Abstract
This report describes and characterizes three novel RNA viruses isolated from dead birds collected during West Nile virus surveillance in Harris County, TX, USA (the Houston metropolitan area). The novel viruses are identified as members of the families Nyamaviridae, Orthomyxoviridae, and Peribunyaviridae and have been designated as San Jacinto virus, Mason Creek virus, and Buffalo Bayou virus, respectively. Their potential public health and/or veterinary importance are still unknown.
1. Introduction
In 2002, when West Nile virus (WNV) was first detected in Texas [1], the Harris County Public Health Mosquito and Vector Control Division (HCMVCD), in collaboration with the University of Texas Medical Branch (UTMB) in Galveston, initiated a surveillance program to monitor WNV activity in wild birds in Harris County and the City of Houston. The avian surveillance program has been described previously [1,2,3]. Basically, it consisted of netting, bleeding, and releasing live wild birds and testing their sera for antibodies to WNV and St. Louis encephalitis virus (SLEV). Dead birds collected in the city and county were also tested for the presence of WNV [1,2].
From 2002 to 2014, a total of 9047 dead birds were submitted to UTMB for culture; 1644 of the birds (18.17%) yielded WNV. Other known viruses recovered from the brain cultures of dead birds from Harris County during this 13-year period included Newcastle disease virus (NDV), eastern equine encephalitis virus (EEEV), Mermet virus (MERV), and Flanders virus (FLAV) [4,5]. In this report, we describe and characterize three additional novel viruses from the families Nyamaviridae, Orthomyxoviridae, and Peribunyaviridae that were also isolated from dead birds in Harris County during the study period.
2. Materials and Methods
2.1. Study Area
Harris County, which includes the city of Houston, is located in the northern coastal plain of the Gulf of Mexico, a 50-m swath along the Texas Gulf Coast. Harris County covers a geographic area of more than 1768 square miles. It is the largest county in Texas; its human population was estimated to be 4,538,000 in 2015. The climate is classified as humid subtropical. The average annual rainfall in Houston is 1145 mm, with an annual mean temperature of 20.5 °C. A more complete description of the study area was given previously [1].
2.2. Collection of Dead Birds
As part of the WNV surveillance program, HCMVCD staff have collected dead birds found in public places or at private residences in the county since 2002 [1,2]. Bird carcasses were identified, tagged with location and date, placed in double plastic bags and stored in ‒70 °C freezers for subsequent transport on dry ice to the World Reference Center for Emerging Viruses and Arboviruses (WRCEVA) in the Department of Pathology at UTMB for testing.
2.3. Culture Methods
After thawing, the skull was opened with sterile surgical instruments and a small piece of brain was removed from each bird in a biosafety level 3 (BSL-3) laboratory. Brain tissue was homogenized using a TissueLyser (Qiagen, Hilde, Germany) in tubes with 1.5–2.0 mL of phosphate-buffered saline, pH 7.4, containing 10% fetal bovine serum, 1% penicillin‒streptomycin‒amphotericin stock (Sigma, St. Louis, MO, USA) and several 3-mm stainless-steel balls. After centrifugation, the supernatant was passed through a 0.20-µm nylon syringe filter (Fisher Scientific, Pittsburgh, PA, USA) to remove bacteria and fungi. Then 150 µL of the filtrate was inoculated into separate 12.5-cm2 flask cultures of Vero E6 and Aedes albopictus C6/36 cells, originally obtained from the American Type Culture Collection (Manassas, VA, USA). After adsorption for 2 h at 28 °C (C6/36) or 37 °C (Vero), 5.0 mL of maintenance medium was added to each flask, and they were held in incubators at 28 °C and 37 °C, respectively. Cell cultures were examined regularly for evidence of viral cytopathic effect (CPE). Two brain cultures, designated TX-9285 (DO 200) and TX-8339 (DO 159), produced CPE in C6/36 cell and Vero cell cultures, respectively, and yielded the three viruses described in this report.
2.4. Evaluation of Pathogenicity in Suckling Mice
Two litters of two-day-old ICR mouse pups (n = 20) were inoculated intracranially with approximately 15 µL of fluid medium from each of the two cell cultures showing CPE. After inoculation, the pups were returned to their dams and examined daily for 14 days for signs of illness or death. The mice were purchased from Harlan Sprague Dawley (Indianapolis, IN, USA); animal work at the University of Texas Medical Branch was carried out under an Institutional Animal Care and Use Committee-approved protocol (no. 9505045; approval date: May 1, 2013)).
2.5. Transmission Electron Microscopy (TEM)
For ultrastructural analysis, infected cells were fixed for at least 1 h in a mixture of 2.5% formaldehyde prepared from paraformaldehyde powder and 0.1% glutaraldehyde in 0.05 M cacodylate buffer (pH 7.3), to which 0.03% picric acid and 0.03% CaCl2 were added. The monolayers were washed in 0.1 M cacodylate buffer, and cells were scraped off and processed further as a pellet. The pellets were post-fixed in 1% OsO4 in 0.1 M cacodylate buffer (pH 7.3) for 1 h, washed with distilled water, and stained in a block with 2% aqueous uranyl acetate for 20 min at 60 °C. The pellets were dehydrated in ethanol, processed through propylene oxide, and embedded in Poly/Bed 812 (Polysciences, Warrington, PA, USA). Ultrathin sections were cut on a Leica EM UC7 µL tramicrotome (Leica Microsystems, Buffalo Grove, IL, USA), stained with lead citrate, and examined with a Phillips 201 transmission electron microscope (FEI Phillips, Hillsboro, OR, USA) at 60 kV.
2.6. RNA Extraction, Viral Genome Sequencing, and Assembly
Viral RNAs were extracted using TRIzol LS reagent (Invitrogen, Carlsbad, CA) as described previously [6]. Viral RNA (~0.9 µg) was fragmented by incubation at 94 °C for 8 min in 19.5 µL of fragmentation buffer (Illumina 15016648). A sequencing library was prepared from the sample RNA using an Illumina TruSeq RNA v2 kit following the manufacturer’s protocol. The samples were sequenced on a HiSeq 1500 using the 2 x 50 paired-end protocol. Reads in fastq format were quality-filtered, and any adapter sequences were removed using Trimmomatic software [7]. The de novo assembly program ABySS [8] was used to assemble the reads into contigs, using several different sets of reads from 25,000 to 1 million read pairs, and kmer values from 20 to 40. The longest contigs were selected from the results, the reads were mapped back to the selected contigs using bowtie2 [9], and finally visualized with the Integrated Genomics Viewer [10] to verify that the assembled contigs were correct. Sequence annotation was determined using the Geneious version 9.1.2 (Biomatters, Auckland, New Zealand) [11]. Sequences were deposited in the GenBank database (http://www.ncbi.nlm.nih.gov/genbank/) under the following access numbers for the isolates SJCV (MK971153), MCRV [MK037473 (PB1), MK037472 (PB2), MK MK037474 (PA), MK037475 (NP), MK037476 (HA) and MN450148 (M)], and BBAV [MK037470 (L), MK037471 (S)]. Raw sequencing data are available upon request to the corresponding author.
2.7. Viral Genome Annotation
The size, organization, terminal regions, open reading frames (ORFs), encoded proteins, and their conserved motifs were annotated using applications available in the Geneious suite. Protein functions were verified using InterPro [12]. The identification of transmembrane regions and signal peptides was performed in the TOPCONS webserver [13] and the identification of glycosylation sites in the NetNglyc 1.0 Server (http://www.dbs.dtu.dk/services/NetNGlyc/). The genetic relatedness of the isolates SJCV and MCRV, as well as the recovered genomes of BBAV and other viruses were first inspected by the Blastx algorithm available from the NCBI (http://www.ncbi.nlm.nih.gov/genbank/).
2.8. Phylogenetic Analyses
Multiple amino acid sequence alignments were conducted using ClustalX [14] or in MUSCLE [15] in MEGA version 7.0. Maximum likelihood phylogenetic trees were inferred in MEGA using the GTR (general time reversible) nucleotide substitution model and a discrete gamma distribution to model evolutionary rate differences among sites rates of substitution among sites. All positions containing gaps were eliminated. Initial trees for the heuristic search were obtained automatically by applying the neighbor-joining and BioNJ algorithms to a matrix of pairwise distances estimates using the maximum composite likelihood (MCL) approach and then selecting the topology with the superior log likelihood value. The phylogenetic robustness of each node was determined using 100 bootstrap replicates and nearest-neighbor branch-swapping. Trees were annotated in MEGA and final mark-up of trees was conducted in Adobe Illustrator CC (Adobe Inc., San Jose, CA, USA).
2.9. Serological Tests
Suckling mouse brain, sucrose‒acetone-extracted antigens, and mouse hyperimmune ascitic fluids (HIAFs) were prepared for isolate DO 200, as well as for Nyamanini (NYMV), Midway (MIDWV), Sierra Nevada (SNVV), and Newcastle disease (NDV) viruses, as described previously [16,17]. The NDV strain used was a 2005 Harris County isolate from a Eurasian collared dove (GenBank EU477193) [4]. Complement fixation (CF) tests were performed by a microtiter technique [18], using two units of guinea pig complement and overnight incubation of the antigen and antibody at 4 °C. CF titers were recorded as the highest dilutions giving CF levels of 3+ or 4+ on a scale of 0 to 4+.
3. Results
3.1. Virus Isolation and Initial Characterization
Sample DO 200 was the brain homogenate from a dead European starling (Sturnus vulgaris) collected in Crosby, Harris County, TX on 20 June 2013 (Table 1). The sample produced CPE in monolayer cultures of Vero E6 cells within 5–6 days after inoculation, but no CPE was observed following the inoculation of C6/36 cell cultures. Culture fluid from the infected Vero cells produced illness in newborn mice 4–5 days after intracranial inoculation.

Table 1.
Summary of tree novel viruses isolated from brains of dead birds in Harris County, TX.
Sample DO 159 was from a partially decomposed carcass of a blue jay (Cyanocitta cristata) collected in Katy, Harris County on 27 July 2011 (Table 1). During dissection, unidentified fly larvae (maggots) were observed in the brain. The homogenized filtered sample produced CPE (large syncytia) in the C6/36 cell culture from six days post-inoculation. No CPE was observed in Vero cells, and inoculation of culture fluid from the infected C6/36 cells did not produce illness after intracranial inoculation of newborn mice.
3.2. Transmission Electron Microscopy
In ultrathin sections of C6/36 cells infected with sample DO 159, numerous highly pleomorphic virus particles (spherical, filamentous, and of irregular shape) were observed to be forming at the cell surface (Figure 1a). Spherical particles were 130‒150 nm and up to 270 nm in diameter (Figure 1b); filamentous particles were 65–85 nm in diameter and could be very long, reaching 1–1.3 µm (Figure 1c). Ultrathin sections of Vero E6 cells infected with sample DO 200 revealed large virus particles, either spherical, from 320 nm to 800 nm in diameter, or pleomorphic and elongated, with sizes ranging from 350 × 540 nm to 280 × 1120 nm and 3280 × 1275 nm (Figure 1d,e). Virus particles formed at the cell surface in large quantities.
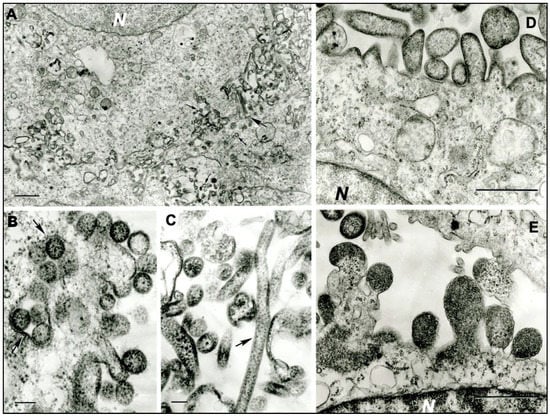
Figure 1.
Ultrastructure of MCRV (DO 159) in infected C6/36 cells, and SJCV (DO 200) in infected Vero E6 cells. (a) Multiple virus particles of MCRV forming at the cell surfaces of adjacent cells. N- cell nucleus. The large arrow shows two very long filamentous virions; small arrows show spherical virions. Bar = 1 µm. (b) Spherical virions of MCRV 130–150 nm in diameter at the cell surface. Arrows show cross-sections of the invaginations of the cell surface containing virions. Bar = 100 nm. (c) Virions of MCRV in extracellular space, a very long one (arrow) and pleomorphic. Bar = 100 nm. (d,e) Virions of SJCV forming at the surface of infected Vero E6 cells. N = fragments of cell nuclei. Bar = 1 µm.
3.3. Nucleotide Sequence Analysis
Illumina sequencing of brain sample DO 200 yielded a single viral contig of 13,295 nt representing the genome of a novel virus named San Jacinto virus (SJCV). Of 12.8 million total read pairs, 961,704 (~7.5%) mapped to the SJCV sequence. A BlastX search of the against NCBI nonredundant protein sequence (nr) database indicated highest homology with the genomes of Midway virus (MIDWV), Nyamanini virus (NYAV), and Sierra Nevada virus (SNVV), each of which is an unsegmented, negative sense (-) single-stranded (ss) RNA virus assigned to the genus Nyavirus, family Nyamiviridae, order Mononegavirales [16,17,19]. The SJCV genome sequence appeared to be complete, except for the extreme 3’ and 5’ terminal regions. The genome contains seven long open reading frames (ORFs), each bounded by relatively conserved consensus sequences corresponding to transcription initiation (3’-CGUUGG-5’) and transcription termination/polyadenylation (3’-AGAAAUCUUUUU-5’) signals (Figure 2a). ClustalX alignments indicated that six of the ORFs encode proteins corresponding to the six recognized nyavirus proteins: ORF1 encodes the nucleoprotein (N) of 402 amino acids (aa) (45 kDa); ORF2 encodes the negative regulator of the viral polymerase (X) of 187 aa (estimated 21.4 kDa); ORF3 encodes the polymerase cofactor (P) of 373 aa (41.7kDa); ORF4 encodes the matrix protein (M) of 192 aa (22.2 kDa); ORF5 encodes the class I transmembrane glycoprotein (G) of 678 aa (76.5 kDa, pre-modification), which contains four predicted glycosylation sites (triangles Figure 2a); and ORF7 encodes the full-length RNA-dependent RNA polymerase (L) of 1943 aa (218.5 kDa). ORF6, which does not occur in the other nyaviruses, encodes a unique protein (designated P6) of 337 aa (38.9 kDa), which displays obvious amino acid sequence identity with the N-terminal domain of the SJCV L protein (23.1%), as well as the L proteins of other nyaviruses (19.8–24.9%) (Figure S1). The extent of sequence conservation indicates that ORF6 has most likely arisen through a partial gene duplication event. The duplicated region precedes the six conserved regions (CRI‒CRVI) containing polymerase, polyribonucleotidyl transferase, and methyl transferase domains that are present in the L proteins of all unsegmented (-) ssRNA viruses and its function has not been clearly defined. Amino acid sequence identity (p-distance) comparisons with the L proteins of other nyamiviruses confirmed that SJCV shares a close relationship (62.4–82.4% identity) with MIDWV, NYAV, and SNVV (Table S1).
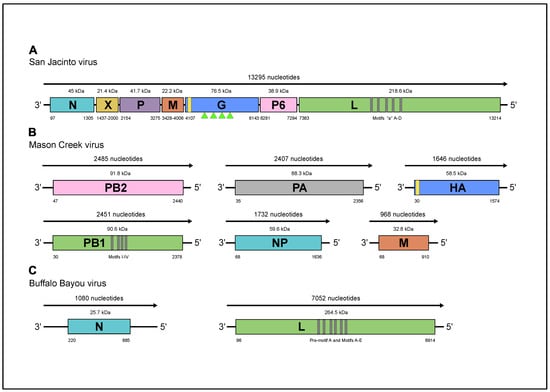
Figure 2.
Schematic representation of the genome organizations (a) SJCV; (b) MCV; and (c) BBAV. Green triangles indicate the location of predicted glycosylation sites.
Illumina sequencing of brain sample DO 159 revealed sequences that BlastX searches indicated represented the genomes of two distinct viruses. Of 12.5 million read pairs, 147,260 (~1.2%) mapped to the genome of a virus that was named Mason Creek virus (MCRV) and 291,205 (~2.3%) mapped to the genome of a virus that was named Buffalo Bayou virus (BBAV). The MCRV genome was represented as six contigs that ranged in size from 968 nt to 2485 nt (Figure 2b). Each contained complete coding sequences that aligned with highest homology to six (-) ssRNA segments (PA, PB1, PB2, NP, HA, and M) of Longchuan virus (LCHV), which was isolated from mosquitoes (Culex quinquefasciatus) collected in Guangdong Province, China (https://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi?id = 2594109), as well as Quaranfil virus (QRFV; genus Quaranjavirus, family Orthomyxoviridae, order Articulavirales) and several other currently unclassified viruses from ticks or birds that have been identified as possible quaranjaviruses [20,21]. The absence of conserved sequences with partial inverse complementary indicated that the 3’ and 5’ terminal noncoding regions were incomplete. The MCRV polymerase complex is formed by three segments: PA encoding a 773-aa protein (88.3 kDa); PB1 encoding a putative protein of 782 aa (90.6 kDa); and PB2 encoding a putative protein of 797 aa (91.8 kDa). The NP segment encodes the nucleoprotein of 523 aa (59.6 kDa). The HA segment encodes the hemagglutinin protein of 514 aa (58.5 kDa); the hemagglutinin is predicted to possess a cleavable signal peptide and three potential N-glycosylation sites. The M segment encodes the matrix protein of 281 aa (32.8 kDa). An exhaustive search of the available contigs failed to detect segment 7 of Wellfleet Bay virus (WFBV), which encodes a putative nonstructural protein VP7 [21], which we used as a surrogate sequence in our attempts to detect the seventh fragment of MRCV. Clustal X alignments of the deduced amino acid sequences of the PA, PB1, PB2, HA, and NP proteins indicated that MCRV was most closely related to LCHV, and more distantly related to QRFV, WFBV, Araguari virus (ARAV), Tjuloc virus (also named Tyulek virus; TLKV) [22], and Beihai orthomyxo-like virus 1 (BhOMV1) [23] (Tables S2‒S6).
The BBAV genome consisted of two (-) ssRNA segments of 1080 nucleotides (S RNA) and 7052 nucleotides (L RNA), each with very long terminal regions (5′ and 3′ NCR) of 219–886 nt and 96–138 nt, respectively (Figure 2c). The absence of conserved sequences with partial inverse complementary indicated that the 3’ and 5’ terminal noncoding regions were incomplete. BlastX searches indicated that the segments aligned with the genome segments of various members of the family Peribunyaviridae. The BBAV L segment encodes for the RNA-dependent RNA polymerase (RdRp) of 2272 amino acids with a predicted molecular weight of 264.5 kDa. The BBAV S segment encodes a nucleoprotein of 221 amino acids and predicted molecular weight of 25.6 kDa, but there was no evidence of an alternative open reading frame encoding a small nonstructural protein (NSs), as occurs in some members of the family Peribunyaviridae [24]. Exhaustive searches of available contigs also failed to detect sequences corresponding to the M segments of other peribunyaviruses that encode the two envelope glycoproteins (Gn and Gc). The BBAV RdRp shares a relatively low level of amino acid sequence identity (p-distance) with those of other peribunyaviruses (29.1–31.4%) (Table S7), but contained all recognized functional domains including pre-motif A and motifs A through E (aa positions between 970 to 1240). The BBAV nucleoprotein is similar in size but shares a relatively low sequence identity (16.8–23.6%) with the other peribunyaviruses (Table S8). Bunya-like viruses lacking an M segment have been reported previously in viral metagenomic studies of invertebrates [23,25]. However, the failure to detect this segment in BBAV and these other viruses may well be a consequence of the divergent nature of the sequences rather than their genuine absence in the viral genomes.
3.4. Complement Fixation Tests
To determine their immunological relationships, CF tests were performed using mouse brain antigens and mouse HIAFs prepared to the SJCV, NYMV, MIDWV, SNVV, and NDV viruses (Table 2). SJCV MIAF had a homologous antibody titer of 1:128, but did not cross-react with the other antigens. Likewise, MIAF to the other viruses did not cross-react with SJCV.
Table 2.
Complement fixation tests of San Jacinto virus (SJCV) with Nyamanini (NYMV), Midway (MIDWV), Sierra Nevada (SNVV), and Newcastle disease (NDV).
3.5. Phylogenetic Analyses
Phylogenetic trees inferred using amino acid sequences for SJCV, MCRV and BBAV grouped the viruses in the families Nyamiviridae, Orthomyxoviridae and Bunyaviridae, respectively. Phylogenetic analysis of the RdRp (L protein) of SJCV and other members of the order Mononegavirales indicated that SJCV clusters with members of the genus Nyavirus, family Nyamiviridae (BSP = 100%) (Figure 3). The genus Nyavirus currently consists of three members: Midway virus (MIDWV; species Midway nyavirus), Nyamanini virus (NYMV; species Nyamanini nyavirus), and Sierra Nevada virus (SNVV; species Sierra Nevada nyavirus) [19]. The sequence data suggest that SJCV should be assigned to a fourth species in the genus Nyavirus. All four of these viruses have been associated with birds and/or ticks [16,17].
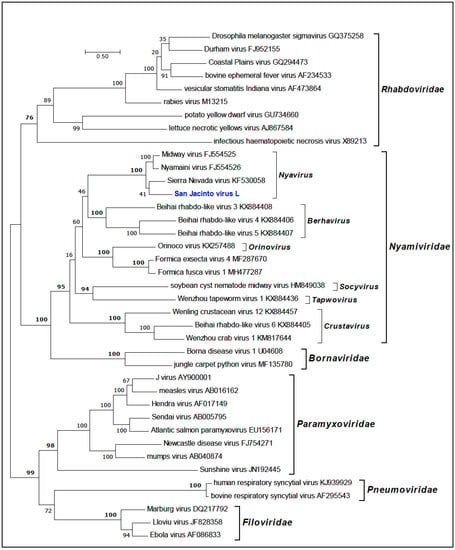
Figure 3.
Maximum likelihood phylogenetic tree of the RNA-dependent RNA polymerase (L protein) amino acid sequences for representative members of the order Mononegavirales with emphasis on the family Nyamiviridae. Branch lengths are proportional to the number of nucleotide substitutions per site. Values for bootstrap support proportion (BSP) were determined from 100 bootstrap iterations. BSP values showing strong support for each of the key nodes are shown in bold. The position in the tree of SJCV is shown in blue. The Genbank accession numbers of all sequences are as shown.
Phylogenetic analysis of the amino sequences of the MCRV RdRp (PB1, PB2, and PA) and nucleoprotein (NP) indicated that it clusters with members of the family Orthomyxoviridae (Figure 4a–d). The Orthomyxoviridae currently comprises seven genera (Quaranjavirus, Thogotovirus, Alphainfluenzavirus, Betainfluenzavirus, Gammainfluenzavirus, Deltainfluenzavirus, and Isfavirus) (https://talk.ictvonline.org/files/master-species-lists/m/msl/8266). Although the members of three of these genera infect birds, MCRV did not fall within any of the existing genera but clustered with high support (BSP >90%) with LCHV (from mosquitoes), as well as Beihai orthomyxo-like virus 1 (from woodlice), Jiujie fly virus (from horseflies), and members (assigned and proposed) of the genus Quaranjavirus [23,25]. The ecological diversity of these viruses suggests that MCRV should be assigned to a new species and genus within the Orthomyxoviridae.
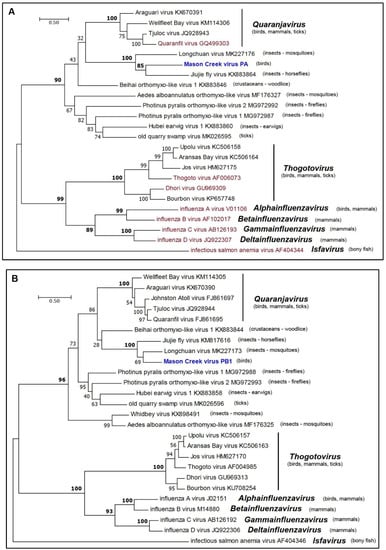
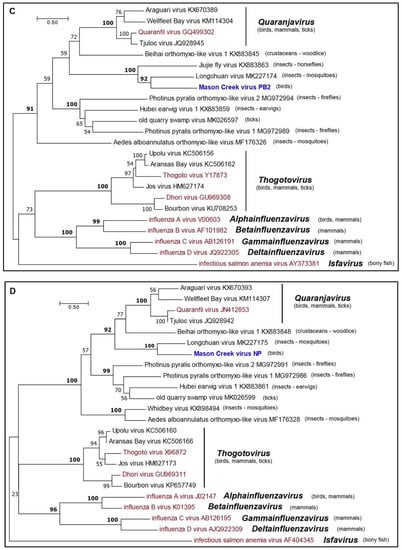
Figure 4.
Maximum likelihood phylogenetic trees for amino acid sequences of representative members of the family Orthomyxoviridae. (a) PA proteins; (b) PB1 proteins; (c) PB2 proteins; and (d) NP proteins. Branch lengths are proportional to the number of nucleotide substitutions per site. Values for bootstrap support proportion (BSP) were determined from 100 bootstrap iterations. BSP values showing strong support for each of the key nodes are shown in bold. In each tree, viruses formally assigned to species are shown in magenta and the position of MCV is shown in blue. The Genbank accession numbers of all sequences are as shown.
Phylogenetic analysis of the amino acid sequences of the BBAV RdRp (L protein) indicated that it is a member of the family Peribunyaviridae [26]. BBAV lies within a well-supported clade that also includes members of the genus Herbevirus (Herbert virus [HEBV; species Herbert herbevirus], Kibale virus [KIBV; species Kibale herbevirus], Tai virus [TAIV; species Tai herbevirus]) [27], Shuangao insect virus 1 (SgIV-1; species Insect shangavirus) [25], and two currently unassigned viruses (Khurdun virus [KHUV] and Akhtuba virus [AKHV]) [28,29] (Figure 5). HEBV, KIBV, and TAIV were each isolated from mosquitoes in Africa and are thought to be insect-specific viruses as they replicate in insect cells but not in vertebrate cells [27]. SgIV-1 was detected by next-generation sequencing of a mixed insect pool containing lacewings (Chrysopidae) and moth flies (Psychoda) in China [23]. KHUV and AKHV were each isolated from birds in Russia, but do not cluster directly with BBAV. The data indicate that BBAV cannot be assigned at this time to any existing genus within the Peribunyaviridae [26] and so appears to represent a unique new species. Moreover, more genetic data are needed to accurately classify BBAV. Lastly, both MCRV and BBAV were isolated from the decomposing brain of a dead bird, which contained fly larvae (maggots). Based on their phenotypic characteristics (growth only in mosquito cells) and their phylogenetic relationship, they would appear to be insect-specific viruses, probably infecting the fly larvae and not the bird.
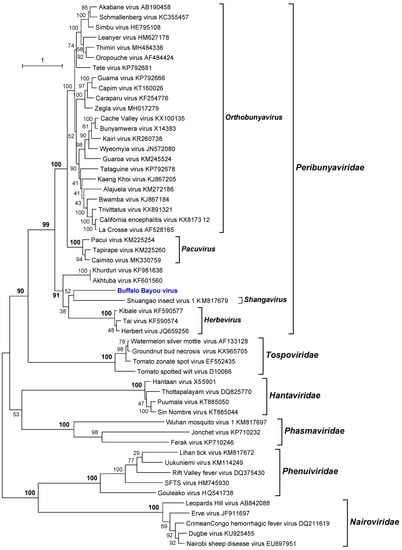
Figure 5.
Maximum likelihood phylogenetic tree of the RNA-dependent RNA polymerase (L protein) amino acid sequences for representative members of the order Bunyavirales with emphasis on the family Peribunyaviridae. Branch lengths are proportional to the number of nucleotide substitutions per site. Values for bootstrap support proportion (BSP) were determined from 100 bootstrap iterations. BSP values showing strong support for each of the key nodes are shown in bold. The position in the tree of BBAV is shown in blue. The Genbank accession numbers of all sequences are as shown.
4. Discussion
The potential public health or veterinary importance of SJCV, MCRV, and BBAV is unknown, as are their prevalence and host range in Harris County. These are subjects for future investigation. Likewise, the pathogenesis of the three viruses should be examined in several avian and possibly mammalian species.
The discovery of the three novel viruses during our WNV surveillance program also reinforces the value of virus culture in arbovirus surveillance programs. Many public health and veterinary diagnostic laboratories now simply screen animal carcasses or pools of hematophagous insects by PCR or other nonculture techniques for the detection of RNA of viruses of interest (i.e., West Nile virus, Saint Louis encephalitis virus, eastern equine encephalitis virus, Venezuelan encephalitis virus, epizootic hemorrhagic disease virus, vesicular stomatitis virus, etc.). While these techniques are very sensitive, provide faster results, are more economical, and reduce the risk of accidental infection of laboratory personnel, they generally detect only the viruses for which one has appropriate primers. These techniques usually do not detect novel or unexpected viruses.
For discovering novel viruses, metagenomic analysis is now the current gold standard, as it allows the detection and genetic characterization of all associated viruses in a sample. However, metagenomic analysis is not a feasible technique for diagnostic laboratories that test thousands of field samples annually because of the per-sample cost, the labor involved in sample preparation, and the sophisticated equipment required. Culture is not ideal, since its sensitivity is dependent on the culture system(s) used and the growth characteristics of the virus(es) present in a sample. Nonetheless, primary culture still allows for the detection and isolation of some novel or unexpected viruses that would otherwise remain unknown.
Supplementary Materials
The following are available online at https://www.mdpi.com/1999-4915/11/10/927/s1, Figure S1. ClustalX amino acid sequence alignment of the SJCV P6 protein and the N-terminal regions of the L proteins of SJCV and other members of the genus Nyavirus. Amino acids conserved between nyavirus L proteins and the SJCV P6 protein are shaded. Table S1. Amino acid sequence homology (p-distances) of the L proteins of SNVV and selected nyamiviruses. Table S2. Amino acid sequence homology (p-distances) of the PA proteins of MCRV and selected orthomyxoviruses. Table S3. Amino acid sequence homology (p-distances) of the PB1 proteins of MCRV and selected orthomyxoviruses. Table S4. Amino acid sequence homology (p-distance) of the PB2 proteins of BBAV and selected orthomyxoviruses. Table S5. Amino acid sequence homology (p-distances) of the NP proteins of BBAV and selected orthomyxoviruses. Table S6. Amino acid sequence homology (p-distances) of the HA proteins of MCRV and selected orthomyxoviruses. Table S7. Amino acid sequence homology (p-distances) of the L proteins of MCRV and selected peribunyaviruses. Table S8. Amino acid sequence homology (p-distances) of the N proteins of MCRV and selected peribunyaviruses.
Author Contributions
Conceptualization, R.B.T., H.G. and N.V.; Data curation, R.B.T., M.R.T.N., S.W., P.J.W. and N.V.; Formal analysis, R.B.T., V.L.P., A.P.A.T.d.R., M.R.T.N., S.W., P.J.W. and N.V.; Funding acquisition, R.B.T.; Investigation, R.B.T., H.G., V.L.P., A.P.A.T.d.R., M.A.C.-G., M.R.T.N., W.M.d.S., M.R., S.P., S.W., T.G.W., J.V., V.S., R.B., P.J.W. and N.V.; Methodology, R.B.T., P.J.W., M.R. and N.V.; Project administration, R.B.T., H.G. and N.V.; Resources, R.B.T., H.G. and N.V.; Supervision, R.B.T. and N.V.; Writing—review & editing, R.B.T., M.R.T.N., V.L.P., S.G.W., P.J.W. and N.V.
Funding
This research was funded by grant R24AI120942 and contracts N01-AI30027, HHS-N2722010000401, and HHSN2720004/D04 from the U.S. National Institutes of Health, and a CNPq grant 302032/2011-8 from the Brazilian National Council for Research and Development. MAC was supported by a Ph.D. scholarship from the Colombian Department of Science (Colciencias Convocatoria 567).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Lillibridge, K.M.; Parsons, R.; Randle, Y.; Travassos da Rosa, A.P.; Guzman, H.; Siirin, M.; Wuithiranyagool, T.; Hailey, C.; Higgs, S.; Bala, A.A.; et al. The 2002 introduction of West Nile virus into Harris County, Texas, an area historically endemic for St. Louis encephalitis. Am. J. Trop. Med. Hyg. 2004, 70, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Wilkerson, L.; Reyna Nava, M.; Battle-Freeman, C.; Travassos da Rosa, A.; Guzman, H.; Tesh, R.; Debboun, M. The role of birds in arboviral disease surveillance in Harris County and the City of Houston, Texas. US Army Med. Dep. J. 2017, (1-17), 1–12. [Google Scholar]
- Mann, B.R.; McMullen, A.R.; Swetnam, D.M.; Salvato, V.; Reyna, M.; Guzman, H.; Bueno, R.; Dennett, J.A.; Tesh, R.B.; Barrett, A.D. Continued evolution of West Nile virus, Houston, Texas, USA, 2002-2012. Emerg. Infect. Dis. 2013, 19, 1418–1427. [Google Scholar] [CrossRef] [PubMed]
- Kim, L.M.; King, D.J.; Guzman, H.; Tesh, R.B.; Travassos da Rosa, A.P.; Bueno, R.; Dennett, J.A.; Afonso, C.L. Biological and phylogenetic characterization of pigeon paramyxovirus serotype 1 circulating in wild North American pigeons and doves. J. Clin. Microbiol. 2008, 46, 3303–3310. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.B.; Gulino, K.M.; Tesh, R.B.; Cui, L.; Fitch, A.; Unnasch, T.R.; Popov, V.L.; Travassos da Rosa, A.P.A.; Guzman, H.; Carrera, J.P.; et al. Characterization of five unclassified orthobunyaviruses (Bunyaviridae) from Africa and the Americas. J. Gen. Virol. 2017, 98, 2258–2266. [Google Scholar] [CrossRef] [PubMed]
- Rio, D.C.; Ares, M.; Hannon, G.J.; Nilsen, T.W. Purification of RNA using TRIzol (TRI reagent). Cold Spring Harb. Protoc. 2010, 2010, pdb prot5439. [Google Scholar] [CrossRef]
- Lohse, M.; Bolger, A.M.; Nagel, A.; Fernie, A.R.; Lunn, J.E.; Stitt, M.; Usadel, B. RobiNA: A user-friendly, integrated software solution for RNA-Seq-based transcriptomics. Nucleic Acids Res. 2012, 40, W622–W627. [Google Scholar] [CrossRef] [PubMed]
- Simpson, J.T.; Wong, K.; Jackman, S.D.; Schein, J.E.; Jones, S.J.; Birol, I. ABySS: A parallel assembler for short read sequence data. Genome Res. 2009, 19, 1117–1123. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Robinson, J.T.; Thorvaldsdottir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative genomics viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Attwood, T.K.; Babbitt, P.C.; Bateman, A.; Bork, P.; Bridge, A.J.; Chang, H.Y.; Dosztanyi, Z.; El-Gebali, S.; Fraser, M.; et al. InterPro in 2017-beyond protein family and domain annotations. Nucleic Acids Res. 2017, 45, D190–D199. [Google Scholar] [CrossRef] [PubMed]
- Tsirigos, K.D.; Peters, C.; Shu, N.; Kall, L.; Elofsson, A. The TOPCONS web server for consensus prediction of membrane protein topology and signal peptides. Nucleic Acids Res. 2015, 43, W401–W407. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC bioinformatics 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed]
- Mihindukulasuriya, K.A.; Nguyen, N.L.; Wu, G.; Huang, H.V.; da Rosa, A.P.; Popov, V.L.; Tesh, R.B.; Wang, D. Nyamanini and midway viruses define a novel taxon of RNA viruses in the order Mononegavirales. J Virol. 2009, 83, 5109–5116. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.B.; Cui, L.; Fitch, A.; Popov, V.; Travassos da Rosa, A.P.; Vasilakis, N.; Tesh, R.B.; Ghedin, E. Whole genome analysis of Sierra Nevada virus, a novel mononegavirus in the family Nyamiviridae. Am. J. Trop. Med. Hyg. 2014, 91, 159–164. [Google Scholar] [CrossRef]
- Beaty, B.J.; Calisher, C.H.; Shope, R.E. Arboviruses. In Diagnostic procedures for viral, rickettsial and chlamydial infections; Schmidt, N.J., Emmons, R.W., Eds.; American Public Health Association: Washington, DC, USA, 1989; pp. 797–856. [Google Scholar]
- Amarasinghe, G.K.; Ayllon, M.A.; Bao, Y.; Basler, C.F.; Bavari, S.; Blasdell, K.R.; Briese, T.; Brown, P.A.; Bukreyev, A.; Balkema-Buschmann, A.; et al. Taxonomy of the order Mononegavirales: Update 2019. Arch. Virol. 2019, 164, 1967–1980. [Google Scholar] [CrossRef]
- Presti, R.M.; Zhao, G.; Beatty, W.L.; Mihindukulasuriya, K.A.; da Rosa, A.P.; Popov, V.L.; Tesh, R.B.; Virgin, H.W.; Wang, D. Quaranfil, Johnston Atoll, and Lake Chad viruses are novel members of the family Orthomyxoviridae. J. Virol. 2009, 83, 11599–11606. [Google Scholar] [CrossRef]
- Allison, A.B.; Ballard, J.R.; Tesh, R.B.; Brown, J.D.; Ruder, M.G.; Keel, M.K.; Munk, B.A.; Mickley, R.M.; Gibbs, S.E.; Travassos da Rosa, A.P.; et al. Cyclic avian mass mortality in the northeastern United States is associated with a novel orthomyxovirus. J. Virol. 2015, 89, 1389–1403. [Google Scholar] [CrossRef]
- L’Vov, D.; Al’khovskii, S.V.; Shchelkanov, M.; Shchetinin, A.M.; Deriabin, P.G.; Aristova, V.A.; Gitel’man, A.K.; Samokhvalov, E.I.; Botikov, A.G. [Taxonomic status of the Tyulek virus (TLKV) (Orthomyxoviridae, Quaranjavirus, Quaranfil group) isolated from the ticks Argas vulgaris Filippova, 1961 (Argasidae) from the birds burrow nest biotopes in the Kyrgyzstan]. Vopr. Virusol. 2014, 59, 28–32. [Google Scholar]
- Shi, M.; Lin, X.D.; Tian, J.H.; Chen, L.J.; Chen, X.; Li, C.X.; Qin, X.C.; Li, J.; Cao, J.P.; Eden, J.S.; et al. Redefining the invertebrate RNA virosphere. Nature 2016, 540, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Haenni, A.L.; de Miranda, J.R.; Falk, B.W.; Goldbach, R.; Mayo, M.A.; Shirako, Y.; Toriyama, S. Family Bunyaviridae. In Virus Taxonomy. Classification and Nomenclature of Viruses. Ninth report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier Academic Press: London, UK, 2012; pp. 694–723. [Google Scholar]
- Li, C.X.; Shi, M.; Tian, J.H.; Lin, X.D.; Kang, Y.J.; Chen, L.J.; Qin, X.C.; Xu, J.; Holmes, E.C.; Zhang, Y.Z. Unprecedented genomic diversity of RNA viruses in arthropods reveals the ancestry of negative-sense RNA viruses. Elife 2015, 4, e05378. [Google Scholar] [CrossRef] [PubMed]
- Abudurexiti, A.; Adkins, S.; Alioto, D.; Alkhovsky, S.V.; Avsic-Zupanc, T.; Ballinger, M.J.; Bente, D.A.; Beer, M.; Bergeron, E.; Blair, C.D.; et al. Taxonomy of the order Bunyavirales: Update 2019. Arch. Virol. 2019, 164, 1949–1965. [Google Scholar] [CrossRef] [PubMed]
- Marklewitz, M.; Zirkel, F.; Rwego, I.B.; Heidemann, H.; Trippner, P.; Kurth, A.; Kallies, R.; Briese, T.; Lipkin, W.I.; Drosten, C.; et al. Discovery of a unique novel clade of mosquito-associated bunyaviruses. J. Virol. 2013, 87, 12850–12865. [Google Scholar] [CrossRef] [PubMed]
- Alkhovsky, S.V.; Shchetinin, A.M.; Lvov, D.K.; Shchelkanov, M.Y.; Deryabin, P.G.; Lvov, D.N.; Samokhvalov, E.I.; Gitelman, A.K.; Botikov, A.G. Khurdun virus (KHURV): A new representative of the genus Orthobunyavirus (Bunyaviridae). Vopr. Virusol. 2013, 58, 10–13. [Google Scholar]
- Quan, P.L.; Deviatkin, A.A.; Markelov, M.; Bhuva, N.; Dedkov, V.G.; Zhuravlev, V.I.; Solovyov, A.; Briese, T.; Lipkin, W.I.; Shipulin, G.A. Akhtuba virus, a new member of the Bunyaviridae family isolated from pintail birds (Anas acuta) in Russia. Available online: https://www.ncbi.nlm.nih.gov/nuccore/KF601560.1 (accessed on 20 August 2019).
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).