Characterization of Three Novel Viruses from the Families Nyamiviridae, Orthomyxoviridae, and Peribunyaviridae, Isolated from Dead Birds Collected during West Nile Virus Surveillance in Harris County, Texas

 , ,
, ,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Collection of Dead Birds
2.3. Culture Methods
2.4. Evaluation of Pathogenicity in Suckling Mice
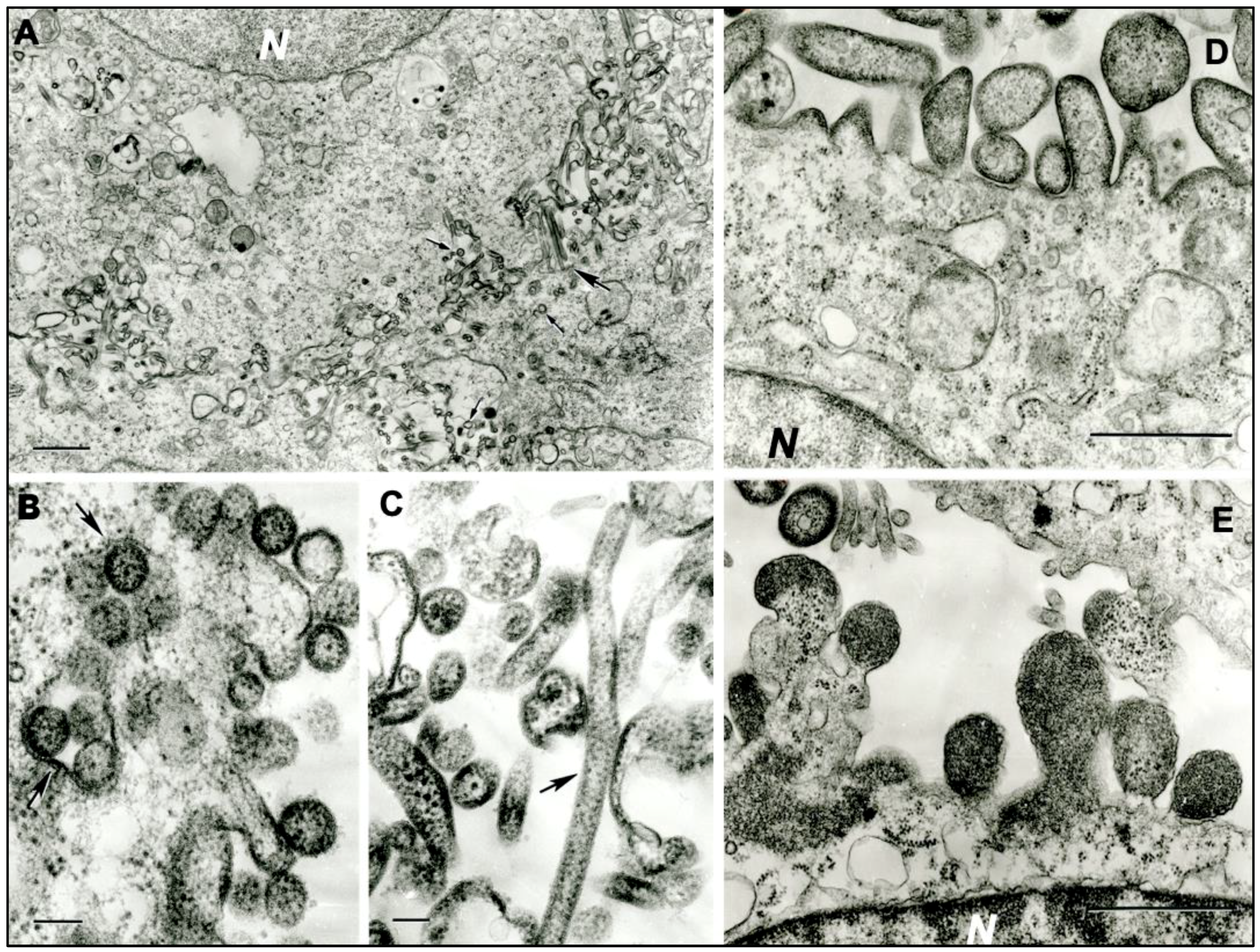
2.5. Transmission Electron Microscopy (TEM)
2.6. RNA Extraction, Viral Genome Sequencing, and Assembly
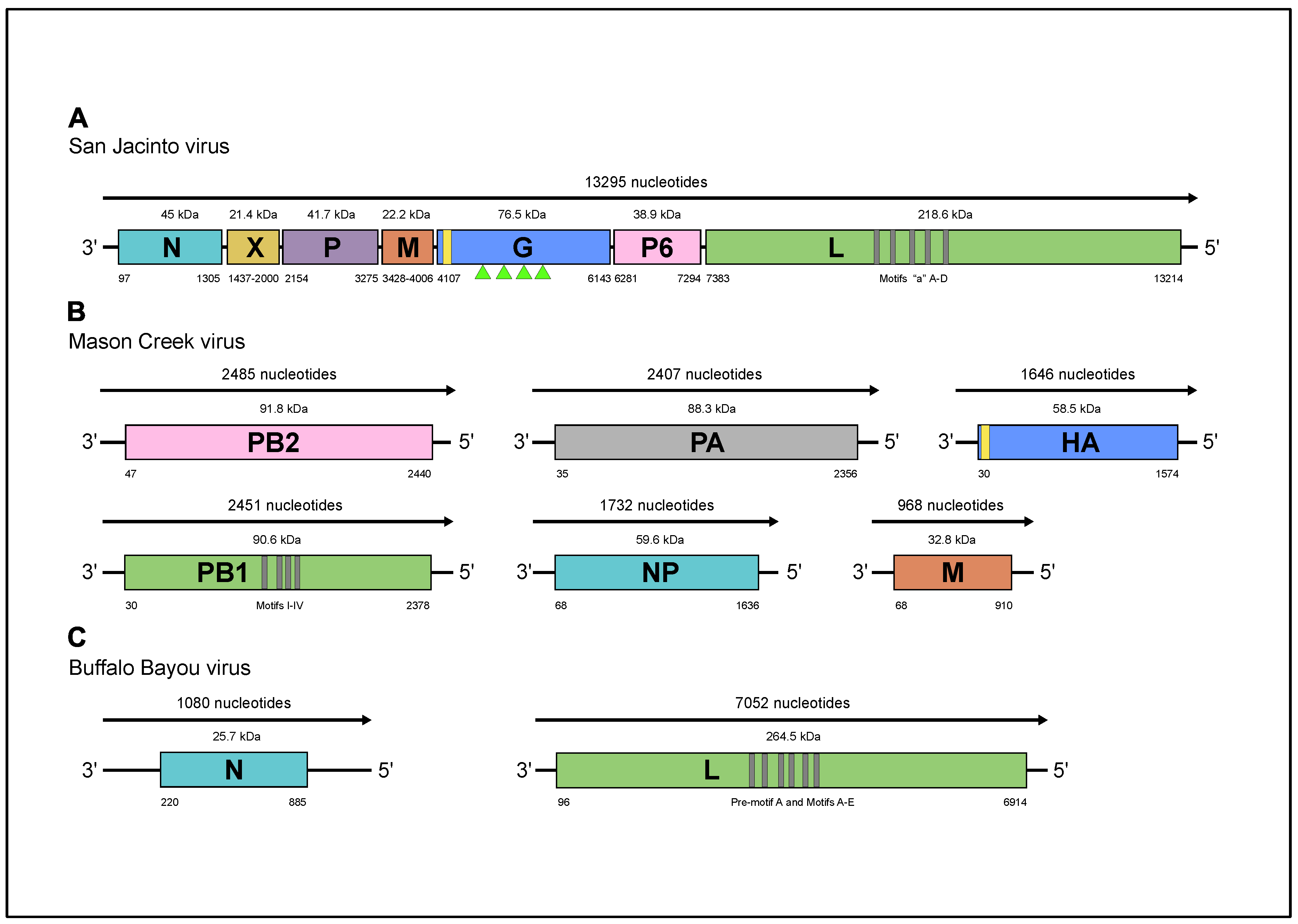
2.7. Viral Genome Annotation
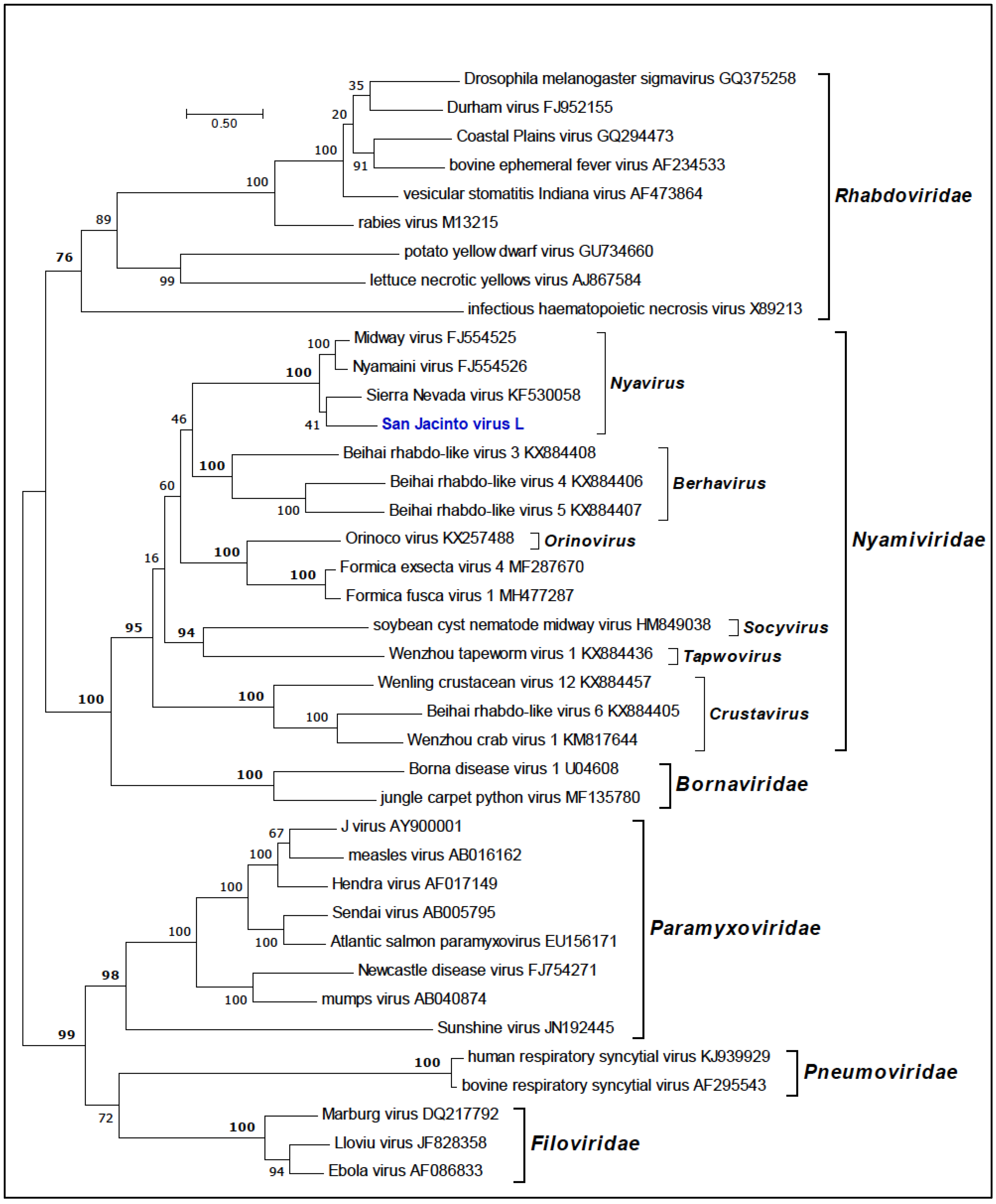
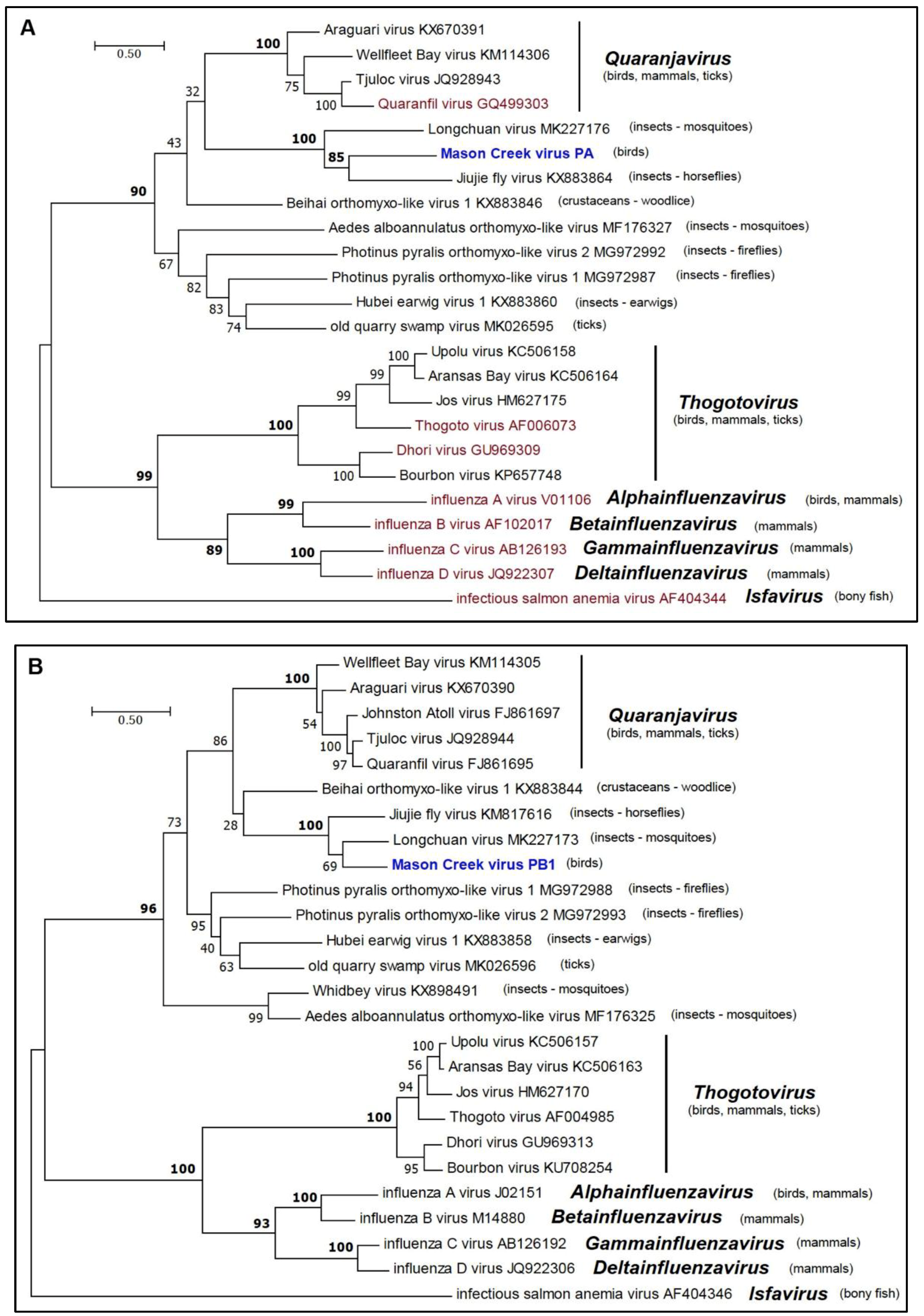
2.8. Phylogenetic Analyses
2.9. Serological Tests
3. Results
3.1. Virus Isolation and Initial Characterization
3.2. Transmission Electron Microscopy
3.3. Nucleotide Sequence Analysis
3.4. Complement Fixation Tests
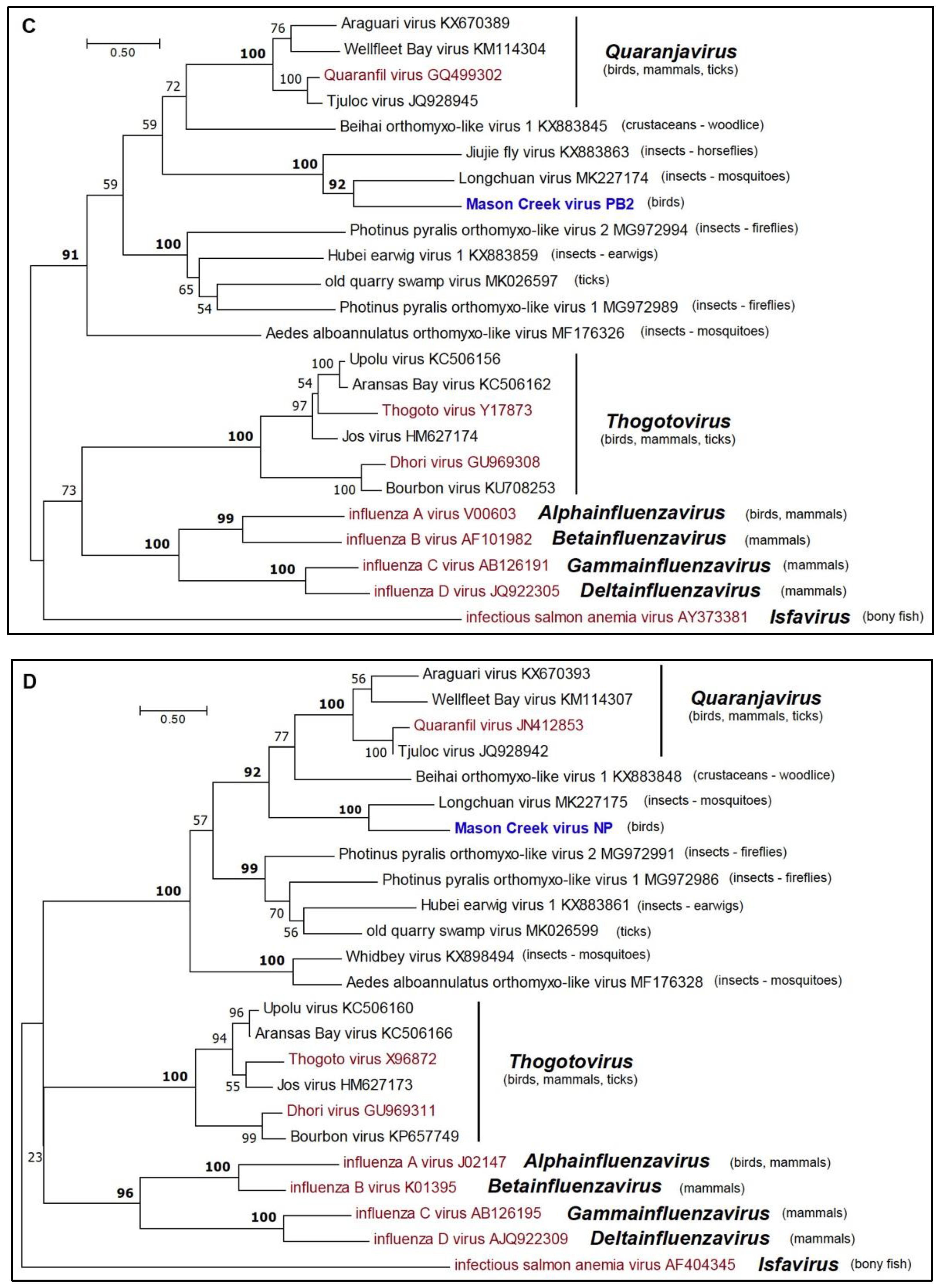
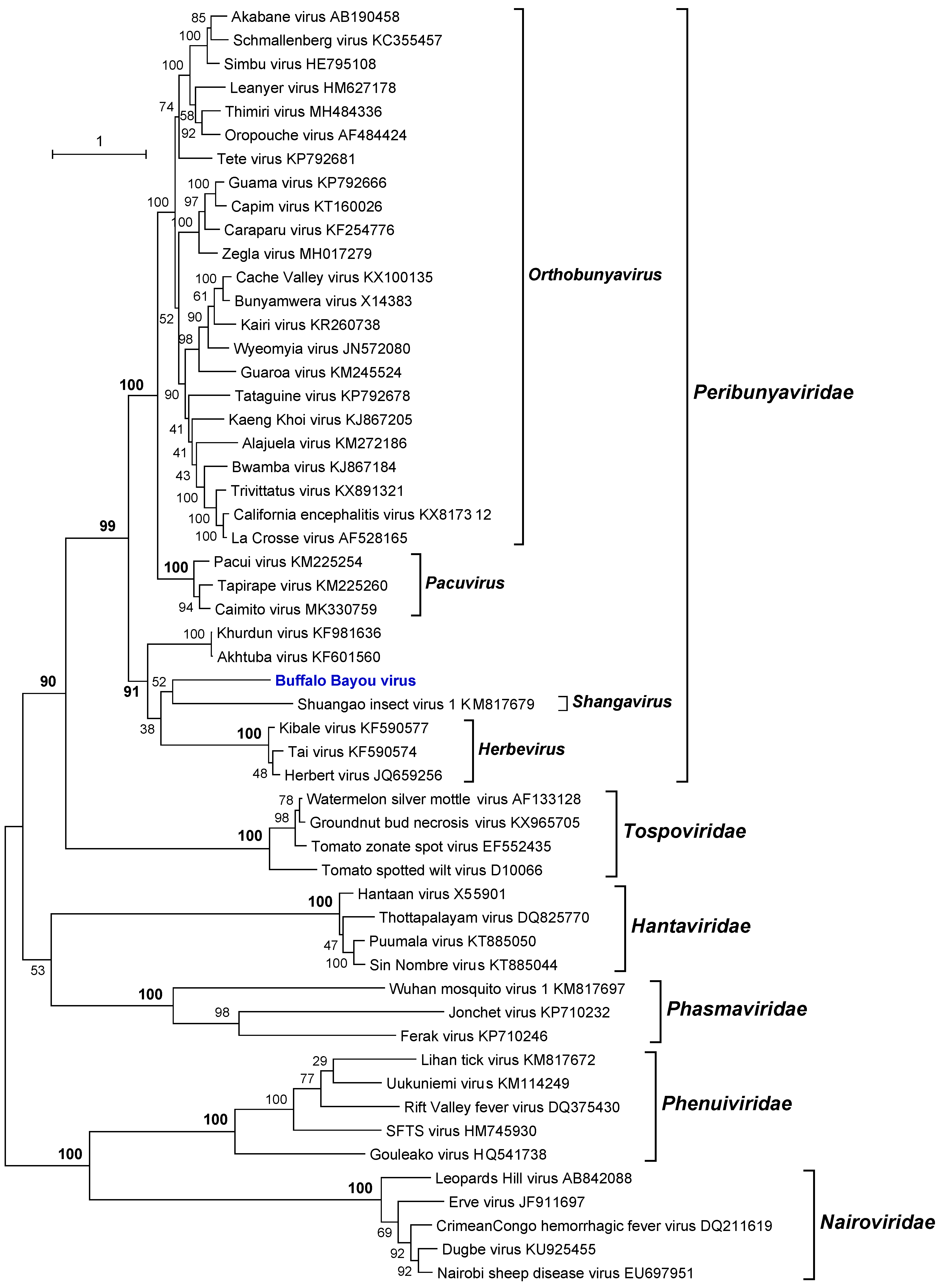
3.5. Phylogenetic Analyses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lillibridge, K.M.; Parsons, R.; Randle, Y.; Travassos da Rosa, A.P.; Guzman, H.; Siirin, M.; Wuithiranyagool, T.; Hailey, C.; Higgs, S.; Bala, A.A.; et al. The 2002 introduction of West Nile virus into Harris County, Texas, an area historically endemic for St. Louis encephalitis. Am. J. Trop. Med. Hyg. 2004, 70, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Wilkerson, L.; Reyna Nava, M.; Battle-Freeman, C.; Travassos da Rosa, A.; Guzman, H.; Tesh, R.; Debboun, M. The role of birds in arboviral disease surveillance in Harris County and the City of Houston, Texas. US Army Med. Dep. J. 2017, (1-17), 1–12. [Google Scholar]
- Mann, B.R.; McMullen, A.R.; Swetnam, D.M.; Salvato, V.; Reyna, M.; Guzman, H.; Bueno, R.; Dennett, J.A.; Tesh, R.B.; Barrett, A.D. Continued evolution of West Nile virus, Houston, Texas, USA, 2002-2012. Emerg. Infect. Dis. 2013, 19, 1418–1427. [Google Scholar] [CrossRef] [PubMed]
- Kim, L.M.; King, D.J.; Guzman, H.; Tesh, R.B.; Travassos da Rosa, A.P.; Bueno, R.; Dennett, J.A.; Afonso, C.L. Biological and phylogenetic characterization of pigeon paramyxovirus serotype 1 circulating in wild North American pigeons and doves. J. Clin. Microbiol. 2008, 46, 3303–3310. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.B.; Gulino, K.M.; Tesh, R.B.; Cui, L.; Fitch, A.; Unnasch, T.R.; Popov, V.L.; Travassos da Rosa, A.P.A.; Guzman, H.; Carrera, J.P.; et al. Characterization of five unclassified orthobunyaviruses (Bunyaviridae) from Africa and the Americas. J. Gen. Virol. 2017, 98, 2258–2266. [Google Scholar] [CrossRef] [PubMed]
- Rio, D.C.; Ares, M.; Hannon, G.J.; Nilsen, T.W. Purification of RNA using TRIzol (TRI reagent). Cold Spring Harb. Protoc. 2010, 2010, pdb prot5439. [Google Scholar] [CrossRef]
- Lohse, M.; Bolger, A.M.; Nagel, A.; Fernie, A.R.; Lunn, J.E.; Stitt, M.; Usadel, B. RobiNA: A user-friendly, integrated software solution for RNA-Seq-based transcriptomics. Nucleic Acids Res. 2012, 40, W622–W627. [Google Scholar] [CrossRef] [PubMed]
- Simpson, J.T.; Wong, K.; Jackman, S.D.; Schein, J.E.; Jones, S.J.; Birol, I. ABySS: A parallel assembler for short read sequence data. Genome Res. 2009, 19, 1117–1123. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Robinson, J.T.; Thorvaldsdottir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative genomics viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Attwood, T.K.; Babbitt, P.C.; Bateman, A.; Bork, P.; Bridge, A.J.; Chang, H.Y.; Dosztanyi, Z.; El-Gebali, S.; Fraser, M.; et al. InterPro in 2017-beyond protein family and domain annotations. Nucleic Acids Res. 2017, 45, D190–D199. [Google Scholar] [CrossRef] [PubMed]
- Tsirigos, K.D.; Peters, C.; Shu, N.; Kall, L.; Elofsson, A. The TOPCONS web server for consensus prediction of membrane protein topology and signal peptides. Nucleic Acids Res. 2015, 43, W401–W407. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC bioinformatics 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed]
- Mihindukulasuriya, K.A.; Nguyen, N.L.; Wu, G.; Huang, H.V.; da Rosa, A.P.; Popov, V.L.; Tesh, R.B.; Wang, D. Nyamanini and midway viruses define a novel taxon of RNA viruses in the order Mononegavirales. J Virol. 2009, 83, 5109–5116. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.B.; Cui, L.; Fitch, A.; Popov, V.; Travassos da Rosa, A.P.; Vasilakis, N.; Tesh, R.B.; Ghedin, E. Whole genome analysis of Sierra Nevada virus, a novel mononegavirus in the family Nyamiviridae. Am. J. Trop. Med. Hyg. 2014, 91, 159–164. [Google Scholar] [CrossRef]
- Beaty, B.J.; Calisher, C.H.; Shope, R.E. Arboviruses. In Diagnostic procedures for viral, rickettsial and chlamydial infections; Schmidt, N.J., Emmons, R.W., Eds.; American Public Health Association: Washington, DC, USA, 1989; pp. 797–856. [Google Scholar]
- Amarasinghe, G.K.; Ayllon, M.A.; Bao, Y.; Basler, C.F.; Bavari, S.; Blasdell, K.R.; Briese, T.; Brown, P.A.; Bukreyev, A.; Balkema-Buschmann, A.; et al. Taxonomy of the order Mononegavirales: Update 2019. Arch. Virol. 2019, 164, 1967–1980. [Google Scholar] [CrossRef]
- Presti, R.M.; Zhao, G.; Beatty, W.L.; Mihindukulasuriya, K.A.; da Rosa, A.P.; Popov, V.L.; Tesh, R.B.; Virgin, H.W.; Wang, D. Quaranfil, Johnston Atoll, and Lake Chad viruses are novel members of the family Orthomyxoviridae. J. Virol. 2009, 83, 11599–11606. [Google Scholar] [CrossRef]
- Allison, A.B.; Ballard, J.R.; Tesh, R.B.; Brown, J.D.; Ruder, M.G.; Keel, M.K.; Munk, B.A.; Mickley, R.M.; Gibbs, S.E.; Travassos da Rosa, A.P.; et al. Cyclic avian mass mortality in the northeastern United States is associated with a novel orthomyxovirus. J. Virol. 2015, 89, 1389–1403. [Google Scholar] [CrossRef]
- L’Vov, D.; Al’khovskii, S.V.; Shchelkanov, M.; Shchetinin, A.M.; Deriabin, P.G.; Aristova, V.A.; Gitel’man, A.K.; Samokhvalov, E.I.; Botikov, A.G. [Taxonomic status of the Tyulek virus (TLKV) (Orthomyxoviridae, Quaranjavirus, Quaranfil group) isolated from the ticks Argas vulgaris Filippova, 1961 (Argasidae) from the birds burrow nest biotopes in the Kyrgyzstan]. Vopr. Virusol. 2014, 59, 28–32. [Google Scholar]
- Shi, M.; Lin, X.D.; Tian, J.H.; Chen, L.J.; Chen, X.; Li, C.X.; Qin, X.C.; Li, J.; Cao, J.P.; Eden, J.S.; et al. Redefining the invertebrate RNA virosphere. Nature 2016, 540, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Haenni, A.L.; de Miranda, J.R.; Falk, B.W.; Goldbach, R.; Mayo, M.A.; Shirako, Y.; Toriyama, S. Family Bunyaviridae. In Virus Taxonomy. Classification and Nomenclature of Viruses. Ninth report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier Academic Press: London, UK, 2012; pp. 694–723. [Google Scholar]
- Li, C.X.; Shi, M.; Tian, J.H.; Lin, X.D.; Kang, Y.J.; Chen, L.J.; Qin, X.C.; Xu, J.; Holmes, E.C.; Zhang, Y.Z. Unprecedented genomic diversity of RNA viruses in arthropods reveals the ancestry of negative-sense RNA viruses. Elife 2015, 4, e05378. [Google Scholar] [CrossRef] [PubMed]
- Abudurexiti, A.; Adkins, S.; Alioto, D.; Alkhovsky, S.V.; Avsic-Zupanc, T.; Ballinger, M.J.; Bente, D.A.; Beer, M.; Bergeron, E.; Blair, C.D.; et al. Taxonomy of the order Bunyavirales: Update 2019. Arch. Virol. 2019, 164, 1949–1965. [Google Scholar] [CrossRef] [PubMed]
- Marklewitz, M.; Zirkel, F.; Rwego, I.B.; Heidemann, H.; Trippner, P.; Kurth, A.; Kallies, R.; Briese, T.; Lipkin, W.I.; Drosten, C.; et al. Discovery of a unique novel clade of mosquito-associated bunyaviruses. J. Virol. 2013, 87, 12850–12865. [Google Scholar] [CrossRef] [PubMed]
- Alkhovsky, S.V.; Shchetinin, A.M.; Lvov, D.K.; Shchelkanov, M.Y.; Deryabin, P.G.; Lvov, D.N.; Samokhvalov, E.I.; Gitelman, A.K.; Botikov, A.G. Khurdun virus (KHURV): A new representative of the genus Orthobunyavirus (Bunyaviridae). Vopr. Virusol. 2013, 58, 10–13. [Google Scholar]
- Quan, P.L.; Deviatkin, A.A.; Markelov, M.; Bhuva, N.; Dedkov, V.G.; Zhuravlev, V.I.; Solovyov, A.; Briese, T.; Lipkin, W.I.; Shipulin, G.A. Akhtuba virus, a new member of the Bunyaviridae family isolated from pintail birds (Anas acuta) in Russia. Available online: https://www.ncbi.nlm.nih.gov/nuccore/KF601560.1 (accessed on 20 August 2019).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Field Number | Proposed Virus Name | Abbreviation | Avian Source, Date Collected | Proposed Classification (Genus; Family) |
|---|---|---|---|---|
| DO-200 | San Jacinto virus | SJCV | Sturnus vulgaris, 20 June 2013 | Nyavirus; Nyamiviridae |
| DO-159 | Buffalo Bayou virus | BBAV | Cyanocitta cristata*, 27 July 2011 | (uncertain); Peribunyaviridae |
| DO-159 | Mason Creek virus | MCRV | Cyanocitta cristata*, 27 July 2011 | Quaranjavirus; Orthomyxoviridae |
| Antigen | Antibody | ||||
|---|---|---|---|---|---|
| NYMV | MIDWV | SNVV | NDV | SJCV | |
| NYMV | 512/128* | 0 | 0 | 0 | 0 |
| MIDWV | 0 | 1024/≥512 | 0 | 0 | 0 |
| SNVV | 0 | 0 | 64/>4 | 0 | 0 |
| NDV | 0 | 0 | 0 | ≥64/≥4 | 0 |
| SJCV | 0 | 0 | 0 | 0 | 128/≥4 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walker, P.J.; Tesh, R.B.; Guzman, H.; Popov, V.L.; Travassos da Rosa, A.P.A.; Reyna, M.; Nunes, M.R.T.; de Souza, W.M.; Contreras-Gutierrez, M.A.; Patroca, S.; et al. Characterization of Three Novel Viruses from the Families Nyamiviridae, Orthomyxoviridae, and Peribunyaviridae, Isolated from Dead Birds Collected during West Nile Virus Surveillance in Harris County, Texas. Viruses 2019, 11, 927. https://doi.org/10.3390/v11100927
Walker PJ, Tesh RB, Guzman H, Popov VL, Travassos da Rosa APA, Reyna M, Nunes MRT, de Souza WM, Contreras-Gutierrez MA, Patroca S, et al. Characterization of Three Novel Viruses from the Families Nyamiviridae, Orthomyxoviridae, and Peribunyaviridae, Isolated from Dead Birds Collected during West Nile Virus Surveillance in Harris County, Texas. Viruses. 2019; 11(10):927. https://doi.org/10.3390/v11100927
Chicago/Turabian StyleWalker, Peter J., Robert B. Tesh, Hilda Guzman, Vsevolod L. Popov, Amelia P.A. Travassos da Rosa, Martin Reyna, Marcio R.T. Nunes, William Marciel de Souza, Maria A. Contreras-Gutierrez, Sandro Patroca, and et al. 2019. "Characterization of Three Novel Viruses from the Families Nyamiviridae, Orthomyxoviridae, and Peribunyaviridae, Isolated from Dead Birds Collected during West Nile Virus Surveillance in Harris County, Texas" Viruses 11, no. 10: 927. https://doi.org/10.3390/v11100927
APA StyleWalker, P. J., Tesh, R. B., Guzman, H., Popov, V. L., Travassos da Rosa, A. P. A., Reyna, M., Nunes, M. R. T., de Souza, W. M., Contreras-Gutierrez, M. A., Patroca, S., Vela, J., Salvato, V., Bueno, R., Widen, S. G., Wood, T. G., & Vasilakis, N. (2019). Characterization of Three Novel Viruses from the Families Nyamiviridae, Orthomyxoviridae, and Peribunyaviridae, Isolated from Dead Birds Collected during West Nile Virus Surveillance in Harris County, Texas. Viruses, 11(10), 927. https://doi.org/10.3390/v11100927