nogales, A.; Piepenbrink, M.S.; Wang, J.; Ortega, S.; Basu, M.; Fucile, C.F.; Treanor, J.J.; Rosenberg, A.F.; Zand, M.S.; Keefer, M.C.; et al. A Highly Potent and Broadly Neutralizing H1 Influenza-Specific Human Monoclonal Antibody. Sci. Rep. 2018, 8, 4374. [Google Scholar] [CrossRef]
- Blanco-Lobo, P.; Nogales, A.; Rodriguez, L.; Martinez-Sobrido, L. Novel Approaches for The Development of Live Attenuated Influenza Vaccines. Viruses 2019, 11, 190. [Google Scholar] [CrossRef]
- Martinez-Sobrido, L.; Peersen, O.; Nogales, A. Temperature Sensitive Mutations in Influenza A Viral Ribonucleoprotein Complex Responsible for the Attenuation of the Live Attenuated Influenza Vaccine. Viruses 2018, 10, 560. [Google Scholar] [CrossRef]
- Nogales, A.; Martinez-Sobrido, L. Reverse Genetics Approaches for the Development of Influenza Vaccines. Int. J. Mol. Sci. 2016, 18, 20. [Google Scholar] [CrossRef]
- Takashita, E.; Kawakami, C.; Morita, H.; Ogawa, R.; Fujisaki, S.; Shirakura, M.; Miura, H.; Nakamura, K.; Kishida, N.; Kuwahara, T.; on behalf of the Influenza Virus Surveillance Group of Japan; et al. Detection of influenza A(H3N2) viruses exhibiting reduced susceptibility to the novel cap-dependent endonuclease inhibitor baloxavir in Japan, December 2018. Euro. Surveill. 2019, 24. [Google Scholar] [CrossRef] [PubMed]
- Earhart, K.C.; Elsayed, N.M.; Saad, M.D.; Gubareva, L.V.; Nayel, A.; Deyde, V.M.; Abdelsattar, A.; Abdelghani, A.S.; Boynton, B.R.; Mansour, M.M.; et al. Oseltamivir resistance mutation N294S in human influenza A(H5N1) virus in Egypt. J. Infect. Public Health 2009, 2, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Omoto, S.; Speranzini, V.; Hashimoto, T.; Noshi, T.; Yamaguchi, H.; Kawai, M.; Kawaguchi, K.; Uehara, T.; Shishido, T.; Naito, A.; et al. Characterization of influenza virus variants induced by treatment with the endonuclease inhibitor baloxavir marboxil. Sci. Rep. 2018, 8, 9633. [Google Scholar] [CrossRef] [PubMed]
- He, G.; Massarella, J.; Ward, P. Clinical pharmacokinetics of the prodrug oseltamivir and its active metabolite Ro 64-0802. Clin. Pharmacokinet. 1999, 37, 471–484. [Google Scholar] [CrossRef]
- Davies, B.E. Pharmacokinetics of oseltamivir: An oral antiviral for the treatment and prophylaxis of influenza in diverse populations. J. Antimicrob. Chemother. 2010, 65, ii5–ii10. [Google Scholar] [CrossRef]
- Horadam, V.W.; Sharp, J.G.; Smilack, J.D.; McAnalley, B.H.; Garriott, J.C.; Stephens, M.K.; Prati, R.C.; Brater, D.C. Pharmacokinetics of amantadine hydrochloride in subjects with normal and impaired renal function. Ann. Intern. Med. 1981, 94, 454–458. [Google Scholar] [CrossRef]
- Ng, K.E. Xofluza (Baloxavir Marboxil) for the Treatment Of Acute Uncomplicated Influenza. P T 2019, 44, 9–11. [Google Scholar]
- Wycoff, K.L. Secretory IgA antibodies from plants. Curr. Pharm. Des. 2005, 11, 2429–2437. [Google Scholar] [CrossRef]
- Rodriguez, M.; Perez, L.; Gavilondo, J.V.; Garrido, G.; Bequet-Romero, M.; Hernandez, I.; Huerta, V.; Cabrera, G.; Perez, M.; Ramos, O.; et al. Comparative in vitro and experimental in vivo studies of the anti-epidermal growth factor receptor antibody nimotuzumab and its aglycosylated form produced in transgenic tobacco plants. Plant. Biotechnol. J. 2013, 11, 53–65. [Google Scholar] [CrossRef]
- Hiatt, A.; Cafferkey, R.; Bowdish, K. Production of antibodies in transgenic plants. Nature 1989, 342, 76–78. [Google Scholar] [CrossRef]
- Lai, H.; He, J.; Engle, M.; Diamond, M.S.; Chen, Q. Robust production of virus-like particles and monoclonal antibodies with geminiviral replicon vectors in lettuce. Plant. Biotechnol. J. 2012, 10, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Hu, W.; Pestka, J.J.; He, S.Y.; Hart, L.P. Expression of a functional antizearalenone single-chain Fv antibody in transgenic Arabidopsis plants. Appl. Environ. Microbiol. 2000, 66, 3499–3505. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.S.; Moon, J.H.; Kim, T.G.; Lee, J.Y. Potent In Vitro and In Vivo Activity of Plantibody Specific for Porphyromonas gingivalis FimA. Clin. Vaccine Immunol. 2016, 23, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, K.; Narimatsu, S.; Ichikawa, S.; Tobisawa, Y.; Kurohane, K.; Niwa, Y.; Kobayashi, H.; Imai, Y. Production of hybrid-IgG/IgA plantibodies with neutralizing activity against Shiga toxin 1. PLoS ONE 2013, 8, e80712. [Google Scholar] [CrossRef] [PubMed]
- Chaisri, U.; Chaicumpa, W. Evolution of Therapeutic Antibodies, Influenza Virus Biology, Influenza, and Influenza Immunotherapy. Biomed. Res. Int. 2018, 2018, 9747549. [Google Scholar] [CrossRef] [PubMed]
- Sully, E.K.; Whaley, K.J.; Bohorova, N.; Bohorov, O.; Goodman, C.; Kim, D.H.; Pauly, M.H.; Velasco, J.; Hiatt, E.; Morton, J.; et al. Chimeric plantibody passively protects mice against aerosolized ricin challenge. Clin. Vaccine Immunol. 2014, 21, 777–782. [Google Scholar] [CrossRef] [PubMed]
- Shafaghi, M.; Maktoobian, S.; Rasouli, R.; Howaizi, N.; Ofoghi, H.; Ehsani, P. Transient Expression of Biologically Active Anti-rabies Virus Monoclonal Antibody in Tobacco Leaves. Iran. J. Biotechnol. 2018, 16, e1774. [Google Scholar] [CrossRef]
- Kopertekh, L.; Meyer, T.; Freyer, C.; Hust, M. Transient plant production of Salmonella Typhimurium diagnostic antibodies. Biotechnol. Rep. (Amst) 2019, 21, e00314. [Google Scholar] [CrossRef]
- Nakanishi, K.; Morikane, S.; Ichikawa, S.; Kurohane, K.; Niwa, Y.; Akimoto, Y.; Matsubara, S.; Kawakami, H.; Kobayashi, H.; Imai, Y. Protection of Human Colon Cells from Shiga Toxin by Plant-based Recombinant Secretory IgA. Sci. Rep. 2017, 7, 45843. [Google Scholar] [CrossRef]
- Lai, H.; Engle, M.; Fuchs, A.; Keller, T.; Johnson, S.; Gorlatov, S.; Diamond, M.S.; Chen, Q. Monoclonal antibody produced in plants efficiently treats West Nile virus infection in mice. Proc. Natl. Acad. Sci. USA 2010, 107, 2419–2424. [Google Scholar] [CrossRef]
- Rattanapisit, K.; Srijangwad, A.; Chuanasa, T.; Sukrong, S.; Tantituvanont, A.; Mason, H.S.; Nilubol, D.; Phoolcharoen, W. Rapid Transient Production of a Monoclonal Antibody Neutralizing the Porcine Epidemic Diarrhea Virus (PEDV) in Nicotiana benthamiana and Lactuca sativa. Planta Med. 2017, 83, 1412–1419. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Velazquez, A.; Lopez-Quesada, A.; Ceballo-Camara, Y.; Cabrera-Herrera, G.; Tiel-Gonzalez, K.; Mirabal-Ortega, L.; Perez-Martinez, M.; Perez-Castillo, R.; Rosabal-Ayan, Y.; Ramos-Gonzalez, O.; et al. Tobacco seeds as efficient production platform for a biologically active anti-HBsAg monoclonal antibody. Transgenic. Res. 2015, 24, 897–909. [Google Scholar] [CrossRef] [PubMed]
- Dubey, K.K.; Luke, G.A.; Knox, C.; Kumar, P.; Pletschke, B.I.; Singh, P.K.; Shukla, P. Vaccine and antibody production in plants: Developments and computational tools. Brief. Funct. Genomics 2018, 17, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Olinger, G.G., Jr.; Pettitt, J.; Kim, D.; Working, C.; Bohorov, O.; Bratcher, B.; Hiatt, E.; Hume, S.D.; Johnson, A.K.; Morton, J.; et al. Delayed treatment of Ebola virus infection with plant-derived monoclonal antibodies provides protection in rhesus macaques. Proc. Natl. Acad. Sci. USA 2012, 109, 18030–18035. [Google Scholar] [CrossRef]
- Nogales, A.; Avila-Perez, G.; Rangel-Moreno, J.; Chiem, K.; DeDiego, M.L.; Martinez-Sobrido, L. A novel fluorescent and bioluminescent Bi-Reporter influenza A virus (BIRFLU) to evaluate viral infections. J. Virol. 2019, 93, e00032-19. [Google Scholar] [CrossRef]
- Nogales, A.; Baker, S.F.; Martinez-Sobrido, L. Replication-competent influenza A viruses expressing a red fluorescent protein. Virology 2015, 476, 206–216. [Google Scholar] [CrossRef]
- Nogales, A.; Rodriguez, L.; DeDiego, M.L.; Topham, D.J.; Martinez-Sobrido, L. Interplay of PA-X and NS1 Proteins in Replication and Pathogenesis of a Temperature-Sensitive 2009 Pandemic H1N1 Influenza A Virus. J. Virol. 2017, 91, e00720-17. [Google Scholar] [CrossRef]
- Wing-Fai Cheung, H.W.; Rebecca, P.; Hirak, S.; Warren, W.; Wakarchuk, D.C.a.M.D.M. Development of fine-control expression vectors for post-translational modification of therapeutic proteins in plants. Mol. Biotechnol. 2020. Submitted. [Google Scholar]
- Castilho, A.; Beihammer, G.; Pfeiffer, C.; Goritzer, K.; Montero-Morales, L.; Vavra, U.; Maresch, D.; Grunwald-Gruber, C.; Altmann, F.; Steinkellner, H.; et al. An oligosaccharyltransferase from Leishmania major increases the N-glycan occupancy on recombinant glycoproteins produced in Nicotiana benthamiana. Plant. Biotechnol. J. 2018, 16, 1700–1709. [Google Scholar] [CrossRef]
- Garabagi, F.; McLean, M.D.; Hall, J.C. Transient and stable expression of antibodies in Nicotiana species. Methods Mol. Biol. 2012, 907, 389–408. [Google Scholar]
- Jensen, E.C. Quantitative analysis of histological staining and fluorescence using ImageJ. Anat Rec. (Hoboken) 2013, 296, 378–381. [Google Scholar] [CrossRef]
- National Research Council (U.S.). Committee for the Update of the Guide for the Care and Use of Laboratory Animals. Guide for the Care and Use of Laboratory Animals, 8th ed.; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Gauger, P.C.; Loving, C.L.; Khurana, S.; Lorusso, A.; Perez, D.R.; Kehrli, M.E., Jr.; Roth, J.A.; Golding, H.; Vincent, A.L. Live attenuated influenza A virus vaccine protects against A(H1N1)pdm09 heterologous challenge without vaccine associated enhanced respiratory disease. Virology 2014, 471–473, 93–104. [Google Scholar] [CrossRef]
- Morgan, S.B.; Hemmink, J.D.; Porter, E.; Harley, R.; Shelton, H.; Aramouni, M.; Everett, H.E.; Brookes, S.M.; Bailey, M.; Townsend, A.M.; et al. Aerosol Delivery of a Candidate Universal Influenza Vaccine Reduces Viral Load in Pigs Challenged with Pandemic H1N1 Virus. J. Immunol. 2016, 196, 5014–5023. [Google Scholar] [CrossRef] [PubMed]
- Robert, S.; Goulet, M.C.; D’Aoust, M.A.; Sainsbury, F.; Michaud, D. Leaf proteome rebalancing in Nicotiana benthamiana for upstream enrichment of a transiently expressed recombinant protein. Plant. Biotechnol. J. 2015, 13, 1169–1179. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, K.G.; Aoki, F.Y.; Osterhaus, A.D.; Trottier, S.; Carewicz, O.; Mercier, C.H.; Rode, A.; Kinnersley, N.; Ward, P. Efficacy and safety of oseltamivir in treatment of acute influenza: A randomised controlled trial. Neuraminidase Inhibitor Flu Treatment Investigator Group. Lancet 2000, 355, 1845–1850. [Google Scholar] [CrossRef]
- McLean, H.Q.; Belongia, E.A.; Kieke, B.A.; Meece, J.K.; Fry, A.M. Impact of Late Oseltamivir Treatment on Influenza Symptoms in the Outpatient Setting: Results of a Randomized Trial. Open Forum Infect. Dis. 2015, 2, ofv100. [Google Scholar] [CrossRef]
- Gomord, V.; Fitchette, A.C.; Menu-Bouaouiche, L.; Saint-Jore-Dupas, C.; Plasson, C.; Michaud, D.; Faye, L. Plant-specific glycosylation patterns in the context of therapeutic protein production. Plant. Biotechnol. J. 2010, 8, 564–587. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Viruses | NT50 (µg/mL) MNA | HAI (µg/mL) | ||
|---|---|---|---|---|
| KPF1-HEK | KPF1-Antx | KPF1-HEK | KPF1-Antx | |
| Brisbane/H1N1 | 0.708 | 0.780 | 0.552 | 0.552 |
| pH1N1 | 0.427 | 0.427 | 1.106 | 0.552 |
| NC/H1N1 | 1.106 | 0.715 | 8.714 | 17.425 |
| PR8/H1N1 | 25.0 | 12.5 | 17.425 | 17.425 |
| WI/H1N1 | 0.195 | 0.195 | 0.271 | 0.271 |
| GA/H1N1 | 0.353 | 0.221 | 0.271 | 0.271 |
| Brisbane/H3N2 | >200 | >200 | ND | ND |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.-G.; Ye, C.; Piepenbrink, M.S.; Nogales, A.; Wang, H.; Shuen, M.; Meyers, A.J.; Martinez-Sobrido, L.; Kobie, J.J. A Broad and Potent H1-Specific Human Monoclonal Antibody Produced in Plants Prevents Influenza Virus Infection and Transmission in Guinea Pigs. Viruses 2020, 12, 167. https://doi.org/10.3390/v12020167
Park J-G, Ye C, Piepenbrink MS, Nogales A, Wang H, Shuen M, Meyers AJ, Martinez-Sobrido L, Kobie JJ. A Broad and Potent H1-Specific Human Monoclonal Antibody Produced in Plants Prevents Influenza Virus Infection and Transmission in Guinea Pigs. Viruses. 2020; 12(2):167. https://doi.org/10.3390/v12020167
Chicago/Turabian StylePark, Jun-Gyu, Chengjin Ye, Michael S. Piepenbrink, Aitor Nogales, Haifeng Wang, Michael Shuen, Ashley J. Meyers, Luis Martinez-Sobrido, and James J. Kobie. 2020. "A Broad and Potent H1-Specific Human Monoclonal Antibody Produced in Plants Prevents Influenza Virus Infection and Transmission in Guinea Pigs" Viruses 12, no. 2: 167. https://doi.org/10.3390/v12020167
APA StylePark, J.-G., Ye, C., Piepenbrink, M. S., Nogales, A., Wang, H., Shuen, M., Meyers, A. J., Martinez-Sobrido, L., & Kobie, J. J. (2020). A Broad and Potent H1-Specific Human Monoclonal Antibody Produced in Plants Prevents Influenza Virus Infection and Transmission in Guinea Pigs. Viruses, 12(2), 167. https://doi.org/10.3390/v12020167