A Metabolomics Approach to Unravel Cricket Paralysis Virus Infection in Silkworm Bm5 Cells

 , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Extraction of Intracellular Metabolites
2.3. Ultrahigh Performance Liquid Chromatography Hyphenated to Quadrupole-Orbitrap High-Resolution Mass Spectrometry (UHPLC-Q-Orbitrap-HRMS) Analysis
2.4. Data Analysis
2.4.1. Untargeted Data Analysis
2.4.2. Targeted Data Analysis
3. Results
3.1. Global Metabolic Changes in Bm5 Cells Following CrPV Infection
3.2. Metabolites in Bm5 Cells Following CrPV Infection
3.3. Modified Metabolic Pathways in CrPV-Infected Bm5 Cells
3.4. Differential Abundance Analysis of Identified Metabolites
3.5. Key Metabolites during CrPV Infection in Bm5 Cells
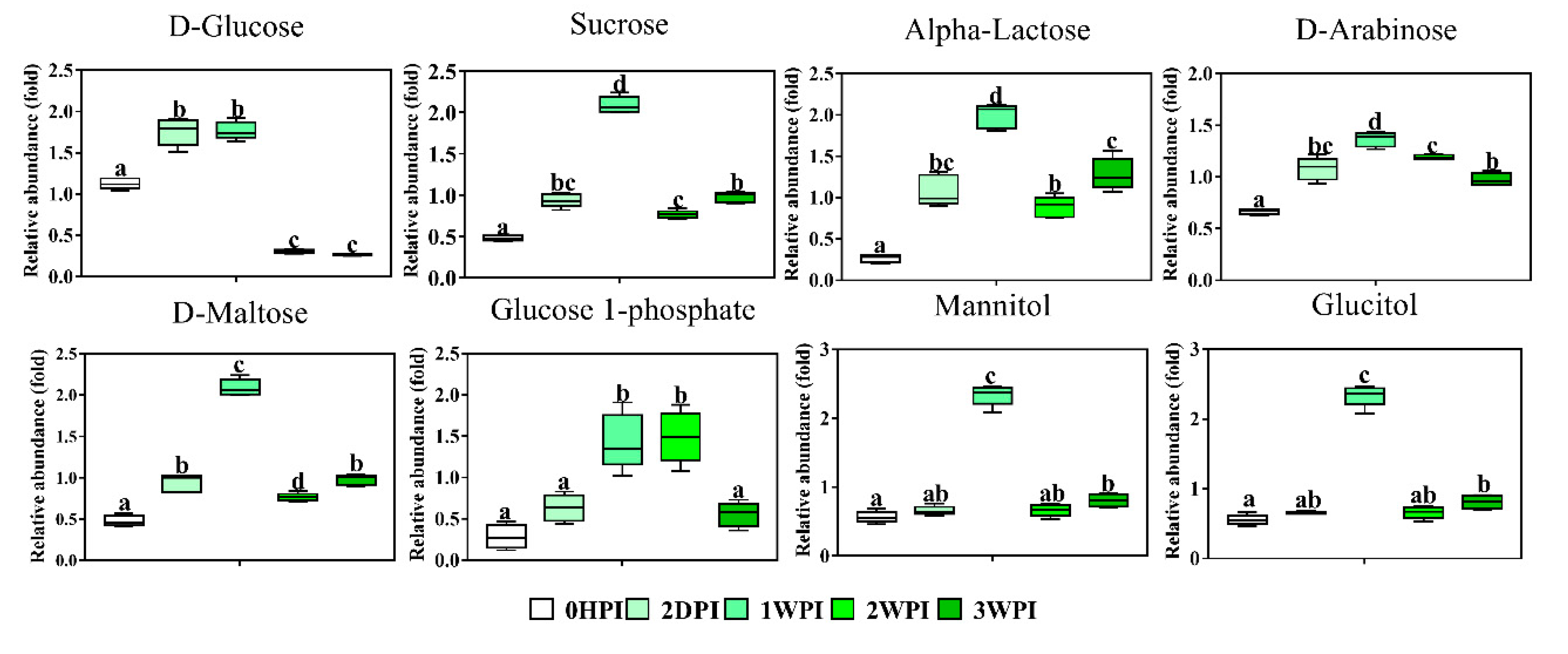
3.5.1. Glucose and Glutamine
3.5.2. Citric Acid
3.5.3. Phosphoenolpyruvate
3.5.4. Vitamins
3.5.5. Purine and Pyrimidine Metabolism
4. Discussion
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Hails, R.S.; Ball, B.V.; Genersch, E. Infection strategies of insect viruses. In Virology and the Honey Bee; Aubert, M., Ball, B., Fries, I., Moritz, R., Milani, N., Bernardinelli, I., Eds.; Office for Official Publications of the European Communities: Luxembourg, 2008; pp. 255–276. [Google Scholar]
- Dimmock, N.J.; Primrose, S.B. Introduction to Modern Virology. In Basic Microbiology; Willey Blackwell: Hoboken, NJ, USA, 2007. [Google Scholar]
- Cory, J.S. Insect virus transmission: Different routes to persistence. Curr. Opin. Insect Sci. 2015, 8, 130–135. [Google Scholar] [CrossRef]
- Goic, B.; Vodovar, N.; Mondotte, J.A.; Monot, C.; Frangeul, L.; Blanc, H.; Gausson, V.; Vera Otarola, J.; Cristofari, G.; Saleh, M.C. RNA-mediated interference and reverse transcription control the persistence of RNA viruses in the insect model Drosophila. Nat. Immunol. 2013, 14, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Roossinck, M.J. The good viruses: Viral mutualistic symbioses. Nat. Rev. Microbiol. 2011, 9, 99. [Google Scholar] [CrossRef] [PubMed]
- Mahy, B.W.J. Strategies of virus persistence. Br. Med. Bull. 1985, 41, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Grozinger, C.M.; Flenniken, M.L. Bee viruses: Ecology, pathogenicity, and impacts. Annu. Rev. Entomol. 2019, 64, 205–226. [Google Scholar] [CrossRef] [PubMed]
- Manchester, M.; Anand, A. Metabolomics: Strategies to define the role of metabolism in virus infection and pathogenesis. In Advances in Virus Research; Elsevier: Amsterdam, The Netherlands, 2017; Volume 98, pp. 57–81. [Google Scholar]
- Rouse, B.T.; Sehrawat, S. Immunity and immunopathology to viruses: What decides the outcome? Nat. Rev. Immunol. 2010, 10, 514–526. [Google Scholar] [CrossRef] [PubMed]
- Buck, M.D.; Sowell, R.T.; Kaech, S.M.; Pearce, E.L. Metabolic Instruction of Immunity. Cell 2017, 169, 570–586. [Google Scholar] [CrossRef] [PubMed]
- Noto, A.; Dessi, A.; Puddu, M.; Mussap, M.; Fanos, V. Metabolomics technology and their application to the study of the viral infection. J. Matern. Fetal Neonatal Med. 2014, 27, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, E.L.; Lagunoff, M. Viral activation of cellular metabolism. Virology 2015, 479, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wu, J.; Du, F.; Chen, X.; Chen, Z.J. Cyclic GMP-AMP synthase is a cytosolic DNA sensor that activates the type I interferon pathway. Science 2013, 339, 786–791. [Google Scholar] [CrossRef] [PubMed]
- Schoeman, J.C.; Hou, J.; Harms, A.C.; Vreeken, R.J.; Berger, R.; Hankemeier, T.; Boonstra, A. Metabolic characterization of the natural progression of chronic hepatitis B. Genome Med. 2016, 8, 64. [Google Scholar] [CrossRef]
- Cui, L.; Lee, Y.H.; Kumar, Y.; Xu, F.; Lu, K.; Ooi, E.E.; Tannenbaum, S.R.; Ong, C.N. Serum metabolome and lipidome changes in adult patients with primary dengue infection. PLoS Negl. Trop. Dis. 2013, 7, e2373. [Google Scholar] [CrossRef] [PubMed]
- Maynard, N.D.; Gutschow, M.V.; Birch, E.W.; Covert, M.W. The virus as metabolic engineer. Biotechnol. J. 2010, 5, 686–694. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.H.; Ivanisevic, J.; Siuzdak, G. Metabolomics: Beyond biomarkers and towards mechanisms. Nat. Rev. Mol. Cell Biol. 2016, 17, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Oldiges, M.; Lütz, S.; Pflug, S.; Schroer, K.; Stein, N.; Wiendahl, C. Metabolomics: Current state and evolving methodologies and tools. Appl. Microbiol. Biotechnol. 2007, 76, 495–511. [Google Scholar] [CrossRef] [PubMed]
- Munger, J.; Bajad, S.U.; Coller, H.A.; Shenk, T.; Rabinowitz, J.D. Dynamics of the cellular metabolome during human cytomegalovirus infection. PLoS Pathog. 2006, 2, e132. [Google Scholar] [CrossRef]
- Fan, W.; Ye, Y.; Chen, Z.; Shao, Y.; Xie, X.; Zhang, W.; Liu, H.P.; Li, C. Metabolic product response profiles of Cherax quadricarinatus towards white spot syndrome virus infection. Dev. Comp. Immunol. 2016, 61, 236–241. [Google Scholar] [CrossRef]
- Su, M.A.; Huang, Y.T.; Chen, I.T.; Lee, D.Y.; Hsieh, Y.C.; Li, C.Y.; Ng, T.H.; Liang, S.Y.; Lin, S.Y.; Huang, S.W.; et al. An invertebrate Warburg effect: A shrimp virus achieves successful replication by altering the host metabolome via the PI3K-Akt-mTOR pathway. PLoS Pathog. 2014, 10, e1004196. [Google Scholar] [CrossRef]
- Munger, J.; Bennett, B.D.; Parikh, A.; Feng, X.J.; McArdle, J.; Rabitz, H.A.; Shenk, T.; Rabinowitz, J.D. Systems level metabolic flux profiling identifies fatty acid synthesis as a target for antiviral therapy. Nat. Biotechnol. 2008, 26, 1179–1186. [Google Scholar] [CrossRef]
- Delgado, T.; Sanchez, E.L.; Camarda, R.; Lagunoff, M. Global metabolic profiling of infection by an oncogenic virus: KSHV induces and requires lipogenesis for survival of latent infection. PLoS Pathog. 2012, 8, e1002866. [Google Scholar] [CrossRef]
- Wang, L.L.; Cappelle, K.; Santos, D.; Van den Broeck, J.; Smagghe, G.; Swevers, L. Short-term persistence precedes pathogenic infection: Infection kinetics of Cricket Paralysis virus in silkworm-derived Bm5 cells. J. Insect Physiol. 2019, 115, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Santos, D.; Wynant, N.; Van den Brande, S.; Verdonckt, T.W.; Mingels, L.; Peeters, P.; Kolliopoulou, A.; Swevers, L.; Van den Broeck, J. Insights into RNAi-based antiviral immunity in Lepidoptera: Acute and persistent infections in Bombyx mori and Trichoplusia ni cell lines. Sci. Rep. 2018, 8, 2423. [Google Scholar] [CrossRef] [PubMed]
- Vanden Bussche, J.; Marzorati, M.; Laukens, D.; Vanhaecke, L. Validated high resolution mass spectrometry-based approach for metabolomic fingerprinting of the human gut phenotype. Anal. Chem. 2015, 87, 10927–10934. [Google Scholar] [CrossRef] [PubMed]
- De Paepe, E.; Van Meulebroek, L.; Rombouts, C.; Huysman, S.; Verplanken, K.; Lapauw, B.; Wauters, J.; Hemeryck, L.Y.; Vanhaecke, L. A validated multi-matrix platform for metabolomic fingerprinting of human urine, feces and plasma using ultra-high performance liquid-chromatography coupled to hybrid orbitrap high-resolution mass spectrometry. Anal. Chim. Acta 2018, 1033, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Ser, Z.; Locasale, J.W. Development and quantitative evaluation of a high-resolution metabolomics technology. Anal. Chem. 2014, 86, 2175–2184. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Ko, B.; Hensley, C.T.; Jiang, L.; Wasti, A.T.; Kim, J.; Sudderth, J.; Calvaruso, M.A.; Lumata, L.; Mitsche, M. Glutamine oxidation maintains the TCA cycle and cell survival during impaired mitochondrial pyruvate transport. Mol. Cell 2014, 56, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Mounce, B.C.; Olsen, M.E.; Vignuzzi, M.; Connor, J.H. Polyamines and their role in virus infection. Microbiol. Mol. Biol. Rev. 2017, 81, e00029-17. [Google Scholar] [CrossRef]
- Fontaine, K.A.; Sanchez, E.L.; Camarda, R.; Lagunoff, M. Dengue virus induces and requires glycolysis for optimal replication. J. Virol. 2015, 89, 2358–2366. [Google Scholar] [CrossRef]
- Maity, S.; Jannasch, A.; Adamec, J.; Nalepa, T.; Höök, T.O.; Sepúlveda, M.S. Starvation causes disturbance in amino acid and fatty acid metabolism in Diporeia. Comp. Biochem. Physiol. B 2012, 161, 348–355. [Google Scholar] [CrossRef]
- Sanchez, E.L.; Pulliam, T.H.; Dimaio, T.A.; Thalhofer, A.B.; Delgado, T.; Lagunoff, M. Glycolysis, glutaminolysis, and fatty acid synthesis are required for distinct stages of Kaposi’s sarcoma-associated herpesvirus lytic replication. J. Virol. 2017, 91, e02237-16. [Google Scholar] [CrossRef]
- Heaton, N.S.; Randall, G. Dengue virus-induced autophagy regulates lipid metabolism. Cell Host Microbe 2010, 8, 422–432. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.; Mandal, A.; Johansson, H.E.; Orjalo, A.V.; Park, M.H. Depletion of cellular polyamines, spermidine and spermine, causes a total arrest in translation and growth in mammalian cells. Proc. Natl. Acad. Sci. USA 2013, 110, 2169–2174. [Google Scholar] [CrossRef] [PubMed]
- Li, M.H.; MacDonald, M.R. Polyamines: Small molecules with a big role in promoting virus Infection. Cell Host Microbe 2016, 20, 123–124. [Google Scholar] [CrossRef] [PubMed]
- Raina, A.; Tuomi, K.; Mäntyjärvi, R. Roles of polyamines in the replication of animal viruses. Med. Biol. 1981, 59, 428–432. [Google Scholar] [PubMed]
- Tuomi, K.; Raina, A.; Mäntyjärvi, R. Synthesis of Semliki forest virus in polyamine depleted baby hamster kidney cells. Biochem. J. 1982, 206, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Berg, J.M.; Tymoczko, J.L.; Stryer, L. Biochemistry; W. H. Freeman and Company: New York, NY, USA, 2002. [Google Scholar]
- Tahiliani, A.G.; Beinlich, C.J. Pantothenic acid in health and disease. In Vitamins & Hormones; Elsevier: Amsterdam, The Netherlands, 1991; Volume 46, pp. 165–228. [Google Scholar]
- Hillyer, J.F. Insect immunology and hematopoiesis. Dev. Comp. Immunol. 2016, 58, 102–118. [Google Scholar] [CrossRef]









{kind=link}
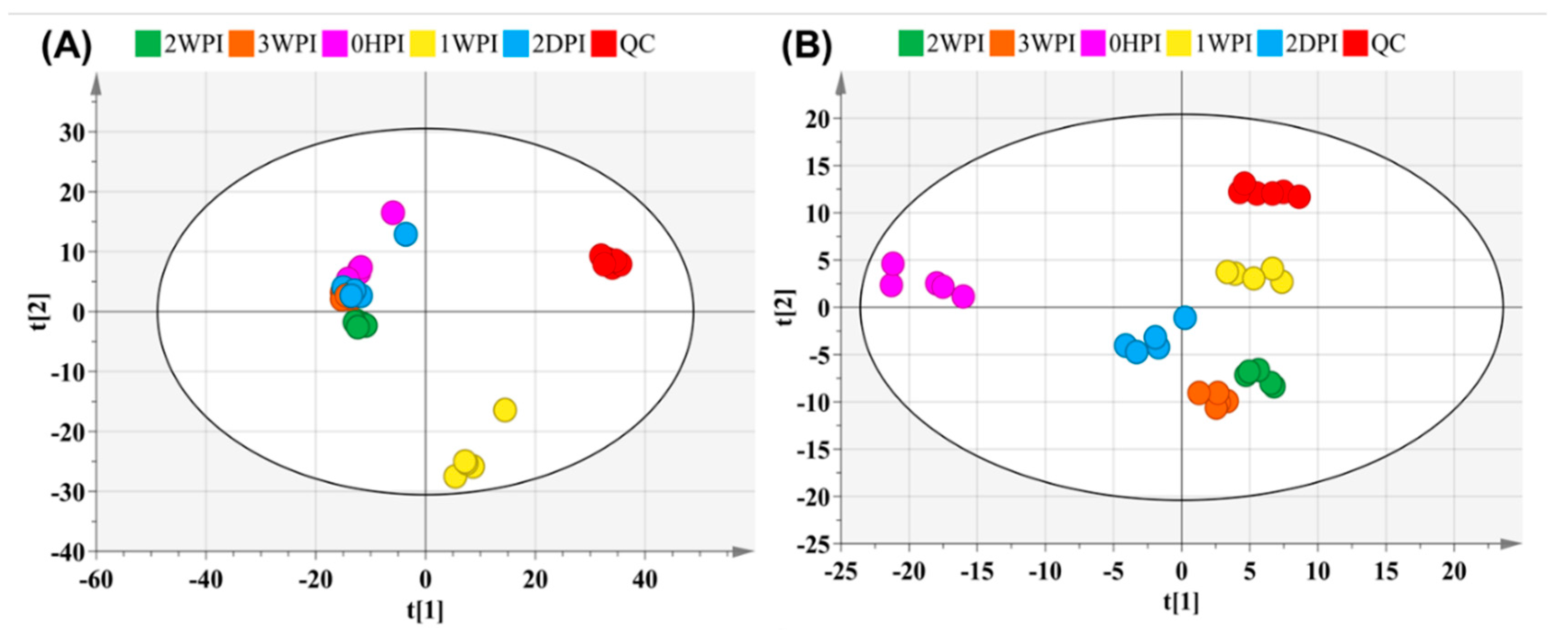{kind=link}
{kind=link}
{kind=link}
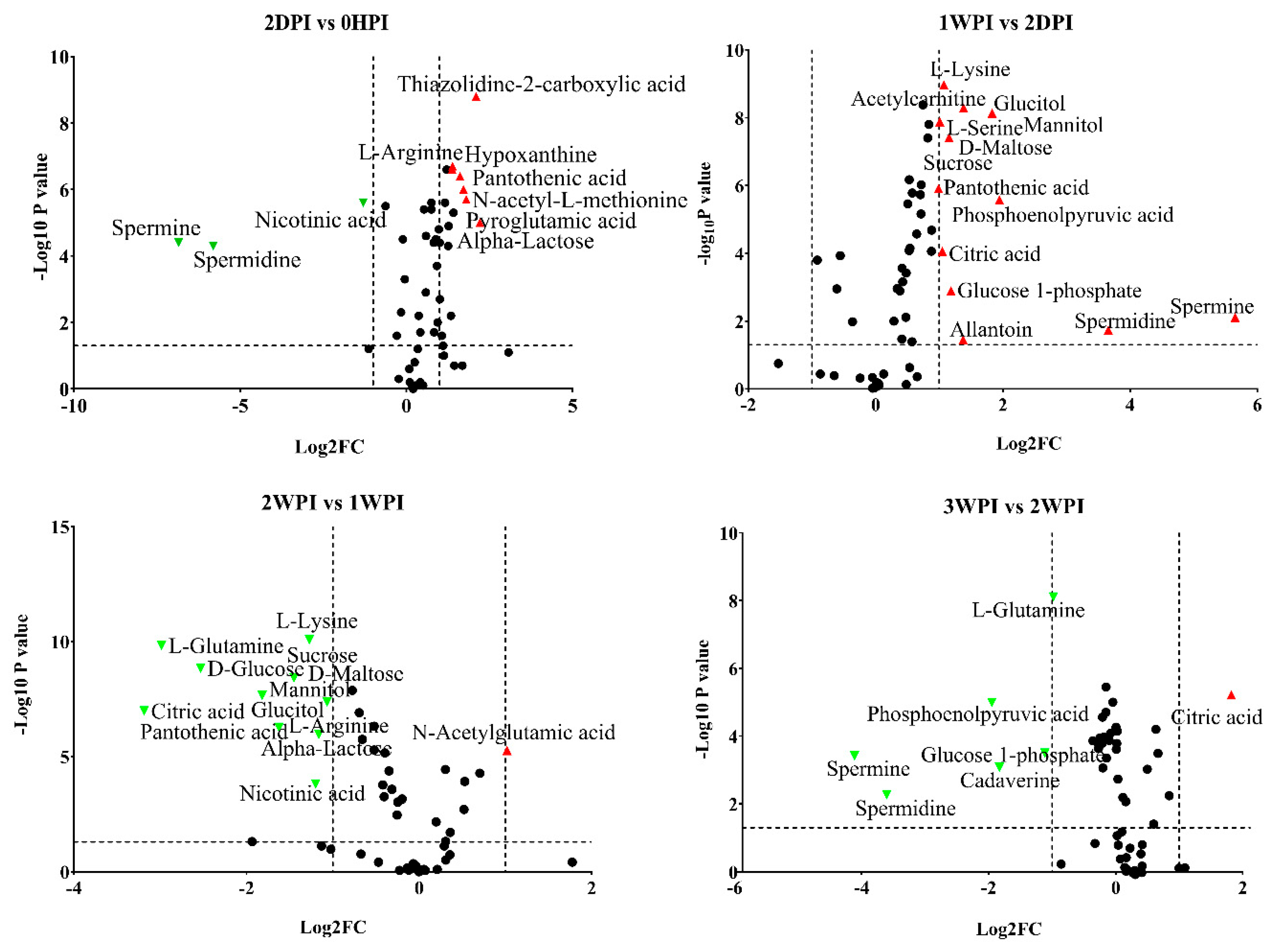{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Numbers of Instances | Numbers of Model Components (to + tp) a | Model Characteristics b | Cross-Validated ANOVA c | Permutation d |
|---|---|---|---|---|---|
| 0HPI vs. 2DPI | 10 | 1 + 1 + 0 | R2Y = 0.993; Q2 = 0.973 | 3.96 × 10−4 | Good |
| 0HPI vs. 1WPI | 10 | 1 + 0 + 0 | R2Y = 1.000; Q2 = 0.995 | 6.16 × 10−9 | Good |
| 0HPI vs. 2WPI | 10 | 1 + 0 + 0 | R2Y = 0.99; Q2 = 0.984 | 5.52 × 10−7 | Good |
| 0HPI vs. 3WPI | 10 | 1 + 0 + 0 | R2Y = 0.996; Q2 = 0.987 | 2.58 × 10−7 | Good |
| 2DPI vs. 1WPI | 10 | 1 + 0 + 0 | R2Y = 0.996; Q2 = 0.988 | 2.15 × 10−4 | Good |
| 1WPI vs. 2WPI | 10 | 1 + 0 + 0 | R2Y = 0.998; Q2 = 0.995 | 1.03 × 10−8 | Good |
| 2WPI vs. 3WPI | 10 | 1 + 1 + 0 | R2Y = 0.998; Q2 = 0.958 | 1.26 × 10−3 | Good |
| 2DPI vs. 2WPI | 10 | 1 + 2 + 0 | R2Y = 0.999; Q2 = 0.970 | 3.79 × 10−3 | Good |
| 2DPI vs. 3WPI | 10 | 1 + 2 + 0 | R2Y = 1.000; Q2 = 0.923 | 2.03 × 10−2 | Good |
| 1WPI vs. 3WPI | 10 | 1 + 0 + 0 | R2Y = 0.999; Q2 = 0.994 | 1.54 × 10−8 | Good |
| 2DPI | 1WPI | 2WPI | 3WPI | |
|---|---|---|---|---|
| Amino Acids | ||||
| L-Arginine | 1.08 | 1.75 | 0.84 | 1.05 |
| L-Asparagine | 0.87 | 1.46 | 1.02 | 0.82 |
| L-Aspartic acid | 1.00 | 1.01 | 1.26 | 0.98 |
| L-Cysteine | 1.02 | 1.87 | 1.41 | 0.88 |
| L-Glutamic acid | 0.96 | 1.70 | 1.05 | 0.85 |
| L-Glutamine | 1.43 | 2.14 | 0.27 | 0.11 |
| L-Histidine | 0.94 | 1.25 | 1.24 | 1.11 |
| L-Isoleucine | 1.08 | 1.55 | 1.25 | 0.87 |
| L-Leucine | 1.08 | 1.55 | 1.25 | 0.87 |
| L-Lysine | 0.96 | 2.02 | 0.84 | 0.95 |
| L-Methionine | 1.03 | 1.34 | 1.32 | 0.93 |
| L-Phenylalanine | 1.08 | 1.58 | 1.33 | 0.96 |
| L-Proline | 1.05 | 1.51 | 1.15 | 0.87 |
| L-Serine | 0.77 | 1.55 | 0.98 | 0.93 |
| L-Threonine | 0.93 | 1.67 | 0.98 | 0.70 |
| L-Tyrosine | 1.11 | 1.73 | 1.29 | 0.87 |
| L-Valine | 1.11 | 1.54 | 1.29 | 0.90 |
| N-acetyl-Methionine | 0.86 | 1.05 | 1.71 | 1.47 |
| N-acetyl-Glutamic acid | 1.25 | 0.82 | 1.66 | 1.11 |
| Pyro-Glutamic acid | 1.11 | 1.81 | 1.26 | 0.95 |
| 4-Hydroxyproline | 1.00 | 1.42 | 1.12 | 0.91 |
| 5-Aminovaleric acid | 1.11 | 1.54 | 1.29 | 0.90 |
| 2-Amino-isobutyric acid | 0.77 | 1.07 | 1.37 | 1.14 |
| Carbohydrates | ||||
| D-Arabinose | 1.08 | 1.37 | 1.19 | 0.98 |
| D-Glucose | 1.75 | 1.77 | 0.31 | 0.27 |
| D-Maltose | 0.94 | 2.09 | 0.77 | 0.98 |
| D-Ribose | 0.94 | 2.09 | 0.77 | 0.98 |
| Alpha-Lactose | 1.08 | 1.99 | 0.89 | 1.28 |
| Glucitol | 0.65 | 2.34 | 0.66 | 0.81 |
| Mannitol | 0.65 | 2.34 | 0.66 | 0.81 |
| Gluconic acid | 1.29 | 0.87 | 1.26 | 0.92 |
| Glucose-1-phosphate | 0.63 | 1.44 | 1.49 | 0.56 |
| Sucrose | 0.94 | 2.09 | 0.77 | 0.98 |
| Other metabolites | ||||
| Allantoin | 0.69 | 1.80 | 1.12 | 1.09 |
| Citric acid | 0.90 | 1.86 | 0.20 | 0.58 |
| Dipropylsulfide | 1.03 | 1.38 | 1.33 | 0.93 |
| Gamma-amino-butyric acid | 0.86 | 1.27 | 1.57 | 1.01 |
| Glycerol-1-phosphate | 0.75 | 1.24 | 1.25 | 1.02 |
| L-Gulunolactone | 1.35 | 0.72 | 0.87 | 0.71 |
| Imidazolepropionic acid | 1.46 | 1.60 | 1.60 | 1.35 |
| Nicotinic acid | 0.96 | 1.27 | 0.55 | 0.49 |
| Pantothenic acid | 1.39 | 2.75 | 0.89 | 0.96 |
| Phosphoenolpyruvic acid | 0.56 | 2.16 | 2.67 | 0.55 |
| Thiazolidine-2-carboxylic acid | 1.37 | 1.07 | 1.53 | 1.02 |
| Hypoxanthine | 1.16 | 1.12 | 1.28 | 1.26 |
| Pipecolic acid | 0.61 | 0.96 | 0.89 | 0.80 |
| Spermidine | 0.22 | 2.77 | 0.73 | 0.05 |
| Spermine | 0.07 | 3.52 | 2.54 | 0.12 |
| Acetylcarnitine | 0.54 | 1.42 | 1.38 | 0.97 |
| Comparison | Pathway | hits | Holm Adjusted P | FDR | Impact |
|---|---|---|---|---|---|
| 2DPI vs. 0HPI | Aminoacyl-tRNA biosynthesis | 15 | 4.38 × 10−9 | 4.38 × 10−9 | 0.14 |
| Arginine and proline metabolism | 7 | 5.25 × 10−3 | 2.66 × 10−3 | 0.42 | |
| Valine, leucine and isoleucine biosynthesis | 4 | 3.27 × 10−2 | 1.12 × 10−2 | 1.00 | |
| 1WPI vs. 0HPI | Aminoacyl-tRNA biosynthesis | 16 | 7.76 × 10−8 | 7.76 × 10−8 | 0.00 |
| Arginine and proline metabolism | 8 | 5.45 × 10−3 | 2.76 × 10−3 | 0.42 | |
| Valine, leucine and isoleucine biosynthesis | 4 | 1.07 × 10−1 | 3.67 × 10−2 | 1.00 | |
| Alanine, aspartate and glutamate metabolism | 5 | 1.41 × 10−1 | 3.67 × 10−2 | 0.73 | |
| 2WPI vs. 0HPI | Aminoacyl-tRNA biosynthesis | 15 | 2.44 × 10−7 | 2.44 × 10−7 | 0.14 |
| Arginine and proline metabolism | 9 | 3.19 × 10−4 | 1.61 × 10−4 | 0.42 | |
| Valine, leucine and isoleucine biosynthesis | 4 | 8.06 × 10−2 | 2.76 × 10−2 | 1.00 | |
| 3WPI vs. 0HPI | Aminoacyl-tRNA biosynthesis | 16 | 5.05 × 10−8 | 5.05 × 10−8 | 0.14 |
| Arginine and proline metabolism | 8 | 4.53 × 10−3 | 2.29 × 10−3 | 0.42 | |
| Valine, leucine and isoleucine biosynthesis | 4 | 9.78 × 10−2 | 3.35 × 10−2 | 1.00 | |
| 1WPI vs. 2DPI | Aminoacyl-tRNA biosynthesis | 16 | 2.69 × 10−9 | 2.69 × 10−9 | 0.14 |
| Valine, leucine and isoleucine biosynthesis | 4 | 5.35 × 10−2 | 2.71 × 10−2 | 1.00 | |
| 2WPI vs. 1WPI | Aminoacyl-tRNA biosynthesis | 14 | 2.22 × 10−8 | 2.22 × 10−8 | 0.14 |
| Arginine and proline metabolism | 6 | 3.14 × 10−2 | 1.59 × 10−2 | 0.37 | |
| 3WPI vs. 2WPI | Aminoacyl-tRNA biosynthesis | 15 | 4.38 × 10−9 | 4.38 × 10−9 | 0.0 |
| Arginine and proline metabolism | 7 | 5.25 × 10−3 | 2.66 × 10−3 | 0.37 | |
| Valine, leucine and isoleucine biosynthesis | 4 | 3.27 × 10−2 | 1.12 × 10−2 | 1.00 | |
| Glutathione metabolism | 5 | 6.10 × 10−2 | 1.50 × 10−2 | 0.12 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.-L.; Swevers, L.; Rombouts, C.; Meeus, I.; Van Meulebroek, L.; Vanhaecke, L.; Smagghe, G. A Metabolomics Approach to Unravel Cricket Paralysis Virus Infection in Silkworm Bm5 Cells. Viruses 2019, 11, 861. https://doi.org/10.3390/v11090861
Wang L-L, Swevers L, Rombouts C, Meeus I, Van Meulebroek L, Vanhaecke L, Smagghe G. A Metabolomics Approach to Unravel Cricket Paralysis Virus Infection in Silkworm Bm5 Cells. Viruses. 2019; 11(9):861. https://doi.org/10.3390/v11090861
Chicago/Turabian StyleWang, Luo-Luo, Luc Swevers, Caroline Rombouts, Ivan Meeus, Lieven Van Meulebroek, Lynn Vanhaecke, and Guy Smagghe. 2019. "A Metabolomics Approach to Unravel Cricket Paralysis Virus Infection in Silkworm Bm5 Cells" Viruses 11, no. 9: 861. https://doi.org/10.3390/v11090861
APA StyleWang, L.-L., Swevers, L., Rombouts, C., Meeus, I., Van Meulebroek, L., Vanhaecke, L., & Smagghe, G. (2019). A Metabolomics Approach to Unravel Cricket Paralysis Virus Infection in Silkworm Bm5 Cells. Viruses, 11(9), 861. https://doi.org/10.3390/v11090861