Generalized Nonlinear Mixed-Effects Individual Tree Crown Ratio Models for Norway Spruce and European Beech



Abstract
:1. Introduction
2. Materials and Methods
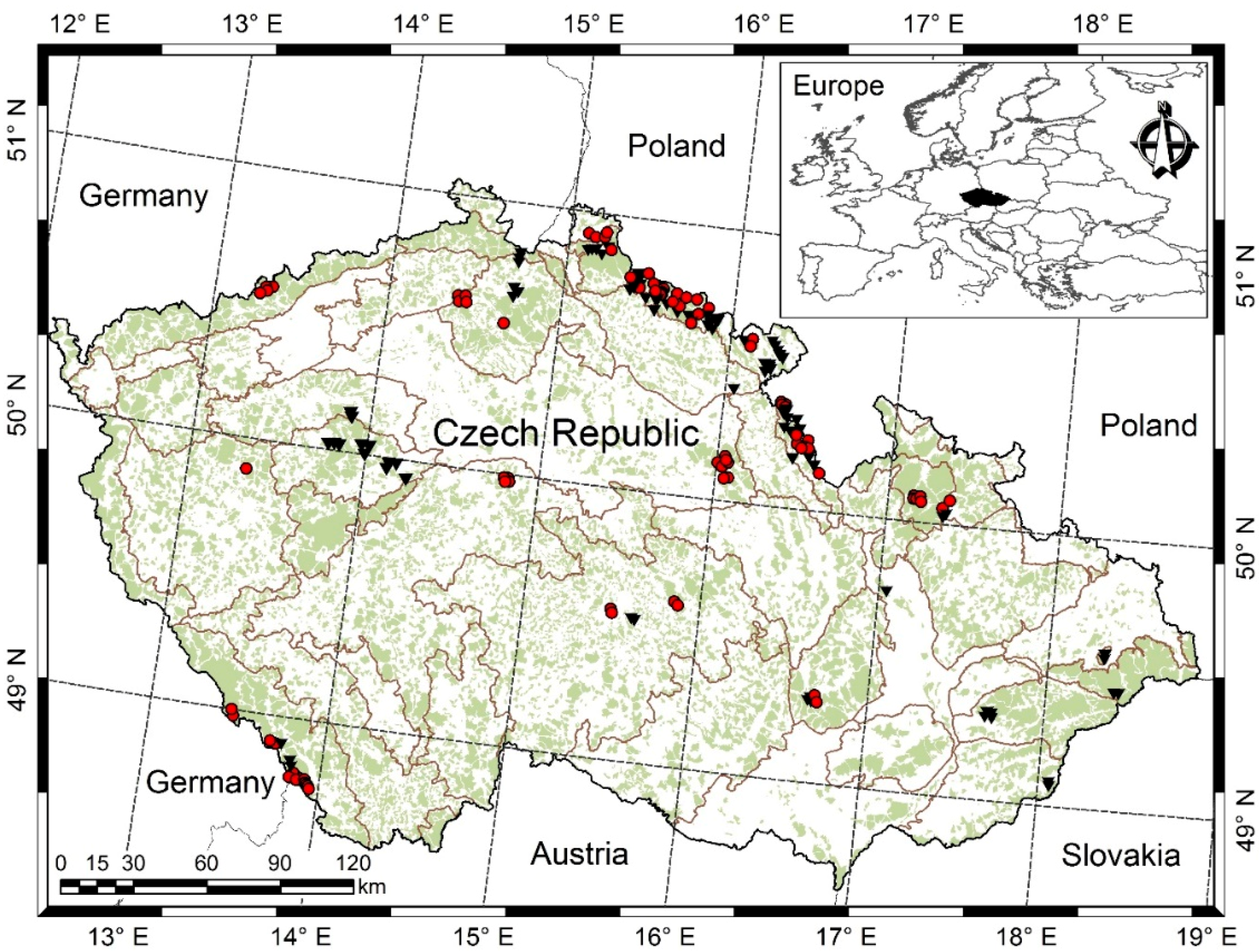
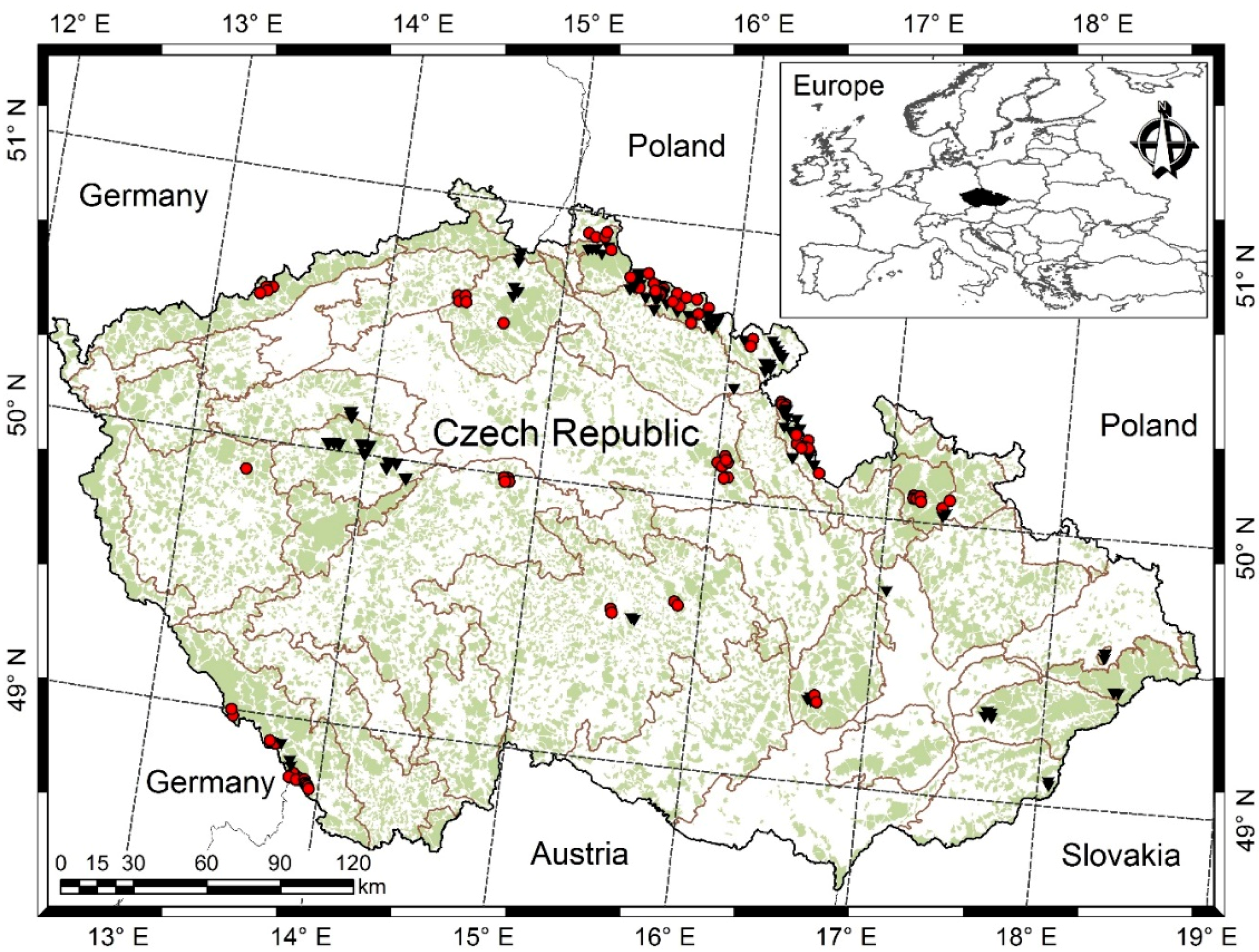
2.1. Sampling and Measurements
2.2. Data Analysis
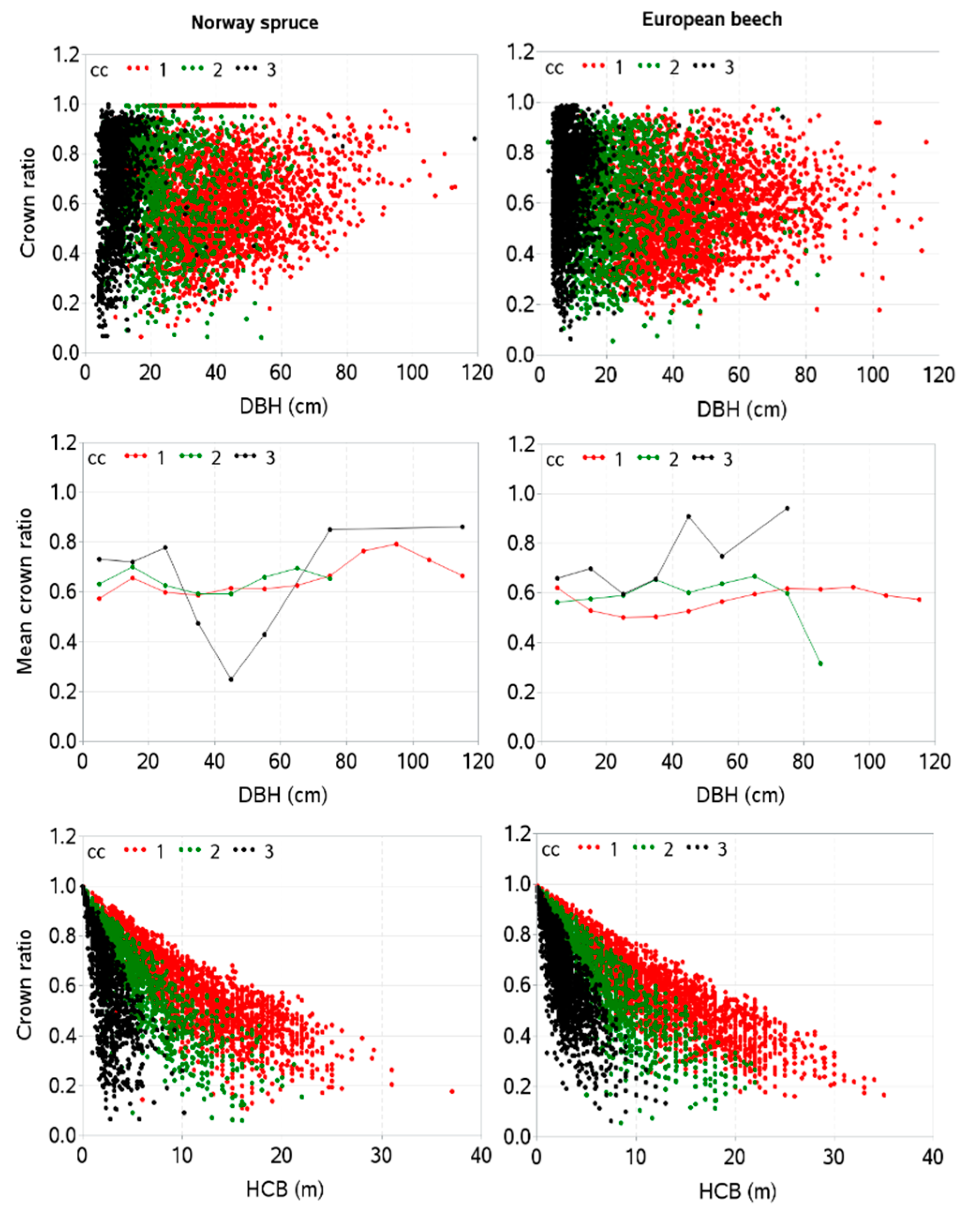
2.2.1. Tree and Stand Measures
- canopy height class I, CC1: height greater than 66% of the tallest tree;
- canopy height class II, CC2: height between 33% and 66% of the tallest tree; and
- canopy height class III, CC3: height less than 33% of the tallest tree.
2.2.2. Model Development
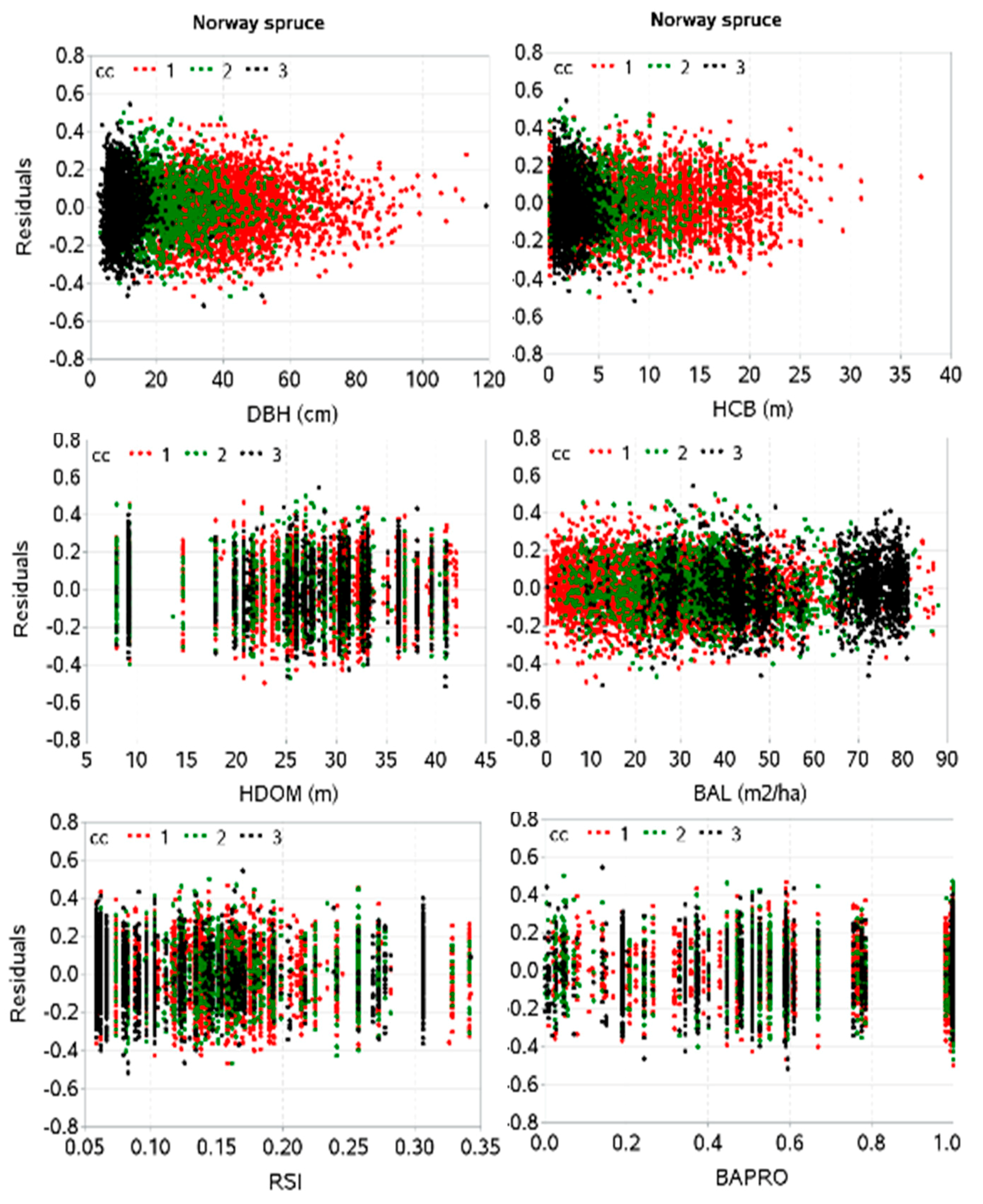
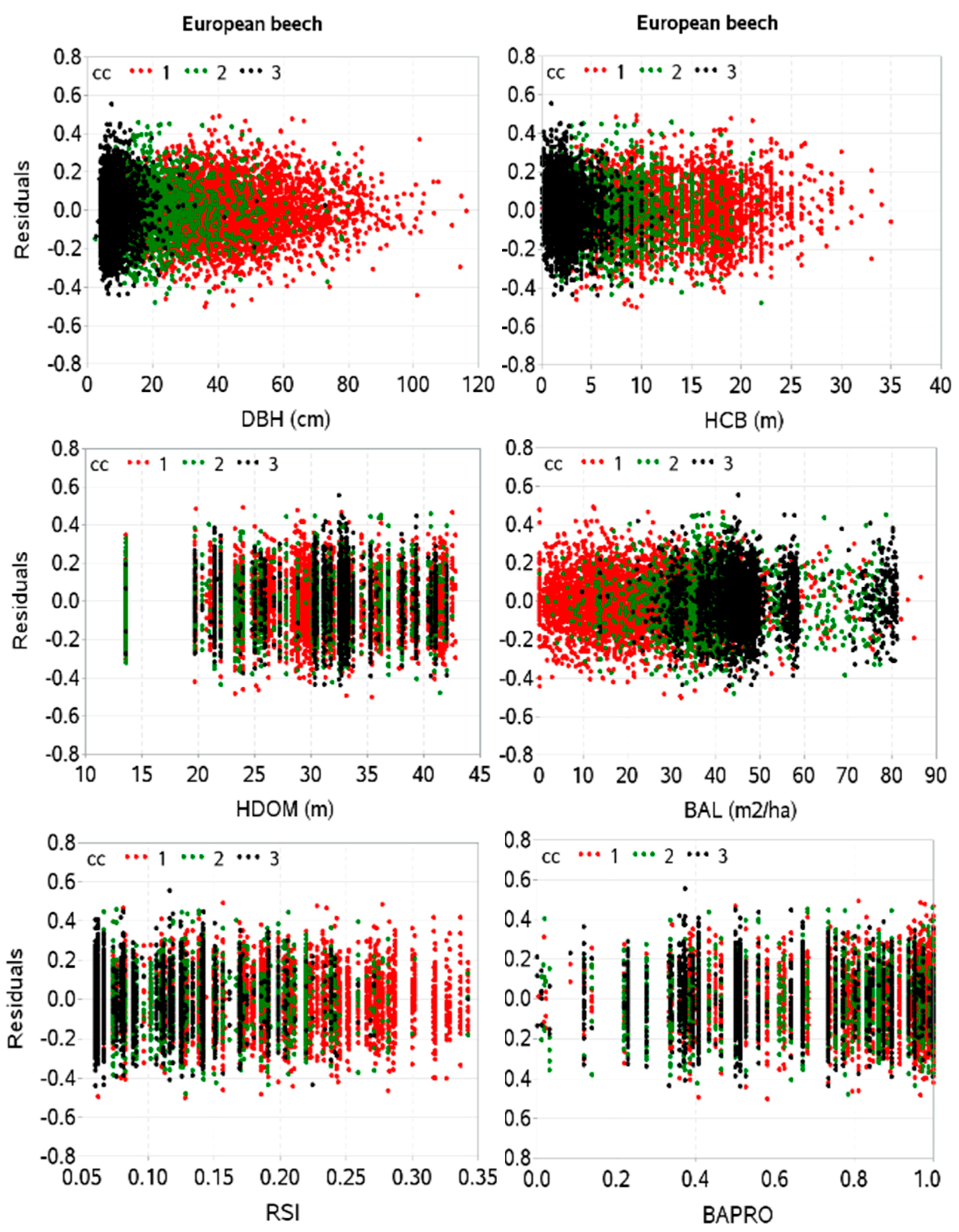
2.3. Estimation of Model Parameters and Evaluation
2.4. The Localizing Mixed-Effects Model and Subject-Specific CR Prediction
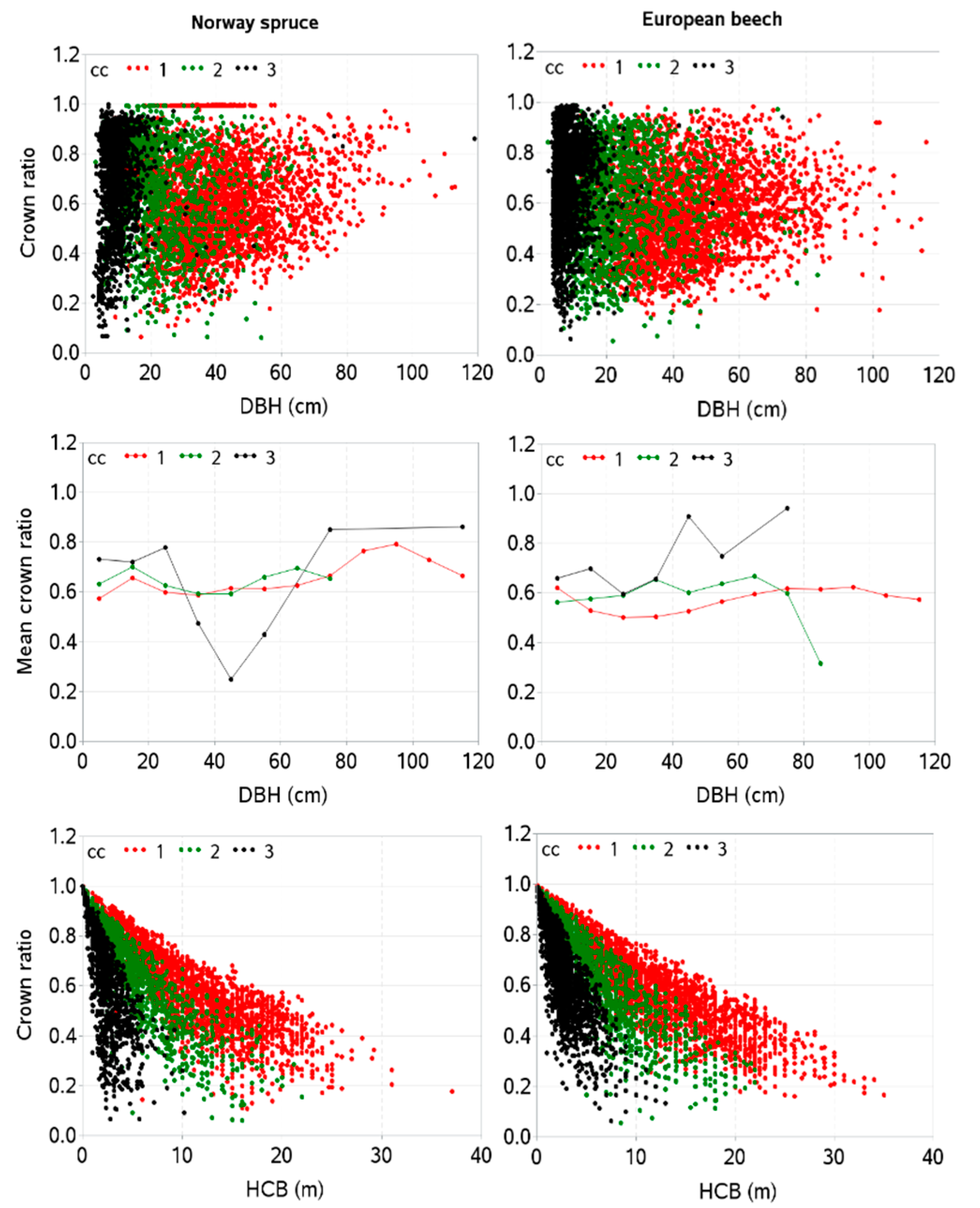
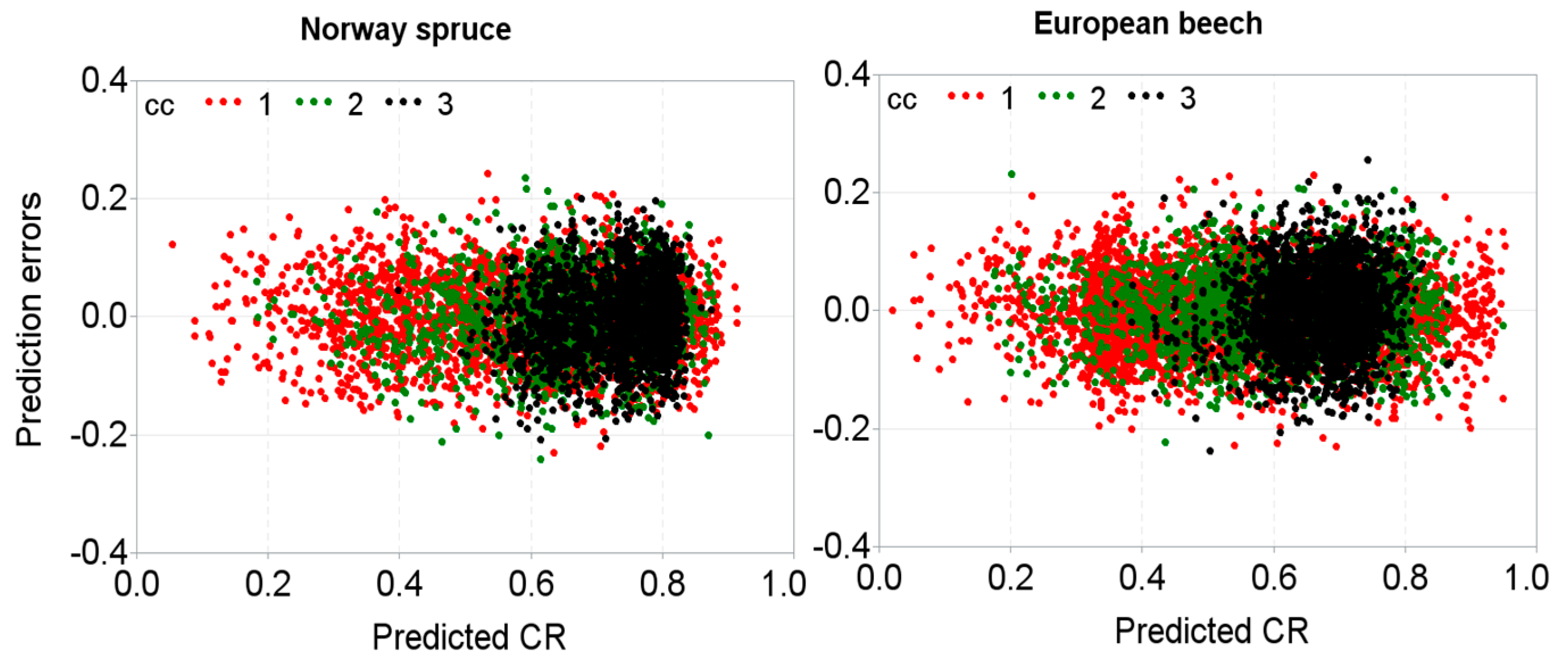
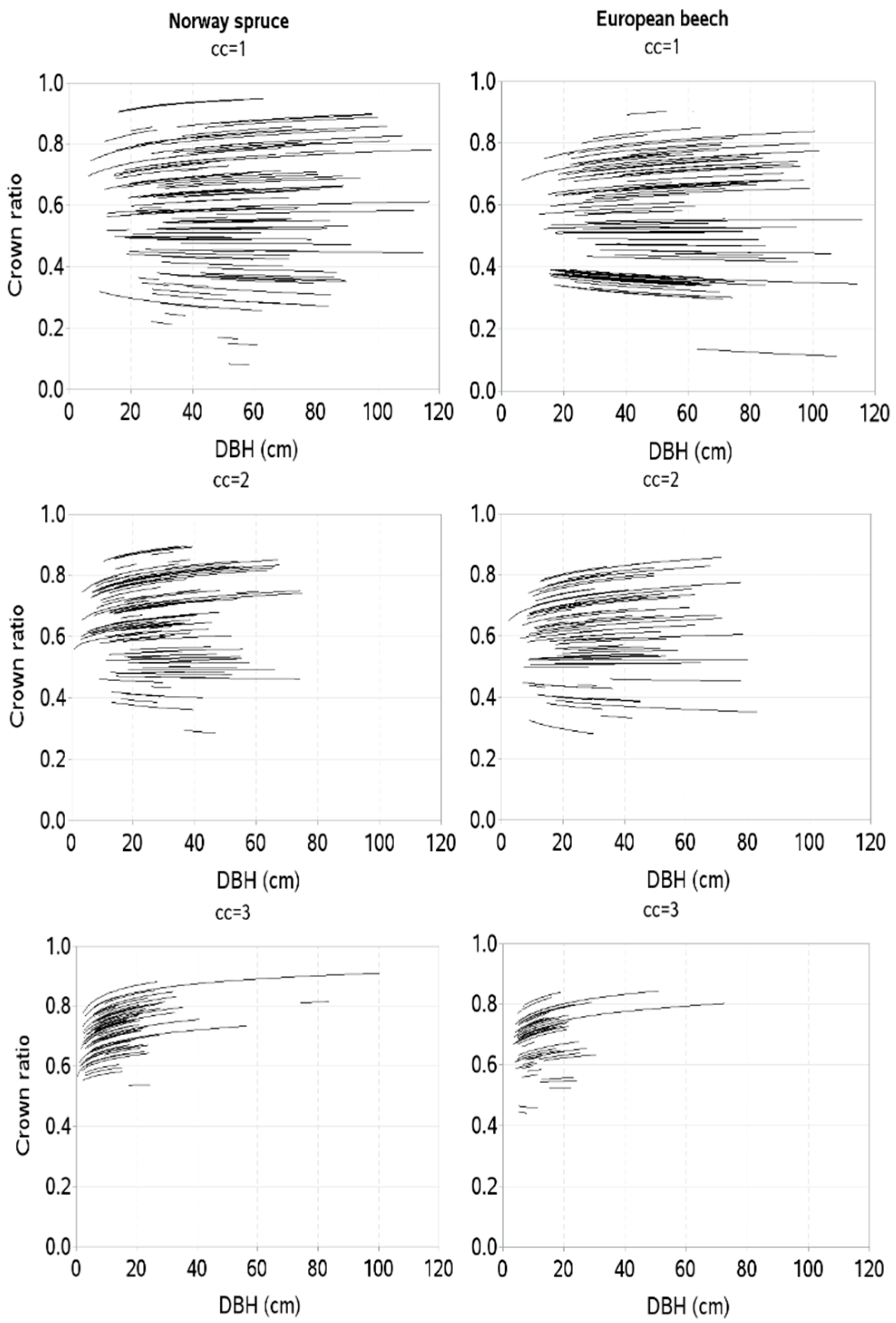
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zarnoch, S.J.; Bechtold, W.A.; Stolte, K.W. Using crown condition variables as indicators of forest health. Can. J. For. Res. 2004, 34, 1057–1070. [Google Scholar] [CrossRef]
- Buckley, T.N.; Cescatti, A.; Farquhar, G.D. What does optimization theory actually predict about crown profiles of photosynthetic capacity when models incorporate greater realism? Plant Cell Environ. 2013, 36, 1547–1563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assmann, E. The Principles of Forest Yield Studies; Pergamon Press: Oxford, UK, 1970; p. 506. [Google Scholar]
- Larocque, G.R.; Marshall, P.L. Crown development in red pine stands. I. Absolute and relative growth measures. Can. J. For. Res. 1994, 24, 762–774. [Google Scholar] [CrossRef]
- Larocque, G.R.; Marshall, P.L. Crown development in red pine stands. II. Relationships with stem growth. Can. J. For. Res. 1994, 24, 775–784. [Google Scholar] [CrossRef]
- Grote, R. Estimation of crown radii and crown projection area from stem size and tree position. Ann. For. Sci. 2003, 60, 393–402. [Google Scholar] [CrossRef] [Green Version]
- Jimenez-Perez, J.; Aguirre, O.; Kramer, H. Tree crown structure indicators in a natural uneven-aged mixed coniferous forest in northeastern Mexico. In Monitoring Science and Technology Symposium: Unifying Knowledge for Sustainability in the Western Hemisphere Proceedings RMRS-P-42CD; Aguirre-Bravo, C., Pellicane, P.J., Burns, D.P., Draggan, S., Eds.; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2006; pp. 649–654. [Google Scholar]
- Hasenauer, H.; Monserud, R.A. A crown ratio model for Austrian forests. For. Ecol. Manag. 1996, 84, 49–60. [Google Scholar] [CrossRef]
- Smith, D.M. The Practice of Silviculture; John Wiley: New York, NY, USA, 1988. [Google Scholar]
- Dyer, M.E.; Burkhart, H.E. Compatible crown ratio and crown height models. Can. J. For. Res. 1987, 17, 572–574. [Google Scholar] [CrossRef]
- Zhao, D.; Kane, M.; Borders, B.E. Crown ratio and relative spacing relationships for loblolly pine plantations. Open J. For. 2012, 2, 101–115. [Google Scholar] [CrossRef]
- Kuprevicius, A.; Auty, D.; Achim, A.; Caspersen, J.P. Quantifying the influence of live crown ratio on the mechanical properties of clear wood. Forestry 2014, 87, 449–458. [Google Scholar] [CrossRef]
- Kershaw, J.A., Jr.; Maguire, D.A.; Hann, D.W. Longevity and duration of radial growth in Douglas-fir branches. Can. J. For. Res. 1990, 20, 1690–1695. [Google Scholar] [CrossRef]
- Navratil, S. Wind damage in thinned stands. In Proceedings of the A Commercial Thinning Workshop, Whitecourt, AB, Canada, 17–18 October 1997; pp. 29–36. [Google Scholar]
- Ritchie, M.W.; Hann, D.W. Equations for Predicting Height to Crown Base for Fourteen Tree Species in Southwest Oregon; Forestry Research Laboratory, Oregon State University: Corvallis, OR, USA, 1987. [Google Scholar]
- Biging, G.S.; Dobbertin, M. Comparison of distance-dependent competition measures for height and basal area growth of individual conifer trees. For. Sci. 1992, 38, 695–720. [Google Scholar]
- Monserud, R.A.; Sterba, H. A basal area increment model for individual trees growing in even- and uneven-aged forest stands in Austria. For. Ecol. Manag. 1996, 80, 57–80. [Google Scholar] [CrossRef]
- Hasenauer, H.; Monserud, R.A. Biased predictions for tree height increment models developed from smoothed ‘data’. Ecol. Model. 1997, 98, 13–22. [Google Scholar] [CrossRef]
- Pretzsch, H.; Biber, P.; Dursky, J. The single tree-based stand simulator SILVA: Construction, application and evaluation. For. Ecol. Manag. 2002, 162, 3–21. [Google Scholar] [CrossRef]
- Saud, P.; Lynch, T.B.; KC, A.; Guldin, J.M. Using quadratic mean diameter and relative spacing index to enhance height-diameter and crown ratio models fitted to longitudinal data. Forestry 2016, 89, 215–229. [Google Scholar] [CrossRef]
- Wykoff, W.R. A basal area increment model for individual conifers in the Northern Rocky Mountain. For. Sci. 1990, 36, 1077–1104. [Google Scholar]
- Sprinz, P.T.; Burkhart, H.E. Relationships between tree crown, stem, and stand characteristics in unthinned loblolly pine plantations. Can. J. For. Res. 1987, 17, 534–538. [Google Scholar] [CrossRef]
- Ritchie, M.W.; Hann, D.W. Equations for Predicting Basal Area Increment for Douglas-Fir and Grand Fir; Forest Research Laboratory, Oregon State University Research Bulletin: Corvallis, OR, USA, 1985; p. 9. [Google Scholar]
- Leites, L.P.; Robinson, A.P.; Crookston, N.L. Accuracy and equivalence testing of crown ratio models and assessment of their impact on diameter growth and basal area increment predictions of two variants of the Forest Vegetation Simulator. Can. J. For. Res. 2009, 39, 655–665. [Google Scholar] [CrossRef]
- Saud, P.; Lynch, T.B.; Guldin, J.M. Twenty five years long survival analysis of an individual shortleaf pine trees. In Proceedings of the 18th Biennial Southern Silvicultural Research Conference, Knoxville, TN, USA, 2–5 March 2015; Schweitzer, C.J., Clatterbuck, W.K., Oswalt, C.M., Eds.; US Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2016; pp. 555–557. [Google Scholar]
- Monserud, R.A.; Sterba, H. Modeling individual tree mortality for Austrian forest species. For. Ecol. Manag. 1999, 113, 109–123. [Google Scholar] [CrossRef]
- Carvalho, J.P.; Parresol, B.R. Additivity in tree biomass components of Pyrenean oak (Quercus pyrenaica Willd.). For. Ecol. Manag. 2003, 179, 269–276. [Google Scholar] [CrossRef]
- Tahvanainen, L. Allometric relationships to estimate above-ground dry-mass and height in Salix. Scand. J. For. Res. 1996, 11, 233–241. [Google Scholar] [CrossRef]
- Jucker, T.; Caspersen, J.; Chave, J.; Antin, C.; Barbier, N.; Bongers, F.; Dalponte, M.; van Ewijk, K.Y.; Forrester, D.I.; Haeni, M.; et al. Allometric equations for integrating remote sensing imagery into forest monitoring programmes. Glob. Chang. Biol. 2017, 23, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Walters, D.K.; Hann, D.W. Taper Equations for Six Conifer Species in Southwest Oregon; Forest Research Laboratory, Oregon State University, Research Bulletin: Corvallis, OR, USA, 1986; Volume 56, p. 41. [Google Scholar]
- Jiang, L.; Liu, R. Segmented taper equations with crown ratio and stand density for Dahurian Larch (Larix gmelinii) in Northeastern China. J. For. Res. 2011, 22, 347–352. [Google Scholar] [CrossRef]
- Gonzalez-Benecke, C.A.; Gezan, S.A.; Samuelson, L.J.; Cropper, W.P.; Leduc, D.J.; Martin, T.A. Estimating Pinus palustris tree diameter and stem volume from tree height, crown area and stand-level parameters. J. For. Res. 2014, 25, 43–52. [Google Scholar] [CrossRef]
- Clutter, J.L.; Fortson, J.C.; Pienaar, L.V.; Brister, G.H.; Bailey, R.L. Timber Management: A Quantitative Approach; Wiley: New York, NY, USA, 1983; p. 333. [Google Scholar]
- McGaughey, R.J. Visualizing forest stand dynamics using the stand visualization system. In Proceedings of the 1997 ACSM/ASPRS Annual Convention and Exposition, Seattle, WA, USA, 7–10 April 1997; pp. 248–257. [Google Scholar]
- Tews, J.; Brose, U.; Grimm, V.; Tielbörger, K.; Wichmann, M.C.; Schwager, M.; Jeltsch, F. Animal species diversity driven by habitat heterogeneity/diversity: The importance of keystone structures. J. Biogeogr. 2004, 31, 79–92. [Google Scholar] [CrossRef]
- Keane, R.E.; Mincemoyer, S.A.; Schmidt, K.M.; Long, D.G.; Garner, J.L. Mapping Vegetation and Fuels for Fire Management on the Gila National Forest Complex, New Mexico; General Technical Report (GTR) RMRS-GTR-46-CD; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 2000; p. 126.
- Oker-Blom, P.; Pukkala, T.; Kuuluvainen, T. Relationship between radiation interception and photosynthesis in forest canopies: Effect of stand structure and latitude. Ecol. Model. 1989, 49, 73–87. [Google Scholar] [CrossRef]
- Pukkala, T.; Becker, P.; Kuuluvainen, T.; Oker-Blom, P. Predicting spatial distribution of direct radiation below forest canopies. Agric. For. Meteorol. 1991, 55, 295–307. [Google Scholar] [CrossRef]
- Gill, S.J.; Biging, G.S.; Murphy, E.C. Modeling conifer tree crown radius and estimating canopy cover. For. Ecol. Manag. 2000, 126, 405–416. [Google Scholar] [CrossRef]
- Bragg, D.C. A local basal area adjustment for crown width prediction. North. J. Appl. For. 2001, 18, 22–28. [Google Scholar]
- Fu, L.; Sun, H.; Sharma, R.P.; Lei, Y.; Zhang, H.; Tang, S. Nonlinear mixed-effects crown width models for individual trees of Chinese fir (Cunninghamia lanceolata) in south-central China. For. Ecol. Manag. 2013, 302, 210–220. [Google Scholar] [CrossRef]
- Sharma, R.P.; Vacek, Z.; Vacek, S. Individual tree crown width models for Norway spruce and European beech in Czech Republic. For. Ecol. Manag. 2016, 366, 208–220. [Google Scholar] [CrossRef]
- Fu, L.; Sharma, R.P.; Hao, K.; Tang, S. A generalized interregional nonlinear mixed-effects crown width model for Prince Rupprecht larch in northern China. For. Ecol. Manag. 2017, 389, 364–373. [Google Scholar] [CrossRef]
- Sharma, R.P.; Bíllek, L.; Vacek, Z.; Vacek, S. Modeling crown width-diameter relationship for Scots pine in the central Europe. Trees 2017, 31, 1875–1889. [Google Scholar] [CrossRef]
- Deleuze, C.; Hervé, J.-C.; Colin, F.; Ribeyrolles, L. Modeling crown shape of Piceaabies: Spacing effects. Can. J. For. Res. 1996, 26, 1957–1966. [Google Scholar] [CrossRef]
- Valentine, H.T.; Amateis, R.L.; Gove, J.H.; Mäkelä, A. Crown-rise and crown-length dynamics: Application to loblolly pine. Forestry 2013, 86, 371–375. [Google Scholar] [CrossRef]
- Taylor, J.E.; Ellis, M.V.; Rayner, L.; Ross, K.A. Variability in allometric relationships for temperate woodland Eucalyptus trees. For. Ecol. Manag. 2016, 360, 122–132. [Google Scholar] [CrossRef]
- Sharma, R.P.; Vacek, Z.; Vacek, S. Modeling tree crown-to-bole diameter ratio for Norway spruce and European beech. Silva Fennica 2017, 51, 1740. [Google Scholar] [CrossRef]
- Soares, P.; Tomé, M. A tree crown ratio prediction equation for eucalypt plantations. Ann. For. Sci. 2001, 58, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Temesgen, H.; LeMay, V.; Mitchell, S.J. Tree crown ratio models for multi-species and multi-layered stands of southeastern British Columbia. For. Chron. 2005, 81, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Maguire, D.A.; Hann, D.W. Constructing models for direct prediction of 5-year crown recession in southwestern Oregon Douglas-fir. Can. J. For. Res. 1990, 20, 1044–1052. [Google Scholar] [CrossRef]
- Hynynen, J. Predicting tree crown ratio for unthinned and thinned Scots pine stands. Can. J. For. Res. 1995, 25, 57–62. [Google Scholar] [CrossRef]
- Fu, L.; Zhang, H.; Lu, J.; Zang, H.; Lou, M.; Wang, G. Multilevel nonlinear mixed-effect crown ratio models for individual trees of Mongolian Oak (Quercus mongolica) in northeast China. PLoS ONE 2015, 10, e0133294. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, J.C.; Bates, D.M. Mixed-Effects Models in S and S-PLUS; Springer: New York, NY, USA, 2000. [Google Scholar]
- Sharma, R.P.; Breidenbach, J. Modeling height-diameter relationships for Norway spruce, Scots pine, and downy birch using Norwegian national forest inventory data. For. Sci. Technol. 2015, 11, 44–53. [Google Scholar] [CrossRef]
- Sharma, R.P.; Vacek, Z.; Vacek, S.; Podrázský, V.; Jansa, V. Modeling individual tree height to crown base of Norway spruce (Picea abies (L.) Karst.) and European beech (Fagus sylvatica L.). PLoS ONE 2017, 12, e0186394. [Google Scholar] [CrossRef] [PubMed]
- Lappi, J.; Bailey, R.L. A height prediction model with random stand and tree parameters-an alternative to traditional site index methods. For. Sci. 1988, 34, 907–927. [Google Scholar]
- Sterba, H.; Rio, M.; Brunner, A.; Condes, S. Effect of species proportion definition on the evaluation of growth in pure vs. mixed stands. For. Syst. 2014, 23, 547–559. [Google Scholar] [CrossRef]
- Pretzsch, H.; Forrester, D.I.; Rötzer, T. Representation of species mixing in forest growth models. A review and perspective. Ecol. Model. 2015, 313, 276–292. [Google Scholar] [CrossRef] [Green Version]
- Pretzsch, H. Canopy space filling and tree crown morphology in mixed-species stands compared with monocultures. For. Ecol. Manag. 2014, 327, 251–264. [Google Scholar] [CrossRef]
- Sharma, R.P.; Vacek, Z.K.; Vacek, S. Modeling individual tree height to diameter ratio for Norway spruce and European beech in Czech Republic. Trees 2016, 30, 1969–1982. [Google Scholar] [CrossRef]
- Wonn, H.T.; O’Hara, K.L. Height: Diameter ratios and stability relationships for four northern rocky mountain tree species. West. J. Appl. For. 2001, 16, 87–94. [Google Scholar]
- Vacek, Z.; Vacek, S.; Bílek, L.; Remeš, J.; Štefančík, I. Changes in horizontal structure of natural beech forests on an altitudinal gradient in the Sudetes. Dendrobiology 2015, 73, 33–45. [Google Scholar] [CrossRef] [Green Version]
- Bulušek, D.; Vacek, Z.; Vacek, S.; Král, J.; Bílek, L.; Králíček, I. Spatial pattern of relict beech (Fagus sylvatica L.) forests in the Sudetes of the Czech Republic and Poland. J. For. Sci. 2016, 62, 293–305. [Google Scholar] [CrossRef]
- Šmelko, Š.S.; Merganič, J. Some methodological aspects of the national forest inventory and monitoring in Slovakia. J. For. Sci. 2008, 54, 476–483. [Google Scholar] [CrossRef]
- Vacek, S.; Bílek, L.; Schwarz, O.; Hejcmanová, P.; Mikeska, M. Effect of Air Pollution on the Health Status of Spruce Stands. Mt. Res. Dev. 2013, 33, 40–50. [Google Scholar] [CrossRef]
- Vacek, Z.; Vacek, S.; Podrázský, V.; Bílek, L.; Štefančík, I.; Moser, W.K.; Bulušek, D.; Král, J.; Remeš, J.; Králíček, I. Effect of tree layer and microsite on the variability of natural regeneration in autochthonous beech forests. Pol. J. Ecol. 2015, 63, 233–246. [Google Scholar] [CrossRef]
- Forest Management Institute (FMI). Inventarizace lesů, Metodika Venkovního Sběru Dat (Forest Inventory, Field Data Collection Methodology); FMI: Brandýs nad Labem, Czech Republic, 2003; p. 136. [Google Scholar]
- Fu, L.; Sharma, R.P.; Wang, G.; Tang, S. Modeling a system of nonlinear additive crown width models applying seemingly unrelated regression for Prince Rupprecht larch in northern China. For. Ecol. Manag. 2017, 386, 71–80. [Google Scholar] [CrossRef]
- Eid, T.; Tuhus, E. Models for individual tree mortality in Norway. For. Ecol. Manag. 2001, 154, 69–84. [Google Scholar] [CrossRef]
- Bollandsås, O.M.; Næsset, E. Weibull models for single-tree increment of Norway spruce, Scots pine, birch and other broadleaves in Norway. Scand. J. For. Res. 2009, 24, 54–66. [Google Scholar] [CrossRef]
- Sharma, R.P.; Brunner, A.; Eid, T.; Øyen, B.-H. Modeling dominant height growth from national forest inventory individual tree data with short time series and large age errors. For. Ecol. Manag. 2011, 262, 2162–2175. [Google Scholar] [CrossRef]
- Monserud, R.A. Height growth and site index curves for inland Douglas-fir based on stem analysis data and forest habitat type. For. Sci. 1984, 30, 943–965. [Google Scholar]
- Corral-Rivas, J.J.; Gonzalez, J.G.A.; Aguirre, O.; Hernandez, F. The effect of competition on individual tree basal area growth in mature stands of Pinus cooperi Blanco in Durango (Mexico). Eur. J. For. Res. 2005, 124, 133–142. [Google Scholar] [CrossRef]
- Schroder, J.; von Gadow, K. Testing a new competition index for Maritime pine in northwestern Spain. Can. J. For. Res. 1999, 29, 280–283. [Google Scholar] [CrossRef]
- Fonseca, T.F.; Duarte, J.C. A silvicultural stand density model to control understory in maritime pine stands. iForest 2017, 10, 829–836. [Google Scholar] [CrossRef] [Green Version]
- Uzoh, F.C.C.; Oliver, W.W. Individual tree diameter increment model for managed even-aged stands of ponderosa pine throughout the western United States using a multilevel linear mixed effects model. For. Ecol. Manag. 2008, 256, 438–445. [Google Scholar] [CrossRef]
- Staudhammer, C.; LeMay, V. Height prediction equations using diameter and stand density measures. For. Chron. 2000, 76, 303–309. [Google Scholar] [CrossRef] [Green Version]
- Montgomery, D.C.; Peck, E.A.; Vining, G.G. Introduction to Linear Regression Analysis, 3rd ed.; Wiley: New York, NY, USA, 2001; p. 641. [Google Scholar]
- Crecente-Campo, F.; Tomé, M.; Soares, P.; Dieguez-Aranda, U. A generalized nonlinear mixed-effects height-diameter model for Eucalyptus globulus L. in northwestern Spain. For. Ecol. Manag. 2010, 259, 943–952. [Google Scholar] [CrossRef]
- Gregoire, T.G.; Schabenberger, O.; Barrett, J.P. Linear modeling of irregularly spaced, unbalanced, longitudinal data from permanent-plot measurements. Can. J. For. Res. 1995, 25, 137–156. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS/ETS 9.1.3 User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2012. [Google Scholar]
- Littell, R.C.; Milliken, G.A.; Stroup, W.W.; Wolfinger, R.D.; Schabenberger, O. SAS for Mixed Models, 2nd ed.; SAS Institute Inc.: Cary, NC, USA, 2006; p. 814. [Google Scholar]
- Akaike, H. A new look at statistical model identification. IEEE Trans. Autom. Control 1972, 19, 716–723. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Inference: A Practical Information-Theoretic Approach; Springer: New York, NY, USA, 2002. [Google Scholar]
- Yang, Y.; Monserud, R.A.; Huang, S. An evaluation of diagnostic tests and their roles in validating forest biometric models. Can. J. For. Res. 2004, 34, 619–629. [Google Scholar] [CrossRef]
- Kozak, A.; Kozak, R. Does cross validation provide additional information in the evaluation of regression models? Can. J. For. Res. 2003, 33, 976–987. [Google Scholar] [CrossRef]
- Zhang, L. Cross-validation of non-linear growth functions for modeling tree height-diameter relationships. Ann. Bot. 1997, 79, 251–257. [Google Scholar] [CrossRef]
- Sharma, R.P.; Vacek, Z.; Vacek, S. Nonlinear mixed effect height-diameter model for mixed species forests in the central part of the Czech Republic. J. For. Sci. 2016, 62, 470–484. [Google Scholar] [Green Version]
- Trincado, G.; VanderSchaaf, C.L.; Burkhart, H.E. Regional mixed-effects height-diameter models for loblolly pine (Pinus taeda L.) plantations. Eur. J. For. Res. 2007, 126, 253–262. [Google Scholar] [CrossRef]
- Fu, L.; Zhang, H.; Sharma, R.P.; Pang, L.; Wang, G. A generalized nonlinear mixed-effects height to crown base model for Mongolian oak in northeast China. For. Ecol. Manag. 2017, 384, 34–43. [Google Scholar] [CrossRef]
- Lynch, T.B.; Saud, P.; Dipesh, K.C.; Will, R.E. Plantation site index comparison of shortleaf pine and loblolly pine in Oklahoma, USA. For. Sci. 2016, 62, 546–552. [Google Scholar] [CrossRef]
- Sharma, M.; Amateis, R.L.; Burkhart, H.E. Top height definition and its effect on site index determination in thinned and unthinned loblolly pine plantations. For. Ecol. Manag. 2002, 168, 163–175. [Google Scholar] [CrossRef]
- Contreras, M.A.; Affleck, D.; Chung, W. Evaluating tree competition indices as predictors of basal area increment in western Montana forests. For. Ecol. Manag. 2011, 262, 1939–1949. [Google Scholar] [CrossRef]
- Coomes, D.A.; Grubb, P.J. Impacts of root competition in forests and woodlands: A theoretical framework and review of experiments. Ecol. Monogr. 2000, 70, 171–207. [Google Scholar] [CrossRef]
- Weiskittel, A.R. Forest Growth and Yield Modeling; Wiley-Blackwell: Hoboken, NJ, USA, 2011; p. 415. [Google Scholar]
- Pretzsch, H.; Biber, P.; Uhl, E.; Dahlhausen, J.; Rötzer, T.; Caldentey, J.; Koike, T.; van Con, T.; Chavanne, A.; Seifert, T.; et al. Crown size and growing space requirement of common tree species in urban centres, parks, and forests. Urban For. Urban Green. 2015, 14, 466–479. [Google Scholar] [CrossRef]
- Bayer, D.; Seifert, S.; Pretzsch, H. Structural crown properties of Norway spruce (Picea abies [L.] Karst.) and European beech (Fagus sylvatica [L.]) in mixed versus pure stands revealed by terrestrial laser scanning. Trees 2013, 27, 1035–1047. [Google Scholar] [CrossRef]
- Viewegh, A.; Kusbach, J.; Mikeska, M. Czech forest ecosystem classification. J. For. Sci. 2003, 49, 74–82. [Google Scholar] [CrossRef]
- Vacek, S.; Zingari, P.C.; Jeník, J.; Simon, J.; Smejkal, J.; Vančura, K. Mountain Forests of the Czech Republic; Inistry of Agriculture of the Czech Republic: Prague, Czech Republic, 2003; p. 320. [Google Scholar]
- Fabrika, M.; Ďurský, J. Algorithms and software solution of thinning models for SIBYLA growth simulator. J. For. Sci. 2005, 51, 431–445. [Google Scholar] [CrossRef]
- Penuelas, J.; Ogaya, R.; Boada, M.S.; Jump, A. Migration, invasion and decline: Changes in recruitment and forest structure in a warming-linked shift of European beech forest in Catalonia (NE Spain). Ecography 2007, 30, 829–837. [Google Scholar] [CrossRef]
- Bolte, A.; Hilbrig, L.; Grundmann, B.; Kampf, F.; Brunet, J.; Roloff, A. Climate change impacts on stand structure and competitive interactions in a southern Swedish spruce—Beech forest. Eur. J. For. Res. 2010, 12, 261–276. [Google Scholar] [CrossRef]
- Saltré, F.; Duputié, A.; Gaucherel, C.; Chuine, I. How climate, migration ability and habitat fragmentation affect the projected future distribution of European beech. Glob. Chang. Biol. 2015, 21, 897–910. [Google Scholar] [CrossRef] [PubMed]
- Harrington, T.B. Silvicultural Approaches for Thinning Southern Pines: Method, Intensity, and Timing; Georgia Forestry Commission: Macon, GA, USA, 2000. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
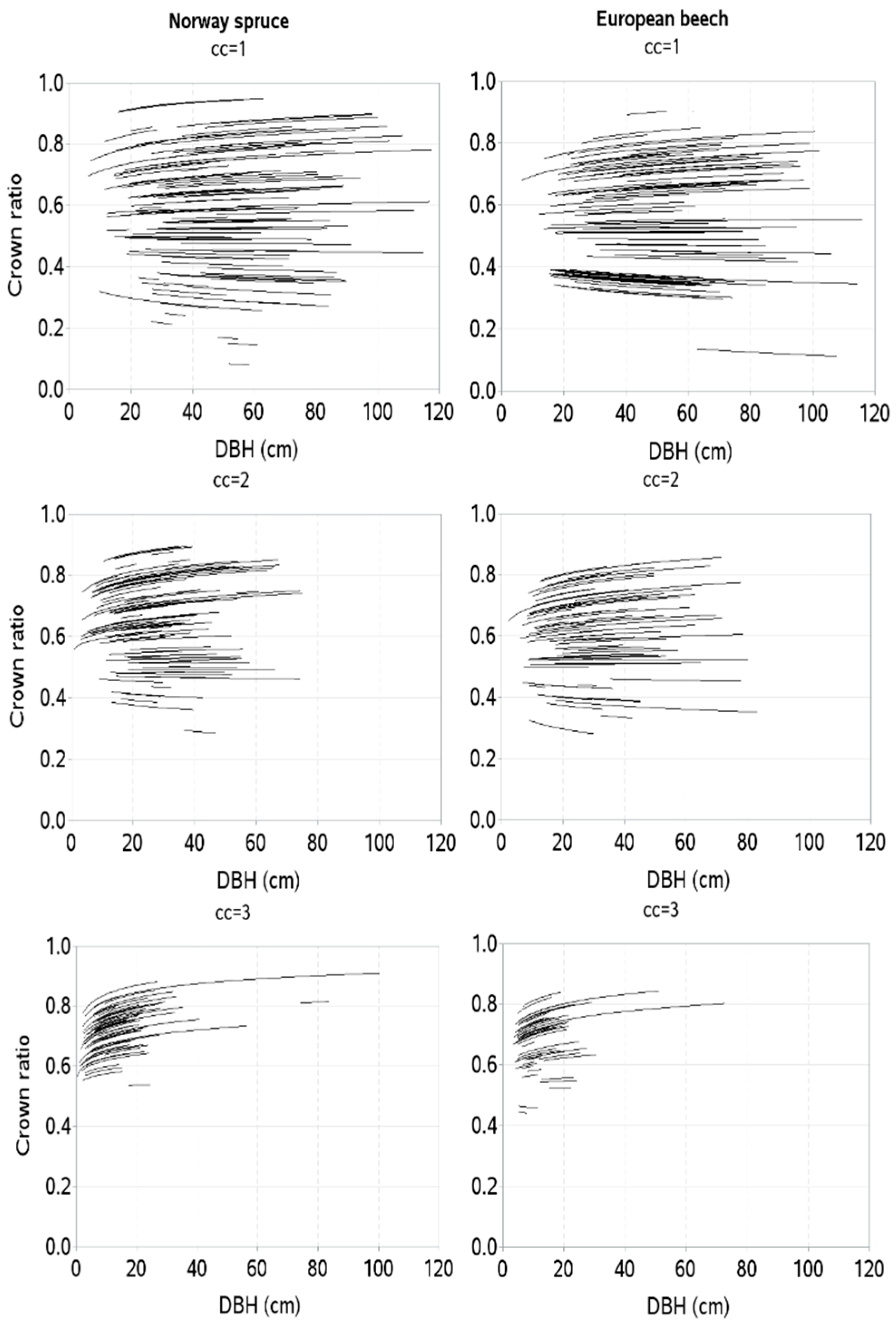{kind=link}
| Variables | Mean ± Std. (Range) | |
|---|---|---|
| Norway Spruce | European Beech | |
| Number of sample plots | 90 (18 monospecific + 72 mixed species) | 88 (18 monospecific + 70 mixed species) |
| Number of sample trees | 6736 | 7933 |
| Number of trees per sample plot | 149 ± 115 (3–430) | 142 ± 124 (3–515) |
| Number of trees per hectare (N ha−1) | 1005 ± 676 (92–2568) | 827 ± 745 (41–2568) |
| Basal area (BA, m2 ha−1) | 47.2 ± 19.4 (7.2–91.4) | 43.3 ± 14 (0.7–89.1) |
| BA proportion of a species (BAPRO) | 0.72 ± 0.31 (0.0005–1) | 0.72 ± 0.24 (0.0004–1) |
| BA of trees larger in diameter than a subject tree (BAL, m2 ha−1) | 36.7 ± 21.2 (0–88.8) | 33.9 ± 16.7 (0–86.3) |
| Quadratic mean DBH (QMD, cm) | 28.9 ± 9.6 (11.9–60.4) | 32.9 ± 11.8 (15.4–87.4) |
| DBH-to-QMD ratio (dq) | 1.2 ± 0.5 (0.1–6.8) | 1.1 ± 0.64 (0.12–5.7) |
| Arithmetic mean DBH (cm) | 24.8 ± 10.5 (9.4–53.9) | 28.6 ± 12.8 (9.4–84.4) |
| DBH sum (DBHSUM, cm) | 4845 ± 1860 (813–9333) | 4172 ± 1911 (675–9333) |
| Dominant diameter (DDOM, cm) | 49.9 ± 13.3 (18.4–77.1) | 56.7 ± 11.2 (24.7–84.3) |
| Dominant height (HDOM, m) | 27.1 ± 8.2 (8–42) | 30.6 ± 6.3 (13.6–42.8) |
| Mean height (m) | 16.2 ± 6.7 (4.6–37.4) | 19.2 ± 7.9 (6.6–42.8) |
| Relative spacing index (RSI) | 0.15 ± 0.07 (0.05–0.44) | 0.15 ± 0.06 (0.06–0.41) |
| Total height (m) | 16.9 ± 9.9 (1.5–48.7) | 19.4 ± 10.3 (1.5–50.6) |
| Diameter at breast height (DBH, cm) | 26.7 ± 18.5 (2.5–119) | 28.3 ± 20.1 (2.9–118.7) |
| Height-to-DBH ratio (HDR, m cm−1) | 0.6 ± 0.3 (0.03–6.6) | 0.8 ± 0.3 (0.1–6.1) |
| Height to crown base (HCB, m) | 6.26 ± 5.7 (0.01–37) | 8.5 ± 6.4 (0.1–35) |
| Crown diameter (CW, m) | 3.7 ± 1.6 (0.5–13.9) | 6.1 ± 3.2 (0.8–24.1) |
| Crown depth (CL, m) | 10.6 ± 6.7 (0.2–35.3) | 10.9 ± 6.4 (0.5–45.2) |
| Crown ratio (CR) | 0.65 ± 0.19 (0.05–0.99) | 0.58 ± 0.18 (0.05–0.99) |
| Model Components | Parameter Estimates | |
|---|---|---|
| Norway Spruce | European Beech | |
| Fixed | ||
| α1 | 0.186249 (0.0372) | 0.343277 (0.0233) |
| α2 | −0.10861 (0.0362) | 0.073573 (0.0233) |
| α3 | −0.2362 (0.0379) | −0.05535 (0.0245) |
| β1 | 0.025698 (0.000945) | 0.026731 (0.000564) |
| β2 | 0.002476 (0.000246) | −0.00229 (0.000218) |
| β3 | 1.106038 (0.0972) | 0.912028 (0.0576) |
| β4 | 0.048479 (0.0149) | −0.19403 (0.0156) |
| β5 | −0.09135 (0.00115) | −0.07912 (0.000764) |
| b2 | 0.194021 (0.009281) | 0.201351 (0.015073) |
| Variance | ||
| σ2ui1 | 1.9746 | 1.8083 |
| σui1ui2 | −0.19082 | −0.08725 |
| σ2ui2 | 0.01296 | 0.00957 |
| σ2 | 0.00998 | 0.00986 |
| Fit statistics | ||
| R2adj | 0.6334 | 0.7203 |
| RMSE | 0.0999 | 0.0993 |
| AIC | −28612 | −36931 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, R.P.; Vacek, Z.; Vacek, S. Generalized Nonlinear Mixed-Effects Individual Tree Crown Ratio Models for Norway Spruce and European Beech. Forests 2018, 9, 555. https://doi.org/10.3390/f9090555
Sharma RP, Vacek Z, Vacek S. Generalized Nonlinear Mixed-Effects Individual Tree Crown Ratio Models for Norway Spruce and European Beech. Forests. 2018; 9(9):555. https://doi.org/10.3390/f9090555
Chicago/Turabian StyleSharma, Ram P., Zdeněk Vacek, and Stanislav Vacek. 2018. "Generalized Nonlinear Mixed-Effects Individual Tree Crown Ratio Models for Norway Spruce and European Beech" Forests 9, no. 9: 555. https://doi.org/10.3390/f9090555
APA StyleSharma, R. P., Vacek, Z., & Vacek, S. (2018). Generalized Nonlinear Mixed-Effects Individual Tree Crown Ratio Models for Norway Spruce and European Beech. Forests, 9(9), 555. https://doi.org/10.3390/f9090555