High-Throughput Method for Detection of Arbovirus Infection of Saliva in Mosquitoes Aedes aegypti and Ae. albopictus

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Mosquito Populations and Rearing
2.2. Chikungunya Virus Isolates, Propagation, and Mosquito Infection
2.3. Transmission Assays
2.4. Viral RNA Isolation and qRT-PCR
2.5. Statistical Analyses
3. Results
3.1. Overall Results
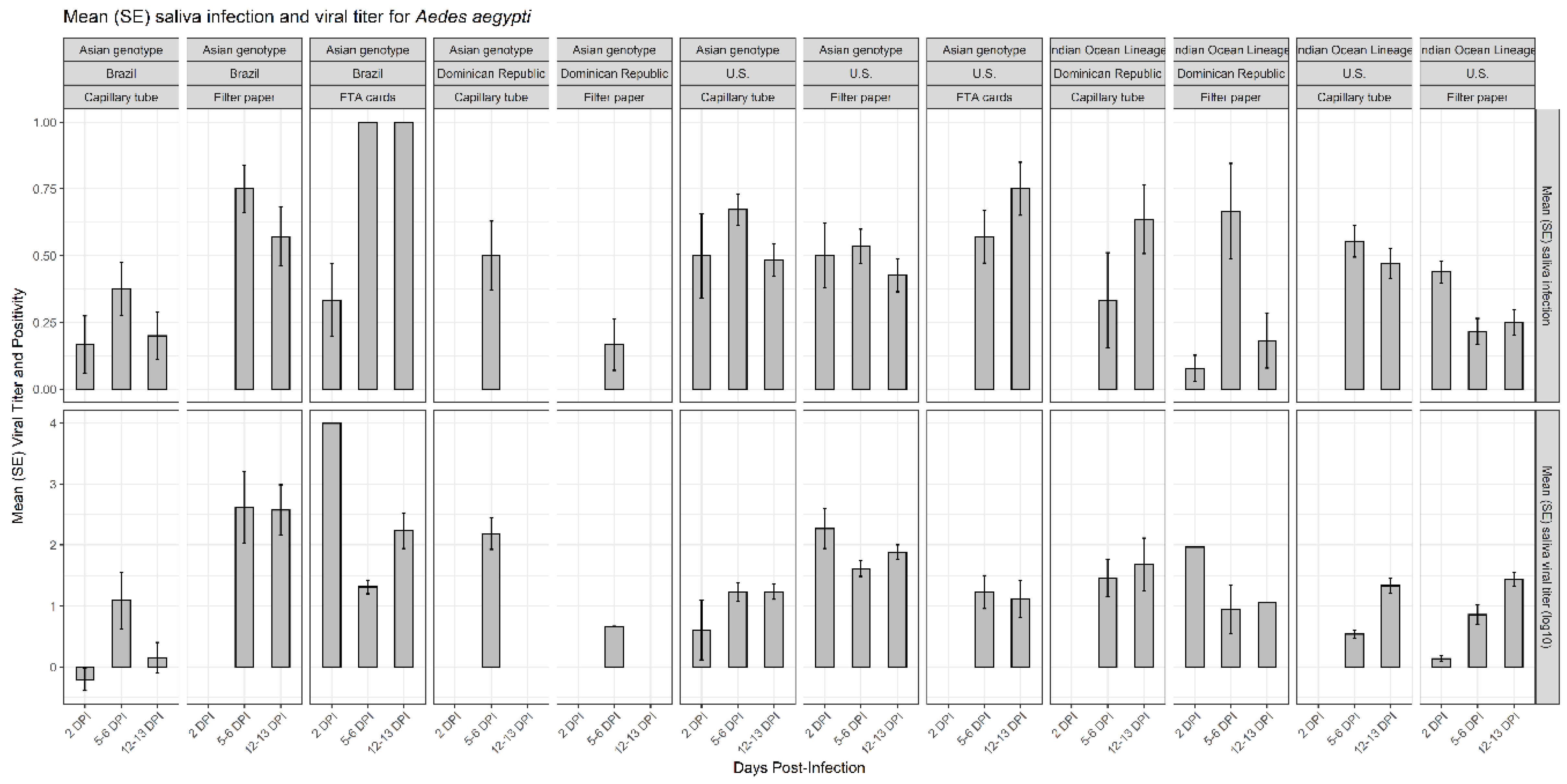
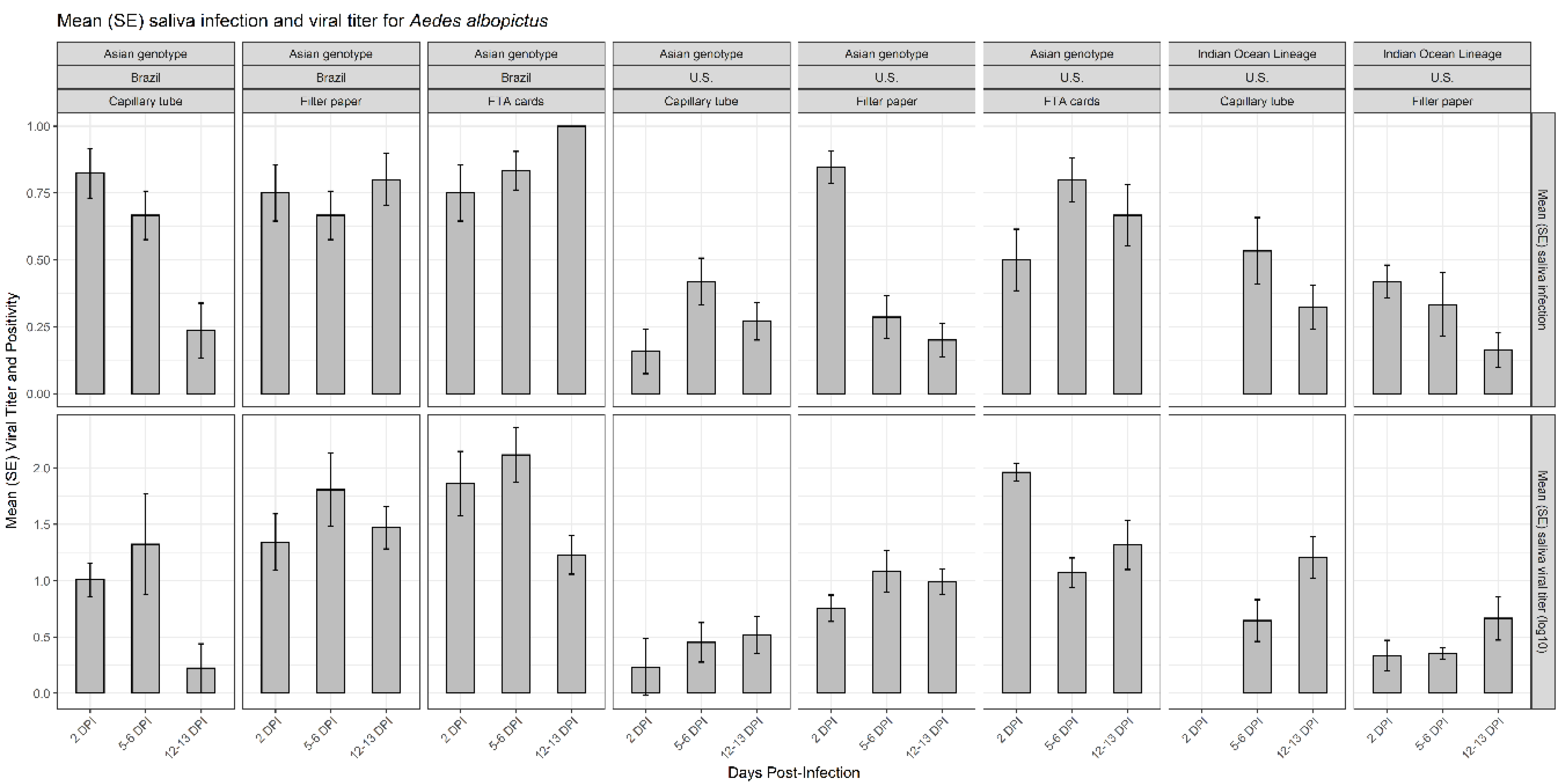
3.2. Effects of Viral Dissemination, Country of Origin, and Days Post-Infection on CHIKV Salivary Positivity and CHIKV Viral Titer on Mosquito Saliva
3.3. Efficacy of Capillary Tube, Filter Paper, and FTA Cards for CHIKV Detection and CHIKV Viral Titer in Mosquito Saliva
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Josseran, L.; Paquet, C.; Zehgnoun, A.; Caillère, N.; Le Tertre, A.; Solet, J.-L.; Ledrans, M. Chikungunya disease outbreak, Reunion Island. Emerg. Infect. Dis. 2006, 12, 1994–1995. [Google Scholar] [CrossRef] [PubMed]
- Chretien, J.-P.; Sergon, K.; Sang, R.; Breiman, R.F.; Anyamba, A.; Bedno, S.A.; Linthicum, K.J.; Tucker, C.J.; Small, J.; Onyango, C.O.; et al. Drought-associated chikungunya emergence along coastal East Africa. Am. J. Trop. Med. Hyg. 2007, 76, 405–407. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.C. Arrival of Chikungunya Virus in the New World: Prospects for Spread and Impact on Public Health. PLoS Neglected Trop. Dis. 2014, 8, e2921. [Google Scholar] [CrossRef] [PubMed]
- Leparc-Goffart, I.; Nougairede, A.; Cassadou, S.; Prat, C.; De Lamballerie, X. Chikungunya in the Americas. Lancet 2014, 383, 514. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Valadere, A.M. Transcontinental Movement of Asian Genotype Chikungunya Virus. Emerg. Infect. Dis. 2014, 20, 1400–1402. [Google Scholar] [CrossRef]
- Honório, N.A.; Camara, D.C.P.; Calvet, G.A.; Brasil, P. Chikungunya: Uma arbovirose em estabelecimento e expansão no Brasil. Cadernos de Saúde Pública 2015, 31, 906–908. [Google Scholar] [CrossRef]
- Faye, O.; Freire, C.C.M.; Iamarino, A.; Faye, O.; De Oliveira, J.V.C.; Diallo, M.; Zanotto, P.M.A.; Sall, A.A. Molecular Evolution of Zika Virus during Its Emergence in the 20th Century. PLoS Neglected Trop. Dis. 2014, 8, e2636. [Google Scholar] [CrossRef]
- Cao-Lormeau, V.-M.; Roche, C.; Teissier, A.; Robin, E.; Berry, A.-L.; Mallet, H.-P.; Sall, A.A.; Musso, D. Zika Virus, French Polynesia, South Pacific, 2013. Emerg. Infect. Dis. 2014, 20, 1084–1086. [Google Scholar] [CrossRef]
- Campos, G.S.; Bandeira, A.C.; Sardi, S.I. Zika Virus Outbreak, Bahia, Brazil. Emerg. Infect. Dis. 2015, 21, 1885–1886. [Google Scholar] [CrossRef]
- Lounibos, L.P.; Kramer, L.D. Invasiveness of Aedes aegypti and Aedes albopictus and Vectorial Capacity for Chikungunya Virus. J. Infect. Dis. 2016, 214, S453–S458. [Google Scholar] [CrossRef]
- Kramer, L.D.; Ciota, A.T. Dissecting vectorial capacity for mosquito-borne viruses. Curr. Opin. Virol. 2015, 15, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Franz, A.W.E.; Sanchez-Vargas, I.; Adelman, Z.N.; Blair, C.D.; Beaty, B.J.; James, A.A.; Olson, K.E. Engineering RNA interference-based resistance to dengue virus type 2 in genetically modified Aedes aegypti. Proc. Natl. Acad. Sci. 2006, 103, 4198–4203. [Google Scholar] [CrossRef]
- Franz, A.W.E.; Sanchez-Vargas, I.; Raban, R.R.; Iv, W.C.B.; James, A.A.; Olson, K.E. Fitness Impact and Stability of a Transgene Conferring Resistance to Dengue-2 Virus following Introgression into a Genetically Diverse Aedes aegypti Strain. PLoS Neglected Trop. Dis. 2014, 8, e2833. [Google Scholar] [CrossRef]
- Brackney, D.E.; Foy, B.D.; Olson, K.E. The effects of midgut serine proteases on dengue virus type 2 infectivity of Aedes aegypti. Am. J. Trop. Med. Hyg. 2008, 79, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Jupatanakul, N.; Sim, S.; Dimopoulos, G. The Insect Microbiome Modulates Vector Competence for Arboviruses. Viruses 2014, 6, 4294–4313. [Google Scholar] [CrossRef]
- Caragata, E.P.; Dutra, H.L.C.; Moreira, L.A. Inhibition of Zika virus by Wolbachia in Aedes aegypti. Microb. Cell 2016, 3, 293–295. [Google Scholar] [CrossRef] [PubMed]
- Conway, M.J.; Colpitts, T.M.; Fikrig, E. Role of the Vector in Arbovirus Transmission. Annu. Rev. Virol. 2014, 1, 71–88. [Google Scholar] [CrossRef]
- Kramer, L.D.; Hardy, J.L.; Presser, S.B.; Houk, E.J. Dissemination Barriers for Western Equine Encephalomyelitis Virus in Culex tarsalis Infected after Ingestion of Low Viral Doses *. Am. J. Trop. Med. Hyg. 1981, 30, 190–197. [Google Scholar] [CrossRef]
- Smith, D.R.; Aguilar, P.V.; Weaver, S.C.; Carrara, A.-S. Evaluation of Methods to Assess Transmission Potential of Venezuelan Equine Encephalitis Virus by Mosquitoes and Estimation of Mosquito Saliva Titers. Am. J. Trop. Med. Hyg. 2005, 73, 33–39. [Google Scholar] [CrossRef]
- Vega-Rúa, A.; Zouache, K.; Girod, R.; Failloux, A.-B.; Lourenço-De-Oliveira, R. High Level of Vector Competence of Aedes aegypti and Aedes albopictus from Ten American Countries as a Crucial Factor in the Spread of Chikungunya Virus. J. Virol. 2014, 88, 6294–6306. [Google Scholar] [CrossRef]
- Alto, B.W.; Wiggins, K.; Eastmond, B.; Velez, D.; Lounibos, L.P.; Lord, C.C. Transmission risk of two chikungunya lineages by invasive mosquito vectors from Florida and the Dominican Republic. PLoS Neglected Trop. Dis. 2017, 11, e0005724. [Google Scholar] [CrossRef] [PubMed]
- Hurlbut, H.S. Mosquito Salivation and Virus Transmission *. Am. J. Trop. Med. Hyg. 1966, 15, 989–993. [Google Scholar] [CrossRef] [PubMed]
- Hall-Mendelin, S.; Ritchie, S.A.; Johansen, C.A.; Zborowski, P.; Cortis, G.; Dandridge, S.; Hall, R.A.; Hurk, A.F.V.D. Exploiting mosquito sugar feeding to detect mosquito-borne pathogens. Proc. Natl. Acad. Sci. 2010, 107, 11255–11259. [Google Scholar] [CrossRef] [PubMed]
- Glushakova, L.G.; Alto, B.W.; Kim, M.-S.; Wiggins, K.; Eastmond, B.; Moussatche, P.; Burkett-Cadena, N.D.; Benner, S.A. Optimization of cationic (Q)-paper for detection of arboviruses in infected mosquitoes. J. Virol. Methods 2018, 261, 71–79. [Google Scholar] [CrossRef]
- Burkhalter, K.L.; Wiggins, K.; Burkett-Cadena, N.D.; Alto, B.W. Laboratory Evaluation of Commercially Available Platforms to Detect West Nile and Zika Viruses From Honey Cards. J. Med Èntomol. 2018, 55, 717–722. [Google Scholar] [CrossRef]
- Honório, N.A.; Wiggins, K.; Câmara, D.C.P.; Eastmond, B.; Alto, B.W. Chikungunya virus vector competency of Brazilian and Florida mosquito vectors. PLoS Neglected Trop. Dis. 2018, 12, e0006521. [Google Scholar] [CrossRef]
- A Zimler, R.; Alto, B.W. Florida Aedes aegypti (Diptera: Culicidae) and Aedes albopictus Vector Competency for Zika Virus. J. Med Èntomol. 2019, 56, 341–346. [Google Scholar] [CrossRef]
- Alto, B.W.; Connelly, C.R.; O’Meara, G.F.; Hickman, D.; Karr, N. Reproductive Biology and Susceptibility of Florida Culex coronator to Infection with West Nile Virus. Vector-Borne Zoonotic Dis. 2014, 14, 606–614. [Google Scholar] [CrossRef]
- Reiskind, M.H.; Pesko, K.; Westbrook, C.J.; Mores, C.N. Susceptibility of Florida mosquitoes to infection with chikungunya virus. Am. J. Trop. Med. Hyg. 2008, 78, 422–425. [Google Scholar] [CrossRef]
- Bustin, S. Absolute quantification of mRNA using real-time reverse transcription polymerase chain reaction assays. J. Mol. Endocrinol. 2000, 25, 169–193. [Google Scholar] [CrossRef]
- Fox, J. Applied Regression Analysis and Generalized Linear Models; SAGE Publications: Thousand Oaks, CA, USA, 2016; ISBN 978-1-4522-0566-3. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. 2018. Available online: https://www.r-project.org/ (accessed on 13 February 2012).
- RStudio Team. RStudio: Integrated Development for R; RStudio, PBC: Boston, MA, USA, 2019; Available online: http://www.rstudio.com/ (accessed on 6 January 2020).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Merritt, R.W.; Clements, A.N. The Biology of Mosquitoes, Volume 1: Development, Nutrition, and Reproduction. A. N. Clements. J. North Am. Benthol. Soc. 1993, 12, 304–305. [Google Scholar] [CrossRef]
- Tabachnick, W.J. Nature, Nurture and Evolution of Intra-Species Variation in Mosquito Arbovirus Transmission Competence. Int. J. Environ. Res. Public Heal. 2013, 10, 249–277. [Google Scholar] [CrossRef] [PubMed]
- Vega-Rúa, A.; Lourenço-De-Oliveira, R.; Mousson, L.; Vazeille, M.; Fuchs, S.; Yebakima, A.; Gustave, J.; Girod, R.; Dusfour, I.; Leparc-Goffart, I.; et al. Chikungunya Virus Transmission Potential by Local Aedes Mosquitoes in the Americas and Europe. PLoS Neglected Trop. Dis. 2015, 9, e0003780. [Google Scholar] [CrossRef]
- Ritchie, S.A.; Cortis, G.; Paton, C.; Townsend, M.; Shroyer, D.; Zborowski, P.; Hall-Mendelin, S.; Hurk, A.F.V.D. A simple non-powered passive trap for the collection of mosquitoes for arbovirus surveillance. J. Med Èntomol. 2013, 50, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Cardona-Ospina, J.A.; Villalba-Miranda, M.F.; Palechor-Ocampo, L.A.; Mancilla, L.I.; Sepúlveda-Arias, J.C. A systematic review of FTA cards® as a tool for viral RNA preservation in fieldwork: Are they safe and effective? Prev. Veter- Med. 2019, 172, 104772. [Google Scholar] [CrossRef]
- Guedes, D.R.; Paiva, M.H.; Donato, M.; Barbosa, P.P.; Krokovsky, L.; Rocha, S.W.D.S.; La Saraiva, K.; Crespo, M.M.; Rezende, T.M.; Wallau, G.L.; et al. Zika virus replication in the mosquito Culex quinquefasciatus in Brazil. Emerg. Microbes Infect. 2017, 6, 1–11. [Google Scholar] [CrossRef]
- Alto, B.W.; Wiggins, K.; Eastmond, B.; Ortiz, S.; Zirbel, K.; Lounibos, L.P. Diurnal Temperature Range and Chikungunya Virus Infection in Invasive Mosquito Vectors. J. Med Èntomol. 2017, 55, 217–224. [Google Scholar] [CrossRef]
- Coffey, L.L.; Failloux, A.-B.; Weaver, S.C. Chikungunya Virus–Vector Interactions. Viruses 2014, 6, 4628–4663. [Google Scholar] [CrossRef]
- Flies, E.J.; Toi, C.; Weinstein, P.; Doggett, S.L.; Williams, C.R. Converting Mosquito Surveillance to Arbovirus Surveillance with Honey-Baited Nucleic Acid Preservation Cards. Vector-Borne Zoonotic Dis. 2015, 15, 397–403. [Google Scholar] [CrossRef]
- Hurk, A.F.V.D.; Hall-Mendelin, S.; Townsend, M.; Kurucz, N.; Edwards, J.; Ehlers, G.; Rodwell, C.; Moore, F.A.; McMahon, J.; Northill, J.A.; et al. Applications of a Sugar-Based Surveillance System to Track Arboviruses in Wild Mosquito Populations. Vector-Borne Zoonotic Dis. 2014, 14, 66–73. [Google Scholar] [CrossRef]
- Burkett-Cadena, N.D.; Gibson, J.; Lauth, M.; Stenn, T.; Acevedo, C.; Xue, R.-D.; McNelly, J.; Northey, E.; Hassan, H.K.; Fulcher, A.; et al. Evaluation of the Honey-Card Technique for Detection of Transmission of Arboviruses in Florida and Comparison With Sentinel Chicken Seroconversion. J. Med Èntomol. 2016, 53, 1449–1457. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Genotype | Species | Country | Detection Method | Positivity (Mean) | Positivity (SE) | Viral Titer (Mean) | Viral Titer (SE) |
|---|---|---|---|---|---|---|---|
| Asian | Aedes aegypti | Brazil | Capillary tube | 0.2679 | 0.0592 | 0.5086 | 0.2369 |
| Filter paper | 0.6667 | 0.063 | 2.5959 | 0.3174 | |||
| FTA cards | 0.8182 | 0.0515 | 2.0199 | 0.1644 | |||
| Dominican Republic | Capillary tube | 0.5 | 0.1291 | 2.1854 | 0.2619 | ||
| Filter paper | 0.1667 | 0.0962 | 0.6653 | 0.0038 | |||
| US | Capillary tube | 0.5725 | 0.0423 | 1.1672 | 0.0998 | ||
| Filter paper | 0.4808 | 0.0416 | 1.8146 | 0.0875 | |||
| FTA cards | 0.6364 | 0.0655 | 1.1783 | 0.1713 | |||
| Aedes albopictus | Brazil | Capillary tube | 0.5902 | 0.063 | 0.9792 | 0.2221 | |
| Filter paper | 0.7333 | 0.0566 | 1.5888 | 0.1563 | |||
| FTA cards | 0.8333 | 0.0477 | 1.8855 | 0.1413 | |||
| US | Capillary tube | 0.2989 | 0.048 | 0.4204 | 0.1068 | ||
| Filter paper | 0.3846 | 0.0473 | 0.8981 | 0.0738 | |||
| FTA cards | 0.6667 | 0.0609 | 1.3243 | 0.0876 | |||
| IOL | Aedes aegypti | Dominican Republic | Capillary tube | 0.5294 | 0.1089 | 1.6296 | 0.3121 |
| Filter paper | 0.2333 | 0.0598 | 1.1342 | 0.1291 | |||
| US | Capillary tube | 0.5115 | 0.0408 | 0.9062 | 0.0778 | ||
| Filter paper | 0.3346 | 0.0276 | 0.5059 | 0.0569 | |||
| Aedes albopictus | US | Capillary tube | 0.3913 | 0.0704 | 0.9577 | 0.1367 | |
| Filter paper | 0.3333 | 0.0443 | 0.384 | 0.0949 |
| Species | Asian Genotype | IOL Genotype | |||||
|---|---|---|---|---|---|---|---|
| Aedes aegypti | Binomial model for CHIKV saliva infection | ||||||
| Effects | Estimate | SE | CI95 | Estimate | SE | CI95 | |
| Intercept | 0.176 | 0.4814 | [0.065, 0.436] | 0.336 | 0.3798 | [0.156, 0.697] | |
| Viral Dissemination (legs) | 1.49 | 0.1 | [1.232, 1.826] | 1.292 | 0.05409 | [1.163, 1.438] | |
| Country: Dom. Republic | 0.874 | 0.5546 | [0.288, 2.582] | (not included in the model) | |||
| Country: US | 1.221 | 0.2773 | [0.708, 2.107] | 1.521 | 0.3443 | [0.785, 3.051] | |
| 5–6 dpi | 1.261 | 0.4211 | [0.547, 2.881] | 0.87 | 0.2503 | [0.532, 1.421] | |
| 12–13 dpi | 0.727 | 0.4148 | [0.32, 1.644] | 0.66 | 0.2535 | [0.4, 1.082] | |
| Gaussian model for CHIKV viral titer in saliva | |||||||
| Intercept | −0.1562 | 0.3079 | [−0.76, 0.447] | 0.5368 | 0.2579 | [0.031, 1.042] | |
| Viral Dissemination (legs) | 0.265 | 0.06485 | [0.138, 0.392] | 0.04749 | 0.0334 | [−0.018, 0.113] | |
| Country: Dom. Republic | 1.219 | 0.4636 | [0.311, 2.128] | (not included in the model) | |||
| Country: US | 0.2706 | 0.1868 | [−0.095, 0.637] | −0.4543 | 0.2263 | [−0.898, −0.011] | |
| 5–6 dpi | 0.02319 | 0.2652 | [−0.497, 0.543] | 0.3832 | 0.1552 | [0.079, 0.687] | |
| 12–13 dpi | 0.1753 | 0.2709 | [−0.356, 0.706] | 1.104 | 0.164 | [0.783, 1.426] | |
| Aedes albopictus | Binomial model for CHIKV saliva infection | ||||||
| Intercept | 1.278 | 0.4299 | [0.549, 2.982] | 0.653 | 0.3225 | [0.342, 1.219] | |
| Viral Dissemination (legs) | 1.396 | 0.1115 | [1.129, 1.751] | 1.035 | 0.1067 | [0.841, 1.281] | |
| Country: US | 0.314 | 0.2973 | [0.174, 0.559] | (not included in the model) | |||
| 5–6 dpi | 0.47 | 0.4101 | [0.205, 1.03] | 1.057 | 0.4683 | [0.417, 2.645] | |
| 12–13 dpi | 0.187 | 0.4255 | [0.079, 0.421] | 0.41 | 0.4248 | [0.175, 0.931] | |
| Gaussian model for CHIKV viral titer in saliva | |||||||
| Intercept | 0.3357 | 0.2489 | [−0.152, 0.823] | 0.2326 | 0.1381 | [−0.038, 0.503] | |
| Viral Dissemination (legs) | 0.2461 | 0.05738 | [0.134, 0.359] | −0.0664 | 0.05264 | [−0.17, 0.037] | |
| Country: US | −0.4006 | 0.1629 | [−0.72, −0.081] | (not included in the model) | |||
| 5–6 dpi | −0.1732 | 0.2037 | [−0.573, 0.226] | 0.5093 | 0.2159 | [0.086, 0.932] | |
| 12–13 dpi | −0.3732 | 0.2208 | [−0.806, 0.06] | 0.8416 | 0.225 | [0.401, 1.283] | |
| Species | Asian Genotype | IOL Genotype | |||||
|---|---|---|---|---|---|---|---|
| Aedes aegypti | Binomial model for CHIKV saliva infection | ||||||
| Effects | Estimate | SE | CI95 | Estimate | SE | CI95 | |
| Intercept | 0.388 | 0.3891 | [0.176, 0.818] | 1.791 | 0.4275 | [0.769, 4.139] | |
| Method: Filter Paper | 1.009 | 0.2433 | [0.626, 1.626] | 0.31 | 0.2511 | [0.188, 0.503] | |
| Method: FTA Card | 3.375 | 0.5205 | [1.275, 10.12] | (not included in the model) | |||
| Country: US | 1.817 | 0.2767 | [1.062, 3.151] | 1.255 | 0.3345 | [0.66, 2.468] | |
| 5–6 dpi | 2.207 | 0.3761 | [1.066, 4.694] | 0.511 | 0.2848 | [0.29, 0.888] | |
| 12–13 dpi | 1.232 | 0.3773 | [0.592, 2.623] | 0.456 | 0.2828 | [0.26, 0.79] | |
| Gaussian model for CHIKV viral titer in saliva | |||||||
| Intercept | 2.112 | 0.1723 | [1.774, 2.45] | 2.041 | 0.3863 | [1.284, 2.798] | |
| Method: Filter Paper | −0.02606 | 0.1221 | [−0.265, 0.213] | 7.09 × 1016 | 0.2202 | [−0.431, 0.431] | |
| Method: FTA Card | −0.01022 | 0.1454 | [−0.295, 0.275] | (not included in the model) | |||
| Country: US | 0.6995 | 0.1162 | [0.472, 0.927] | −0.807 | 0.299 | [−1.393, −0.221] | |
| 5–6 dpi | 1.722 | 0.1623 | [1.404, 2.04] | 0.2116 | 0.2527 | [−0.284, 0.707] | |
| 12–13 dpi | 1.64 | 0.1648 | [1.317, 1.963] | 0.8245 | 0.2474 | [0.34, 1.309] | |
| Aedes albopictus | Binomial model for CHIKV saliva infection | ||||||
| Intercept | 2.219 | 0.3419 | [1.148, 4.412] | 1.732 | 0.5437 | [0.604, 5.145] | |
| Method: Filter Paper | 1.662 | 0.3253 | [0.881, 3.168] | 0.417 | 0.479 | [0.158, 1.048] | |
| Method: FTA Card | 3.835 | 0.5279 | [1.427, 11.641] | (not included in the model) | |||
| Country: US | 0.309 | 0.3004 | [0.17, 0.553] | ||||
| 5–6 dpi | 0.799 | 0.3555 | [0.395, 1.6] | 0.675 | 0.5224 | [0.235, 1.849] | |
| 12–13 dpi | 0.34 | 0.3656 | [0.164, 0.692] | 0.272 | 0.4935 | [0.098, 0.69] | |
| Gaussian model for CHIKV viral titer in saliva | |||||||
| Intercept | 2.785 | 0.1863 | [2.419, 3.15] | 1.744 | 0.3994 | [0.961, 2.527] | |
| Method: Filter Paper | 0.02291 | 0.164 | [−0.298, 0.344] | 2.54 × 1015 | 0.3411 | [−0.669, 0.669] | |
| Method: FTA Card | −0.1018 | 0.1774 | [−0.45, 0.246] | (not included in the model) | |||
| Country: US | −0.2115 | 0.1418 | [−0.489, 0.066] | ||||
| 5–6 dpi | 1.332 | 0.1717 | [0.996, 1.669] | 1.188 | 0.4018 | [0.4, 1.976] | |
| 12–13 dpi | 1.391 | 0.1781 | [1.042, 1.74] | 1.341 | 0.3398 | [0.675, 2.007] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Honório, N.A.; Câmara, D.C.P.; Wiggins, K.; Eastmond, B.; Alto, B.W. High-Throughput Method for Detection of Arbovirus Infection of Saliva in Mosquitoes Aedes aegypti and Ae. albopictus. Viruses 2020, 12, 1343. https://doi.org/10.3390/v12111343
Honório NA, Câmara DCP, Wiggins K, Eastmond B, Alto BW. High-Throughput Method for Detection of Arbovirus Infection of Saliva in Mosquitoes Aedes aegypti and Ae. albopictus. Viruses. 2020; 12(11):1343. https://doi.org/10.3390/v12111343
Chicago/Turabian StyleHonório, Nildimar Alves, Daniel Cardoso Portela Câmara, Keenan Wiggins, Bradley Eastmond, and Barry Wilmer Alto. 2020. "High-Throughput Method for Detection of Arbovirus Infection of Saliva in Mosquitoes Aedes aegypti and Ae. albopictus" Viruses 12, no. 11: 1343. https://doi.org/10.3390/v12111343
APA StyleHonório, N. A., Câmara, D. C. P., Wiggins, K., Eastmond, B., & Alto, B. W. (2020). High-Throughput Method for Detection of Arbovirus Infection of Saliva in Mosquitoes Aedes aegypti and Ae. albopictus. Viruses, 12(11), 1343. https://doi.org/10.3390/v12111343