Generation of Monoclonal Antibodies against Variable Epitopes of the M Protein of Rabies Virus

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses and Cells
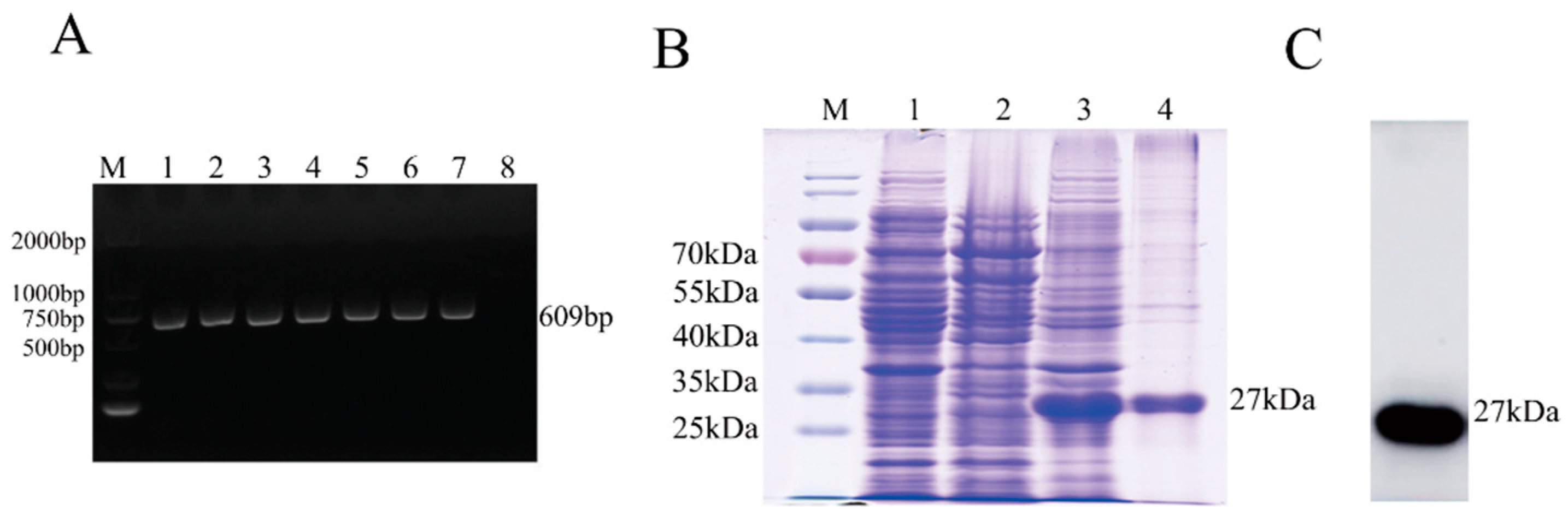
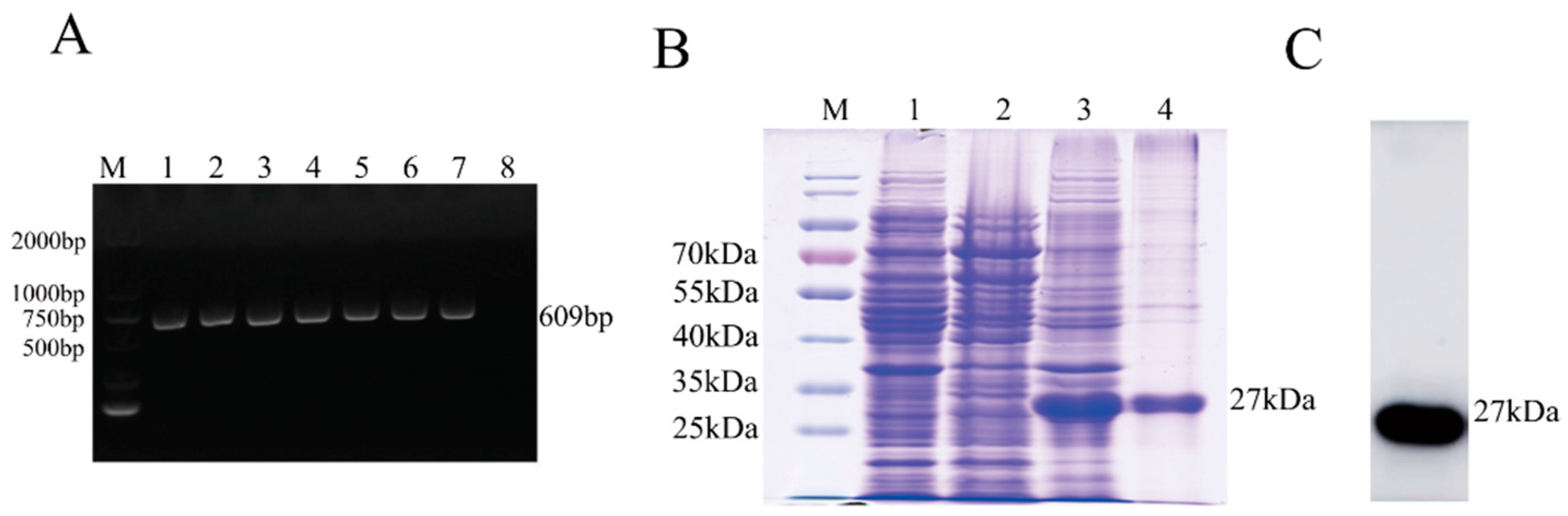
2.2. Expression and Purification of Recombinant M Protein
2.3. Preparation of Monoclonal Antibodies Against the M Protein
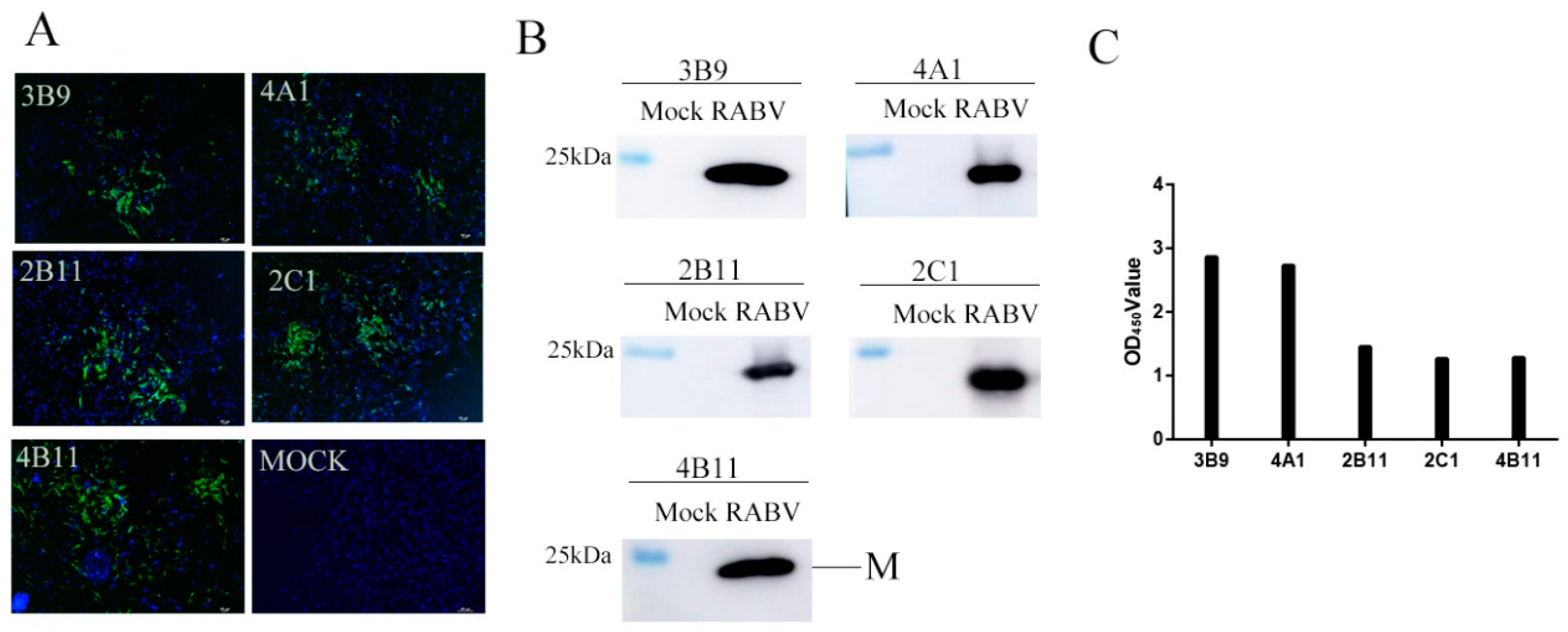
2.4. Screening of Anti-RABV M Antibodies Using Indirect ELISA
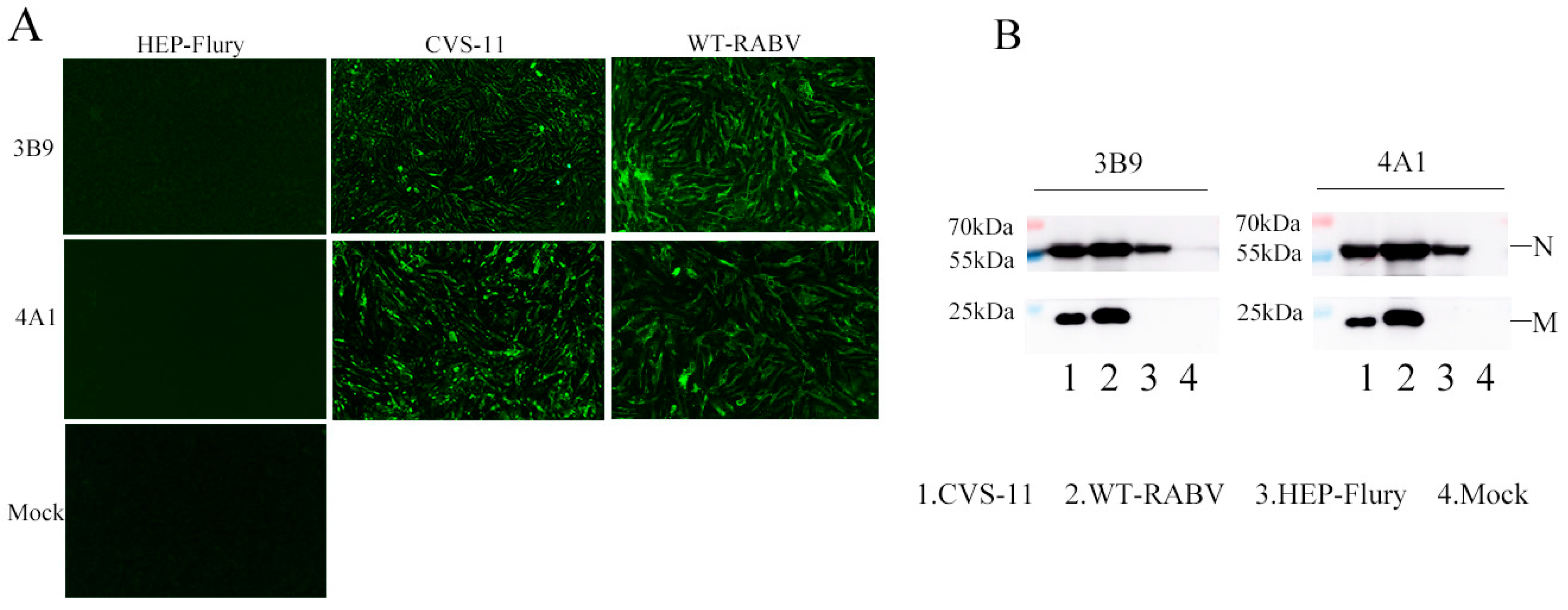
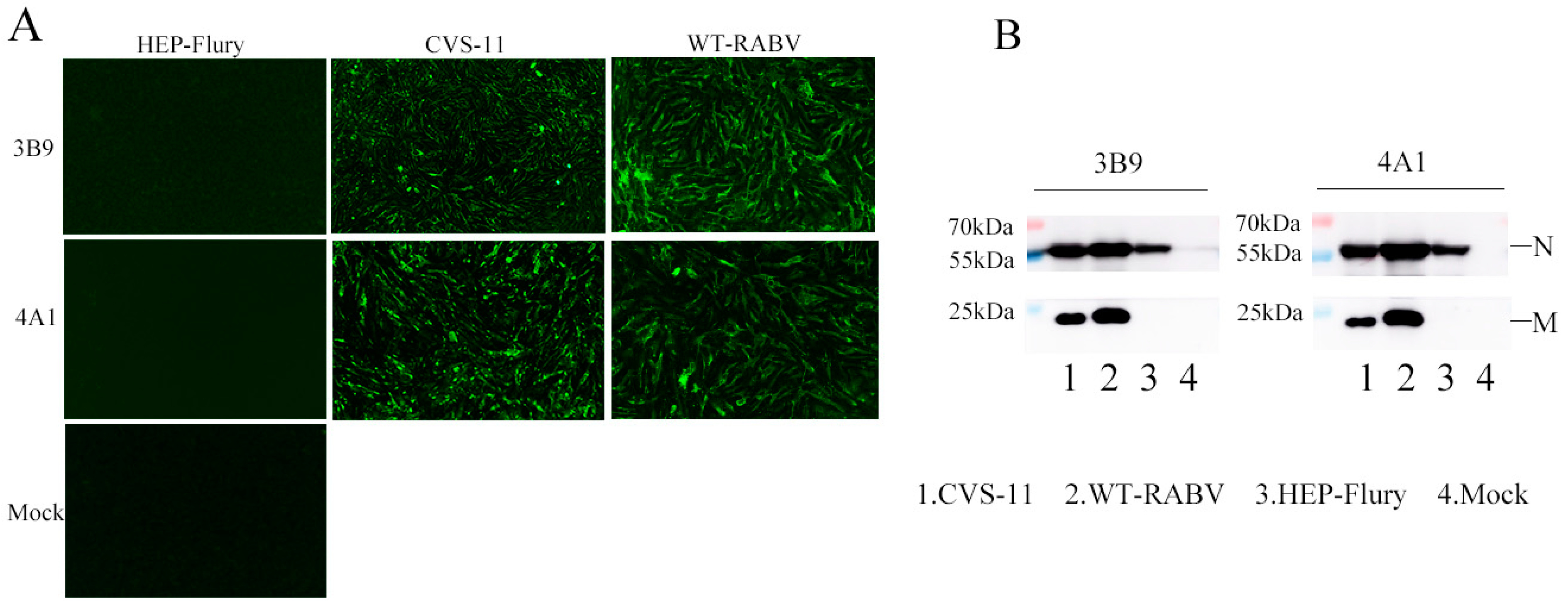
2.5. SDS-PAGE and Western Blotting
2.6. Indirect Immunofluorescence Assay
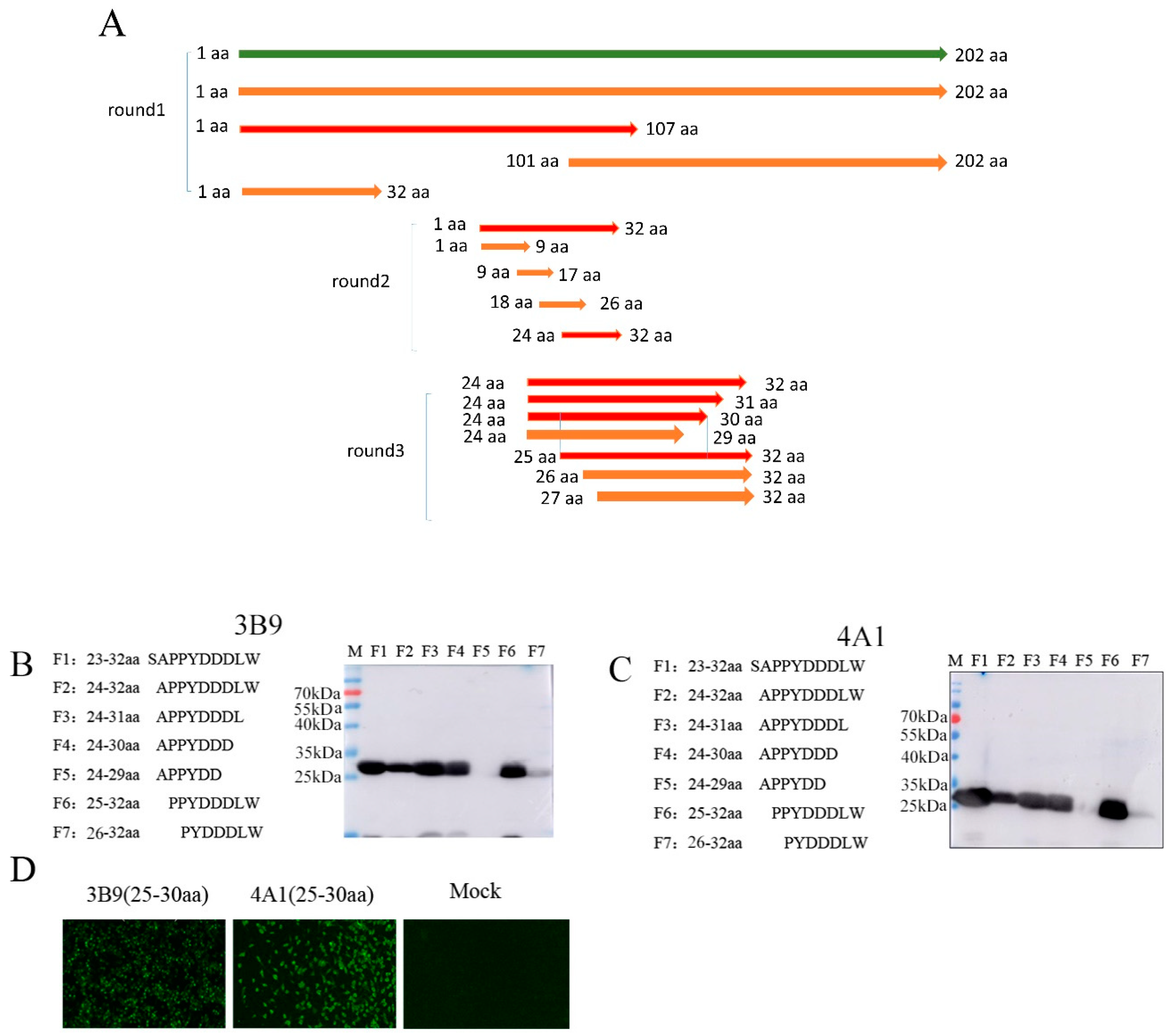
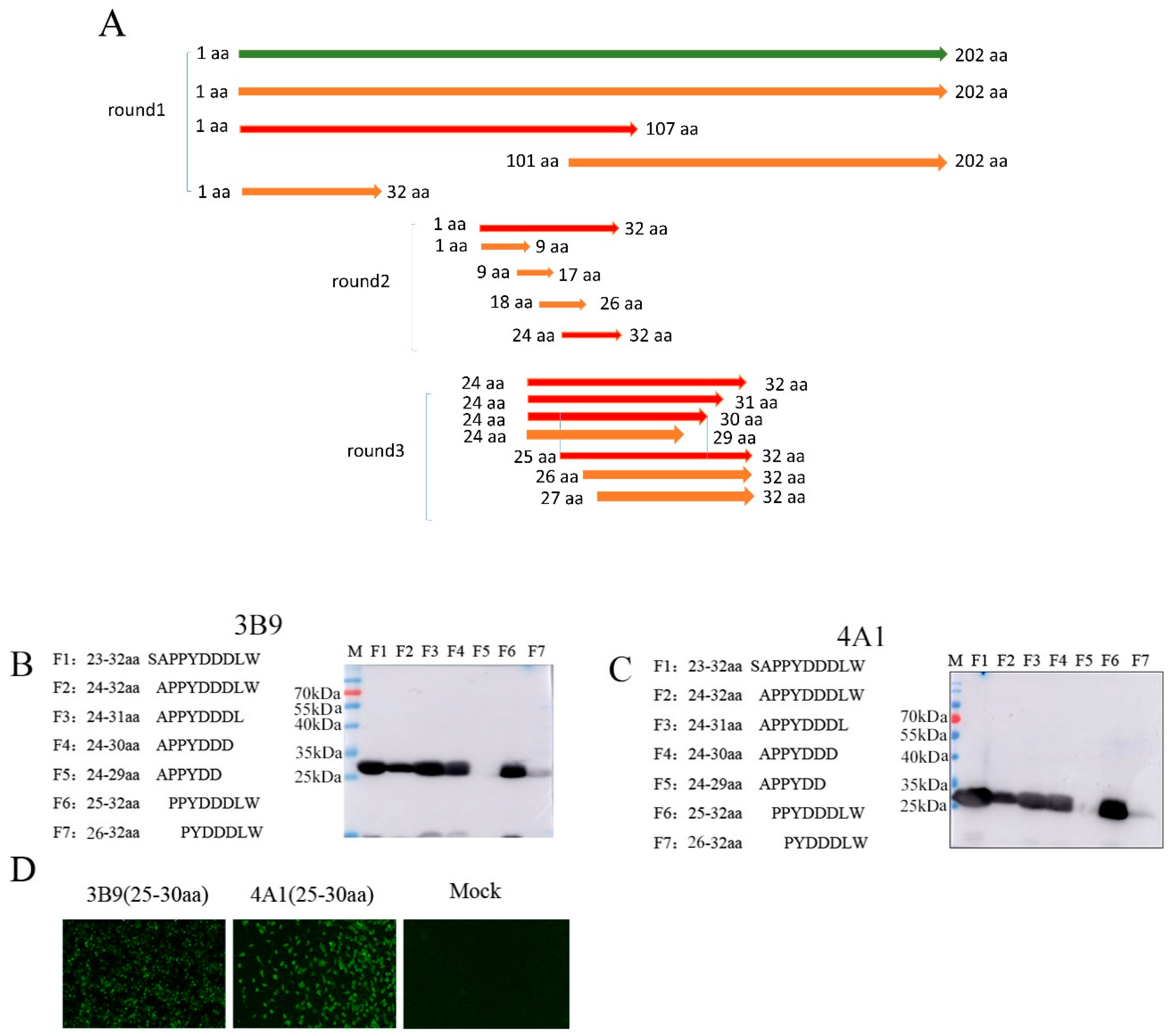
2.7. Identification of M Protein Epitopes
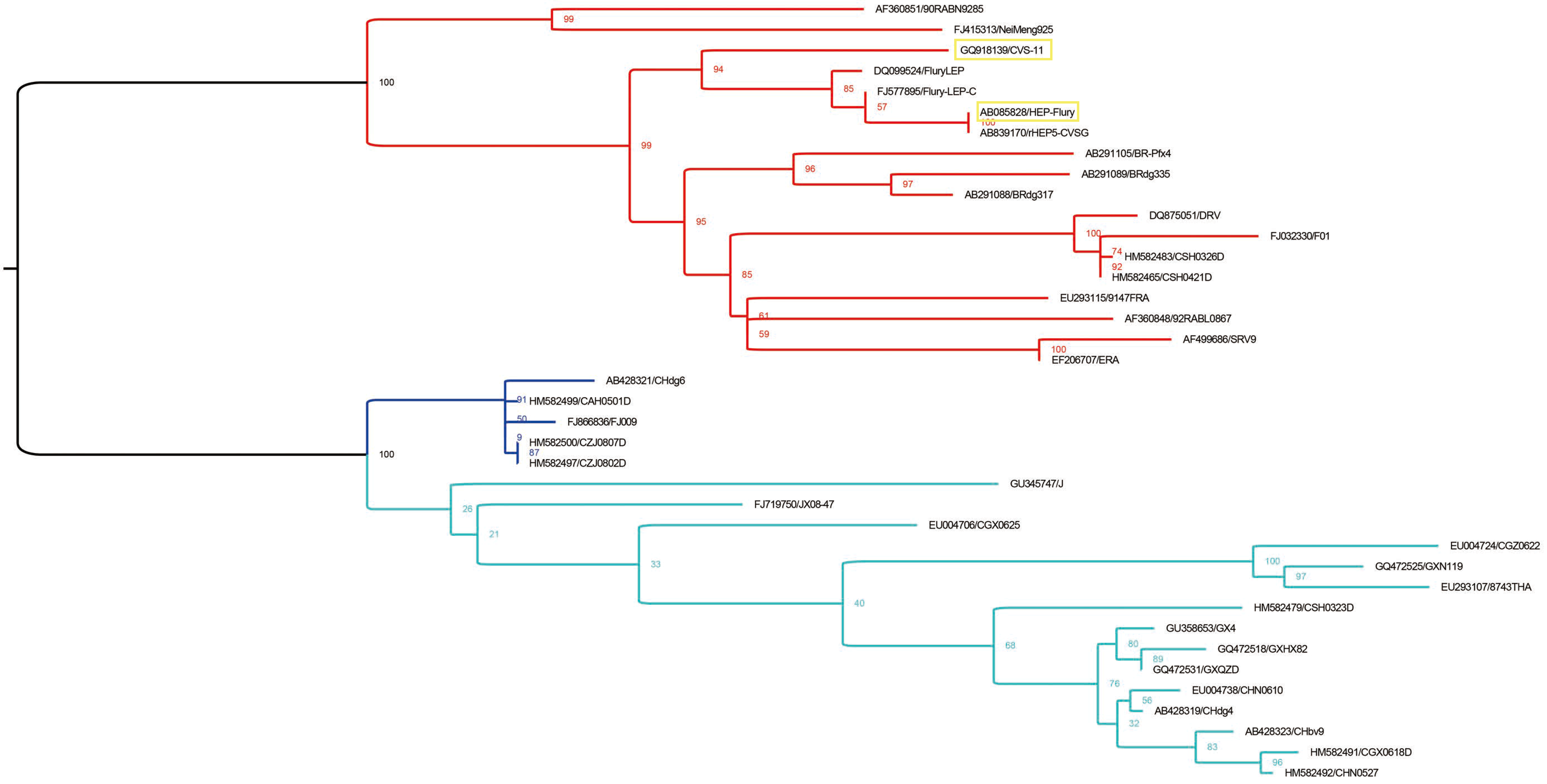
2.8. Alignment of Epitope Sequences of the M Protein
2.9. Application of mAbs in the Detection of RABV by Immunohistochemistry
3. Results
3.1. Expression and Purification of the M Protein
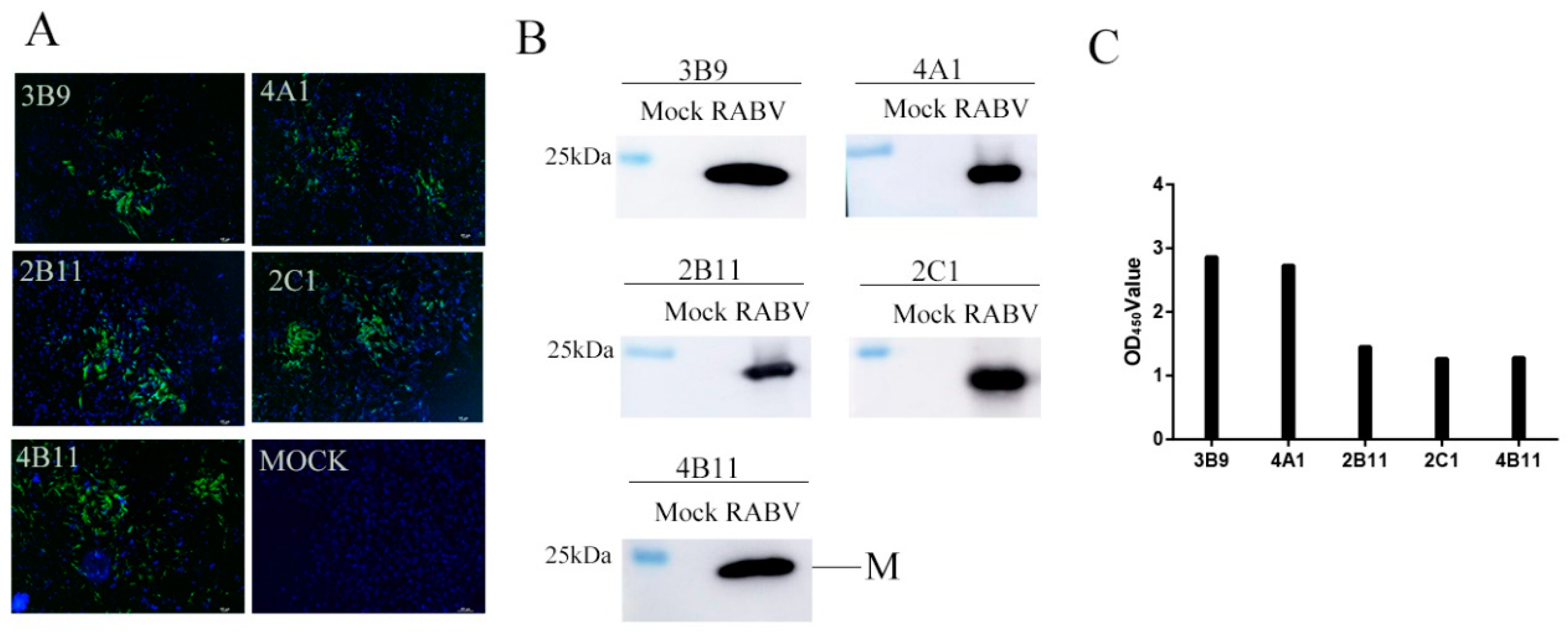
3.2. Generation and Characterization of mAbs Against the M Protein
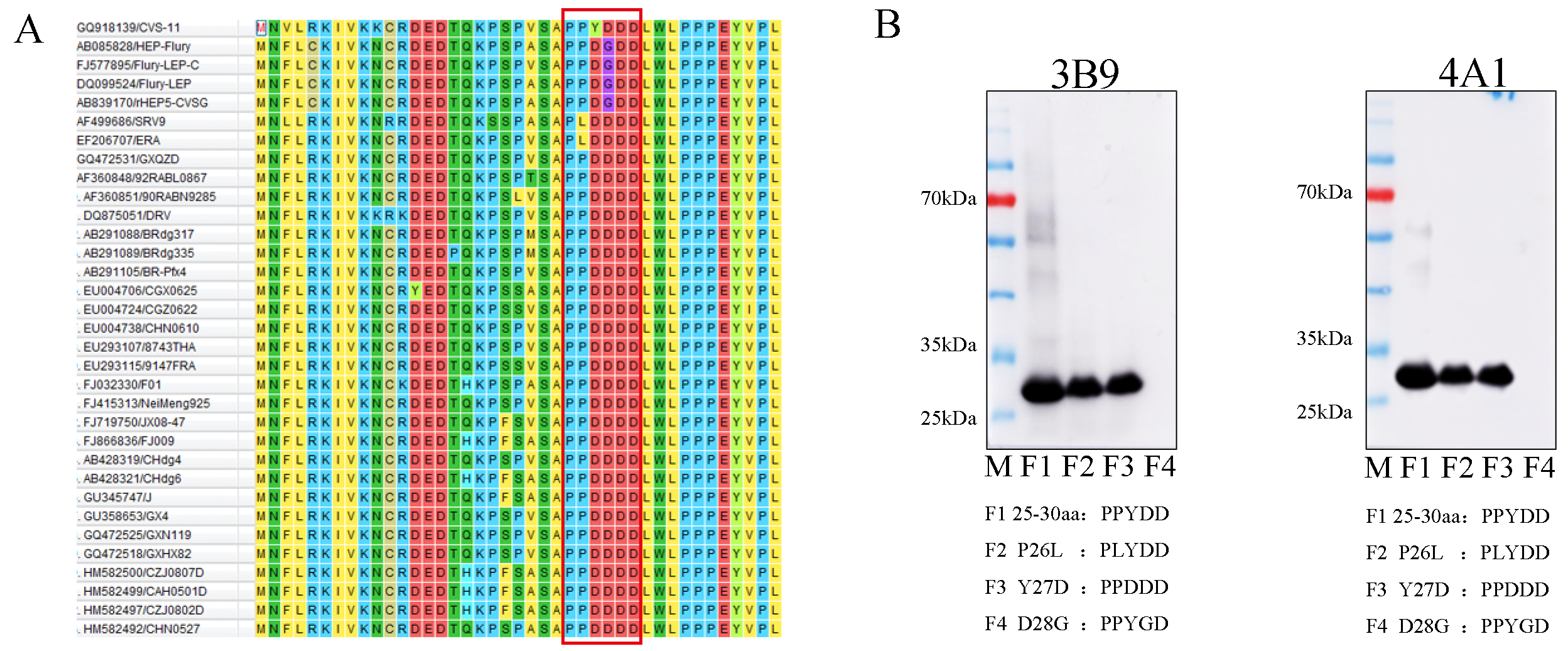
3.3. Mapping of the Epitopes of M Protein
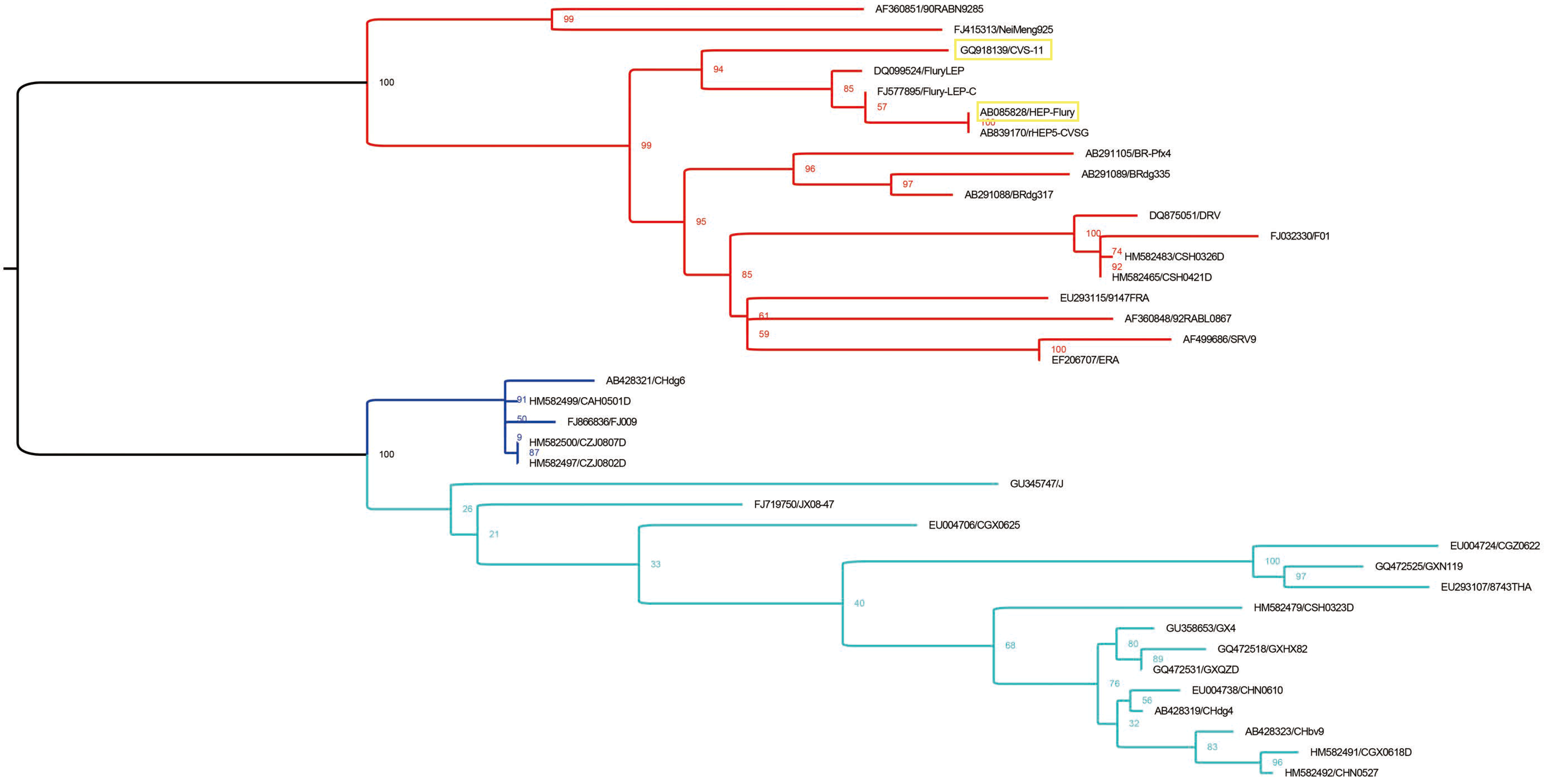
3.4. Alignment of M Protein Epitope Sequences
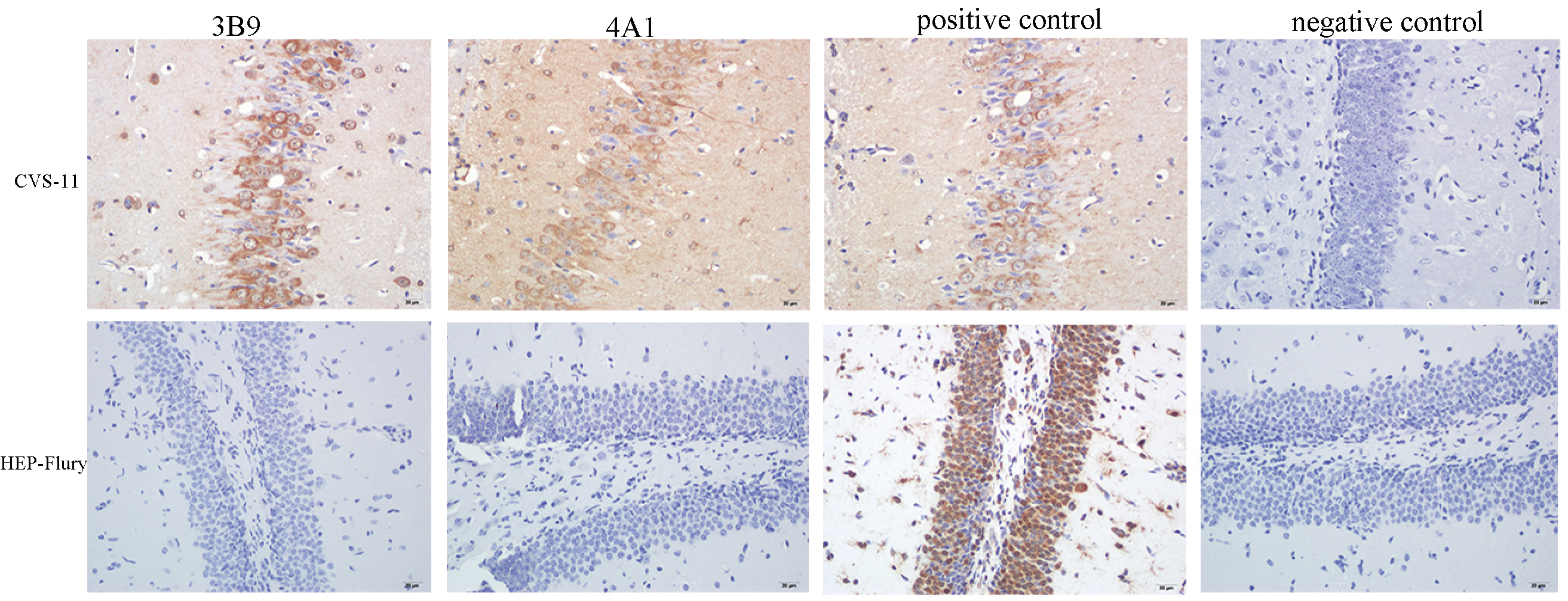
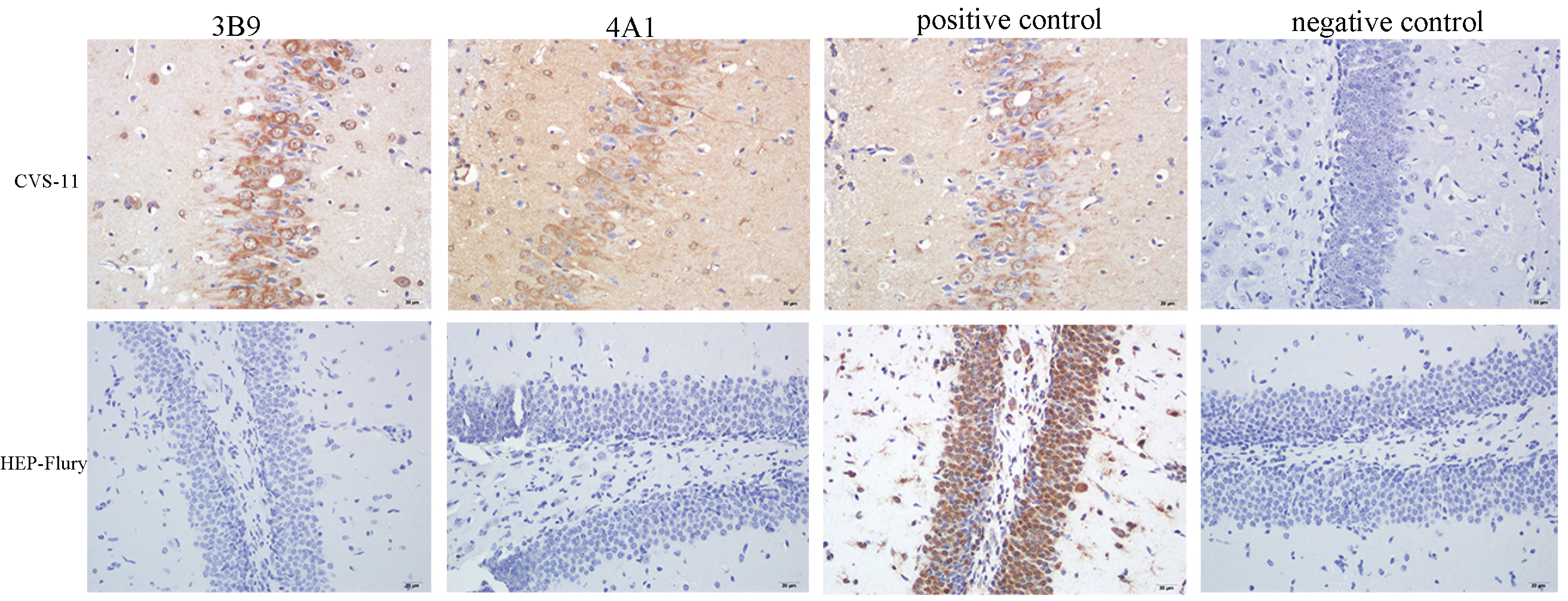
3.5. Reactivity of mAbs with RABV in the Brain Tissue of Infected Mice Using Immunohistochemistry (IHC)
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wilde, H.; Hemachudha, T.; Tantawichien, T.; Khawplod, P. Rabies and other lyssavirus diseases. Lancet 2004, 363, 1906, author reply 1907. [Google Scholar] [CrossRef]
- Jacob, Y.; Badrane, H.; Ceccaldi, P.E.; Tordo, N. Cytoplasmic dynein LC8 interacts with lyssavirus phosphoprotein. J. Virol. 2000, 74, 10217–10222. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Wang, R.; Ji, S.; Su, S.; Zhou, J. One Health strategies for rabies control in rural areas of China. Lancet Infect. Dis. 2017, 17, 365–367. [Google Scholar] [CrossRef]
- Schnell, M.J.; McGettigan, J.P.; Wirblich, C.; Papaneri, A. The cell biology of rabies virus: Using stealth to reach the brain. Nat. Rev. Microbiol. 2010, 8, 51–61. [Google Scholar] [CrossRef]
- Tordo, N.; Poch, O.; Ermine, A.; Keith, G. Primary structure of leader RNA and nucleoprotein genes of the rabies genome: Segmented homology with VSV. Nucleic Acids Res. 1986, 14, 2671–2683. [Google Scholar] [CrossRef] [PubMed]
- Ameyama, S.; Toriumi, H.; Takahashi, T.; Shimura, Y.; Nakahara, T.; Honda, Y.; Mifune, K.; Uchiyama, T.; Kawai, A. Monoclonal antibody #3-9-16 recognizes one of the two isoforms of rabies virus matrix protein that exposes its N-terminus on the virion surface. Microbiol. Immunol. 2003, 47, 639–651. [Google Scholar] [PubMed]
- Odenwald, W.F.; Arnheiter, H.; Dubois-Dalcq, M.; Lazzarini, R.A. Stereo images of vesicular stomatitis virus assembly. J. Virol. 1986, 57, 922–932. [Google Scholar]
- Morimoto, K.; Patel, M.; Corisdeo, S.; Hooper, D.C.; Fu, Z.F.; Rupprecht, C.E.; Koprowski, H.; Dietzschold, B. Characterization of a unique variant of bat rabies virus responsible for newly emerging human cases in North America. Proc. Natl. Acad. Sci. USA 1996, 93, 5653–5658. [Google Scholar] [CrossRef]
- Whitt, M.A. Generation of VSV pseudotypes using recombinant DeltaG-VSV for studies on virus entry, identification of entry inhibitors, and immune responses to vaccines. J. Virol. Methods 2010, 169, 365–374. [Google Scholar] [CrossRef]
- Mebatsion, T.; Weiland, F.; Conzelmann, K.K. Matrix protein of rabies virus is responsible for the assembly and budding of bullet-shaped particles and interacts with the transmembrane spike glycoprotein G. J. Virol. 1999, 73, 242–250. [Google Scholar] [PubMed]
- Okumura, A.; Harty, R.N. Rabies virus assembly and budding. Adv. Virus Res. 2011, 79, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Finke, S.; Mueller-Waldeck, R.; Conzelmann, K.K. Rabies virus matrix protein regulates the balance of virus transcription and replication. J. Gen. Virol. 2003, 84, 1613–1621. [Google Scholar] [CrossRef] [PubMed]
- Pulmanausahakul, R.; Li, J.; Schnell, M.J.; Dietzschold, B. The glycoprotein and the matrix protein of rabies virus affect pathogenicity by regulating viral replication and facilitating cell-to-cell spread. J. Virol. 2008, 82, 2330–2338. [Google Scholar] [CrossRef] [PubMed]
- Kassis, R.; Larrous, F.; Estaquier, J.; Bourhy, H. Lyssavirus matrix protein induces apoptosis by a TRAIL-dependent mechanism involving caspase-8 activation. J. Virol. 2004, 78, 6543–6555. [Google Scholar] [CrossRef] [PubMed]
- Zan, J.; Liu, J.; Zhou, J.W.; Wang, H.L.; Mo, K.K.; Yan, Y.; Xu, Y.B.; Liao, M.; Su, S.; Hu, R.L.; et al. Rabies virus matrix protein induces apoptosis by targeting mitochondria. Exp. Cell Res. 2016, 347, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Wang, L.; Tao, X.; Li, H.; Rayner, S.; Liang, G.; Tang, Q. Genetic diversity and molecular evolution of the rabies virus matrix protein gene in China. Infect. Genet. Evol. 2013, 16, 248–253. [Google Scholar] [CrossRef]
- Bi, Z.; Xia, X.; Wang, Y.; Mei, Y. Development and characterization of neutralizing monoclonal antibodies against canine distemper virus hemagglutinin protein. Microbiol. Immunol. 2015, 59, 202–208. [Google Scholar] [CrossRef]
- Ito, N.; Mita, T.; Shimizu, K.; Ito, Y.; Masatani, T.; Nakagawa, K.; Yamaoka, S.; Abe, M.; Okadera, K.; Minamoto, N.; et al. Amino acid substitution at position 95 in rabies virus matrix protein affects viral pathogenicity. J. Vet. Med. Sci. 2011, 73, 1363–1366. [Google Scholar] [CrossRef]
- Bi, Z.; Wang, Y.; Pan, Q.; Xia, X.; Xu, L. Development of CDV-specific monoclonal antibodies for differentiation of variable epitopes of nucleocapsid protein. Vet. Microbiol. 2017, 211, 84–91. [Google Scholar] [CrossRef]
- Peterson, N.C. Behavioral, clinical, and physiologic analysis of mice used for ascites monoclonal antibody production. Comp. Med. 2000, 50, 516–526. [Google Scholar]
- Schagger, H.; von Jagow, G. Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal. Biochem. 1987, 166, 368–379. [Google Scholar] [CrossRef]
- Zhang, J.; Ruan, X.; Zan, J.; Zheng, X.; Yan, Y.; Liao, M.; Zhou, J. Efficient generation of monoclonal antibodies against major structural proteins of rabies virus with suckling mouse brain antigen. Monoclon. Antib. Immunodiagn. Immunother. 2014, 33, 94–100. [Google Scholar] [CrossRef]
- Feng, M.; Yuan, Z.; Xia, W.; Huang, X.; Wang, X.; Yan, Y.; Liao, M.; Zhou, J. Monoclonal antibody against the universal M2 epitope of influenza A virus. Appl. Microbiol. Biotechnol. 2018, 102, 5645–5656. [Google Scholar] [CrossRef]
- Rupprecht, C.E.; Hanlon, C.A.; Hemachudha, T. Rabies re-examined. Lancet Infect. Dis. 2002, 2, 327–343. [Google Scholar] [CrossRef]
- Davis, B.M.; Rall, G.F.; Schnell, M.J. Everything You Always Wanted to Know About Rabies Virus (But Were Afraid to Ask). Annu. Rev. Virol. 2015, 2, 451–471. [Google Scholar] [CrossRef] [PubMed]
- Wiktor, T.J.; Koprowski, H. Monoclonal antibodies against rabies virus produced by somatic cell hybridization: detection of antigenic variants. Proc. Natl. Acad. Sci. USA 1978, 75, 3938–3942. [Google Scholar] [CrossRef]
- Marissen, W.E.; Kramer, R.A.; Rice, A.; Weldon, W.C.; Niezgoda, M.; Faber, M.; Slootstra, J.W.; Meloen, R.H.; Clijsters-van der Horst, M.; Visser, T.J.; et al. Novel rabies virus-neutralizing epitope recognized by human monoclonal antibody: fine mapping and escape mutant analysis. J. Virol. 2005, 79, 4672–4678. [Google Scholar] [CrossRef] [PubMed]
- Hiramatsu, K.; Mifune, K.; Mannen, K.; Nishizono, A.; Kawano, H.; Ito, Y.; Kawai, A. Mapping of the antigenic determinants recognized by monoclonal antibodies against the M2 protein of rabies virus. Virology 1992, 187, 472–479. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fragment | Primer Sequence (5′–3′) |
|---|---|
| 1–203 aa | GGGGTACCATGAACGTTCTACGCAAGAT (HindIII) |
| CCCAAGCTTTTATTCTAGAAGCAGAGAAGAG (KpnI) | |
| 1–107 aa | GGGGTACCATGAACGTTCTACGCAAGAT (HindIII) |
| CCCAAGCTTAGGTACTGGAGCTCCTGATAAA (KpnI) | |
| 101–203 aa | GGGGTACCTTATCAGGAGCTCCAGTACCT (HindIII) |
| CCCAAGCTTTTATTCTAGAAGCAGAGAAGAG (KpnI) | |
| 1–32 aa | GGGGTACCATGAACGTTCTACGCAAGATA (HindIII) |
| CCCAAGCTTCCACAGGTCATCGTCATACGG (KpnI) | |
| 26–36 aa | GGGGTACCTATGACGATGACCTGTGGCTTCCACCTCCT (HindIII) |
| CCCAAGCTTAGGAGGTGGAAGCCACAGGTCATCGTCATACGG (KpnI) | |
| 33–203 aa | GGGGTACCTGACCTGTGGCTTCCACCTC (HindIII) |
| CCCAAGCTTTTATTCTAGAAGCAGAGAAGAG (KpnI) | |
| 1–17 aa | GGGGTACCATGAACGTTCTACGCAAGAT (HindIII) |
| TAGGGATGAGGACACTCAAAAGCTTGGG (KpnI) | |
| 17–26 aa | GGGGTACCCAAAAGCCCTCTCCTGTGT (HindIII) |
| CCCAAGCTTCCACAGGTCATCGTCATACGG (KpnI) |
| Fragment | Primer Sequence (5′–3′) |
|---|---|
| 17–25 aa | CCAAAAGCCCTCTCCTGTGTCAGCCCCTA |
| CATGGGTTTTCGGGAGAGGACACAGTCGGGGATTCGA | |
| 24–32 aa | CGCCCCTCCGTATGACGATGACCTGTGGA |
| CATGGCGGGGAGGCATACTGCTACTGGACACCTTCGA | |
| 24–31 aa | GATCCGCCCCTCCGTATGACGATGACCTGC |
| TCGAGCAGGTCATCGTCATACGGAGGGGCG | |
| 24–30 aa | GATCCGCCCCTCCGTATGACGATGACC |
| TCGAGGTCATCGTCATACGGAGGGGCG | |
| 24–29 aa | GATCCGCCCCTCCGTATGACGATC |
| TCGAGATCGTCATACGGAGGGGCG | |
| 24–28 aa | GATCCGCCCCTCCGTATGACC |
| TCGAGGTCATACGGAGGGGCG | |
| 25–32 aa | GATCCCCTCCGTATGACGATGACCTGTGGC |
| TCGAGCCACAGGTCATCGTCATACGGAGGG | |
| 26–32 aa | GATCCCCGTATGACGATGACCTGTGGC |
| TCGAGCCACAGGTCATCGTCATACGGG | |
| 27–32 aa | GATCCTATGACGATGACCTGTGGC |
| TCGAGCCACAGGTCATCGTCATAG | |
| 28–32 aa | GATCCGACGATGACCTGTGGC |
| 25–30 aa | TCGAGCCACAGGTCATCGTCG GATCCCCTCCGTATGACGATGACC TCGAGGTCATCGTCATACGGAGGG |
| 25–30 aa (pCMV-Flag) | AGCTTCCTCCGTATGACGATGACGGTAC AGTCATCGTCATACGGAGGC |
| Fragment | Primer Sequence (5′–3′) |
|---|---|
| P26L mutation | GATCCCCTCTGTATGACGATGACC |
| TCGAGGTCATCGTCATACAGAGGG | |
| Y27D mutation | GATCCCCTCCGGATGACGATGACC |
| TCGAGGTCATCGTCATCCGGAGGG | |
| D28G mutation | GATCCCCTCCGTATGGCGATGACC |
| TCGAGGTCATCGCCATACGGAGGG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Zhao, W.; He, W.; Wang, N.; Su, J.; Ji, S.; Chen, J.; Wang, D.; Zhou, J.; Su, S. Generation of Monoclonal Antibodies against Variable Epitopes of the M Protein of Rabies Virus. Viruses 2019, 11, 375. https://doi.org/10.3390/v11040375
Liu J, Zhao W, He W, Wang N, Su J, Ji S, Chen J, Wang D, Zhou J, Su S. Generation of Monoclonal Antibodies against Variable Epitopes of the M Protein of Rabies Virus. Viruses. 2019; 11(4):375. https://doi.org/10.3390/v11040375
Chicago/Turabian StyleLiu, Jie, Wen Zhao, Wanting He, Ningning Wang, Jingyin Su, Senlin Ji, Jian Chen, Dong Wang, Jiyong Zhou, and Shuo Su. 2019. "Generation of Monoclonal Antibodies against Variable Epitopes of the M Protein of Rabies Virus" Viruses 11, no. 4: 375. https://doi.org/10.3390/v11040375
APA StyleLiu, J., Zhao, W., He, W., Wang, N., Su, J., Ji, S., Chen, J., Wang, D., Zhou, J., & Su, S. (2019). Generation of Monoclonal Antibodies against Variable Epitopes of the M Protein of Rabies Virus. Viruses, 11(4), 375. https://doi.org/10.3390/v11040375