Impact of Zika Virus Infection on Human Neural Stem Cell MicroRNA Signatures

 , , , , ,
, , , , ,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
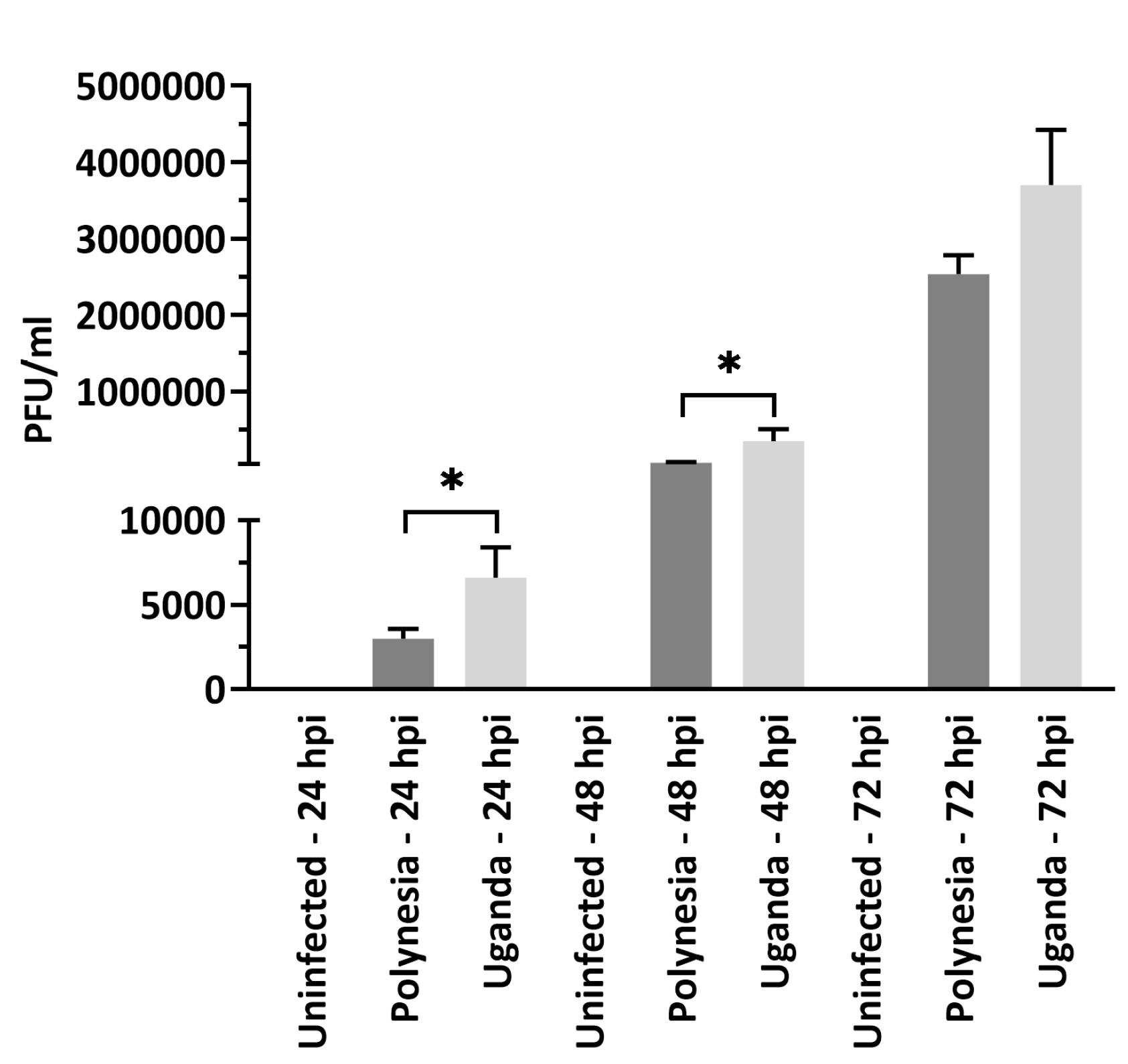
2.2. Infection
2.3. Plaque-Forming-Assay
2.4. MiRNAs: Sequencing and Data Analysis
2.5. Microarray-Based Gene Expression Analysis
2.6. Real-Time Quantitative PCR
2.7. Oxyblot
2.8. Immunofluorescence Microscopy
2.9. Statistical Analysis
3. Results
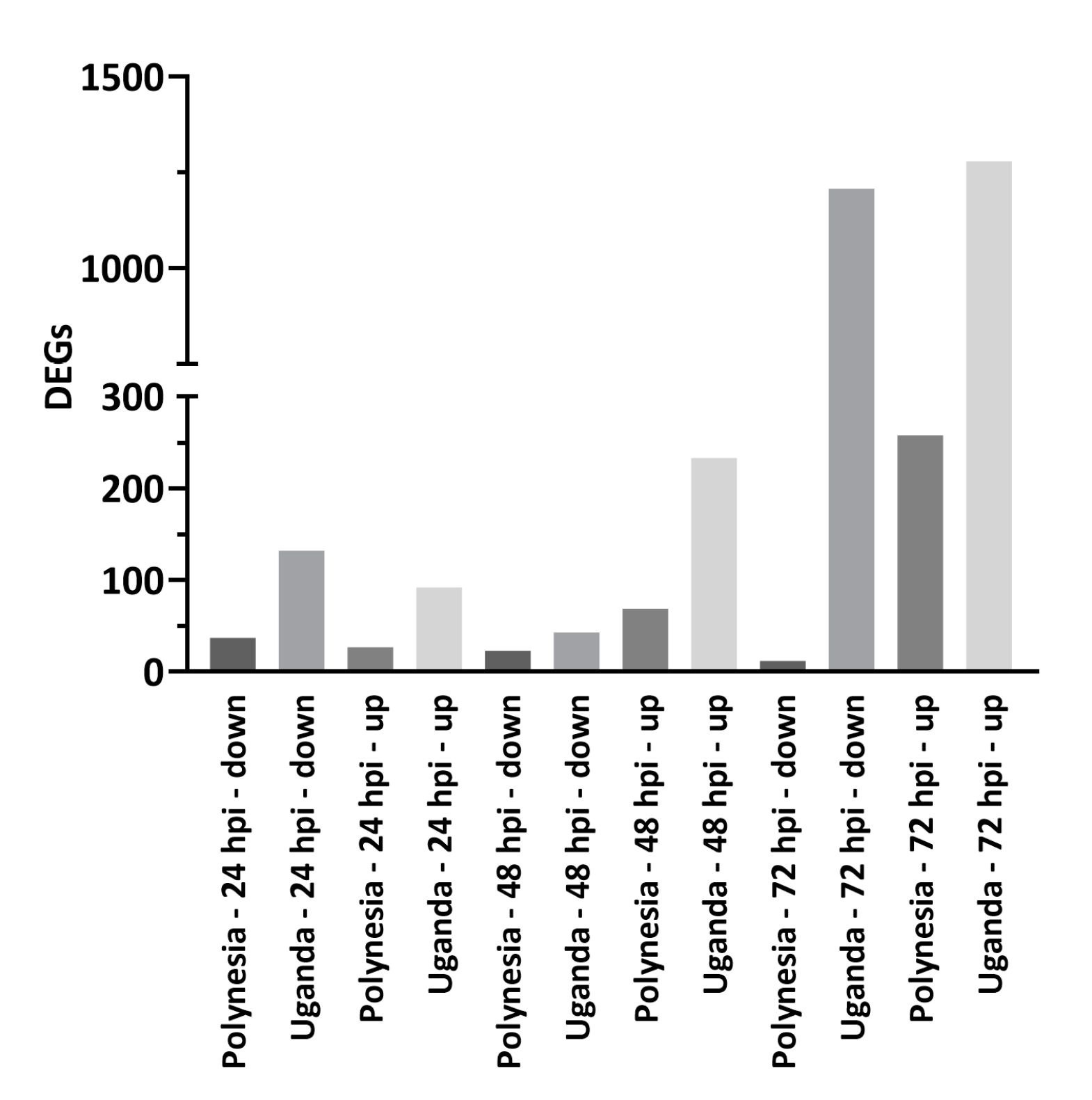
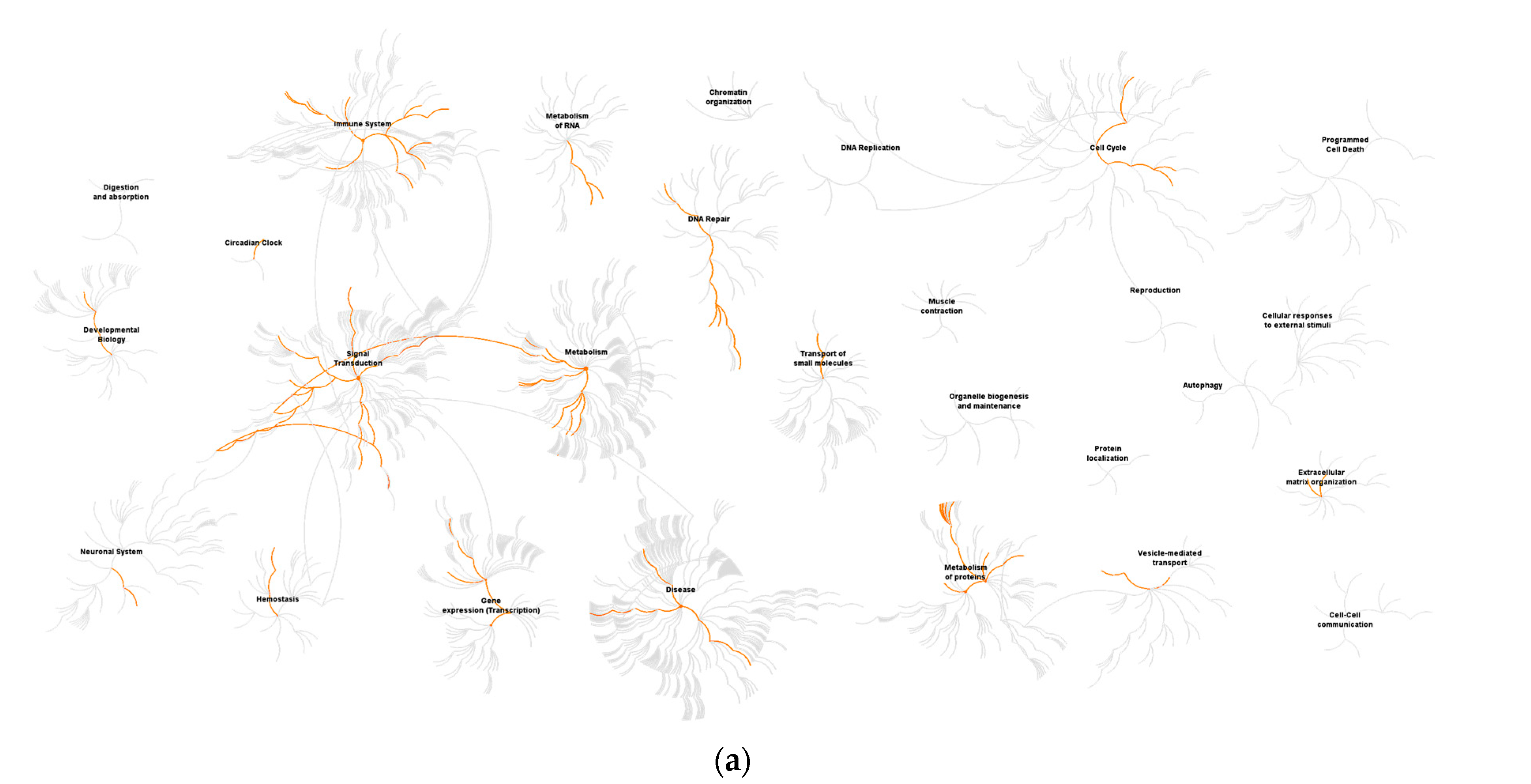
3.1. ZIKV Uganda and French Polynesia Alter Gene Expression in lt-NES® Cells
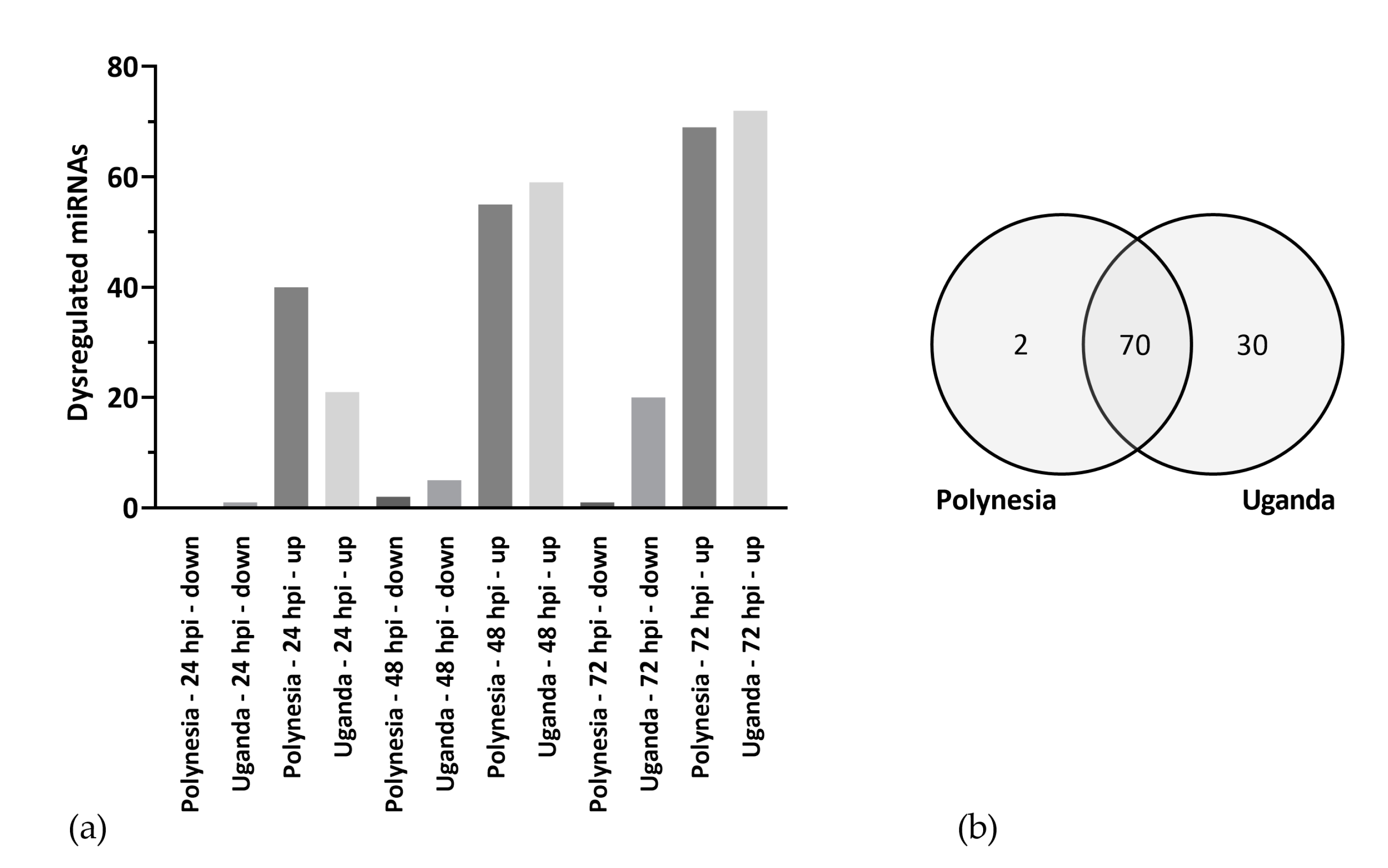
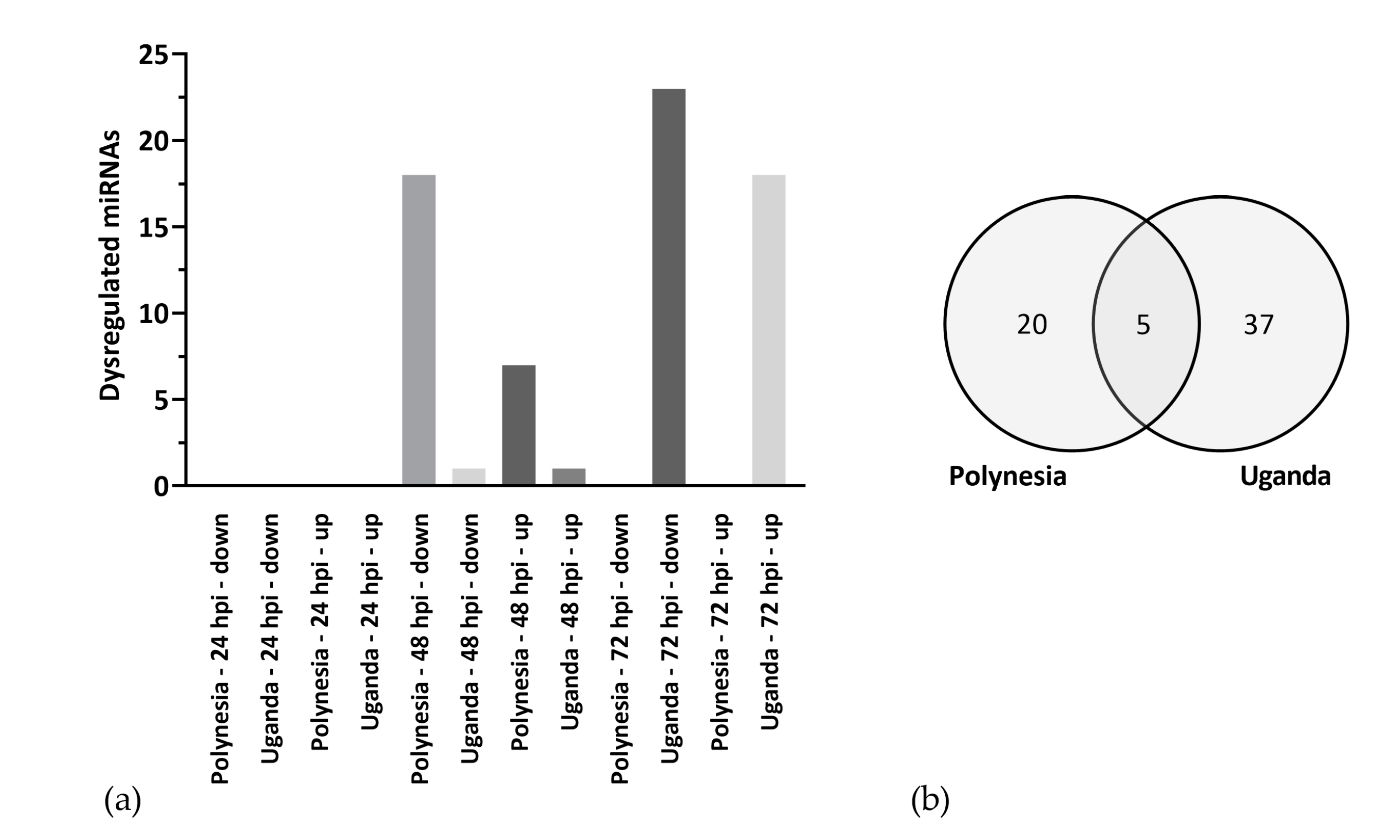
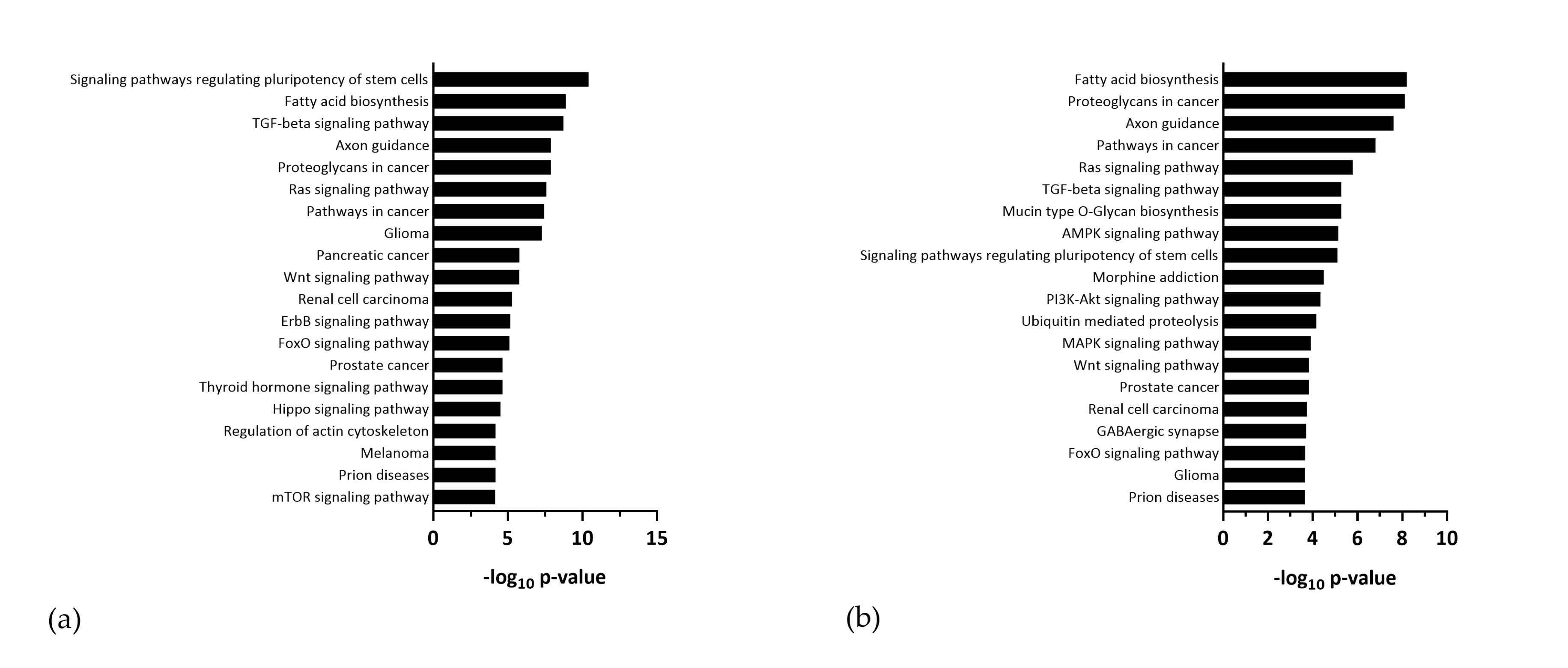
3.2. Dysregulated miRNAs Play a Role in Neurodevelopment and Cell Cycle
3.3. ZIKV Modifies the Host miRNA Cargo in Secreted EVs
3.4. Dysregulated miRNAs and mRNAs Involved in Oxidative Stress
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dick, G.W.A.; Kitchen, S.F.; Haddow, A.J. Zika Virus (I). Isolations and serological specificity. Trans. R. Soc. Trop. Med. Hyg. 1952, 46, 509–520. [Google Scholar] [CrossRef]
- Cauchemez, S.; Besnard, M.; Bompard, P.; Dub, T.; Guillemette-Artur, P.; Eyrolle-Guignot, D.; Salje, H.; Van Kerkhove, M.D.; Abadie, V.; Garel, C.; et al. Association between Zika virus and microcephaly in French Polynesia, 2013–15: A retrospective study. Lancet 2016, 387, 2125–2132. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Velez, J.O.; Lambert, A.J.; Johnson, A.J.; Stanfield, S.M.; Duffy, M.R. Genetic and Serologic Properties of Zika Virus Associated with an Epidemic, Yap State, Micronesia 2007. CDC Emerg. Infect. Dis. J. 2008, 14, 8. [Google Scholar] [CrossRef] [PubMed]
- Calvet, G.; Aguiar, R.S.; Melo, A.; A Sampaio, S.; De Filippis, I.; Fabri, A.; Araujo, E.S.M.; De Sequeira, P.C.; De Mendonça, M.C.L.; De Oliveira, L.; et al. Detection and sequencing of Zika virus from amniotic fluid of fetuses with microcephaly in Brazil: A case study. Lancet Infect. Dis. 2016, 16, 653–660. [Google Scholar] [CrossRef]
- Driggers, R.W.; Ho, C.-Y.; Korhonen, E.M.; Kuivanen, S.; Jääskeläinen, A.J.; Smura, T.; Rosenberg, A.; Hill, D.A.; DeBiasi, R.L.; Vezina, G.; et al. Zika Virus Infection with Prolonged Maternal Viremia and Fetal Brain Abnormalities. N. Engl. J. Med. 2016, 374, 2142–2151. [Google Scholar] [CrossRef] [PubMed]
- Mlakar, J.; Korva, M.; Tul, N.; Popovic, M.; Poljšak-Prijatelj, M.; Mraz, J.; Kolenc, M.; Rus, K.R.; Vipotnik, T.V.; Vodušek, V.F.; et al. Zika Virus Associated with Microcephaly. N. Engl. J. Med. 2016, 374, 951–958. [Google Scholar] [CrossRef]
- Oehler, E.; Watrin, L.; Larre, P.; Leparc-Goffart, I.; Lastère, S.; Valour, F.; Baudouin, L.; Mallet, H.P.; Musso, D.; Ghawche, F. Zika virus infection complicated by Guillain-Barré syndrome—Case report, French Polynesia, December 2013. Eurosurveillance 2014, 19, 20720. [Google Scholar] [CrossRef]
- Schuler-Faccini, L.; Ribeiro, E.M.; Feitosa, I.M.; Horovitz, D.D.; Cavalcanti, D.P.; Pessoa, A.; Doriqui, M.J.R.; Neri, J.I.; Neto, J.M.D.P.; Wanderley, H.Y.; et al. Possible Association Between Zika Virus Infection and Microcephaly—Brazil, 2015. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 59–62. [Google Scholar] [CrossRef]
- Zanluca, C.; De Melo, V.C.A.; Mosimann, A.L.P.; Dos Santos, G.I.V.; Dos Santos, C.N.D.; Luz, K. First report of autochthonous transmission of Zika virus in Brazil. Memórias Inst. Oswaldo Cruz 2015, 110, 569–572. [Google Scholar] [CrossRef]
- Zhang, Q.; Sun, K.; Chinazzi, M.; Piontti, A.P.Y.; Dean, N.E.; Rojas, D.P.; Merler, S.; Mistry, D.; Poletti, P.; Rossi, L.; et al. Spread of Zika virus in the Americas. Proc. Natl. Acad. Sci. USA 2017, 114, E4334–E4343. [Google Scholar] [CrossRef]
- Jaeger, A.S.; Murrieta, R.A.; Goren, L.R.; Crooks, C.M.; Moriarty, R.V.; Weiler, A.M.; Rybarczyk, S.; Semler, M.R.; Huffman, C.; Mejia, A.; et al. Zika viruses of African and Asian lineages cause fetal harm in a mouse model of vertical transmission. PLoS Neglected Trop. Dis. 2019, 13, e0007343. [Google Scholar] [CrossRef] [PubMed]
- Meda, N.; Salinas, S.; Kagoné, T.; Simonin, Y.; Van De Perre, P. Zika virus epidemic: Africa should not be neglected. Lancet 2016, 388, 337–338. [Google Scholar] [CrossRef]
- Heymann, D.L.; Hodgson, A.; Sall, A.A.; O Freedman, D.; Staples, J.E.; Althabe, F.; Baruah, K.; Mahmud, G.; Kandun, N.; Vasconcelos, P.F.C.; et al. Zika virus and microcephaly: Why is this situation a PHEIC? Lancet 2016, 387, 719–721. [Google Scholar] [CrossRef]
- Dang, J.; Tiwari, S.K.; Lichinchi, G.; Qin, Y.; Patil, V.S.; Eroshkin, A.M.; Rana, T.M. Zika Virus Depletes Neural Progenitors in Human Cerebral Organoids through Activation of the Innate Immune Receptor TLR3. Cell Stem Cell 2016, 19, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Xu, D.; Ye, Q.; Hong, S.; Jiang, Y.; Liu, X.; Zhang, N.; Shi, L.; Qin, C.-F.; Xu, Z. Zika Virus Disrupts Neural Progenitor Development and Leads to Microcephaly in Mice. Cell Stem Cell 2016, 19, 672. [Google Scholar] [CrossRef]
- Ming, G.-L.; Tang, H.; Song, H. Advances in Zika Virus Research: Stem Cell Models, Challenges, and Opportunities. Cell Stem Cell 2016, 19, 690–702. [Google Scholar] [CrossRef]
- Tang, H.; Hammack, C.; Ogden, S.C.; Wen, Z.; Qian, X.; Li, Y.; Yao, B.; Shin, J.; Zhang, F.; Lee, E.M.; et al. Zika Virus Infects Human Cortical Neural Progenitors and Attenuates Their Growth. Cell Stem Cell 2016, 18, 587–590. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef]
- Fu, G.; Brkić, J.; Hayder, H.; Peng, C. MicroRNAs in Human Placental Development and Pregnancy Complications. Int. J. Mol. Sci. 2013, 14, 5519–5544. [Google Scholar] [CrossRef]
- Liu, P.; Pu, J.; Zhang, J.; Chen, Z.; Wei, K.; Shi, L. Bioinformatic analysis of miR-4792 regulates Radix Tetrastigma hemsleyani flavone to inhibit proliferation, invasion, and induce apoptosis of A549 cells. Onco Targets Ther. 2019, 12, 1401–1412. [Google Scholar] [CrossRef]
- Yao, S. MicroRNA biogenesis and their functions in regulating stem cell potency and differentiation. Biol. Proced. Online 2016, 18, 8. [Google Scholar] [CrossRef]
- Gurunathan, S.; Kang, M.-H.; Jeyaraj, M.; Qasim, M.; Kim, J.-H. Review of the Isolation, Characterization, Biological Function, and Multifarious Therapeutic Approaches of Exosomes. Cells 2019, 8, 307. [Google Scholar] [CrossRef] [PubMed]
- Devhare, P.B.; Sasaki, R.; Shrivastava, S.; Di Bisceglie, A.M.; Ray, R.; Ray, R.B. Exosome-Mediated Intercellular Communication between Hepatitis C Virus-Infected Hepatocytes and Hepatic Stellate Cells. J. Virol. 2017, 91, e02225-16. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Akbar, I.; Kumari, B.; Vrati, S.; Basu, A.; Banerjee, A. Japanese Encephalitis Virus-induced let-7a/b interacted with the NOTCH—TLR 7 pathway in microglia and facilitated neuronal death via caspase activation. J. Neurochem. 2019, 149, 518–534. [Google Scholar] [CrossRef] [PubMed]
- Dang, J.W.; Tiwari, S.K.; Qin, Y.; Rana, T.M. Genome-wide Integrative Analysis of Zika-Virus-Infected Neuronal Stem Cells Reveals Roles for MicroRNAs in Cell Cycle and Stemness. Cell Rep. 2019, 27, 3618–3628. [Google Scholar] [CrossRef]
- Kozak, R.A.; Majer, A.; Biondi, M.J.; Medina, S.J.; Goneau, L.W.; Sajesh, B.V.; Slota, J.A.; Zubach, V.; Severini, A.; Safronetz, D.; et al. MicroRNA and mRNA Dysregulation in Astrocytes Infected with Zika Virus. Viruses 2017, 9, 297. [Google Scholar] [CrossRef]
- Valadão, A.L.C.; Aguiar, R.S.; De Arruda, L.B. Interplay between Inflammation and Cellular Stress Triggered by Flaviviridae Viruses. Front. Microbiol. 2016, 7, 1233. [Google Scholar] [CrossRef]
- Kobayashi, M.; Li, L.; Iwamoto, N.; Nakajima-Takagi, Y.; Kaneko, H.; Nakayama, Y.; Eguchi, M.; Wada, Y.; Kumagai, Y.; Yamamoto, M. The Antioxidant Defense System Keap1-Nrf2 Comprises a Multiple Sensing Mechanism for Responding to a Wide Range of Chemical Compounds. Mol. Cell. Biol. 2008, 29, 493–502. [Google Scholar] [CrossRef]
- Ungvari, Z.; Csiszar, A. Resveratrol Confers Endothelial Protection in Insulin-Dependent Diabetes Mellitus. Cardiovasc. Drugs Ther. 2011, 25, 111–113. [Google Scholar] [CrossRef]
- Yamamoto, M.; Kensler, T.W.; Motohashi, H. The KEAP1-NRF2 System: A Thiol-Based Sensor-Effector Apparatus for Maintaining Redox Homeostasis. Physiol. Rev. 2018, 98, 1169–1203. [Google Scholar] [CrossRef]
- Bender, D.; Hildt, E. Effect of Hepatitis Viruses on the Nrf2/Keap1-Signaling Pathway and Its Impact on Viral Replication and Pathogenesis. Int. J. Mol. Sci. 2019, 20, 4659. [Google Scholar] [CrossRef]
- Carvajal-Yepes, M.; Himmelsbach, K.; Schaedler, S.; Ploen, D.; Krause, J.; Ludwig, L.; Weiss, T.S.; Klingel, K.; Hildt, E. Hepatitis C Virus Impairs the Induction of Cytoprotective Nrf2 Target Genes by Delocalization of Small Maf Proteins. J. Biol. Chem. 2011, 286. [Google Scholar] [CrossRef]
- Zhang, Z.; Rong, L.; Li, Y.-P. Flaviviridae Viruses and Oxidative Stress: Implications for Viral Pathogenesis. Oxid. Med. Cell. Longev. 2019, 2019, 1409582. [Google Scholar] [CrossRef] [PubMed]
- Koch, P.; Opitz, T.; Steinbeck, J.A.; Ladewig, J.; Brustle, O. A rosette-type, self-renewing human ES cell-derived neural stem cell with potential for in vitro instruction and synaptic integration. Proc. Natl. Acad. Sci. USA 2009, 106, 3225–3230. [Google Scholar] [CrossRef] [PubMed]
- Falk, A.; Koch, P.; Kesavan, J.; Takashima, Y.; Ladewig, J.; Alexander, M.; Wiskow, O.; Tailor, J.; Trotter, M.; Pollard, S.; et al. Capture of Neuroepithelial-Like Stem Cells from Pluripotent Stem Cells Provides a Versatile System for In Vitro Production of Human Neurons. PLoS ONE 2012, 7, e29597. [Google Scholar] [CrossRef] [PubMed]
- Rehbach, K.; Kesavan, J.; Hauser, S.; Ritzenhofen, S.; Jungverdorben, J.; Schüle, R.; Schöls, L.; Peitz, M.; Brüstle, O. Multiparametric rapid screening of neuronal process pathology for drug target identification in HSP patient-specific neurons. Sci. Rep. 2019, 9, 9615. [Google Scholar] [CrossRef] [PubMed]
- Tenoever, B.R. RNA viruses and the host microRNA machinery. Nat. Rev. Genet. 2013, 11, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Vlachos, I.S.; Zagganas, K.; Paraskevopoulou, M.D.; Georgakilas, G.; Karagkouni, D.; Vergoulis, T.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA-miRPath v3.0: Deciphering microRNA function with experimental support. Nucleic Acids Res. 2015, 43, W460–W466. [Google Scholar] [CrossRef]
- Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Reczko, M.; Maragkakis, M.; Alexiou, P.; Grosse, I.; Hatzigeorgiou, A.G. Functional microRNA targets in protein coding sequences. Bioinformatics 2012, 28, 771–776. [Google Scholar] [CrossRef]
- Agarwal, V.; Bell, G.W.; Nam, J.-W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4, e05005. [Google Scholar] [CrossRef] [PubMed]
- Krek, A.; Grün, D.; Poy, M.N.; Wolf, R.; Rosenberg, L.; Epstein, E.J.; MacMenamin, P.; Da Piedade, I.; Gunsalus, K.C.; Stoffel, M.; et al. Combinatorial microRNA target predictions. Nat. Genet. 2005, 37, 495–500. [Google Scholar] [CrossRef]
- Sethupathy, P.; Corda, B.; Hatzigeorgiou, A.G. TarBase: A comprehensive database of experimentally supported animal microRNA targets. RNA 2005, 12, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Wang, X. miRDB: A microRNA target prediction and functional annotation database with a wiki interface. RNA 2008, 14, 1012–1017. [Google Scholar] [CrossRef] [PubMed]
- Croft, D.; Mundo, A.F.; Haw, R.; Milacic, M.; Weiser, J.; Wu, G.; Caudy, M.; Garapati, P.; Gillespie, M.; Kamdar, M.R.; et al. The Reactome pathway knowledgebase. Nucleic Acids Res. 2013, 42, D472–D477. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Elgner, F.; Sabino, C.; Basic, M.; Ploen, D.; Grünweller, A.; Hildt, E. Inhibition of Zika Virus Replication by Silvestrol. Viruses 2018, 10, 149. [Google Scholar] [CrossRef]
- Davis, A.P.; Grondin, C.J.; Johnson, R.J.; Sciaky, D.; King, B.L.; McMorran, R.; Wiegers, J.; Wiegers, T.C.; Mattingly, C.J. The Comparative Toxicogenomics Database: Update 2017. Nucleic Acids Res. 2016, 45, D972–D978. [Google Scholar] [CrossRef]
- Li, M.; Wang, P.; Zheng, Z.; Hu, K.; Zhang, M.; Guan, X.; Fu, M.; Zhang, D.; Wang, W.; Xiao, G.; et al. Japanese encephalitis virus counteracts BST2 restriction via its envelope protein E. Virology 2017, 510, 67–75. [Google Scholar] [CrossRef]
- Cho, H.; Shrestha, B.; Sen, G.C.; Diamond, M.S. A Role for Ifit2 in Restricting West Nile Virus Infection in the Brain. J. Virol. 2013, 87, 8363–8371. [Google Scholar] [CrossRef]
- Xie, H.; Zhao, Y.; Caramuta, S.; Larsson, C.; Lui, W.-O. miR-205 Expression Promotes Cell Proliferation and Migration of Human Cervical Cancer Cells. PLoS ONE 2012, 7, e46990. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, X. miR-4792 inhibits epithelial–mesenchymal transition and invasion in nasopharyngeal carcinoma by targeting FOXC1. Biochem. Biophys. Res. Commun. 2015, 468, 863–869. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.-T.; Liu, S.-Y.; Leu, J.-D.; Chang, C.-Y.; Chiou, S.-H.; Lee, T.-C.; Lee, Y.-J. Arsenic trioxide-mediated suppression of miR-182-5p is associated with potent anti-oxidant effects through up-regulation of SESN2. Oncotarget 2018, 9, 16028–16042. [Google Scholar] [CrossRef] [PubMed]
- Lowey, B.; Hertz, L.; Chiu, S.; Valdez, K.; Li, Q.; Liang, T.J.; Weinman, S.; Saito, T. Hepatitis C Virus Infection Induces Hepatic Expression of NF-κB-Inducing Kinase and Lipogenesis by Downregulating miR-122. mBio 2019, 10, 19. [Google Scholar] [CrossRef]
- Zhu, X.; He, Z.; Hu, Y.; Wen, W.; Lin, C.; Yu, J.; Pan, J.; Li, R.; Deng, H.; Liao, S.; et al. MicroRNA-30e* Suppresses Dengue Virus Replication by Promoting NF-κB–Dependent IFN Production. PLoS Negl. Trop. Dis. 2014, 8, e3088. [Google Scholar] [CrossRef]
- Azouz, F.; Arora, K.; Krause, K.; Nerurkar, V.R.; Kumar, M. Integrated MicroRNA and mRNA Profiling in Zika Virus-Infected Neurons. Viruses 2019, 11, 162. [Google Scholar] [CrossRef]
- Nutt, C.; Adams, P. Zika in Africa—The invisible epidemic? Lancet 2017, 389, 1595–1596. [Google Scholar] [CrossRef]
- Simonin, Y.; Loustalot, F.; Desmetz, C.; Foulongne, V.; Constant, O.; Fournier-Wirth, C.; Leon, F.; Molès, J.-P.; Goubaud, A.; Lemaitre, J.-M.; et al. Zika Virus Strains Potentially Display Different Infectious Profiles in Human Neural Cells. EBioMedicine 2016, 12, 161–169. [Google Scholar] [CrossRef]
- Sheridan, M.A.; Balaraman, V.; Schust, D.J.; Ezashi, T.; Roberts, R.M.; Franz, A.W.E. African and Asian strains of Zika virus differ in their ability to infect and lyse primitive human placental trophoblast. PLoS ONE 2018, 13, e0200086. [Google Scholar] [CrossRef]
- Simonin, Y.; van Riel, D.; Van de Perre, P.; Rockx, B.; Salinas, S. Differential virulence between Asian and African lineages of Zika virus. PLoS Negl. Trop. Dis. 2017, 11. [Google Scholar] [CrossRef]
- Anfasa, F.; Siegers, J.Y.; Van Der Kroeg, M.; Mumtaz, N.; Raj, V.S.; De Vrij, F.M.S.; Widagdo, W.; Gabriel, G.; Salinas, S.; Simonin, Y.; et al. Phenotypic Differences between Asian and African Lineage Zika Viruses in Human Neural Progenitor Cells. mSphere 2017, 2, e00292-17. [Google Scholar] [CrossRef] [PubMed]
- Lomelino, C.L.; Andring, J.T.; McKenna, R.; Kilberg, M.S. Asparagine synthetase: Function, structure, and role in disease. J. Biol. Chem. 2017, 292, 19952–19958. [Google Scholar] [CrossRef]
- Schleinitz, D.; Seidel, A.; Stassart, R.; Klammt, J.; Hirrlinger, P.G.; Winkler, U.; Köhler, S.; Heiker, J.T.; Schönauer, R.; Bialek, J.; et al. Novel Mutations in the Asparagine Synthetase Gene (ASNS) Associated With Microcephaly. Front. Genet. 2018, 9, 245. [Google Scholar] [CrossRef] [PubMed]
- Richards, N.G.J.; Kilberg, M.S. Asparagine Synthetase Chemotherapy. Annu. Rev. Biochem. 2006, 75, 629–654. [Google Scholar] [CrossRef]
- Slonchak, A.; Clarke, B.; MacKenzie, J.; Amarilla, A.A.; Setoh, Y.X.; Khromykh, A.A. West Nile virus infection and interferon alpha treatment alter the spectrum and the levels of coding and noncoding host RNAs secreted in extracellular vesicles. BMC Genom. 2019, 20, 474. [Google Scholar] [CrossRef]
- Yoshikawa, F.S.Y.; Teixeira, F.M.E.; Sato, M.N.; Oliveira, L.M.S. Delivery of microRNAs by Extracellular Vesicles in Viral Infections: Could the News be Packaged? Cells 2019, 8, 611. [Google Scholar] [CrossRef]
- A Aldinger, K.; Lehmann, O.J.; Hudgins, L.; Chizhikov, V.V.; Bassuk, A.G.; Ades, L.C.; Krantz, I.D.; Dobyns, W.B.; Millen, K.J. FOXC1 is required for normal cerebellar development and is a major contributor to chromosome 6p25.3 Dandy-Walker malformation. Nat. Genet. 2009, 41, 1037–1042. [Google Scholar] [CrossRef] [PubMed]
- Haldipur, P.; Dang, D.; Aldinger, K.A.; Janson, O.K.; Guimiot, F.; Adle-Biasette, H.; Dobyns, W.B.; Siebert, J.R.; Russo, R.; Millen, K.J. Phenotypic outcomes in Mouse and Human Foxc1 dependent Dandy-Walker cerebellar malformation suggest shared mechanisms. eLife 2017, 6, 321. [Google Scholar] [CrossRef]
- Pool, K.-L.; Adachi, K.; Karnezis, S.; Salamon, N.; Romero, T.; Nielsen-Saines, K.; Pone, S.; Boechat, M.; Aibe, M.; Da Silva, T.G.; et al. Association Between Neonatal Neuroimaging and Clinical Outcomes in Zika-Exposed Infants From Rio de Janeiro, Brazil. JAMA Netw. Open 2019, 2, e198124. [Google Scholar] [CrossRef]
- Ito, Y.A.; Goping, I.S.; Berry, F.; Walter, M.A. Dysfunction of the stress-responsive FOXC1 transcription factor contributes to the earlier-onset glaucoma observed in Axenfeld-Rieger syndrome patients. Cell Death Dis. 2014, 5, e1069. [Google Scholar] [CrossRef]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24, 1583. [Google Scholar] [CrossRef] [PubMed]
- Medvedev, R.; Ploen, D.; Spengler, C.; Elgner, F.; Ren, H.; Bunten, S.; Hildt, E. HCV-induced oxidative stress by inhibition of Nrf2 triggers autophagy and favors release of viral particles. Free Radic. Biol. Med. 2017, 110, 300–315. [Google Scholar] [CrossRef] [PubMed]
- Olagnier, D.; Peri, S.; Steel, C.; Van Montfoort, N.; Chiang, C.; Beljanski, V.; Slifker, M.; He, Z.; Nichols, C.N.; Lin, R.; et al. Cellular Oxidative Stress Response Controls the Antiviral and Apoptotic Programs in Dengue Virus-Infected Dendritic Cells. PLoS Pathog. 2014, 10, e1004566. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Bowman, J.W.; Jung, J.U. Autophagy during viral infection—A double-edged sword. Nat. Rev. Genet. 2018, 16, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Dreux, M.; Chisari, F.V. Viruses and the autophagy machinery. Cell Cycle 2010, 9, 1295–1307. [Google Scholar] [CrossRef] [PubMed]
- Itoha, K.; Chibabc, T.; Takahashia, S.; Ishiia, T.; Igarashia, K.; Katoha, Y.; Oyaked, T.; Hayashid, N.; Satohe, K.; Hatayamae, I.; et al. An Nrf2/Small Maf Heterodimer Mediates the Induction of Phase II Detoxifying Enzyme Genes through Antioxidant Response Elements. Biochem. Biophys. Res. Commun. 1997, 236, 313–322. [Google Scholar] [CrossRef]
- Rushworth, S.A.; MacEwan, D.; O’Connell, M.A. Lipopolysaccharide-induced expression of NAD(P)H:quinone oxidoreductase 1 and heme oxygenase-1 protects against excessive inflammatory responses in human monocytes. J. Immunol. 2008, 181, 6730–6737. [Google Scholar] [CrossRef]
- Vasiliou, V.; Ross, D.; Nebert, D.W. Update of the NAD(P)H:quinone oxidoreductase (NQO) gene family. Hum. Genom. 2006, 2, 329–335. [Google Scholar] [CrossRef]
- Shin, B.Y.; Jin, S.H.; Cho, I.J.; Ki, S.H. Nrf2-ARE pathway regulates induction of Sestrin-2 expression. Free Radic. Biol. Med. 2012, 53, 834–841. [Google Scholar] [CrossRef]
- Ding, B.; Parmigiani, A.; Divakaruni, A.S.; Archer, K.; Murphy, A.N.; Budanov, A.V. Sestrin2 is induced by glucose starvation via the unfolded protein response and protects cells from non-canonical necroptotic cell death. Sci. Rep. 2016, 6, 22538. [Google Scholar] [CrossRef]
- Jegal, K.H.; Park, S.M.; Cho, S.S.; Byun, S.H.; Ku, S.-K.; Kim, S.C.; Ki, S.H.; Cho, I.J. Activating transcription factor 6-dependent sestrin 2 induction ameliorates ER stress-mediated liver injury. Biochim. Biophys. Acta Bioenerg. 2017, 1864, 1295–1307. [Google Scholar] [CrossRef] [PubMed]
- Carr, M.; Gonzalez, G.; Martinelli, A.; Wastika, C.E.; Ito, K.; Orba, Y.; Sasaki, M.; Hall, W.W.; Sawa, H. Upregulated expression of the antioxidant sestrin 2 identified by transcriptomic analysis of Japanese encephalitis virus-infected SH-SY5Y neuroblastoma cells. Virus Genes 2019, 55, 630–642. [Google Scholar] [CrossRef] [PubMed]
- Pasha, M.; Eid, A.H.; Eid, A.A.; Gorin, Y.; Munusamy, S. Sestrin2 as a Novel Biomarker and Therapeutic Target for Various Diseases. Available online: https://www.hindawi.com/journals/omcl/2017/3296294/ (accessed on 3 June 2020).
- Soontornniyomkij, V.; Soontornniyomkij, B.; Moore, D.J.; Gouaux, B.; Masliah, E.; Tung, S.; Vinters, H.V.; Grant, I.; Achim, C.L. Antioxidant sestrin-2 redistribution to neuronal soma in human immunodeficiency virus-associated neurocognitive disorders. J. Neuroimmune Pharmacol. 2012, 7, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Xiao, T.; Zhang, L.; Huang, Y.; Shi, Y.; Wang, J.; Ji, Q.; Ye, J.; Lin, Y.; Liu, H. Sestrin2 increases in aortas and plasma from aortic dissection patients and alleviates angiotensin II-induced smooth muscle cell apoptosis via the Nrf2 pathway. Life Sci. 2019, 218, 132–138. [Google Scholar] [CrossRef]
- Ye, J.; Wang, M.; Xu, Y.; Liu, J.; Jiang, H.; Wang, Z.; Lin, Y.; Wan, J. Sestrins increase in patients with coronary artery disease and associate with the severity of coronary stenosis. Clin. Chim. Acta 2017, 472, 51–57. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GeneSymbol | Fold Change ZIKV Uganda | p-Value ZIKV Uganda | Fold Change ZIKV Polynesia | p-Value ZIKV Polynesia |
|---|---|---|---|---|
| COL4A1 | −2.1 | 0.0093 | −1.2 | 0.635 |
| MIR17HG | −2.3 | 0.0497 | −1.3 | 0.749 |
| TUBA1A | −2.2 | 2.56 × 10−4 | −1.5 | 0.256 |
| SLC2A1 | −2.3 | 0.0239 | 1.0 | 0.806 |
| ASNS | 9.6 | 0.0314 | 2.9 | 0.006 |
| STAMBP | 2.5 | 1.07 × 10−4 | 1.4 | 0.975 |
| GeneSymbol | Fold Change ZIKV Uganda 24 hpi | Fold Change ZIKV Uganda 48 hpi | Fold Change ZIKV Uganda 72 hpi |
|---|---|---|---|
| OASL | 1.6 | 49.9 | 766.7 |
| CH25H | 2.5 | 46.8 | 611.3 |
| IFNB1 | 2.3 | 37.9 | 554.3 |
| CXCL10 | −1.3 | 40.8 | 521.3 |
| IFIT2 | 1.0 | 30.7 | 218.8 |
| IFIH1 | 2.1 | 38.1 | 176.4 |
| PRDM16 | 8.1 | 23.5 | 141.9 |
| CCL5 | −1.3 | 10.3 | 117.1 |
| CXCL11 | 1.1 | 12.8 | 104.1 |
| GSC | 1.2 | 9.0 | 60.4 |
| BST2 | 1.8 | 26.6 | 59.0 |
| IFI44 | 2.4 | 20.8 | 55.5 |
| PKD1L2 | 2.8 | 9.6 | 49.9 |
| DHX58 | 1.3 | 8.3 | 46.3 |
| ATP4A | 1.1 | 3.7 | 38.3 |
| TMEM71 | 2.5 | 8.7 | 35.8 |
| CEACAM1 | 1.4 | 6.8 | 35.8 |
| NEURL3 | 1.2 | 10.5 | 34.0 |
| IFI6 | 1.0 | 9.1 | 33.8 |
| GeneSymbol | Fold Change ZIKV Polynesia 24 hpi | Fold Change ZIKV Polynesia 48 hpi | Fold Change ZIKV Polynesia 72 hpi |
|---|---|---|---|
| BST2 | 1.2 | 10.2 | 459.5 |
| OAS1 | −1.0 | 2.7 | 261.2 |
| IFIH1 | 1.5 | 11.8 | 219.1 |
| IFNB1 | 1.4 | 4.3 | 211.2 |
| OASL | −1.1 | 6.3 | 193.1 |
| CXCL10 | 1.0 | 4.2 | 147.1 |
| IFITM1 | 1.4 | 4.8 | 133.1 |
| HERC6 | 1.1 | 3.2 | 130.7 |
| CH25H | 1.5 | 3.0 | 126.7 |
| IFI44 | 2.1 | 11.6 | 124.7 |
| IFI6 | −1.0 | 3.7 | 118.3 |
| SAMD9L | −1.0 | 6.2 | 105.6 |
| IFIT2 | 1.3 | 5.1 | 96.8 |
| MX1 | 1.1 | 4.3 | 90.2 |
| ISG15 | −1.0 | 5.1 | 67.5 |
| OAS2 | −1.1 | 2.4 | 54.4 |
| DDX60 | 1.0 | 4.3 | 52.0 |
| GO Category | p-Value | #Genes |
|---|---|---|
| Gene expression | 2.49 × 10−5 | 12 |
| Cellular nitrogen compound metabolic process | 0.00013 | 37 |
| Macromolecular complex assembly | 0.00162 | 13 |
| Protein complex assembly | 0.01012 | 11 |
| Cellular component assembly | 0.01986 | 14 |
| Biosynthetic process | 0.02673 | 28 |
| Biological process | 0.02901 | 90 |
| Negative regulation of oxidoreductase activity | 0.03821 | 2 |
| In utero embryonic development | 0.03821 | 7 |
| DEG | Fold Change ZIKV Polynesia |
|---|---|
| PNPT1 | 3.36 |
| FOS | 5.18 |
| XBP1 | 2.06 |
| PPARGC1A | 2.06 |
| KLF4 | 6.28 |
| PML | 3.76 |
| TNFAIP3 | 4.96 |
| STAT1 | 17.09 |
| SLC7A11 | 2.11 |
| JUN | 2.10 |
| DEG | Fold Change ZIKV Polynesia |
|---|---|
| XBP1 | 2.06 |
| EIF2AK2 | 4.72 |
| ASNS | 2.13 |
| TRIB3 | 2.84 |
| DDIT3 | 5.17 |
| ATF3 | 5.56 |
| HERPUD1 | 2.48 |
| PPP1R15A | 2.31 |
| FBXO6 | 2.33 |
| CHAC1 | 4.46 |
| miRNA ID | Gene Targets (ID) |
|---|---|
| hsa-miR-26b-5p | TRIB3, FOS, HERPUD1, TNFAIP3, JUN, ASNS, CHAC1, ATF3 |
| hsa-let-7g-5p | TRIB3, FBXO6, CHAC1 |
| hsa-miR-17-5p | FOS, TNFAIP3, JUN, KLF4 |
| hsa-miR-221-3p | FOS, PPP1R15A |
| hsa-miR-16-5p | HERPUD1, PML, EIF2AK2, JUN, ASNS, PPP1R15A, CHAC1, STAT1 |
| hsa-miR-106b-5p | TNFAIP3 |
| hsa-miR-374a-5p | TNFAIP3, EIF2AK2 |
| hsa-miR-93-5p | TNFAIP3, JUN |
| hsa-miR-20a-5p | TNFAIP3 |
| hsa-miR-186-5p | EIF2AK2, JUN, ASNS, CHAC1 |
| hsa-miR-32-5p | JUN, KLF4 |
| hsa-miR-218-5p | CHAC1 |
| hsa-miR-340-5p | PPARGC1A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tabari, D.; Scholl, C.; Steffens, M.; Weickhardt, S.; Elgner, F.; Bender, D.; Herrlein, M.-L.; Sabino, C.; Semkova, V.; Peitz, M.; et al. Impact of Zika Virus Infection on Human Neural Stem Cell MicroRNA Signatures. Viruses 2020, 12, 1219. https://doi.org/10.3390/v12111219
Tabari D, Scholl C, Steffens M, Weickhardt S, Elgner F, Bender D, Herrlein M-L, Sabino C, Semkova V, Peitz M, et al. Impact of Zika Virus Infection on Human Neural Stem Cell MicroRNA Signatures. Viruses. 2020; 12(11):1219. https://doi.org/10.3390/v12111219
Chicago/Turabian StyleTabari, Denna, Catharina Scholl, Michael Steffens, Sandra Weickhardt, Fabian Elgner, Daniela Bender, Marie-Luise Herrlein, Catarina Sabino, Vesselina Semkova, Michael Peitz, and et al. 2020. "Impact of Zika Virus Infection on Human Neural Stem Cell MicroRNA Signatures" Viruses 12, no. 11: 1219. https://doi.org/10.3390/v12111219
APA StyleTabari, D., Scholl, C., Steffens, M., Weickhardt, S., Elgner, F., Bender, D., Herrlein, M.-L., Sabino, C., Semkova, V., Peitz, M., Till, A., Brüstle, O., Hildt, E., & Stingl, J. (2020). Impact of Zika Virus Infection on Human Neural Stem Cell MicroRNA Signatures. Viruses, 12(11), 1219. https://doi.org/10.3390/v12111219