Ten-Year Responses of Underplanted Northern Red Oak to Silvicultural Treatments, Herbivore Exclusion, and Fertilization


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Descriptions
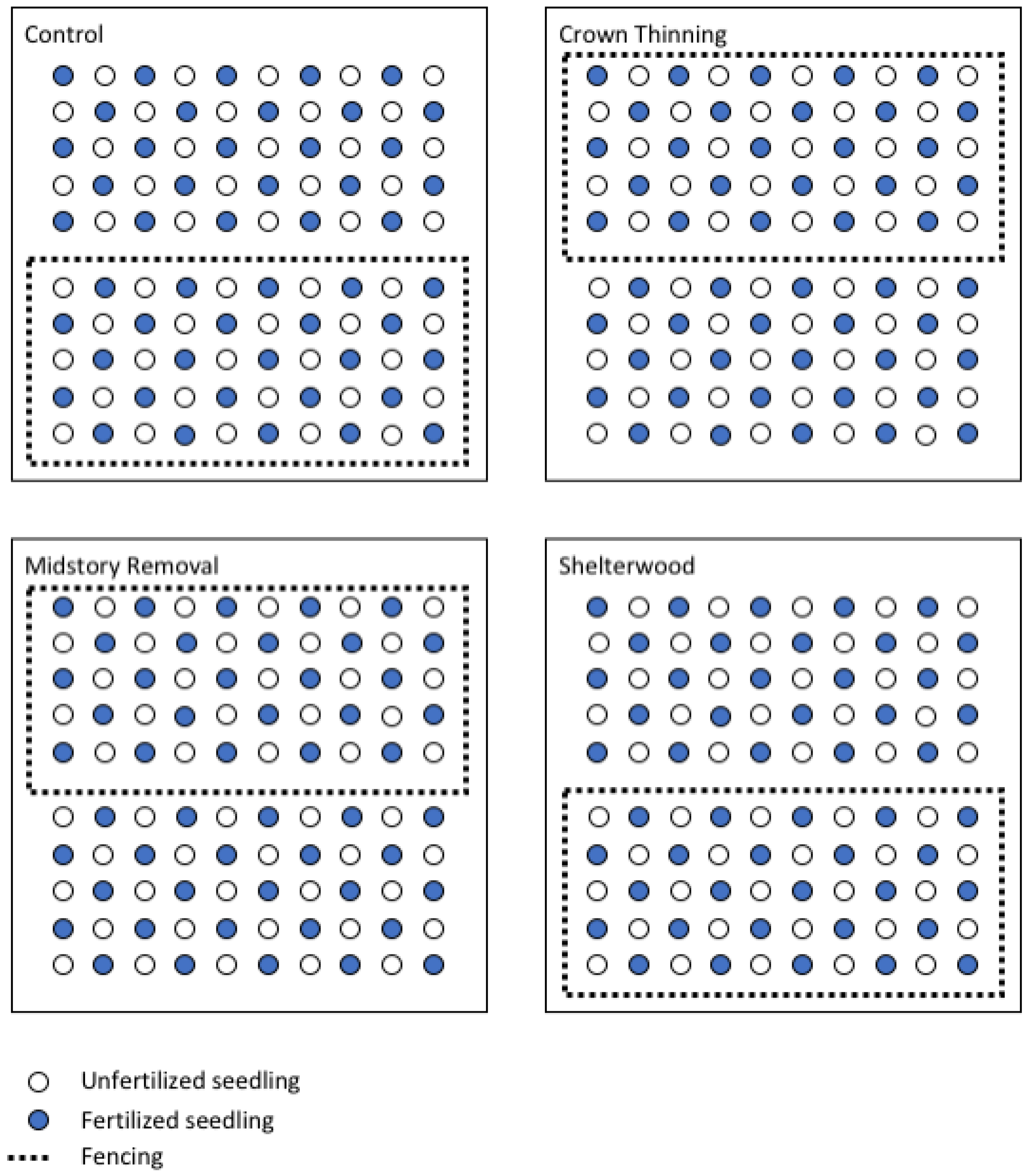
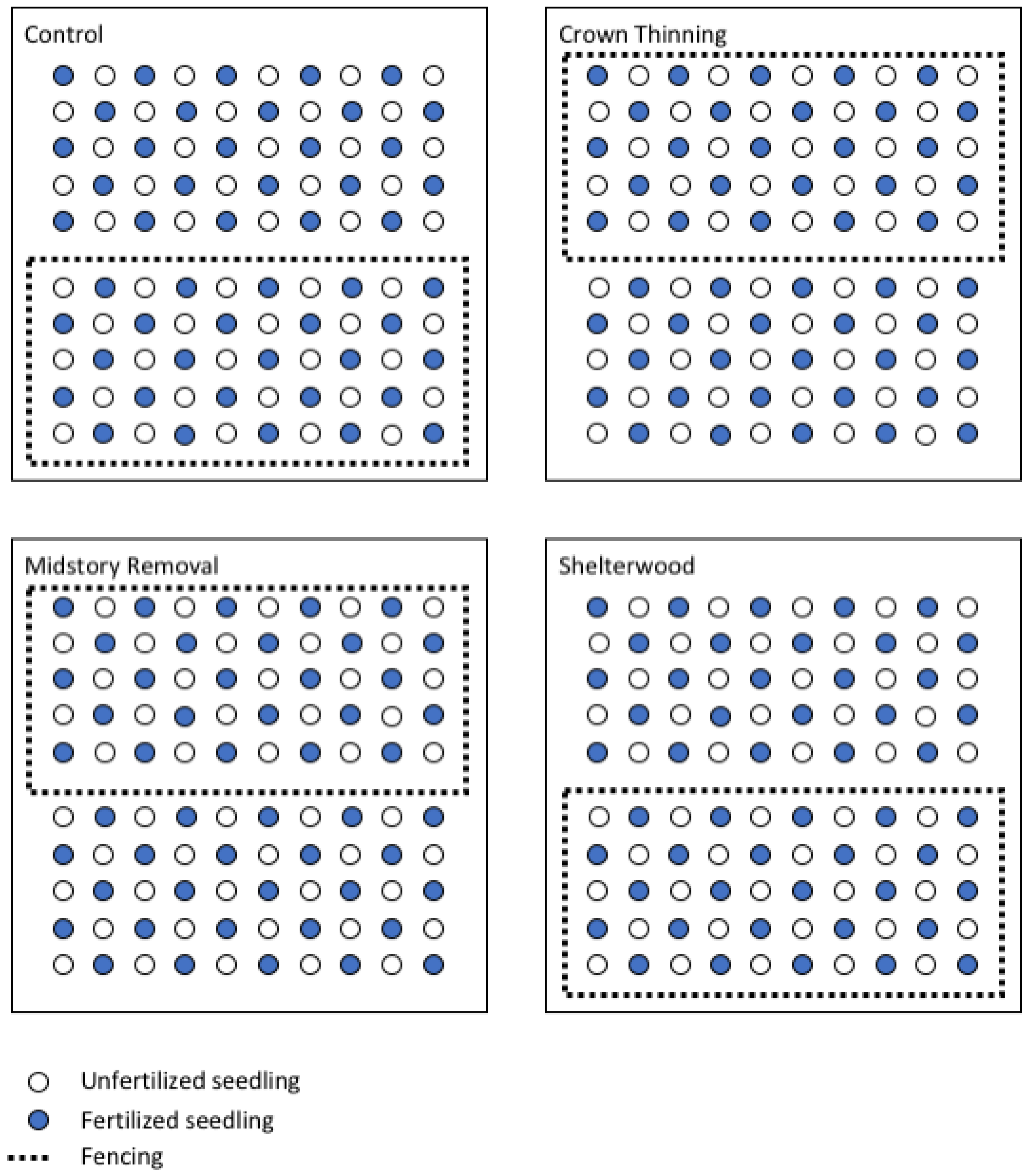
2.2. Experimental Design
2.3. Data Collection
2.4. Statistical Analysis
3. Results
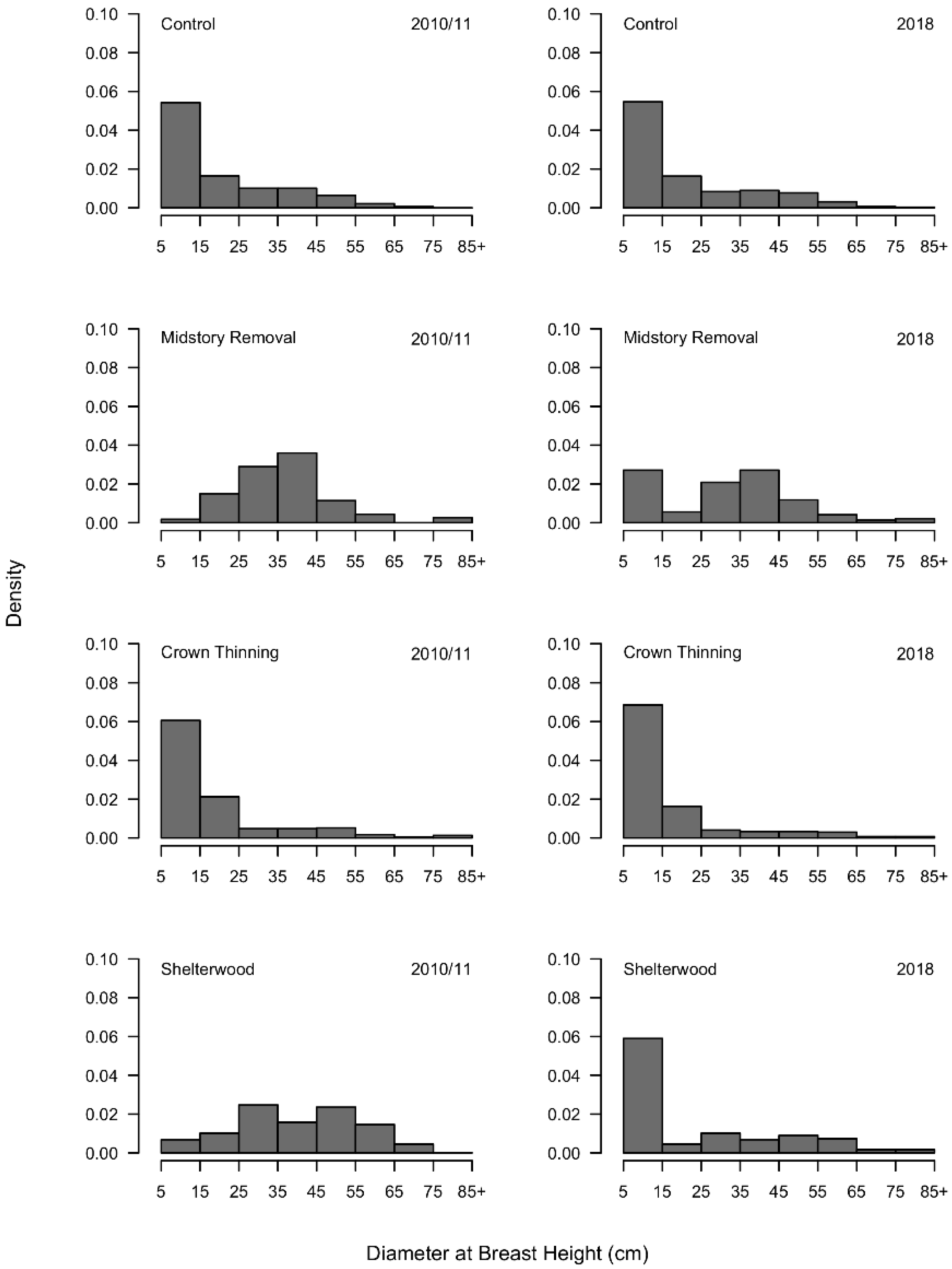
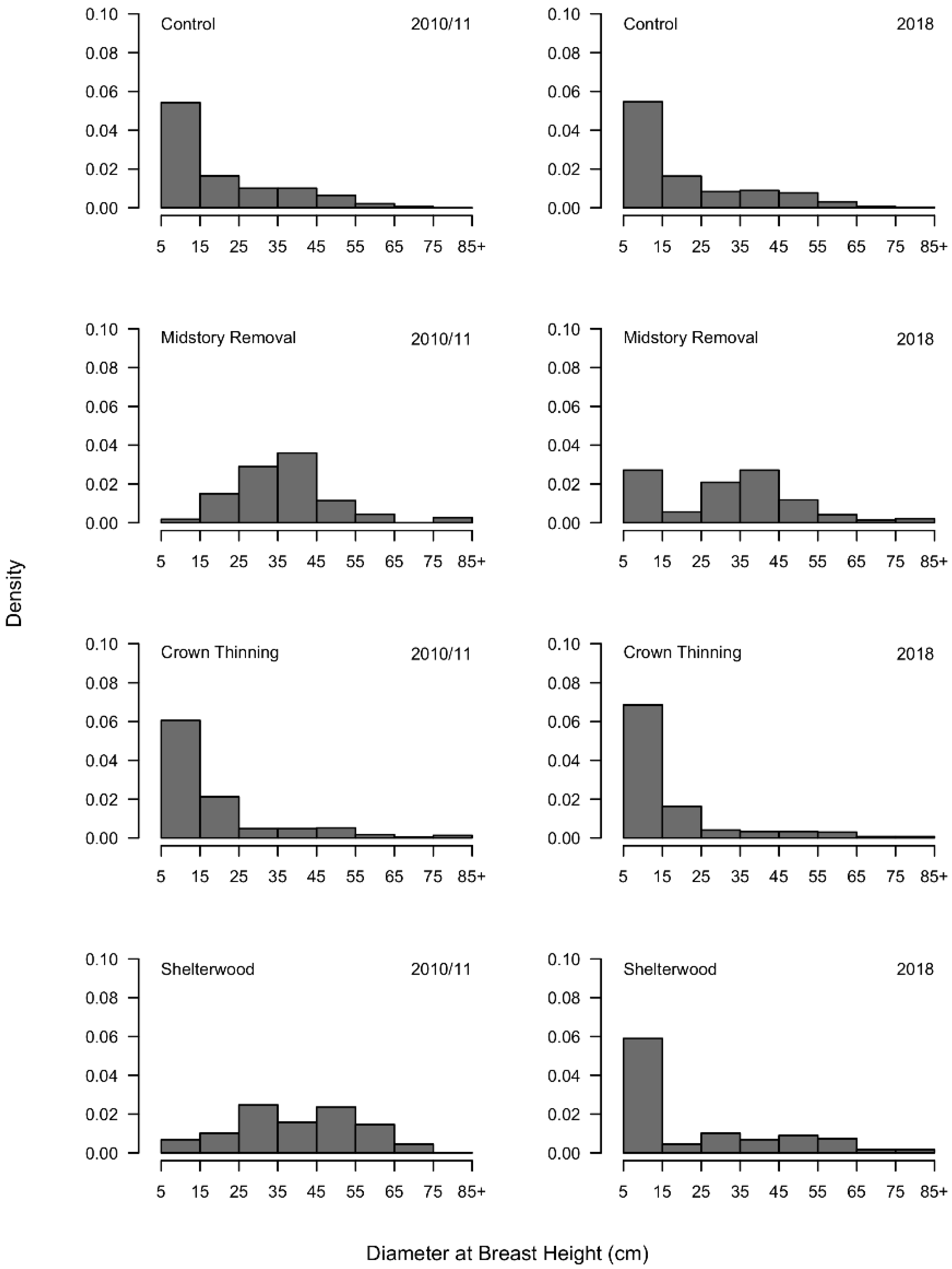
3.1. Overstory Conditions
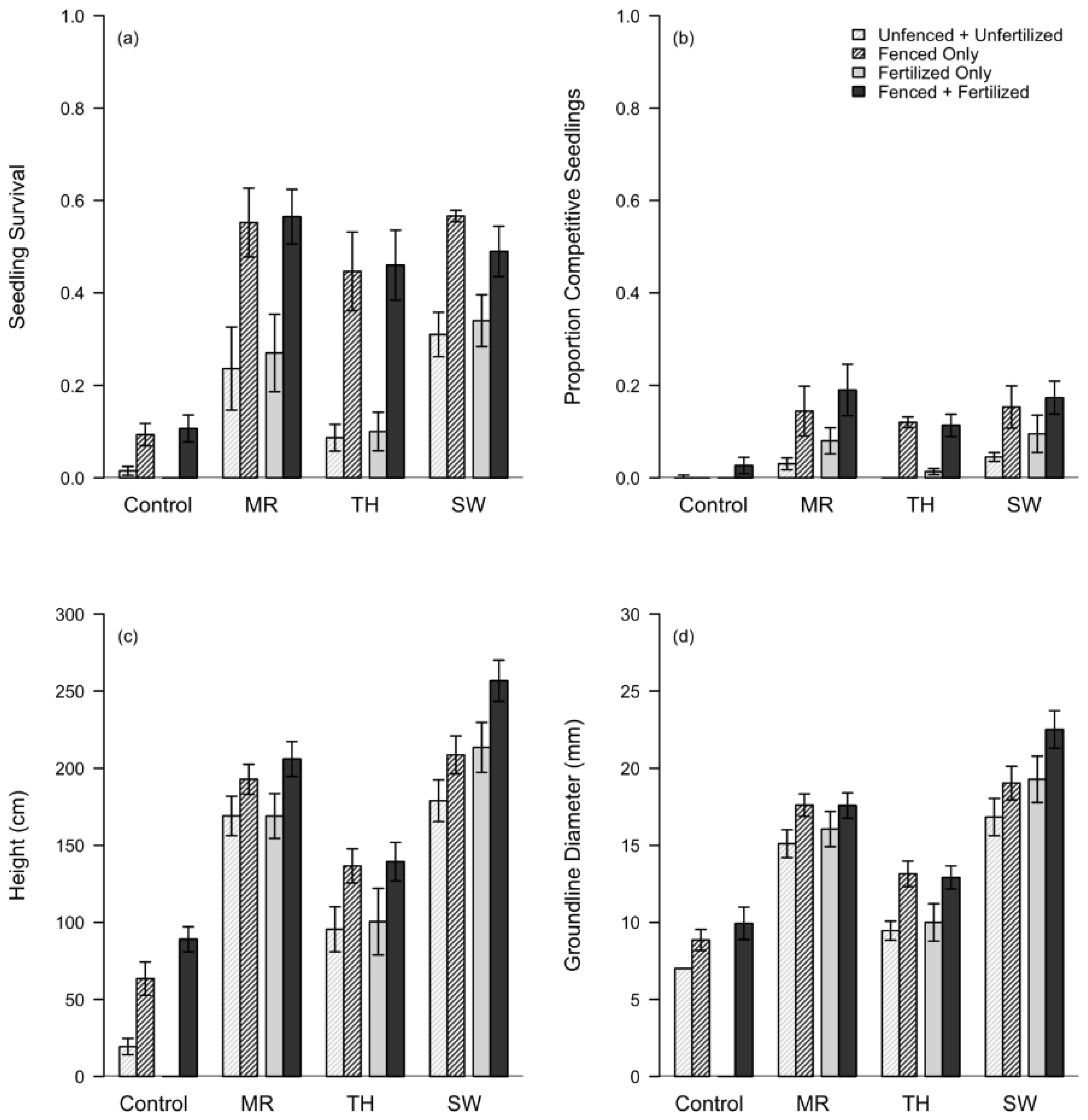
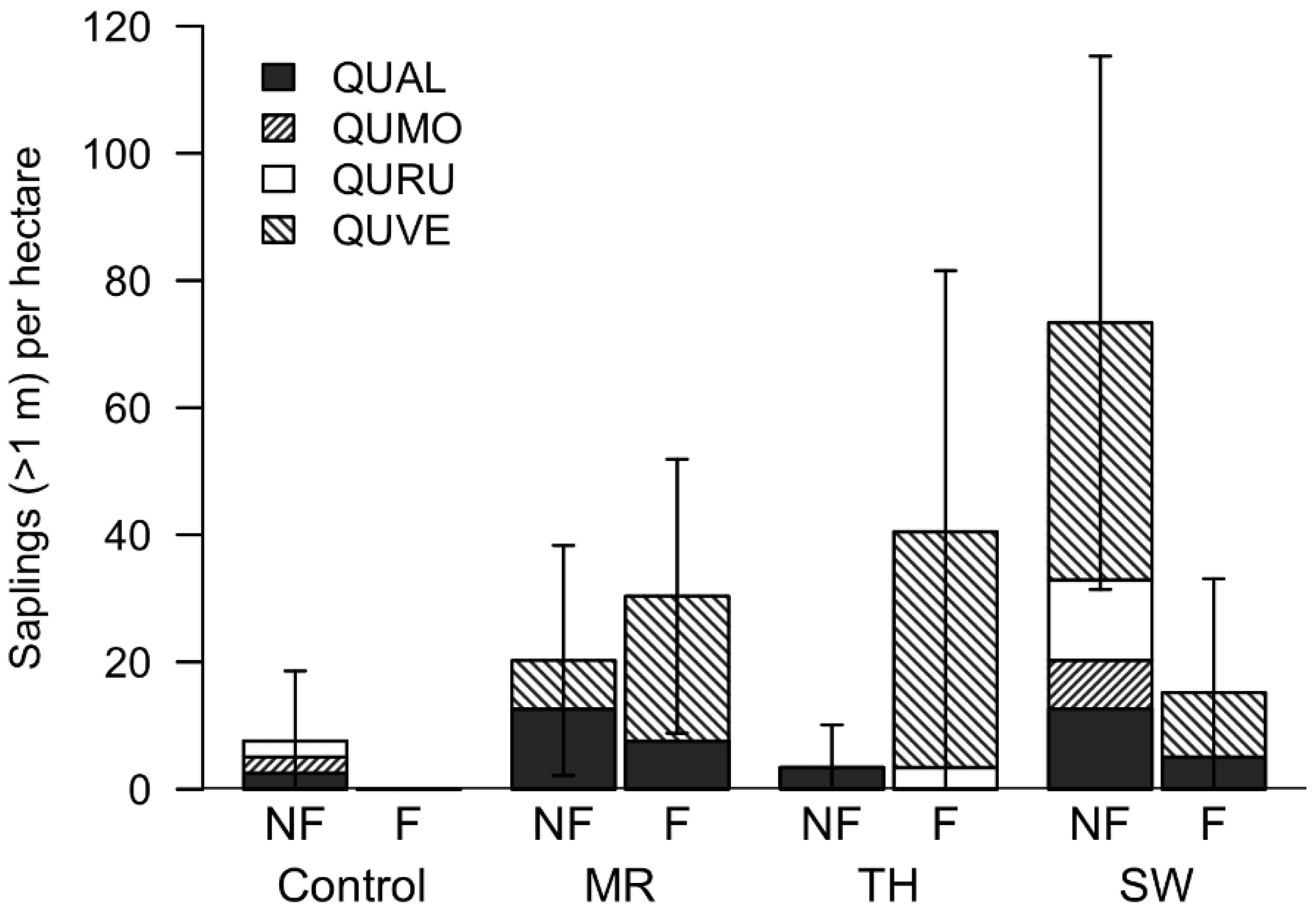
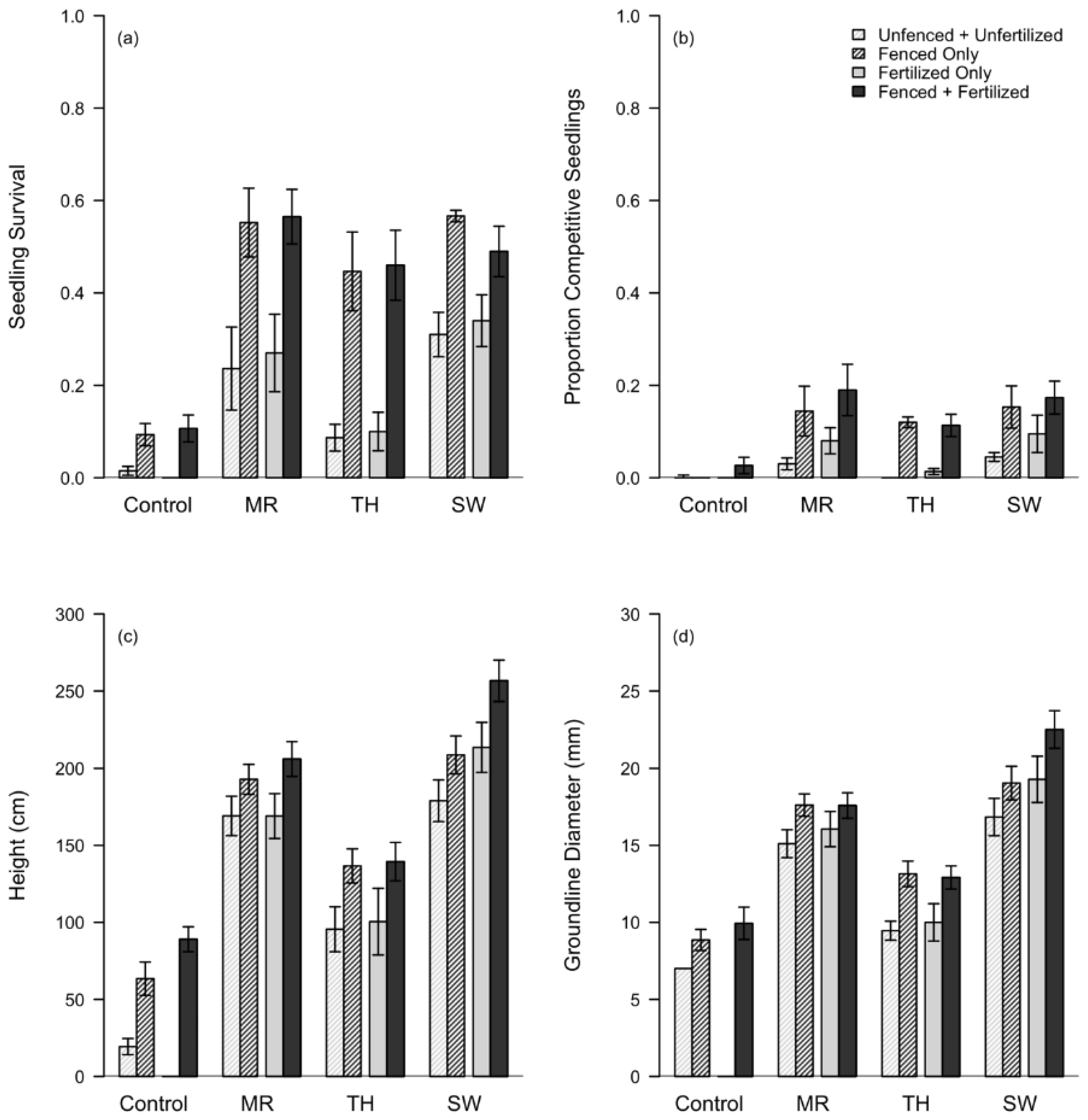
3.2. Artificial Regeneration
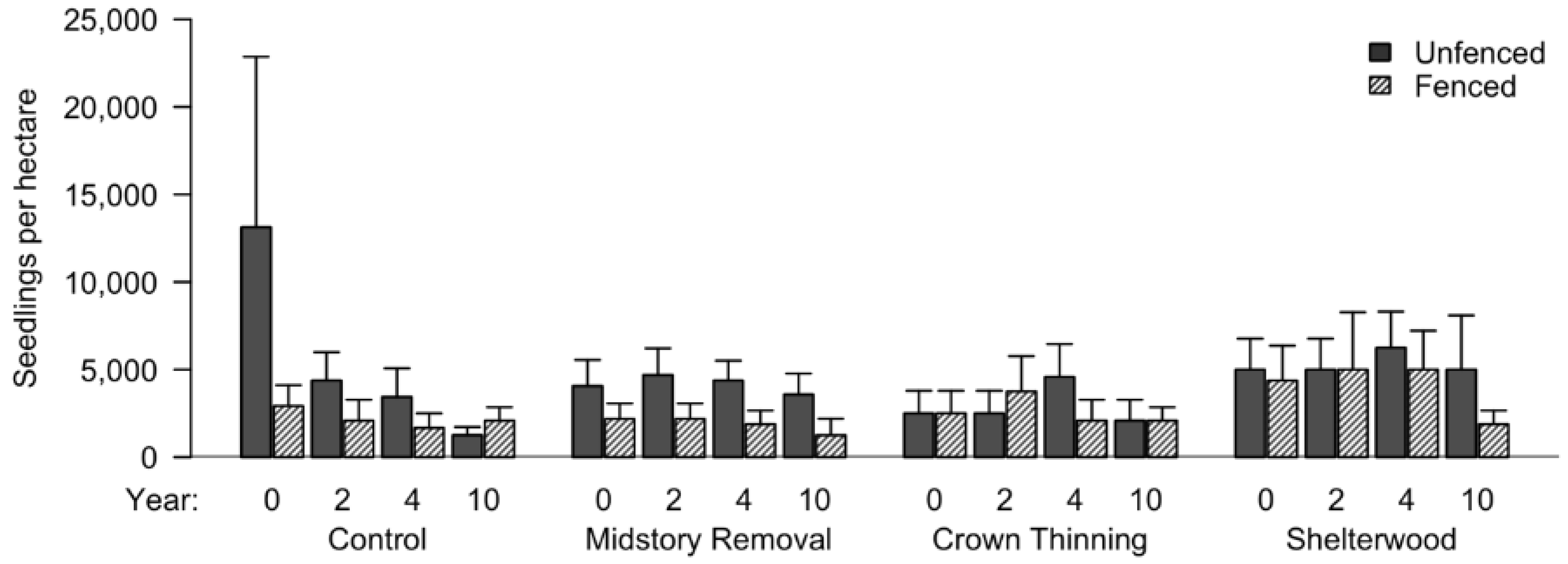
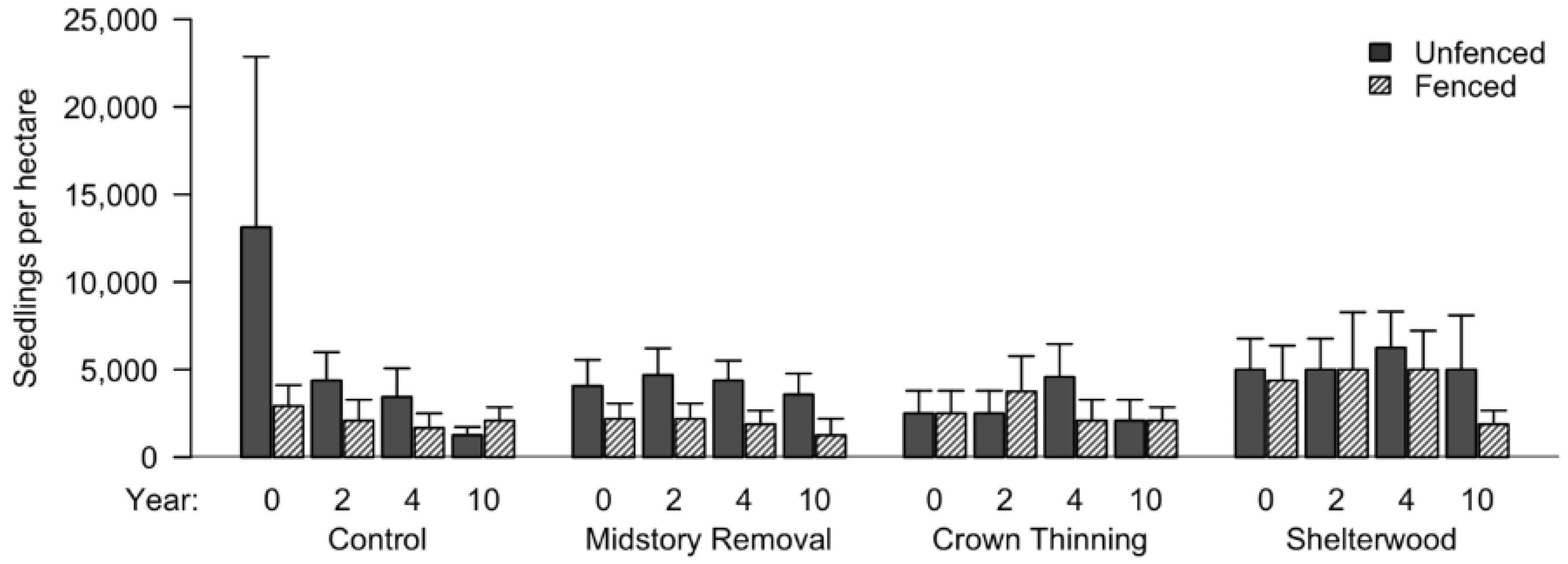
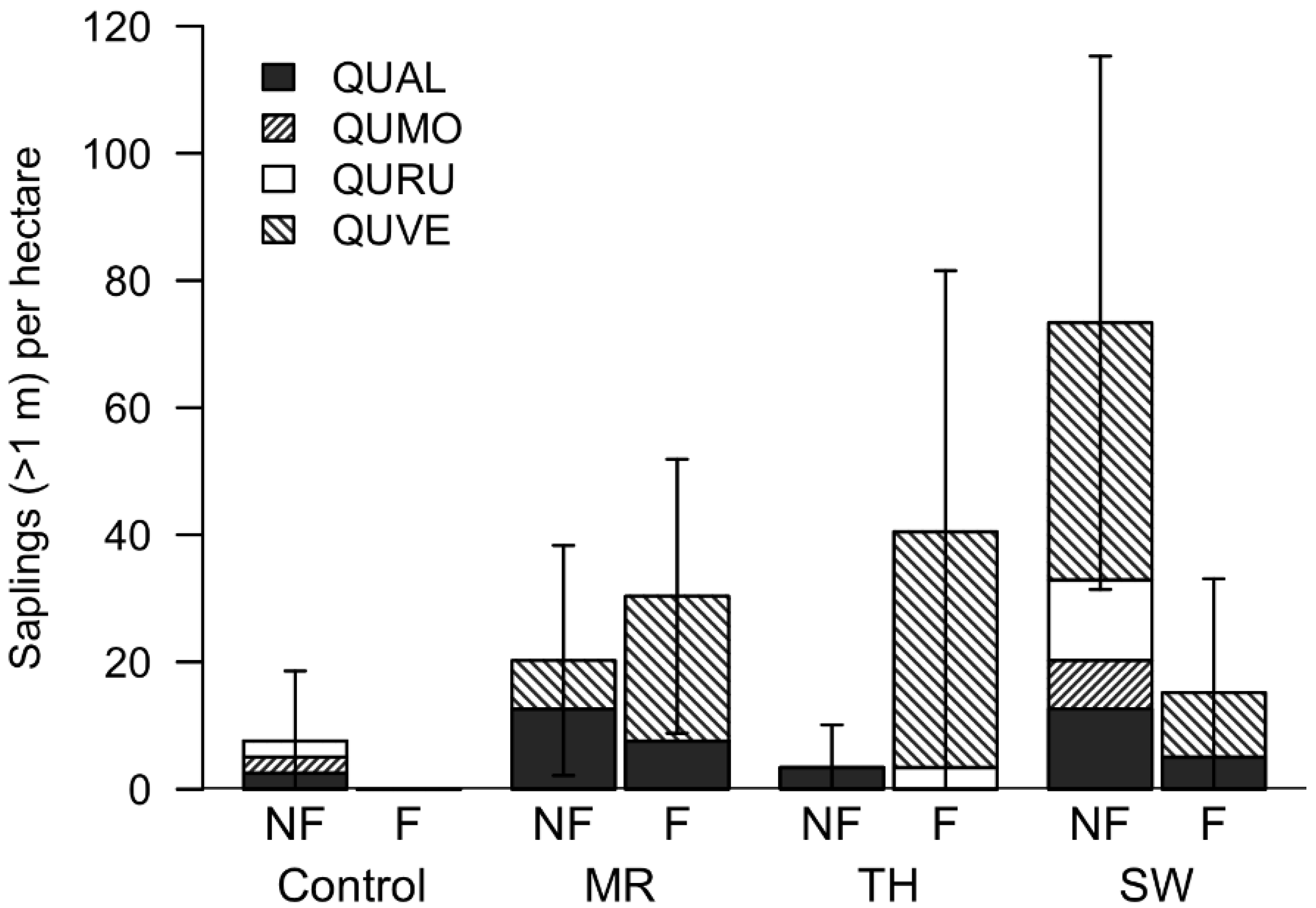
3.3. Natural Regeneration
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Hanberry, B.B.; Nowacki, G.J. Oaks were the historical foundation genus of the east-central United States. Quat. Sci. Rev. 2016, 145, 94–103. [Google Scholar] [CrossRef]
- Clark, F.B. An historical perspective of oak regeneration. In Oak Regeneration: Serious Problems Practical Recommendations; Loftis, D.L., McGee, C.E., Eds.; USDA Forest Service Gen. Tech. Rep. SE-84; U.S. Department of Agriculture, Forest Service, Southeastern Forest Experiment Station: Asheville, NC, USA, 1993. [Google Scholar]
- Lashley, M.A.; Harper, C.A.; Bates, G.E.; Keyser, P.D. Forage availability for white-tailed deer following silvicultural treatments in hardwood forests. J. Wildl. Manag. 2011, 75, 1467–1476. [Google Scholar] [CrossRef]
- Lorimer, C.G. Causes of the oak regeneration problem. In Oak Regeneration: Serious Problems, Practical Recommendations; Loftis, D.L., McGee, C.E., Eds.; USDA Forest Service Gen. Tech. Rep. SE-84; U.S. Department of Agriculture, Forest Service, Southeastern Forest Experiment Station: Asheville, NC, USA, 1993; pp. 14–39. [Google Scholar]
- Russell, F.L.; Zippin, D.B.; Fowler, N.L. Effects of White-Tailed Deer (Odocoileus Virginianus) on Plants, Plant Populations and Communities: A Review. Am. Midl. Nat. 2001, 146, 1–26. [Google Scholar] [CrossRef]
- Fei, S.; Kong, N.; Steiner, K.C.; Moser, W.K.; Steiner, E.B. Change in oak abundance in the eastern United States from 1980 to 2008. For. Ecol. Manag. 2011, 262, 1370–1377. [Google Scholar] [CrossRef]
- Nowacki, G.J.; Abrams, M.D. The Demise of Fire and “Mesophication” of Forests in the Eastern United States. Bioscience 2008, 58, 123–138. [Google Scholar] [CrossRef] [Green Version]
- Lhotka, J.M.; Loewenstein, E.F. Influence of canopy structure on the survival and growth of underplanted seedlings. New For. 2008, 35, 89–104. [Google Scholar] [CrossRef]
- Jenkins, M.A.; Parker, G.R. Composition and diversity of woody vegetation in silvicultural openings of southern Indiana forests. For. Ecol. Manag. 1998, 109, 57–74. [Google Scholar] [CrossRef]
- Keyser, T.L.; Loftis, D.L. Long-term effects of single-tree selection cutting on structure and composition in upland mixed-hardwood forests of the southern Appalachian Mountains. Forestry 2013, 86, 255–265. [Google Scholar] [CrossRef]
- Raymond, P.; Munson, A.D.; Ruel, J.C.; Nolet, P. Group and single-tree selection cutting in mixed tolerant hardwood-white pine stands: Early establishment dynamics of white pine and associated species. For. Chron. 2003, 79, 1093–1106. [Google Scholar] [CrossRef]
- Morrissey, R.C.; Jacobs, D.F.; Seifert, J.R.; Kershaw, J.A. Overstory species composition of naturally regenerated clearcuts in an ecological classification framework. Plant Ecol. 2010, 208, 21–34. [Google Scholar] [CrossRef]
- Schweitzer, C.J.; Dey, D.C. Forest structure, composition, and tree diversity response to a gradient of regeneration harvests in the mid-Cumberland Plateau escarpment region, USA. For. Ecol. Manag. 2011, 262, 1729–1741. [Google Scholar] [CrossRef]
- Loftis, D.L. A shelterwood method for regenerating red oak in the Southern Appalachians. For. Sci. 1990, 36, 917–929. [Google Scholar]
- Kellner, K.F.; Swihart, R.K. Timber harvest and drought interact to impact oak seedling growth and survival in the Central Hardwood Forest. Ecosphere 2016, 7, 1–15. [Google Scholar] [CrossRef]
- Brose, P.H.; Van Lear, D.H. Responses of hardwood advance regeneration to seasonal prescribed fires in oak-dominated shelterwood stands. Can. J. For. Res. 1998, 28, 331–339. [Google Scholar] [CrossRef]
- Brose, P.H.; Gottschalk, K.W.; Horsley, S.B.; Knopp, P.D.; Kochenderfer, J.N.; McGuiness, B.J.; Miller, G.W.; Ristau, T.E.; Stoleson, S.H.; Stout, S.L. Prescribing Regeneration Treatments for Mixed-Oak Forests in the Mid-Atlantic Region; USDA Forest Service Gen. Tech. Rep. NRS-33; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2008.
- Gordon, A.M.; Simpson, J.A.; Williams, P.A. Six-year response of red oak seedlings planted under a shelterwood in central Ontario. Can. J. For. Res. 1995, 25, 603–613. [Google Scholar] [CrossRef]
- Dey, D.C.; Gardiner, E.S.; Schweitzer, C.J.; Kabrick, J.M.; Jacobs, D.F. Underplanting to sustain future stocking of oak (Quercus) in temperate deciduous forests. New For. 2012, 43, 955–978. [Google Scholar] [CrossRef]
- Dey, D.C.; Spetich, M.A.; Weigel, D.R.; Johnson, P.S.; Graney, D.L.; Kabrick, J.M. A suggested approach for design of oak (Quercus L.) regeneration research considering regional differences. New For. 2009, 37, 123–135. [Google Scholar] [CrossRef]
- Sander, I.L.; Graney, D.L. Regenerating oaks in the central states. In Oak Regeneration: Serious Problems, Practical Recommendations.; Loftis, D.L., McGee, C.E., Eds.; USDA Forest Service Gen. Tech. Rep. SE-84; U.S. Department of Agriculture, Forest Service, Southeastern Forest Experiment Station: Asheville, NC, USA, 1993. [Google Scholar]
- Jarnemo, A.; Minderman, J.; Bunnefeld, N.; Zidar, J.; Mansson, J. Managing landscapes for multiple objectives: Alternative forage can reduce the conflict between deer and forestry. Ecosphere 2014, 5. [Google Scholar] [CrossRef]
- Murray, B.D.; Webster, C.R.; Jenkins, M.A.; Saunders, M.R.; Haulton, G.S. Ungulate impacts on herbaceous-layer plant communities in even-aged and uneven-aged managed forests. Ecosphere 2016, 7, 1–20. [Google Scholar] [CrossRef]
- Miller, B.F.; Campbell, T.A.; Laseter, B.R.; Ford, W.M.; Miller, K.V. White-tailed deer herbivory and timber harvesting rates: Implications for regeneration success. For. Ecol. Manag. 2009, 258, 1067–1072. [Google Scholar] [CrossRef] [Green Version]
- Royo, A.A.; Kramer, D.W.; Miller, K.V.; Nibbelink, N.P.; Stout, S.L. Spatio-temporal variation in foodscapes modifies deer browsing impact on vegetation. Landsc. Ecol. 2017, 32, 2281–2295. [Google Scholar] [CrossRef]
- Schreffler, A.M.; Sharpe, W.E. Effects of lime, fertilizer, and herbicide on forest soil and soil solution chemistry, hardwood regeneration, and hardwood growth following shelterwood harvest. For. Ecol. Manag. 2003, 177, 471–484. [Google Scholar] [CrossRef]
- Tripler, C.E.; Canham, C.D.; Inouye, R.S.; Schnurr, J.L. Soil nitrogen availability, plant luxury consumption, and herbivory by white-tailed deer. Oecologia 2002, 133, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Burney, O.T.; Jacobs, D.F. Species selection—A fundamental silvicultural tool to promote forest regeneration under high animal browsing pressure. For. Ecol. Manag. 2018, 408, 67–74. [Google Scholar] [CrossRef]
- Cha, D.H.; Appel, H.M.; Frost, C.J.; Schultz, J.C.; Steiner, K.C. Red oak responses to nitrogen addition depend on herbivory type, tree family, and site. For. Ecol. Manag. 2010, 259, 1930–1937. [Google Scholar] [CrossRef]
- Kellner, K.F.; Riegel, J.K.; Swihart, R.K. Effects of silvicultural disturbance on acorn infestation and removal. New For. 2014, 45, 265–281. [Google Scholar] [CrossRef]
- Zaczek, J.J.; Lhotka, J.M. Seedling reproduction established with soil scarification within an oak overwood after overstory removal. North J. Appl. For. 2004, 21, 5–11. [Google Scholar]
- Spetich, M.A.; Dey, D.C.; Johnson, P.S. Shelterwood-planted northern red oaks: Integrated costs and options. J. Appl. For. 2009, 33, 182–186. [Google Scholar]
- Homoya, M.A.; Abrell, D.B.; Aldrich, J.R.; Post, T.W. The natural regions of Indiana. Proc. Indiana Acad. Sci. 1984, 94, 245–268. [Google Scholar] [CrossRef]
- Parrott, D.L.; Lhotka, J.M.; Stringer, J.W.; Dillaway, D.N. Seven-Year Effects of Midstory Removal on Natural and Underplanted Oak Reproduction. North J. Appl. For. 2012, 29, 182–190. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 15 March 2018).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biometr. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed]
- Knoot, T.G.; Schulte, L.A.; Rickenbach, M. Oak conservation and restoration on private forestlands: Negotiating a social-ecological landscape. Environ. Manag. 2010, 45, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Sander, I.L.; Johnson, P.S.; Rogers, R. Evaluating Oak Advance Reproduction in the Missouri Ozarks; USDA Forest Service Gen. Tech Rep. NC-251; U.S. Department of Agriculture, Forest Service, North Central Forest Experiment Station: St. Paul, MN, USA, 1984; 16p.
- Healy, W.M.; Lewis, A.M.; Boose, E.F. Variation of red oak acorn production. For. Ecol. Manag. 1999, 116, 1–11. [Google Scholar] [CrossRef]
- Lhotka, J.M.; Loewenstein, E.F. Effect of midstory removal on understory light availability and the 2-year response of underplanted cherrybark oak seedlings. South J. Appl. For. 2009, 33, 171–177. [Google Scholar]
- Craig, J.M.; Lhotka, J.M.; Stringer, J.W. Evaluating Initial Responses of Natural and Underplanted Oak Reproduction and a Shade-Tolerant Competitor to Midstory Removal. For. Sci. 2014, 60, 1164–1171. [Google Scholar] [CrossRef]
- Miller, G.W.; Brose, P.H.; Gottschalk, K.W. Advanced Oak Seedling Development as Influenced by Shelterwood Treatments, Competition Control, Deer Fencing, and Prescribed Fire. J. For. 2017, 115, 179–189. [Google Scholar] [CrossRef]
- Stange, E.E.; Shea, K.L. Effects of Deer Browsing, Fabric Mats, and Tree Shelters on Quercus rubra Seedlings. Restor. Ecol. 1998, 6, 29–34. [Google Scholar] [CrossRef]
- Jenkins, L.H.; Jenkins, M.A.; Webster, C.R.; Zollner, P.A.; Shields, J.M. Herbaceous layer response to 17 years of controlled deer hunting in forested natural areas. Biol. Conserv. 2014, 175, 119–128. [Google Scholar] [CrossRef]
- Jenkins, L.H.; Murray, B.D.; Jenkins, M.A.; Webster, C.R. Woody regeneration response to over a decade of deer population reductions in Indiana state parks 1. J. Torrey Bot. Soc. 2015, 142, 205–219. [Google Scholar] [CrossRef]
- Wakeland, B.; Swihart, R.K. Ratings of white-tailed deer preferences for woody browse in Indiana. Proc. Indiana Acad. Sci. 2009, 118, 96–101. [Google Scholar]
- Kellner, K.F.; Swihart, R.K. Simulation of oak early life history and interactions with disturbance via an individual-based model, SOEL. PLoS ONE 2017, 12, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.S.; Hale, P.E.; Ford, W.M.; Wentworth, J.M.; French, J.R.; Anderson, O.F.; Pullen, G.B. White-tailed deer foraging in relation to successional stage, overstory type and management of southern Appalachian forests. Am. Midl. Nat. 1995, 133, 18–35. [Google Scholar] [CrossRef]
- Burney, O.T.; Jacobs, D.F. Ungulate herbivory of boreal and temperate forest regeneration in relation to seedling mineral nutrition and secondary metabolites. New For. 2013, 44, 753–768. [Google Scholar] [CrossRef]
- Schuler, J.L.; Robison, D.J. Performance of northern red oak enrichment plantings in naturally regenerating Southern Appalachian hardwood stands. New For. 2010, 40, 119–130. [Google Scholar] [CrossRef]
- Lorimer, C.G.; Chapman, J.W.; Lambert, W.D. Tall understorey vegetation as a factor in the poor development of oak seedlings beneath mature stands. J. Ecol. 1994, 82, 227–237. [Google Scholar] [CrossRef]
- Dalgleish, H.J.; Lichti, N.I.; Schmedding, N.; Swihart, R.K. Exposure to herbivores increases seedling growth and survival of American chestnut (Castanea dentata) through decreased interspecific competition in canopy gaps. Restor. Ecol. 2015, 23, 655–661. [Google Scholar] [CrossRef]
- Ross-Davis, A.; Broussard, S. A typology of family forest owners in North Central Indiana. North J. Appl. 2007, 24, 282–289. [Google Scholar]
- Kittredge, D.B.; Finley, A.O.; Foster, D.R. Timber harvesting as ongoing disturbance in a landscape of diverse ownership. For. Ecol. Manag. 2003, 180, 425–442. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Basal Area | Initial BA Change | |||
|---|---|---|---|---|---|
| 2007 (Y0) (m2 ha−1) | 2009 (Y2) | 2011 (Y4) † | 2018 (Y10) | Percent | |
| Control | 26.7 ± 3.2 | 28.5 ± 3.0 | 28.4 ± 2.7 | 23.9 ± 2.0 | +7.0 ± 5.3 a |
| Midstory Removal | 27.4 ± 3.1 | 21.3 ± 2.8 | 21.2 ± 2.6 | 20.8 ± 1.9 | −24.1 ± 5.0 b |
| Thinning | 30.1 ± 3.4 | 20.4 ± 3.2 | 20.7 ± 2.9 | 19.3 ± 2.1 | −29.2 ± 5.7 b |
| Shelterwood | 30.0 ± 3.1 | 20.2 ± 2.8 | 19.9 ± 2.6 | 17.9 ± 1.8 | −36.1 ± 5.0 b |
| Response | Fixed Effects | Estimate | SE | z Value | Significance | 95% Conf. Int. | |
|---|---|---|---|---|---|---|---|
| Lower | Upper | ||||||
| Survival | |||||||
| Silvicultural Treatment | |||||||
| Midstory Removal | 3.817 a | 0.630 | 6.055 | *** | 2.233 | 5.401 | |
| Thinning | 2.781 b | 0.659 | 4.218 | *** | 1.125 | 4.438 | |
| Shelterwood | 4.317 a | 0.629 | 6.865 | *** | 2.737 | 5.897 | |
| Fencing | 2.838 | 0.686 | 4.136 | *** | 1.114 | 4.562 | |
| Fertilization | 0.015 | 0.092 | 0.160 | −0.217 | 0.246 | ||
| Silvi. × Fence | |||||||
| MR × Fn | −1.328 a | 0.641 | −2.071 | −2.939 | 0.283 | ||
| TH × Fn | −0.712 a | 0.663 | −1.075 | −2.377 | 0.952 | ||
| SW × Fn | −1.968 b | 0.639 | −3.082 | * | −3.572 | −0.364 | |
| Competitiveness | |||||||
| Silvicultural Treatment | |||||||
| Midstory Removal † | - | - | |||||
| Thinning | −2.000 a | 0.820 | −2.439 | · | −4.181 | 0.181 | |
| Shelterwood | 0.341 b | 0.437 | 0.780 | −0.821 | 1.503 | ||
| Fencing | 1.790 | 0.401 | 4.464 | *** | 0.724 | 2.856 | |
| Fertilization | 0.974 | 0.313 | 3.106 | * | 0.140 | 1.807 | |
| Silvi × Fence | |||||||
| TH × Fn | 1.831 a | 0.778 | 2.353 | −0.238 | 3.899 | ||
| SW × Fn | −0.318 b | 0.358 | −0.890 | −1.269 | 0.633 | ||
| Fert × Fence | −0.795 | 0.357 | −2.227 | −1.744 | 0.154 | ||
| Silvicultural Treatment | Fence | Fertilized | Predicted Probability of Survival | Predicted Probability of Competitiveness |
|---|---|---|---|---|
| SW | Y | Y | 0.531 | 0.161 |
| MR | Y | Y | 0.565 | 0.158 |
| SW | Y | N | 0.527 | 0.138 |
| TH | Y | Y | 0.461 | 0.137 |
| MR | Y | N | 0.562 | 0.135 |
| TH | Y | N | 0.457 | 0.117 |
| SW | N | Y | 0.321 | 0.089 |
| MR | N | Y | 0.223 | 0.065 |
| SW | N | N | 0.318 | 0.035 |
| MR | N | N | 0.220 | 0.025 |
| TH | N | Y | 0.092 | 0.009 |
| TH | N | N | 0.091 | 0.004 |
| Control | Y | Y | 0.097 | -- |
| Control | Y | N | 0.096 | -- |
| Control | N | Y | 0.006 | -- |
| Control | N | N | 0.006 | -- |
| Response | Fixed Effects | Estimate | SE | t Value | Signif. | 95% Conf. Int. | |
|---|---|---|---|---|---|---|---|
| Lower | Upper | ||||||
| Height (cm) | |||||||
| Silvicultural Treatment | |||||||
| Midstory Removal | 1.135 | 0.218 | 5.205 | *** | 0.560 | 1.710 | |
| Thinning | 0.877 | 0.226 | 3.881 | *** | 0.282 | 1.472 | |
| Shelterwood | 1.288 | 0.217 | 5.929 | *** | 0.715 | 1.860 | |
| Fencing | 0.299 | 0.068 | 4.379 | *** | 0.119 | 0.479 | |
| Fertilization | 0.537 | 0.228 | 2.357 | −0.063 | 1.138 | ||
| Silvi. × Fert. | |||||||
| MR × Fert. | −0.502 | 0.227 | −2.215 | −1.100 | 0.095 | ||
| TH × Fert. | −0.539 | 0.235 | −2.289 | −1.160 | 0.081 | ||
| SW × Fert. | −0.310 | 0.226 | −1.370 | −0.907 | 0.286 | ||
| GLD (mm) | |||||||
| Silvicultural Treatment | |||||||
| Midstory Removal | 0.516 ab | 0.117 | 4.416 | *** | 0.214 | 0.818 | |
| Thinning | 0.282 a | 0.121 | 2.336 | −0.030 | 0.595 | ||
| Shelterwood | 0.664 b | 0.117 | 5.692 | *** | 0.363 | 0.965 | |
| Fencing | 0.184 | 0.045 | 4.054 | *** | 0.067 | 0.301 | |
| Fertilization | 0.049 | 0.050 | 0.979 | −0.081 | 0.179 | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frank, G.S.; Rathfon, R.A.; Saunders, M.R. Ten-Year Responses of Underplanted Northern Red Oak to Silvicultural Treatments, Herbivore Exclusion, and Fertilization. Forests 2018, 9, 571. https://doi.org/10.3390/f9090571
Frank GS, Rathfon RA, Saunders MR. Ten-Year Responses of Underplanted Northern Red Oak to Silvicultural Treatments, Herbivore Exclusion, and Fertilization. Forests. 2018; 9(9):571. https://doi.org/10.3390/f9090571
Chicago/Turabian StyleFrank, Graham S., Ronald A. Rathfon, and Michael R. Saunders. 2018. "Ten-Year Responses of Underplanted Northern Red Oak to Silvicultural Treatments, Herbivore Exclusion, and Fertilization" Forests 9, no. 9: 571. https://doi.org/10.3390/f9090571
APA StyleFrank, G. S., Rathfon, R. A., & Saunders, M. R. (2018). Ten-Year Responses of Underplanted Northern Red Oak to Silvicultural Treatments, Herbivore Exclusion, and Fertilization. Forests, 9(9), 571. https://doi.org/10.3390/f9090571