Exogenous Oxalic Acid and Citric Acid Improve Lead (Pb) Tolerance of Larix olgensis A. Henry Seedlings

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Harvest and Seedling Culture
2.2. Pb Treatment and Organic Acid Addition
2.3. Plant Assays
2.4. Data Analysis
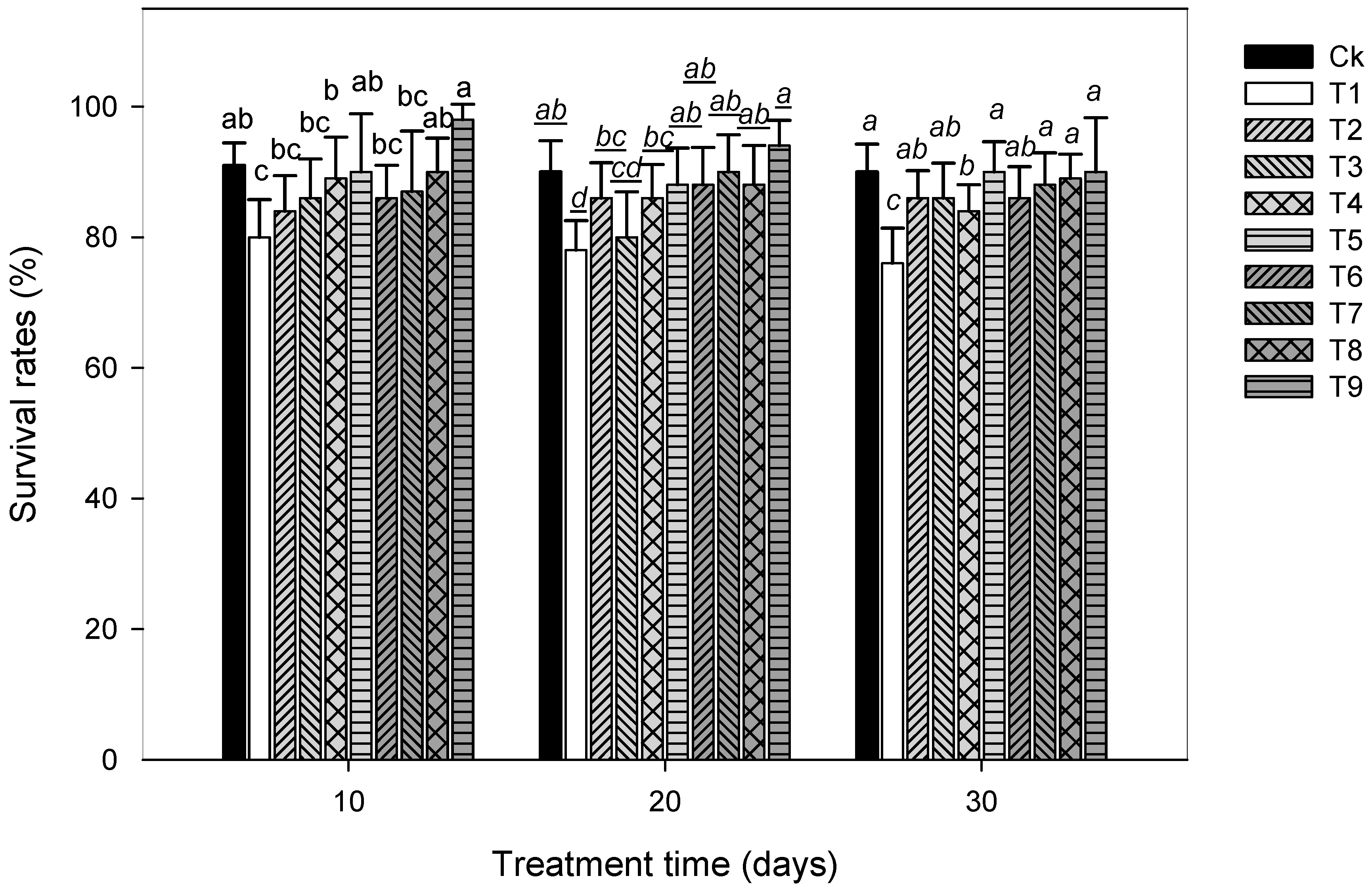
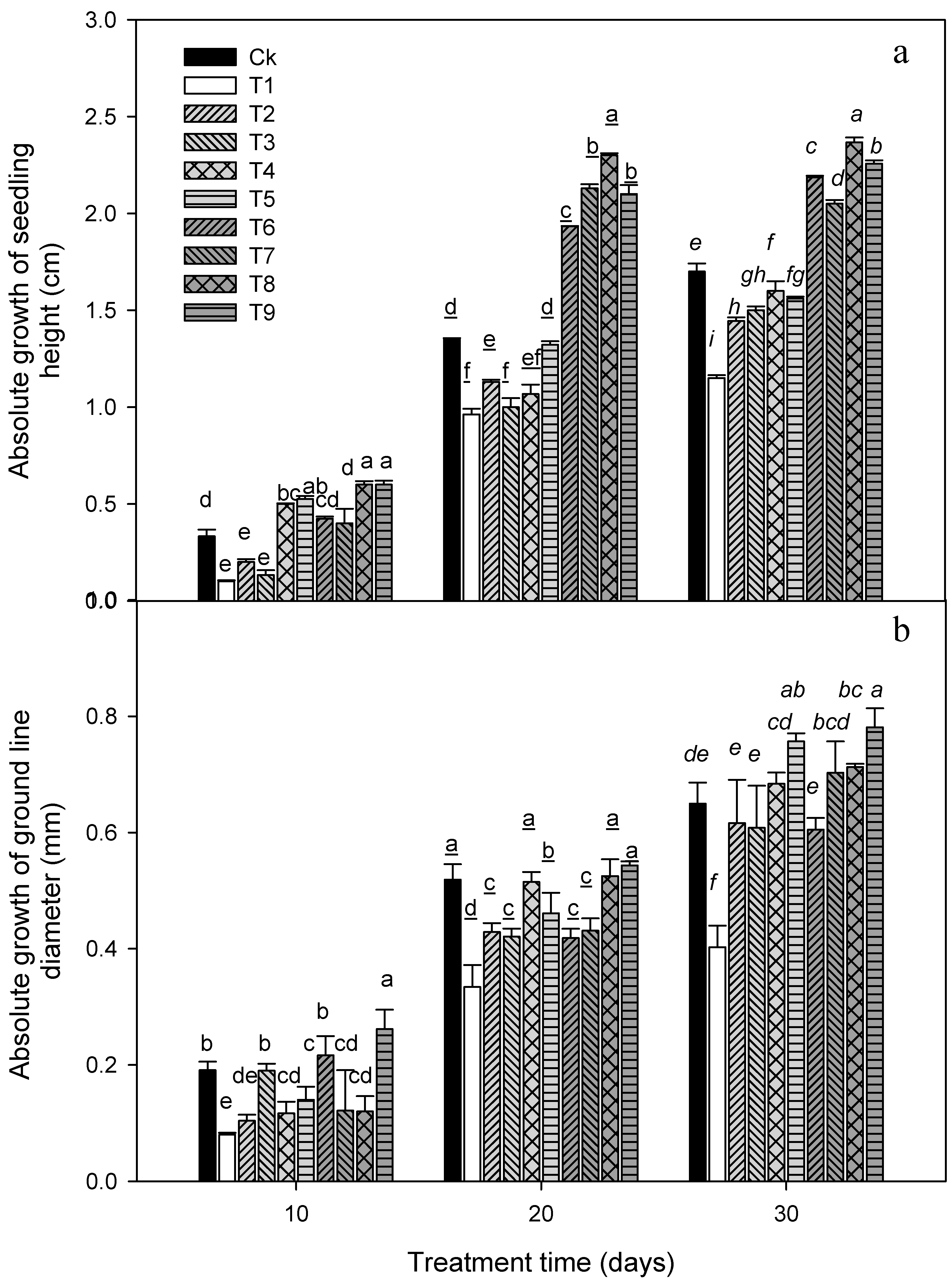
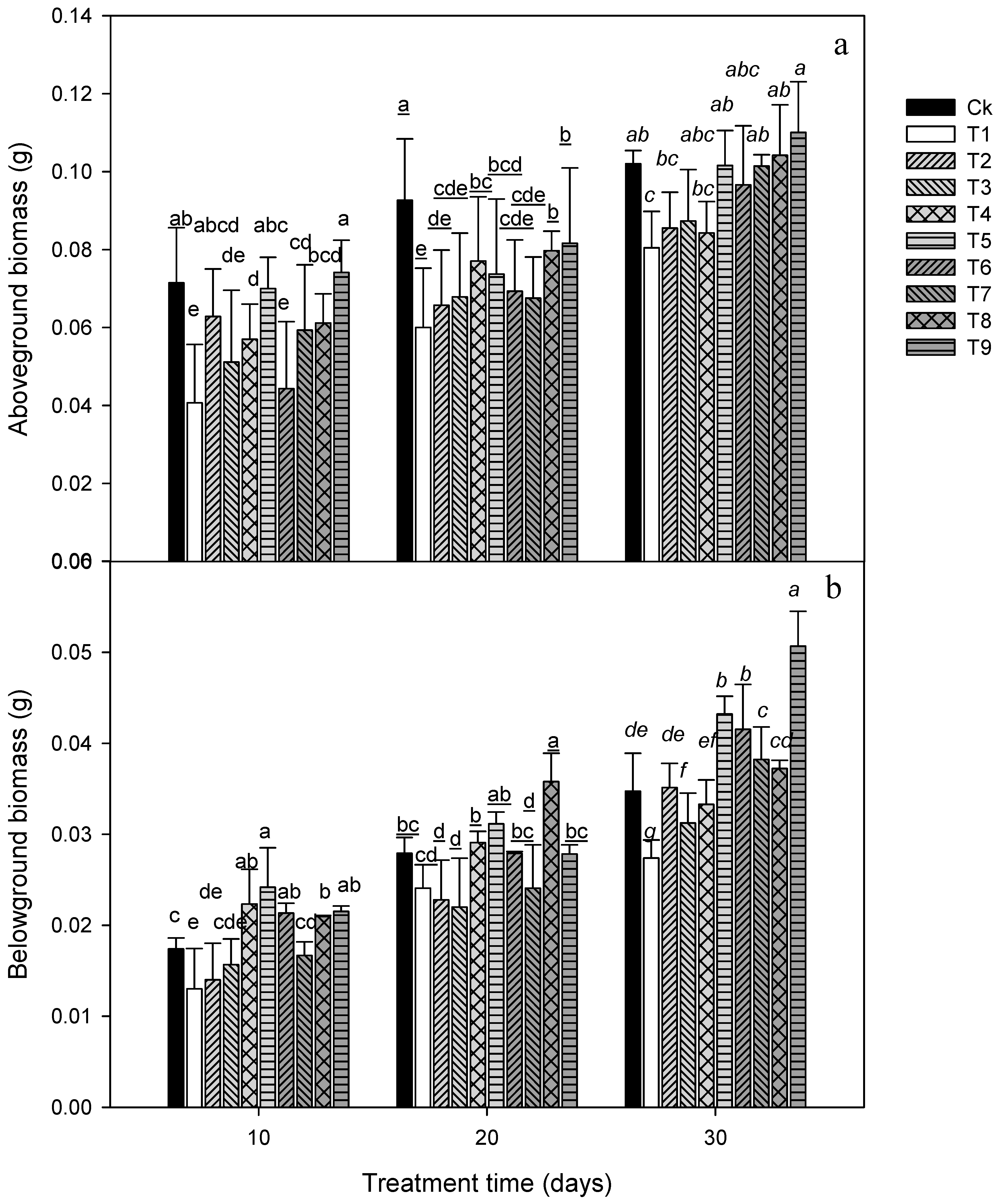
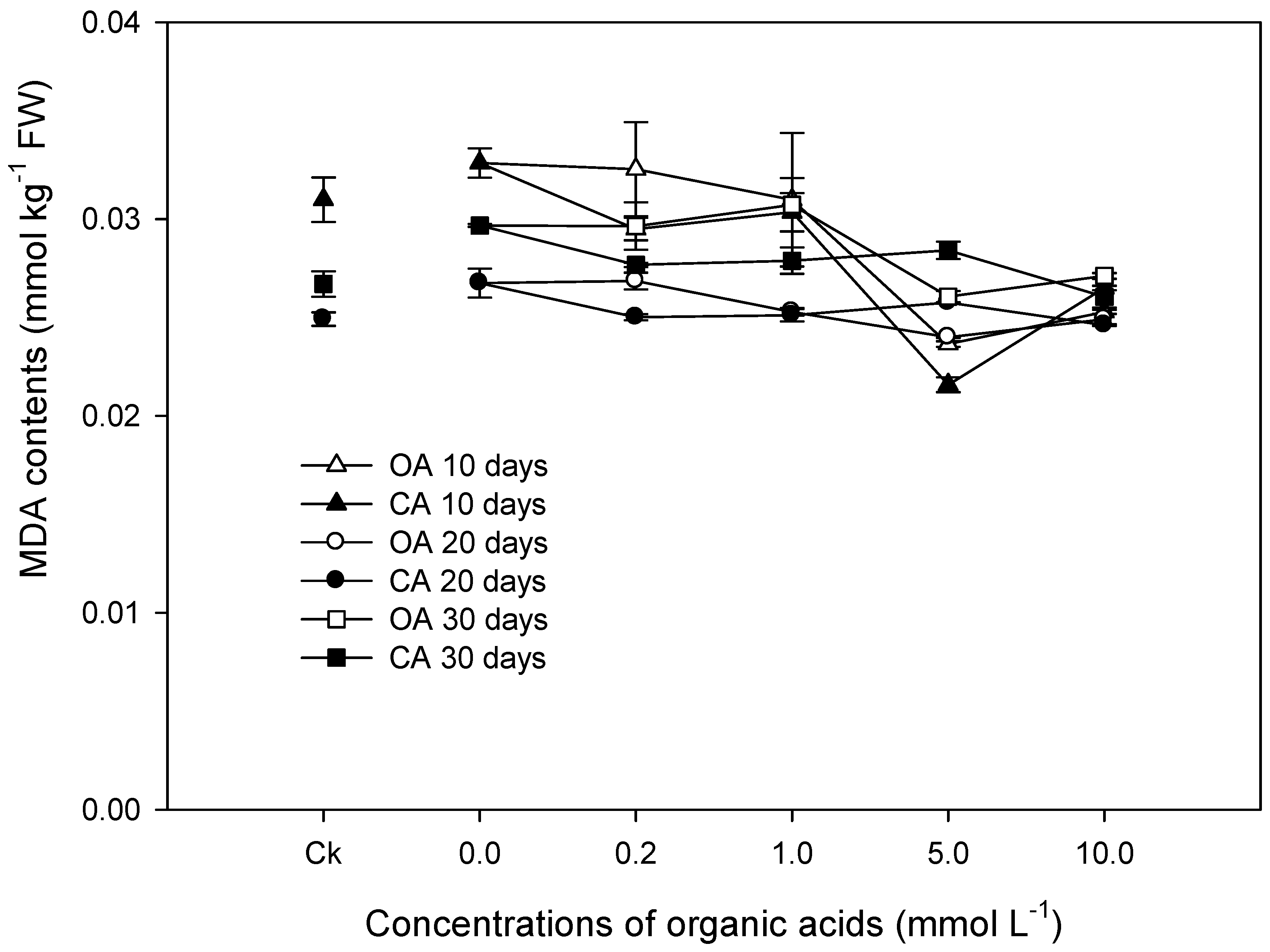
3. Results
3.1. Plant Growth Attributes
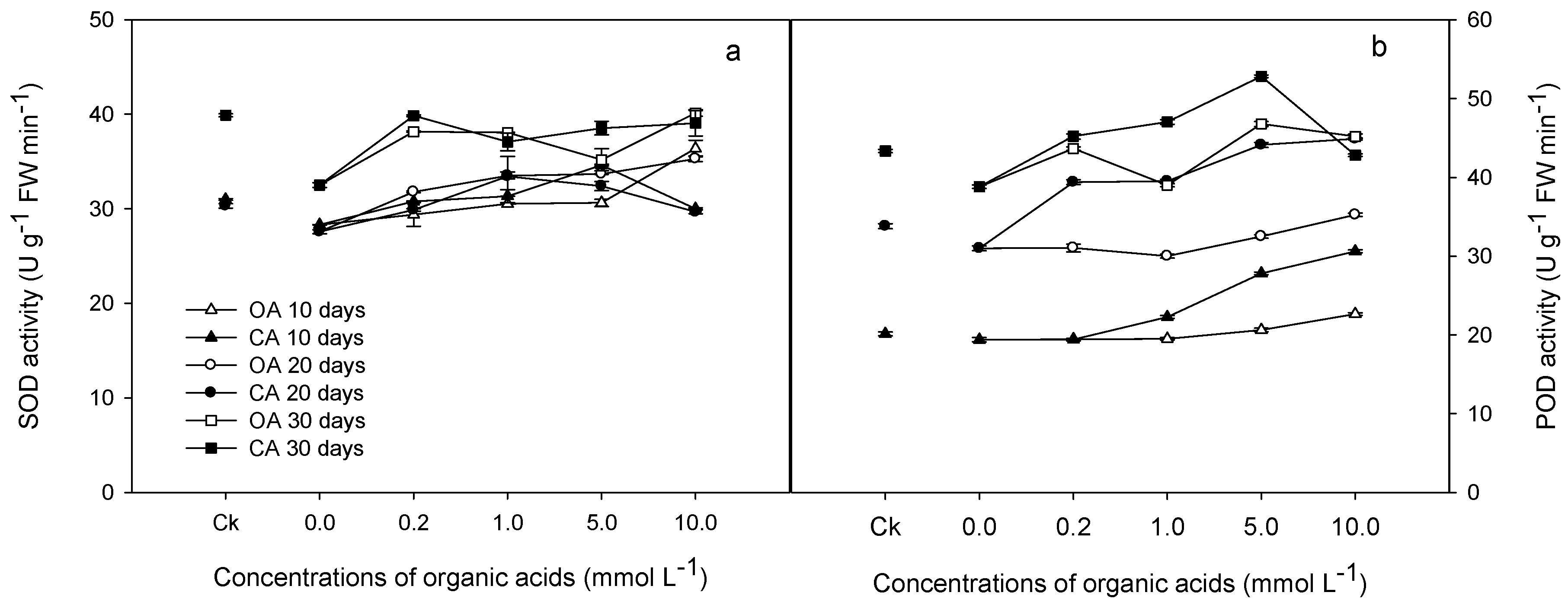
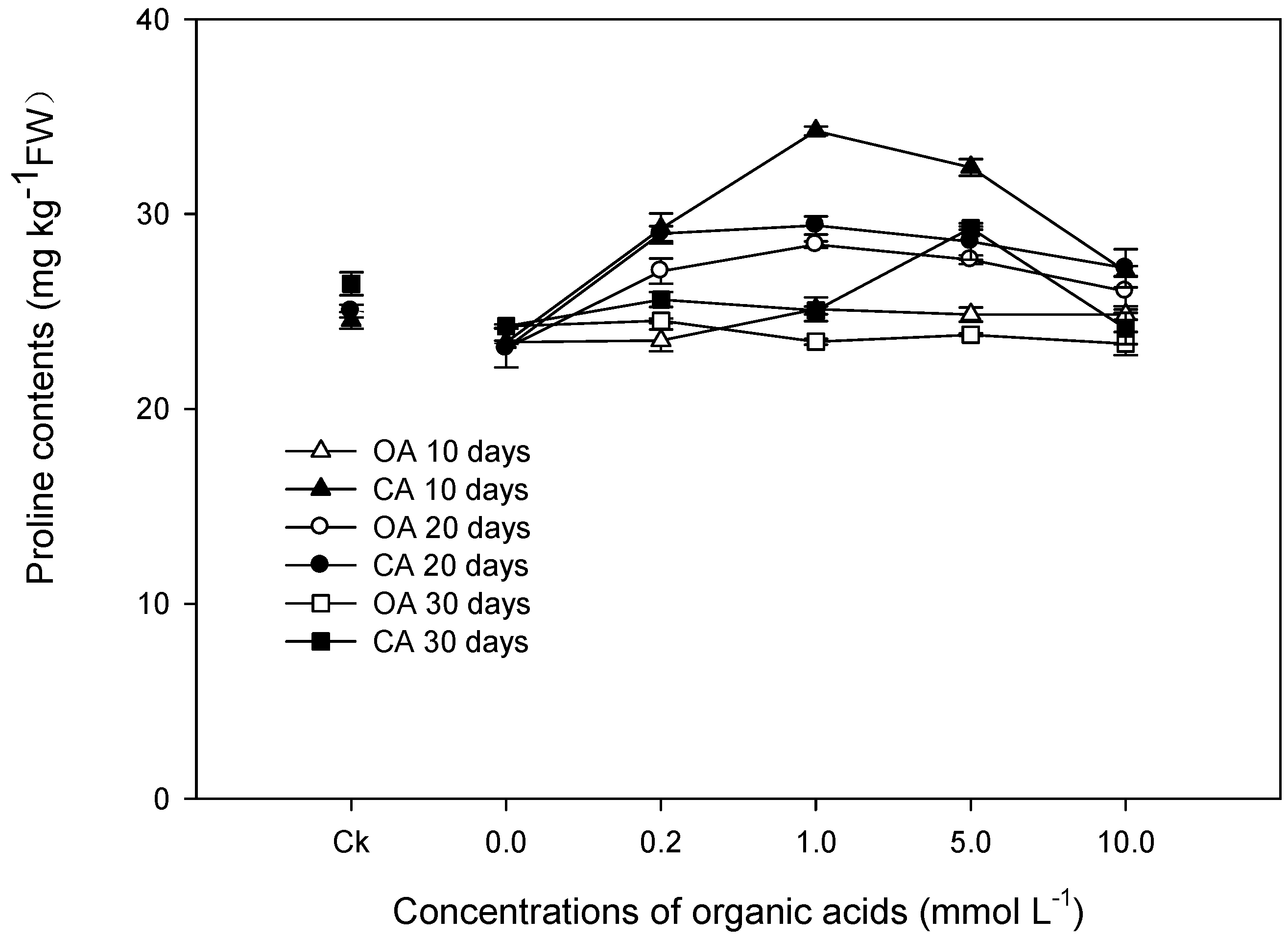
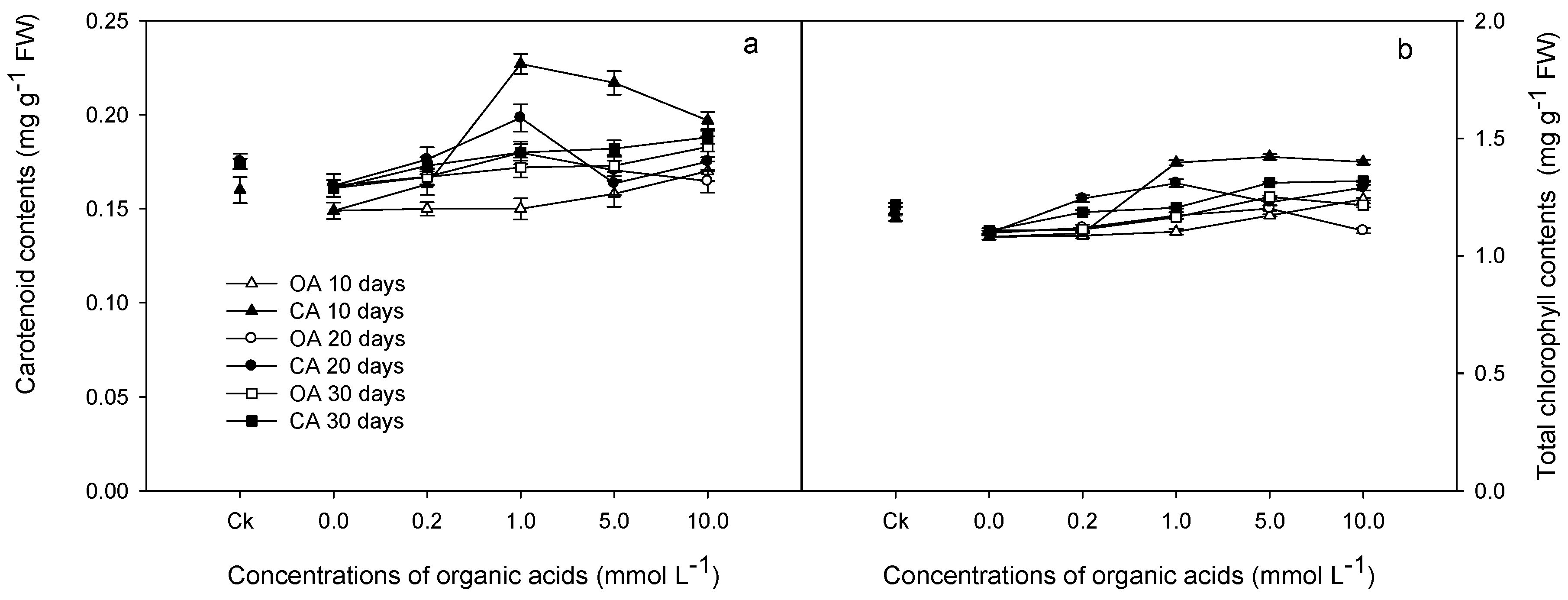
3.2. Physiological Responses
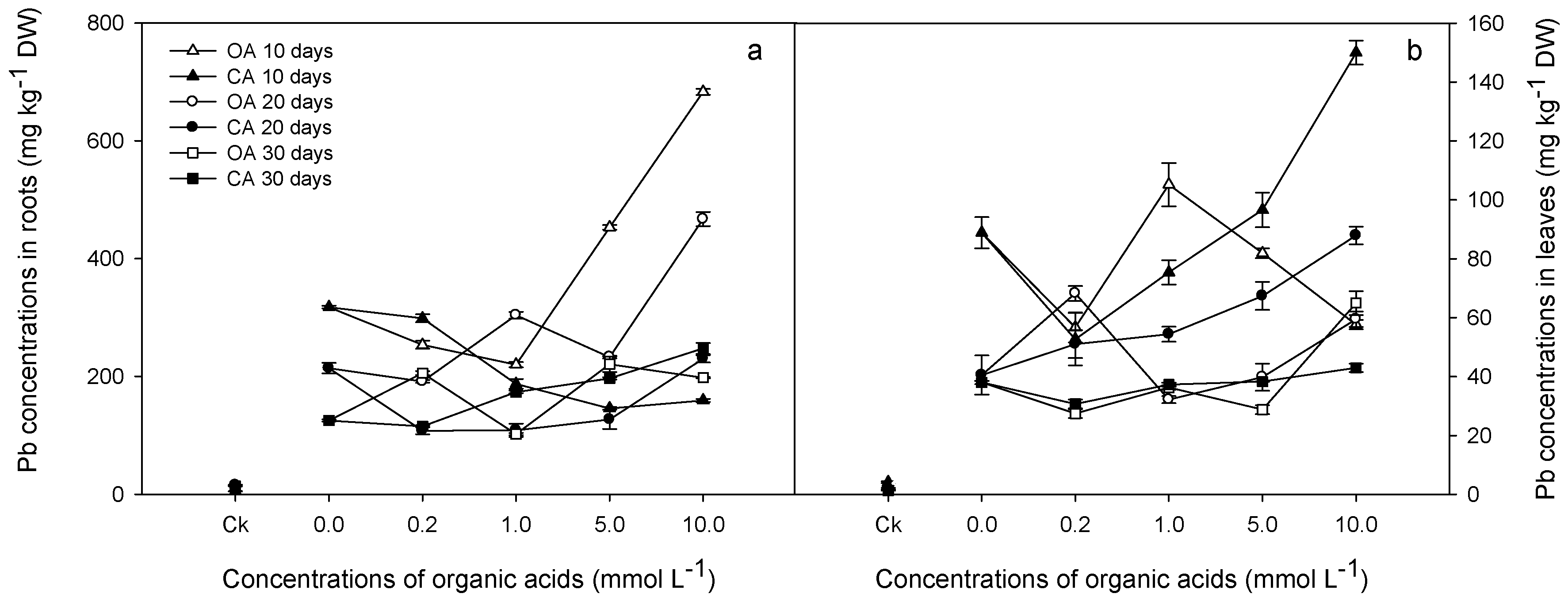
3.3. Pb Accumulation and Distribution
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kidd, P.S.; Domínguez-Rodríguez, M.J.; Díez, J.; Monterroso, C. Bioavailability and plant accumulation of heavy metals and phosphorus in agricultural soils amended by long-term application of sewage sludge. Chemosphere 2007, 66, 1458–1467. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.L.; Huang, S.Z.; Yuan, H.Y.; Zhao, J.Z.; Gu, J.G. Organic acids on the growth, anatomical structure, biochemical parameters and heavy metal accumulation of Iris lactea var. chinensis seedling growing in Pb mine tailings. Ecotoxicology 2013, 22, 1033–1042. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.Q.; Huang, S.Z.; Yu, S.L.; Gu, J.G.; Zhao, J.Z.; Han, Y.L.; Fu, J.J. The physiological response and sub-cellular location of lead and cadmium in Iris pseudacorus L. Ecotoxicology 2010, 19, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Ferrand, E.; Schreck, E.; Dumat, C. Behavior and impact of zirconium in the soil-plant system: Plant uptake and phytotoxicity. Rev. Environ. Contam. Toxicol. 2013, 221, 107–127. [Google Scholar] [PubMed]
- Pourrut, B.; Shahid, M.; Dumat, C.; Winterton, P.; Pinelli, E. Lead uptake, toxicity, and detoxification in plants. Rev. Environ. Contam. Toxicol. 2011, 213, 113–136. [Google Scholar] [PubMed]
- Zhong, Z.Y.; Lin, K.; Nora, F.Y.T. Lead stress in seedlings of Avicennia marina, a common mangrove species in South China, with and without cotyledons. Aquat. Bot. 2010, 92, 112–118. [Google Scholar]
- Yan, J.; Han, X.Z.; Ji, Z.J.; Li, Y.; Wang, E.T.; Xie, Z.H.; Chen, W.F. Abundance and diversity of sovbean-nodulating rhizobia in black soil are impacted by land use and crop management. Appl. Environ. Microb. 2014, 80, 5394–5402. (In Chinese) [Google Scholar] [CrossRef] [PubMed]
- López-Millán, A.F.; Sagardoy, R.; Solanas, M.; Abadía, A.; Abadía, J. Cadmium toxicity in tomato (Lycopersicon esculentum) plants grown in hydroponics. Environ. Exp. Bot. 2009, 65, 376–385. [Google Scholar] [CrossRef]
- Fu, X.P.; Dou, C.M.; Hu, S.P.; Chen, X.C.; Shi, J.Y.; Chen, Y.X. A review of progress in roles of organic acids on heavy metal resistance and detoxification in plants. Chin. J. Plant Ecol. 2010, 534, 1354–1358. (In Chinese) [Google Scholar]
- Hall, J.L. Cellular mechanism for heavy metal detoxification and tolerance. J. Exp. Bot. 2002, 53, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Shakoor, M.B.; Ali, S.; Hameed, A.; Farid, M.; Hussain, S.; Yasmeen, T.; Najeeb, U.; Bharwana, S.A.; Abbasi, G.H. Citric acid improves lead (Pb) phytoextraction in Brassica napus L. by mitigating Pb-induced morphological and biochemical damages. Ecotox. Environ. Saf. 2014, 109, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Li, H.Y.; Liu, Y.G.; Zeng, G.M.; Zhou, L.; Wang, X.; Wang, Y.Q.; Wang, C.L.; Hu, X.J.; Xu, W.H. Enhanced efficiency of cadmium removal by Boehmeria nivea (L.) Gaud in the presence of exogenous citric and oxalic acids. J. Environ. Sci. 2014, 26, 2508–2516. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Y.; Tong, H.Y.; Huang, S.Z.; Yuan, H.Y. Effects of citric acid and oxalic acid on the growth and physiology of Iris lactea var. Chinensis under Pb stress. Chin. J. Ecol. 2010, 29, 1340–1346. (In Chinese) [Google Scholar]
- Ruley, A.T.; Sharma, N.C.; Sahi, S.V.; Singh, S.R.; Sajwan, K.S. Effects of lead and chelators on growth, photosynthetic activity and Pb uptake in Sesbania drummondii grown in soil. Environ. Pollut. 2006, 144, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.M.; Song, Z.L.; Parr, J.F.; Wang, H.L. Occluded C in rice phytoliths: Implications to biogeochemical carbon sequestration. Plant Soil 2013, 370, 615–623. [Google Scholar] [CrossRef]
- Yang, J.Y.; Teskey, R.O.; Wang, C.K. Stem CO2 efflux of ten species in temperate forests in Northeastern China. Trees 2012, 26, 1225–1235. [Google Scholar] [CrossRef]
- Song, J.F.; Cui, X.Y. Analysis of organic acids in selected forest litters of Northeast China. J. For. Res. 2003, 14, 285–289. [Google Scholar]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2006; World Soil Resource Reports 103; FAO: Rome, Italy, 2006; pp. 53–65. [Google Scholar]
- Hu, R.; Sun, K.; Su, X.; Pan, Y.X.; Zhang, Y.F.; Wang, X.P. Physiological responses and tolerance mechanisms to Pb in two xerophils: Salsola passerina Bunge and Chenopodium album L. J. Hazard. Mater. 2012, 205–206, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Song, J.F.; Cui, X.Y. Sorption and biodegradation behavior of organic acids with low molecular weight in dark brown forest soils. Chin. J. Soil Sci. 2007, 38, 1118–1122. (In Chinese) [Google Scholar]
- Li, H.S.; Sun, Q.; Zhao, S.J.; Zhang, W.H. Principles and Techniques of Plant Physiological Biochemical Experiment; Higher Education Press: Beijing, China, 2000; pp. 134–263. (In Chinese) [Google Scholar]
- Song, J.F.; Ma, R.; Huang, W.B.; Yang, D.; Cui, X.Y. Exogenous organic acids protect Changbai Larch (Larix olgensis) seedlings against cadmium toxicity. Fresen. Environ. Bull. 2014, 23, 3459–3467. [Google Scholar]
- Song, J.F.; Cui, X.Y.; Wang, Z.Q. Effects of low molecular-weight organic acids on P, Fe and K availability of dark brown forest soils and absorption of forest seedlings. J. Soil Water Conserv. 2011, 25, 123–127. (In Chinese) [Google Scholar]
- Pang, X.Q.; Cui, X.J.; Sun, Z.F.; Qi, Z.X. Application of Foliar fertilizer Potassium dihydrogen phosphate in culture of larch seedlings. Pr. For. Technol. 2010, 3, 22. (In Chinese) [Google Scholar]
- Xiao, Y.; Tang, Y.K.; Cao, Y.P.; Wang, J.G. Application of surfactants in fertilizer for foliage dressing and its progress. Phosphate Compd. Fert. 2003, 18, 14–16. (In Chinese) [Google Scholar]
- Song, J.F.; Yang, J.Y.; Cui, X.Y. Effects of low molecular-weight organic acids/salts on availability of lead, zinc and arsenic in mixed metal-polluted soil. J. Soil Water Conserv. 2010, 24, 108–112, 118. (In Chinese) [Google Scholar]
- Shu, X.; Yin, L.Y.; Zhang, Q.F.; Wang, W.B. Effect of Pb toxicity on leaf growth, antioxidant enzyme activities, and photosynthesis in cuttings and seedlings of Jatropha curcas L. Environ. Sci. Pollut. Res. 2012, 19, 893–902. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Huang, R.D.; Cao, Z.Q. Effects of Pb stress on the physiological and biochemical traits of maize. J. Maize Sci. 2005, 13, 61–64. [Google Scholar]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Najeeb, U.; Jilani, G.; Ali, S.; Sarwar, M.; Xu, L.; Zhou, W.J. Insight into cadmium induced physiological and ultra-structural disorders in Juncus effusus L. and its remediation through exogenous citric acid. J. Hazard. Mater. 2011, 186, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.R.; Xie, Y.; Jin, G.F.; Fu, J.M.; Li, H.Y. Growth responses of two tall fescue cultivars to Pb stress and their metal accumulation characteristics. Ecotoxicology 2015, 24, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.Y.; Dong, Y.J.; Kong, J.; Xu, L.L.; Liu, S. Effects of application of salicylic acid alleviates cadmium toxicity in perennial ryegrass. Plant Growth Regul. 2015, 75, 695–706. [Google Scholar] [CrossRef]
- Singh, I.; Shah, K. Evidences for suppression of cadmium induced oxidative stress in presence of sulphosalicylic acid in rice seedlings. Plant Growth Regul. 2015, 76, 99–110. [Google Scholar] [CrossRef]
- Fan, H.L.; Hong, W.; Wu, C.Z.; Chen, C.; Li, J. Effects of water stress on the physiological indexes of carbon and nitrogen metabolization of Liriope muscari (Decne.) Bailey. J. Fujian Agric. For. Univ. (Nat. Sci. Ed.) 2012, 41, 454–458. (In Chinese) [Google Scholar]
- Ali, E.; Maodzeka, A.; Hussain, N.; Shamsi, I.H.; Jiang, L.X. The alleviation of cadmium toxicity in oilseed rape (Brassica napus) by the application of salicylic acid. Plant Growth Regul. 2015, 75, 641–655. [Google Scholar] [CrossRef]
- Metwally, A.; Finkermeier, I.; Georgi, M.; Dietz, K.J. Salicylic acid alleviates the cadmium toxicity in barley seedlings. Plant Physiol. 2003, 132, 272–281. [Google Scholar] [CrossRef] [PubMed]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, J.; Markewitz, D.; Wu, S.; Sang, Y.; Duan, C.; Cui, X. Exogenous Oxalic Acid and Citric Acid Improve Lead (Pb) Tolerance of Larix olgensis A. Henry Seedlings. Forests 2018, 9, 510. https://doi.org/10.3390/f9090510
Song J, Markewitz D, Wu S, Sang Y, Duan C, Cui X. Exogenous Oxalic Acid and Citric Acid Improve Lead (Pb) Tolerance of Larix olgensis A. Henry Seedlings. Forests. 2018; 9(9):510. https://doi.org/10.3390/f9090510
Chicago/Turabian StyleSong, JinFeng, Daniel Markewitz, Shaoping Wu, Ying Sang, Chengwei Duan, and XiaoYang Cui. 2018. "Exogenous Oxalic Acid and Citric Acid Improve Lead (Pb) Tolerance of Larix olgensis A. Henry Seedlings" Forests 9, no. 9: 510. https://doi.org/10.3390/f9090510
APA StyleSong, J., Markewitz, D., Wu, S., Sang, Y., Duan, C., & Cui, X. (2018). Exogenous Oxalic Acid and Citric Acid Improve Lead (Pb) Tolerance of Larix olgensis A. Henry Seedlings. Forests, 9(9), 510. https://doi.org/10.3390/f9090510