IFN-λ Decreases Murid Herpesvirus-4 Infection of the Olfactory Epithelium but Fails to Prevent Virus Reactivation in the Vaginal Mucosa


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Experiments
2.2. Viruses
2.3. Cell Culture
2.4. CRISPR Cas 9 Editing
2.5. ISGs and Viral Genome Quantification
2.6. Flow Cytometry
2.7. Vectors
2.8. Interferons
2.9. Immunohistochemistry
2.10. Statistics
3. Results
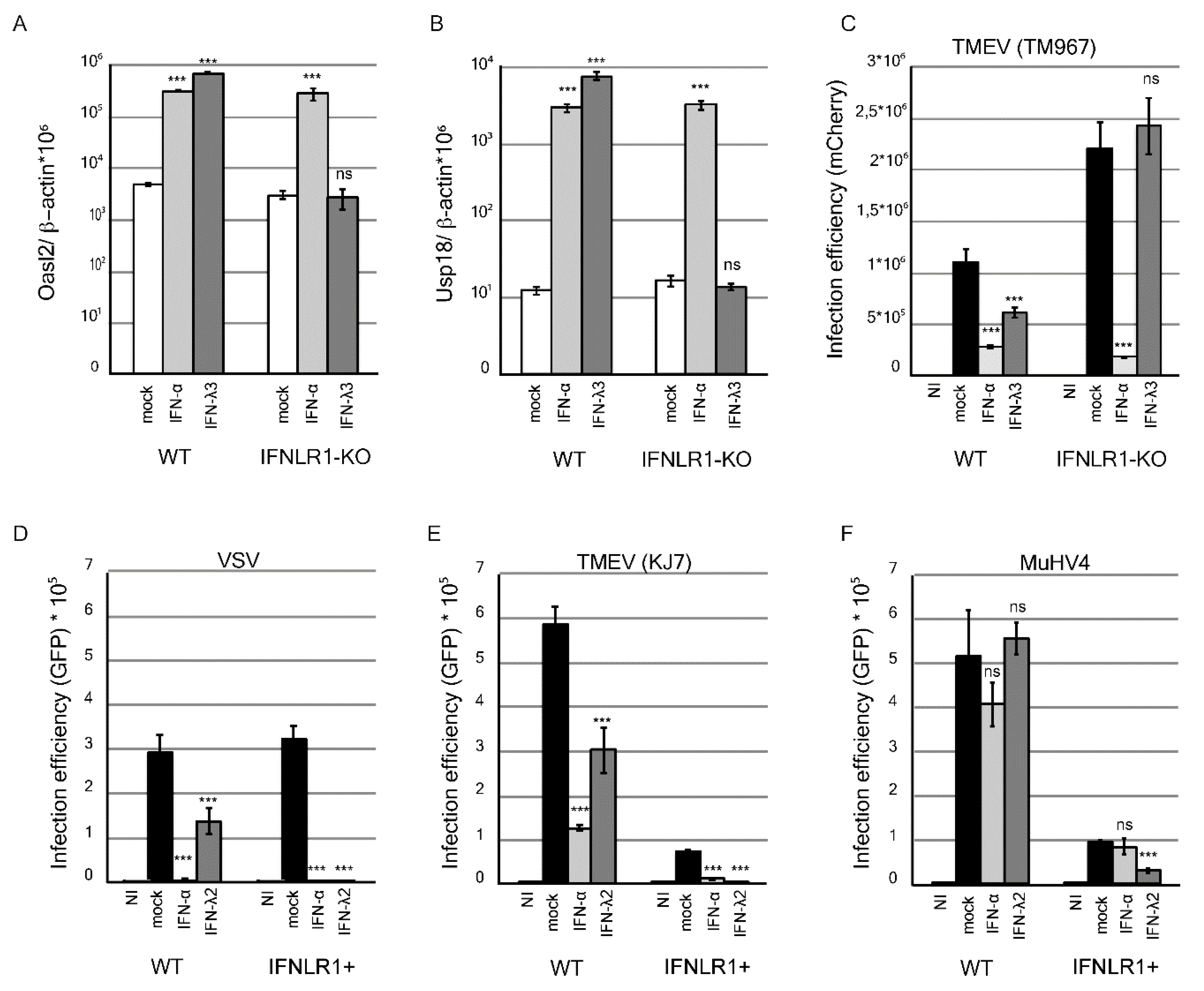
3.1. IFN-λ Decreases MuHV-4 Infection in Epithelial Cells Overexpressing IFNLR1
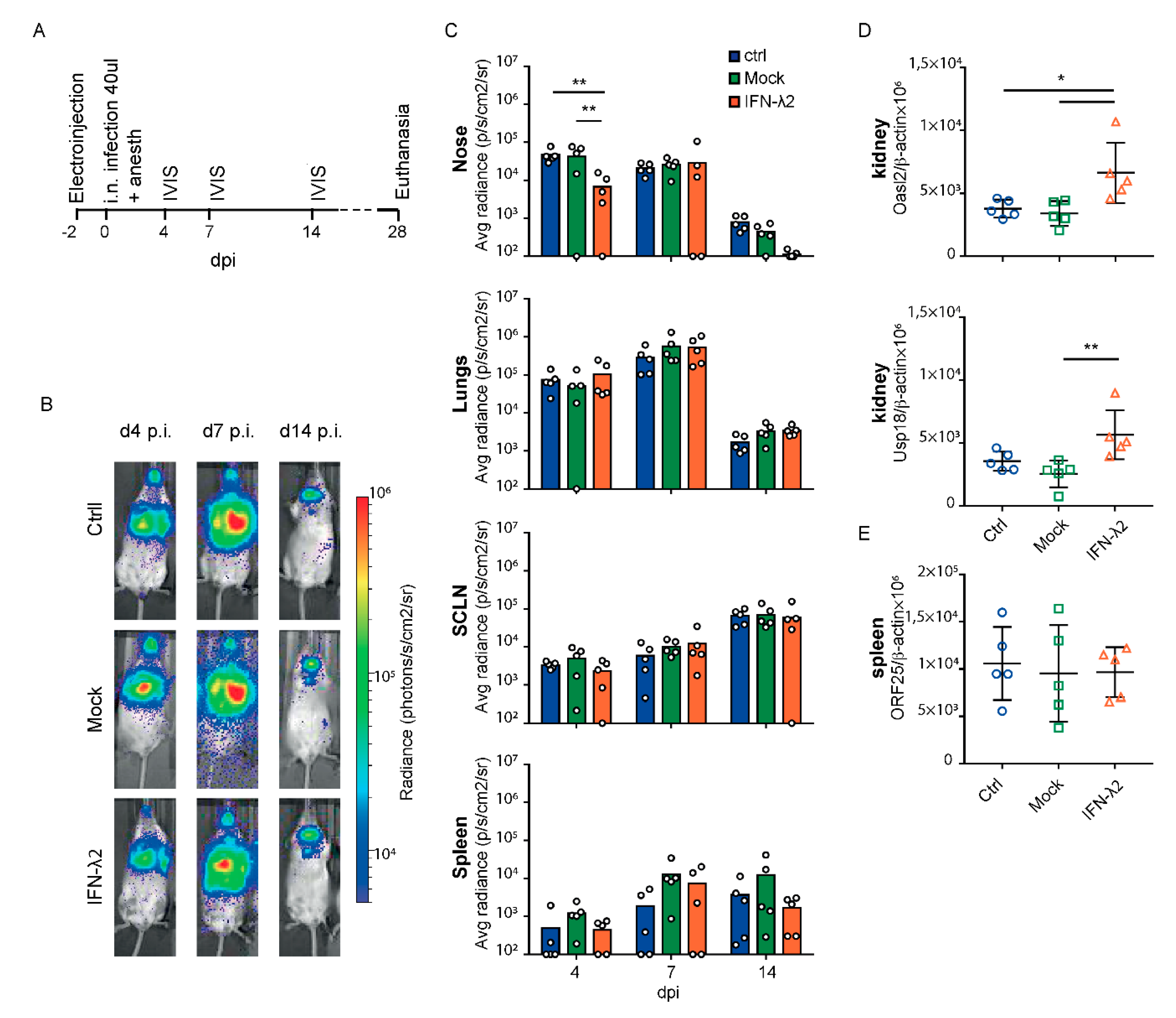
3.2. IFN-λ Decreases Nasal but not Lung MuHV-4 Infection
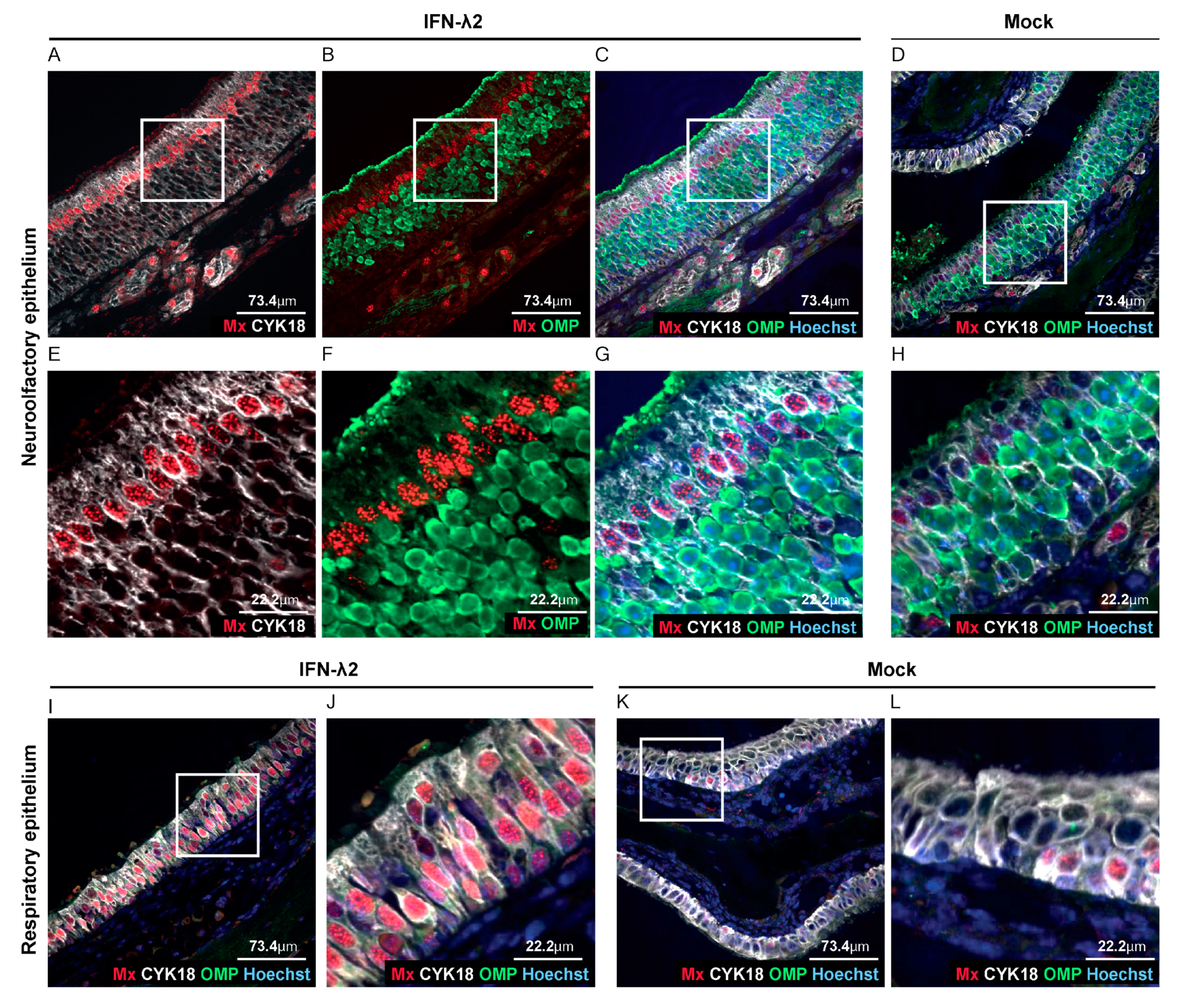
3.3. Sustentacular Cells of the Olfactory Epithelium Respond to IFN-λ Whereas Olfactory Neurons Do Not
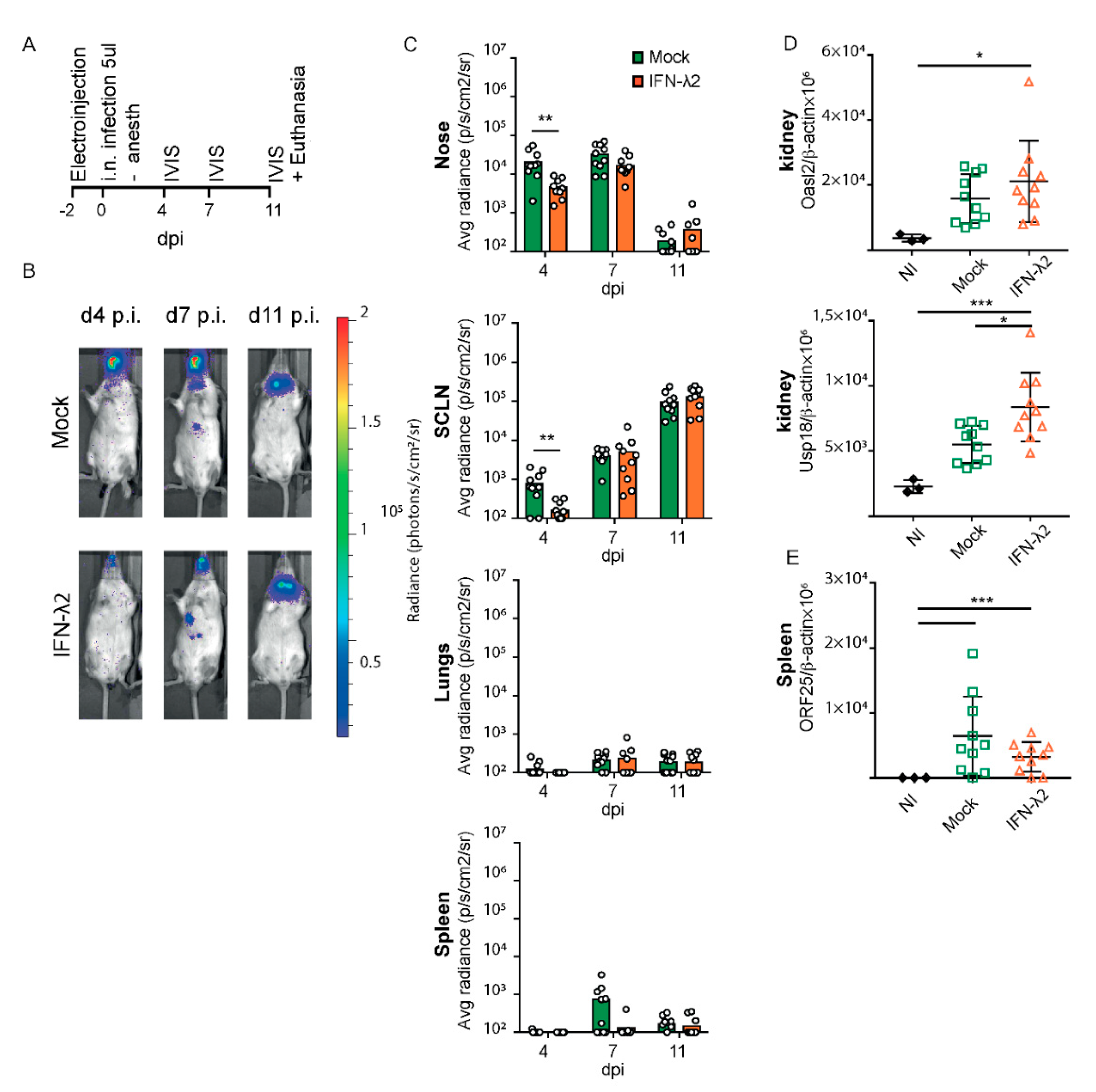
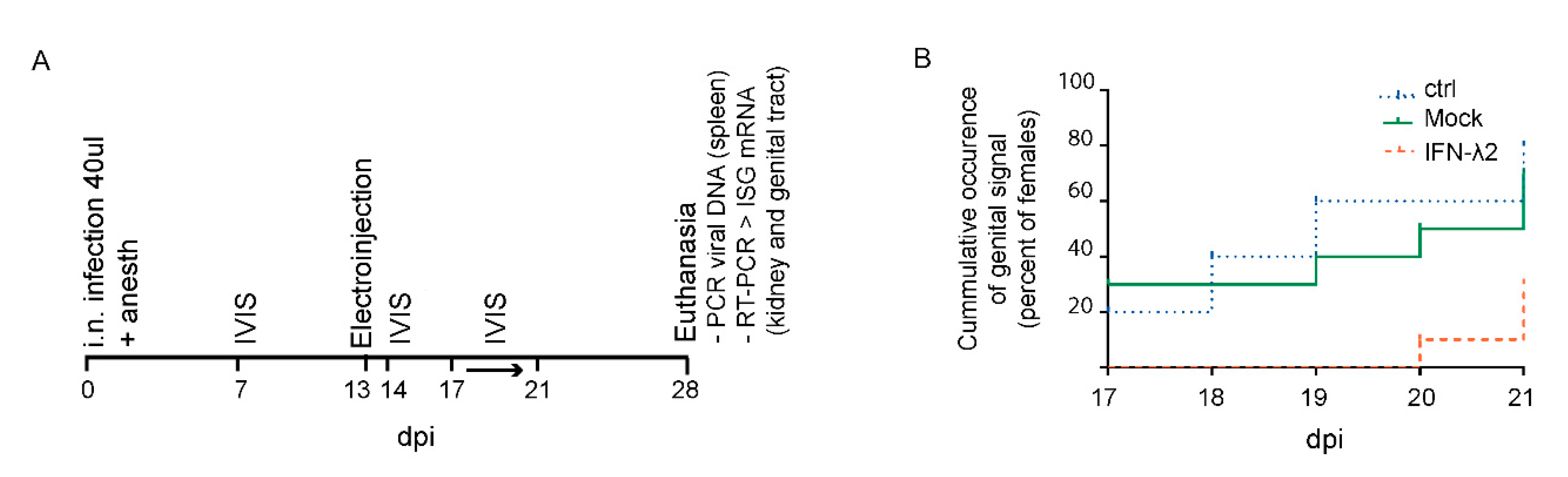
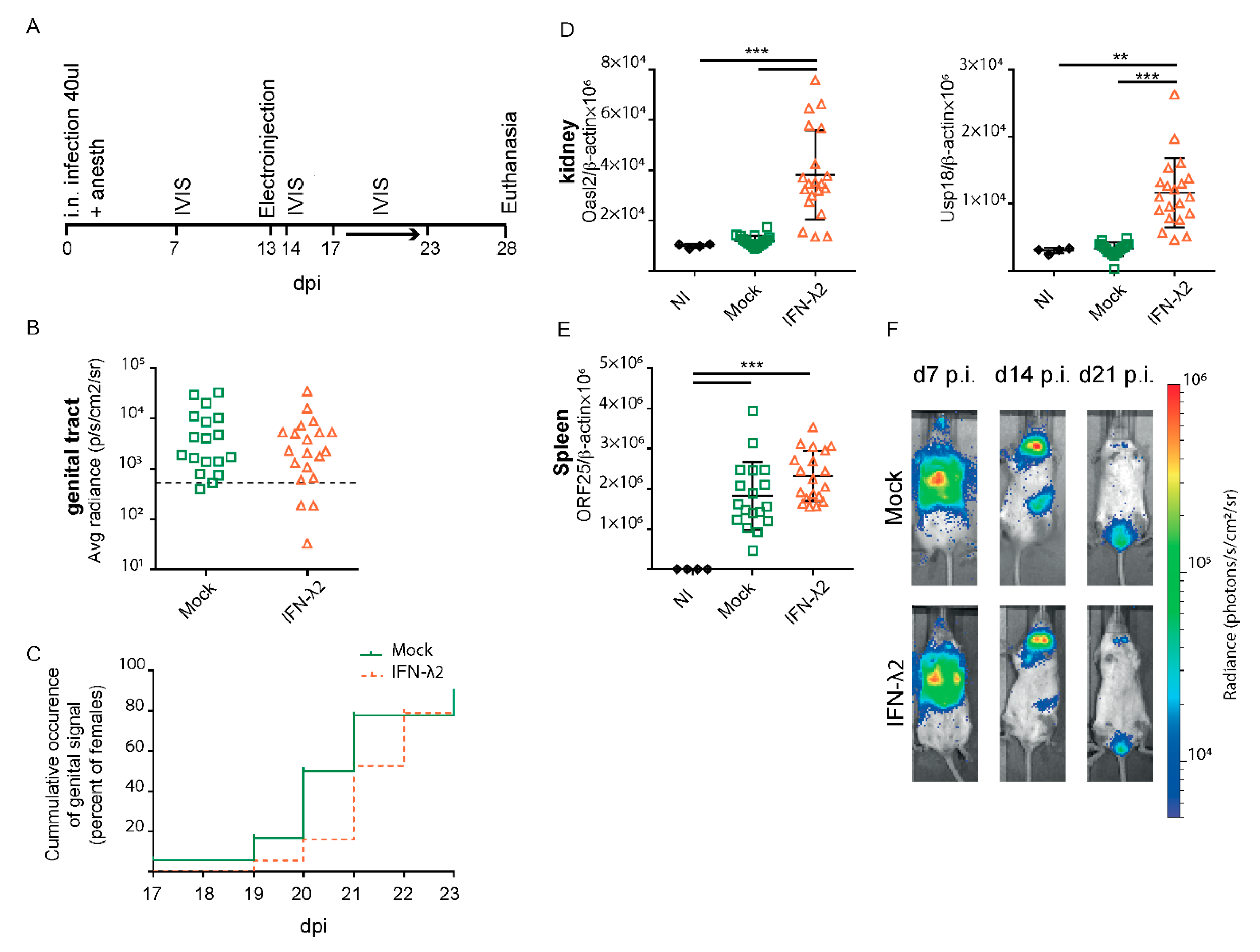
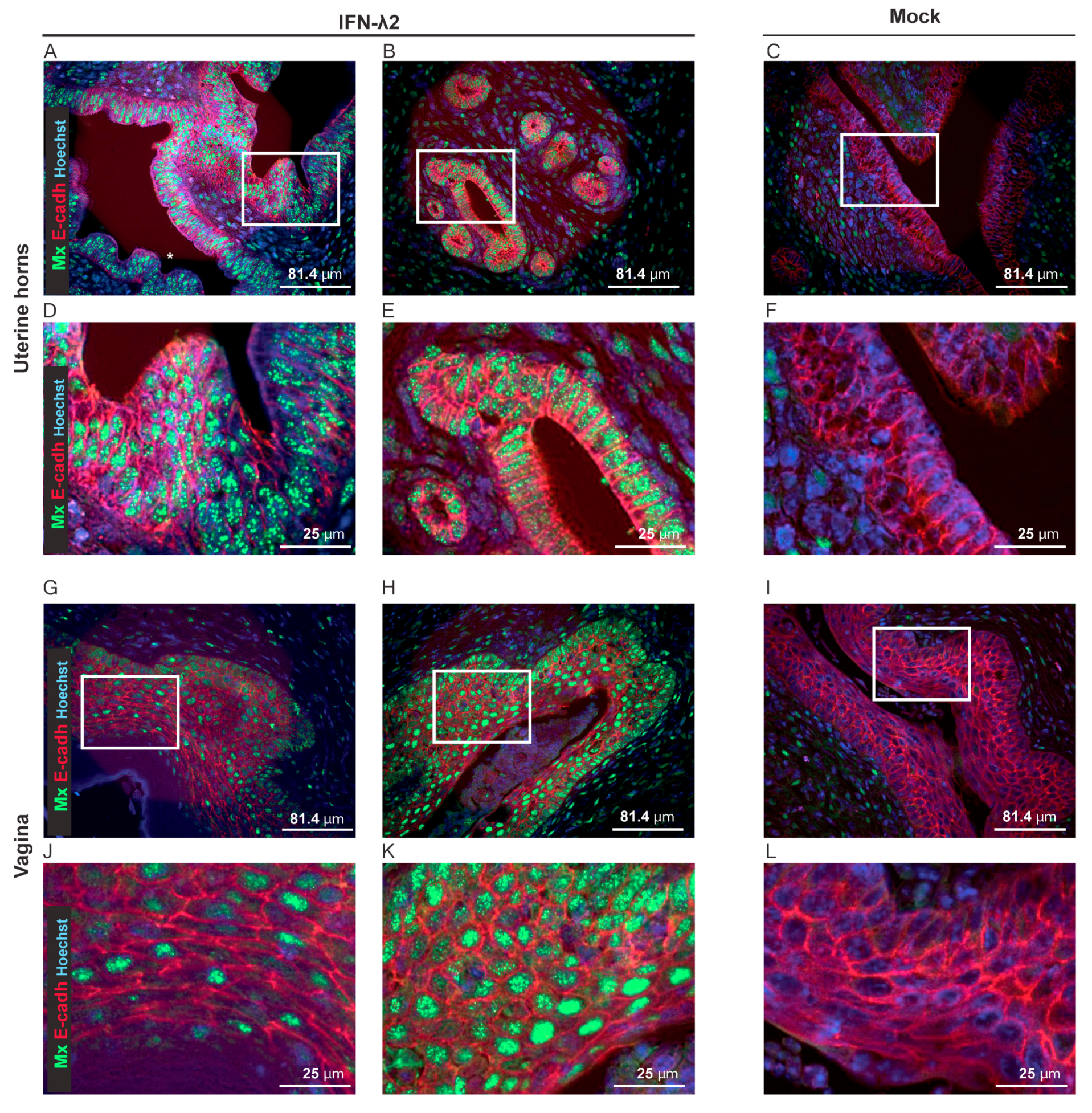
3.4. IFN-λ Expresion Fails to Prevent Genital Reactivation from Latency in Female Mice
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Henle, G.; Henle, W.; Clifford, P.; Diehl, V.; Kafuko, G.W.; Kirya, B.G.; Klein, G.; Morrow, R.H.; Munube, G.M.; Pike, P.; et al. Antibodies to epstein-barr virus in burkitt’s lymphoma and control groups. J. Natl. Cancer Inst. 1969, 43, 1147–1157. [Google Scholar] [PubMed]
- Verma, S.C.; Robertson, E.S. Molecular biology and pathogenesis of kaposi sarcoma-associated herpesvirus. FEMS Microbiol. Lett. 2003, 222, 155–163. [Google Scholar] [CrossRef]
- Young, L.S.; Rickinson, A.B. Epstein-barr virus: 40 years on. Nat. Rev. Cancer 2004, 4, 757–768. [Google Scholar] [CrossRef] [PubMed]
- Mesri, E.A.; Cesarman, E.; Boshoff, C. Kaposi’s sarcoma and its associated herpesvirus. Nat. Rev. Cancer 2010, 10, 707–719. [Google Scholar] [CrossRef] [PubMed]
- Lang, D.J.; Garruto, R.M.; Gajdusek, D.C. Early acquisition of cytomegalovirus and epstein-barr virus antibody in several isolated melanesian populations. Am. J. Epidemiol. 1977, 105, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Dunmire, S.K.; Grimm, J.M.; Schmeling, D.O.; Balfour, H.H., Jr.; Hogquist, K.A. The incubation period of primary epstein-barr virus infection: Viral dynamics and immunologic events. PLoS Pathog. 2015, 11, e1005286. [Google Scholar] [CrossRef] [PubMed]
- Blackbourn, D.J.; Lennette, E.T.; Ambroziak, J.; Mourich, D.V.; Levy, J.A. Human herpesvirus 8 detection in nasal secretions and saliva. J. Infect. Dis. 1998, 177, 213–216. [Google Scholar] [CrossRef] [PubMed]
- Davison, A.J. Evolution of sexually transmitted and sexually transmissible human herpesviruses. Ann. N. Y. Acad. Sci. 2011, 1230, E37–E49. [Google Scholar] [CrossRef] [PubMed]
- Kedes, D.H.; Operskalski, E.; Busch, M.; Kohn, R.; Flood, J.; Ganem, D. The seroepidemiology of human herpesvirus 8 (kaposi’s sarcoma-associated herpesvirus): Distribution of infection in ks risk groups and evidence for sexual transmission. Nat. Med. 1996, 2, 918–924. [Google Scholar] [CrossRef] [PubMed]
- Barton, E.; Mandal, P.; Speck, S.H. Pathogenesis and host control of gammaherpesviruses: Lessons from the mouse. Annu. Rev. Immunol. 2011, 29, 351–397. [Google Scholar] [CrossRef]
- Sunil-Chandra, N.P.; Efstathiou, S.; Nash, A.A. Murine gammaherpesvirus 68 establishes a latent infection in mouse b lymphocytes in vivo. J. Gen. Virol. 1992, 73, 3275–3279. [Google Scholar] [CrossRef] [PubMed]
- Frederico, B.; Milho, R.; May, J.S.; Gillet, L.; Stevenson, P.G. Myeloid infection links epithelial and b cell tropisms of murid herpesvirus-4. PLoS Pathog. 2012, 8, e1002935. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Collins, C.M.; Mendel, J.B.; Iwakoshi, N.N.; Speck, S.H. Gammaherpesvirus-driven plasma cell differentiation regulates virus reactivation from latently infected b lymphocytes. PLoS Pathog. 2009, 5, e1000677. [Google Scholar] [CrossRef] [PubMed]
- Gillet, L.; Frederico, B.; Stevenson, P.G. Host entry by gamma-herpesviruses—lessons from animal viruses? Curr. Opin. Virol. 2015, 15, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Milho, R.; Smith, C.M.; Marques, S.; Alenquer, M.; May, J.S.; Gillet, L.; Gaspar, M.; Efstathiou, S.; Simas, J.P.; Stevenson, P.G. In vivo imaging of murid herpesvirus-4 infection. J. Gen. Virol. 2009, 90, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Francois, S.; Vidick, S.; Sarlet, M.; Desmecht, D.; Drion, P.; Stevenson, P.G.; Vanderplasschen, A.; Gillet, L. Illumination of murine gammaherpesvirus-68 cycle reveals a sexual transmission route from females to males in laboratory mice. PLoS Pathog. 2013, 9, e1003292. [Google Scholar] [CrossRef]
- Ye, L.; Schnepf, D.; Staeheli, P. Interferon-lambda orchestrates innate and adaptive mucosal immune responses. Nat. Rev. Immunol. 2019. [Google Scholar] [CrossRef]
- Ank, N.; West, H.; Bartholdy, C.; Eriksson, K.; Thomsen, A.R.; Paludan, S.R. Lambda interferon (ifn-lambda), a type iii ifn, is induced by viruses and ifns and displays potent antiviral activity against select virus infections in vivo. J. Virol. 2006, 80, 4501–4509. [Google Scholar] [CrossRef]
- Baldridge, M.T.; Lee, S.; Brown, J.J.; McAllister, N.; Urbanek, K.; Dermody, T.S.; Nice, T.J.; Virgin, H.W. Expression of ifnlr1 on intestinal epithelial cells is critical to the antiviral effects of interferon lambda against norovirus and reovirus. J. Virol. 2017, 91, e02079-16. [Google Scholar] [CrossRef]
- Rocha-Pereira, J.; Jacobs, S.; Noppen, S.; Verbeken, E.; Michiels, T.; Neyts, J. Interferon lambda (ifn-lambda) efficiently blocks norovirus transmission in a mouse model. Antivir. Res. 2018, 149, 7–15. [Google Scholar] [CrossRef]
- Kotenko, S.V.; Gallagher, G.; Baurin, V.V.; Lewis-Antes, A.; Shen, M.; Shah, N.K.; Langer, J.A.; Sheikh, F.; Dickensheets, H.; Donnelly, R.P. Ifn-lambdas mediate antiviral protection through a distinct class ii cytokine receptor complex. Nat. Immunol. 2003, 4, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, P.; Kindsvogel, W.; Xu, W.; Henderson, K.; Schlutsmeyer, S.; Whitmore, T.E.; Kuestner, R.; Garrigues, U.; Birks, C.; Roraback, J.; et al. Il-28, il-29 and their class ii cytokine receptor il-28r. Nat. Immunol. 2003, 4, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Sommereyns, C.; Paul, S.; Staeheli, P.; Michiels, T. Ifn-lambda (ifn-lambda) is expressed in a tissue-dependent fashion and primarily acts on epithelial cells in vivo. PLoS Pathog. 2008, 4, e1000017. [Google Scholar] [CrossRef] [PubMed]
- Blazek, K.; Eames, H.L.; Weiss, M.; Byrne, A.J.; Perocheau, D.; Pease, J.E.; Doyle, S.; McCann, F.; Williams, R.O.; Udalova, I.A. Ifn-lambda resolves inflammation via suppression of neutrophil infiltration and il-1beta production. J. Exp. Med. 2015, 212, 845–853. [Google Scholar] [CrossRef] [PubMed]
- Broggi, A.; Tan, Y.; Granucci, F.; Zanoni, I. Ifn-lambda suppresses intestinal inflammation by non-translational regulation of neutrophil function. Nat. Immunol. 2017, 18, 1084–1093. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, V.; Dutta, O.; McElrath, C.; Du, P.; Chang, Y.J.; Cicciarelli, B.; Pitler, A.; Whitehead, I.; Obar, J.J.; Durbin, J.E.; et al. Type iii interferon is a critical regulator of innate antifungal immunity. Sci. Immunol. 2017, 2, eaan5357. [Google Scholar] [CrossRef] [PubMed]
- Koltsida, O.; Hausding, M.; Stavropoulos, A.; Koch, S.; Tzelepis, G.; Ubel, C.; Kotenko, S.V.; Sideras, P.; Lehr, H.A.; Tepe, M.; et al. Il-28a (ifn-lambda2) modulates lung dc function to promote th1 immune skewing and suppress allergic airway disease. EMBO Mol. Med. 2011, 3, 348–361. [Google Scholar] [CrossRef] [PubMed]
- Dumoutier, L.; Tounsi, A.; Michiels, T.; Sommereyns, C.; Kotenko, S.V.; Renauld, J.C. Role of the interleukin (il)-28 receptor tyrosine residues for antiviral and antiproliferative activity of il-29/interferon-lambda 1: Similarities with type i interferon signaling. J. Biol. Chem. 2004, 279, 32269–32274. [Google Scholar] [CrossRef] [PubMed]
- Randall, R.E.; Goodbourn, S. Interferons and viruses: An interplay between induction, signalling, antiviral responses and virus countermeasures. J. Gen. Virol. 2008, 89, 1–47. [Google Scholar] [CrossRef]
- Hwang, S.; Kim, K.S.; Flano, E.; Wu, T.T.; Tong, L.M.; Park, A.N.; Song, M.J.; Sanchez, D.J.; O’Connell, R.M.; Cheng, G.; et al. Conserved herpesviral kinase promotes viral persistence by inhibiting the irf-3-mediated type i interferon response. Cell Host Microbe 2009, 5, 166–178. [Google Scholar] [CrossRef]
- Kang, H.R.; Cheong, W.C.; Park, J.E.; Ryu, S.; Cho, H.J.; Youn, H.; Ahn, J.H.; Song, M.J. Murine gammaherpesvirus 68 encoding open reading frame 11 targets tank binding kinase 1 to negatively regulate the host type i interferon response. J. Virol. 2014, 88, 6832–6846. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sheridan, V.; Polychronopoulos, L.; Dutia, B.M.; Ebrahimi, B. A shutoff and exonuclease mutant of murine gammaherpesvirus-68 yields infectious virus and causes rna loss in type i interferon receptor knockout cells. J. Gen. Virol. 2014, 95, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Shin, Y.C.; Means, R.E.; Jung, J.U. Inhibition of interferon-mediated antiviral activity by murine gammaherpesvirus 68 latency-associated m2 protein. J. Virol. 2004, 78, 12416–12427. [Google Scholar] [CrossRef] [PubMed]
- Leang, R.S.; Wu, T.T.; Hwang, S.; Liang, L.T.; Tong, L.; Truong, J.T.; Sun, R. The anti-interferon activity of conserved viral dutpase orf54 is essential for an effective mhv-68 infection. PLoS Pathog. 2011, 7, e1002292. [Google Scholar] [CrossRef] [PubMed]
- Lopusna, K.; Benkoczka, T.; Luptak, J.; Matuskova, R.; Lukacikova, L.; Oveckova, I.; Rezuchova, I. Murine gammaherpesvirus targets type i ifn receptor but not type iii ifn receptor early in infection. Cytokine 2016, 83, 158–170. [Google Scholar] [CrossRef]
- Tan, C.S.; Lawler, C.; May, J.S.; Belz, G.T.; Stevenson, P.G. Type i interferons direct gammaherpesvirus host colonization. PLoS Pathog. 2016, 12, e1005654. [Google Scholar] [CrossRef] [PubMed]
- Lawler, C.; Milho, R.; May, J.S.; Stevenson, P.G. Rhadinovirus host entry by co-operative infection. PLoS Pathog. 2015, 11, e1004761. [Google Scholar] [CrossRef]
- Milho, R.; Frederico, B.; Efstathiou, S.; Stevenson, P.G. A heparan-dependent herpesvirus targets the olfactory neuroepithelium for host entry. PLoS Pathog. 2012, 8, e1002986. [Google Scholar] [CrossRef]
- Lawler, C.; Stevenson, P.G. Type i interferon signaling to dendritic cells limits murid herpesvirus 4 spread from the olfactory epithelium. J. Virol. 2017, 91, e00951-17. [Google Scholar] [CrossRef]
- Klinkhammer, J.; Schnepf, D.; Ye, L.; Schwaderlapp, M.; Gad, H.H.; Hartmann, R.; Garcin, D.; Mahlakoiv, T.; Staeheli, P. Ifn-lambda prevents influenza virus spread from the upper airways to the lungs and limits virus transmission. Elife 2018, 7, e33354. [Google Scholar] [CrossRef]
- Muir, A.J.; Arora, S.; Everson, G.; Flisiak, R.; George, J.; Ghalib, R.; Gordon, S.C.; Gray, T.; Greenbloom, S.; Hassanein, T.; et al. A randomized phase 2b study of peginterferon lambda-1a for the treatment of chronic hcv infection. J. Hepatol. 2014, 61, 1238–1246. [Google Scholar] [CrossRef] [PubMed]
- Davidson, S.; McCabe, T.M.; Crotta, S.; Gad, H.H.; Hessel, E.M.; Beinke, S.; Hartmann, R.; Wack, A. Ifnlambda is a potent anti-influenza therapeutic without the inflammatory side effects of ifnalpha treatment. EMBO Mol. Med. 2016, 8, 1099–1112. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, S.; Wavreil, F.; Schepens, B.; Gad, H.H.; Hartmann, R.; Rocha-Pereira, J.; Neyts, J.; Saelens, X.; Michiels, T. Species specificity of type iii interferon activity and development of a sensitive luciferase-based bioassay for quantitation of mouse ifn-λ. J. Interferon Cytokine Res. 2018, 38, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Jnaoui, K.; Michiels, T. Adaptation of theiler’s virus to l929 cells: Mutations in the putative receptor binding site on the capsid map to neutralization sites and modulate viral persistence. Virology 1998, 244, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Adler, H.; Messerle, M.; Wagner, M.; Koszinowski, U.H. Cloning and mutagenesis of the murine gammaherpesvirus 68 genome as an infectious bacterial artificial chromosome. J. Virol. 2000, 74, 6964–6974. [Google Scholar] [CrossRef] [PubMed]
- Zeippen, C.; Javaux, J.; Xiao, X.; Ledecq, M.; Mast, J.; Farnir, F.; Vanderplasschen, A.; Stevenson, P.; Gillet, L. The major envelope glycoprotein of murid herpesvirus 4 promotes sexual transmission. J. Virol. 2017, 91, e00235-17. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.; Mercer, K.; Greenbaum, D.; Bronson, R.T.; Crowley, D.; Tuveson, D.A.; Jacks, T. Somatic activation of the k-ras oncogene causes early onset lung cancer in mice. Nature 2001, 410, 1111–1116. [Google Scholar] [CrossRef]
- Aaronson, S.A.; Todaro, G.J. Development of 3t3-like lines from balb-c mouse embryo cultures: Transformation susceptibility to sv40. J. Cell Physiol. 1968, 72, 141–148. [Google Scholar] [CrossRef]
- DuBridge, R.B.; Tang, P.; Hsia, H.C.; Leong, P.M.; Miller, J.H.; Calos, M.P. Analysis of mutation in human cells by using an epstein-barr virus shuttle system. Mol. Cell Biol. 1987, 7, 379–387. [Google Scholar] [CrossRef]
- Ran, F.A.; Hsu, P.D.; Lin, C.Y.; Gootenberg, J.S.; Konermann, S.; Trevino, A.E.; Scott, D.A.; Inoue, A.; Matoba, S.; Zhang, Y.; et al. Double nicking by rna-guided crispr cas9 for enhanced genome editing specificity. Cell 2013, 154, 1380–1389. [Google Scholar] [CrossRef]
- Optimized CRISPR Design. Available online: http://crispr.mit.edu (accessed on 12 October 2014).
- Chomczynski, P.; Sacchi, N. Single-step method of rna isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Paul, S.; Michiels, T. Cardiovirus leader proteins are functionally interchangeable and have evolved to adapt to virus replication fitness. J. Gen. Virol. 2006, 87, 1237–1246. [Google Scholar] [CrossRef] [PubMed]
- Hermant, P.; Francius, C.; Clotman, F.; Michiels, T. Ifn-epsilon is constitutively expressed by cells of the reproductive tract and is inefficiently secreted by fibroblasts and cell lines. PLoS ONE 2013, 8, e71320. [Google Scholar] [CrossRef] [PubMed]
- Van Pesch, V.; Lanaya, H.; Renauld, J.C.; Michiels, T. Characterization of the murine alpha interferon gene family. J. Virol. 2004, 78, 8219–8228. [Google Scholar] [CrossRef] [PubMed]
- Van Pesch, V.; Michiels, T. Characterization of interferon-alpha 13, a novel constitutive murine interferon-alpha subtype. J. Biol. Chem. 2003, 278, 46321–46328. [Google Scholar] [CrossRef]
- Chhatbar, C.; Detje, C.N.; Grabski, E.; Borst, K.; Spanier, J.; Ghita, L.; Elliott, D.A.; Jordao, M.J.C.; Mueller, N.; Sutton, J.; et al. Type i interferon receptor signaling of neurons and astrocytes regulates microglia activation during viral encephalitis. Cell reports 2018, 25, 118–129 e114. [Google Scholar] [CrossRef]
- Detje, C.N.; Lienenklaus, S.; Chhatbar, C.; Spanier, J.; Prajeeth, C.K.; Soldner, C.; Tovey, M.G.; Schluter, D.; Weiss, S.; Stangel, M.; et al. Upon intranasal vesicular stomatitis virus infection, astrocytes in the olfactory bulb are important interferon beta producers that protect from lethal encephalitis. J. Virol. 2015, 89, 2731–2738. [Google Scholar] [CrossRef]
- Parakkal, P.F.; Gregoire, A.T. Differentiation of vaginal epithelium in the normal and hormone-treated rhesus monkey. Biol. Reprod. 1972, 6, 117–130. [Google Scholar] [CrossRef][Green Version]
- Miyagawa, S.; Iguchi, T. Epithelial estrogen receptor 1 intrinsically mediates squamous differentiation in the mouse vagina. Proc. Natl. Acad. Sci. USA 2015, 112, 12986–12991. [Google Scholar] [CrossRef]
- Wira, C.R.; Rodriguez-Garcia, M.; Patel, M.V. The role of sex hormones in immune protection of the female reproductive tract. Nat. Rev. Immunol. 2015, 15, 217–230. [Google Scholar] [CrossRef]
- Caine, E.A.; Scheaffer, S.M.; Arora, N.; Zaitsev, K.; Artyomov, M.N.; Coyne, C.B.; Moley, K.H.; Diamond, M.S. Interferon lambda protects the female reproductive tract against zika virus infection. Nat. Commun. 2019, 10, 280. [Google Scholar] [CrossRef] [PubMed]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jacobs, S.; Zeippen, C.; Wavreil, F.; Gillet, L.; Michiels, T. IFN-λ Decreases Murid Herpesvirus-4 Infection of the Olfactory Epithelium but Fails to Prevent Virus Reactivation in the Vaginal Mucosa. Viruses 2019, 11, 757. https://doi.org/10.3390/v11080757
Jacobs S, Zeippen C, Wavreil F, Gillet L, Michiels T. IFN-λ Decreases Murid Herpesvirus-4 Infection of the Olfactory Epithelium but Fails to Prevent Virus Reactivation in the Vaginal Mucosa. Viruses. 2019; 11(8):757. https://doi.org/10.3390/v11080757
Chicago/Turabian StyleJacobs, Sophie, Caroline Zeippen, Fanny Wavreil, Laurent Gillet, and Thomas Michiels. 2019. "IFN-λ Decreases Murid Herpesvirus-4 Infection of the Olfactory Epithelium but Fails to Prevent Virus Reactivation in the Vaginal Mucosa" Viruses 11, no. 8: 757. https://doi.org/10.3390/v11080757
APA StyleJacobs, S., Zeippen, C., Wavreil, F., Gillet, L., & Michiels, T. (2019). IFN-λ Decreases Murid Herpesvirus-4 Infection of the Olfactory Epithelium but Fails to Prevent Virus Reactivation in the Vaginal Mucosa. Viruses, 11(8), 757. https://doi.org/10.3390/v11080757