Ecological Factors Affecting White Pine, Red Oak, Bitternut Hickory and Black Walnut Underplanting Success in a Northern Temperate Post-Agricultural Forest


 ,
,
Abstract

1. Introduction
2. Materials and Methods
2.1. Study Site Description
2.2. Experimental Design
2.3. Measurements of Survival, Tree Growth and Deer Browsing
2.4. Measurements of Shelterwood Characteristics
2.4.1. Overstory Structure and Composition
2.4.2. Relative Cover of Understory Vegetation
2.4.3. Soil Characteristics
2.5. Statistical Analyses
2.5.1. Red Oak and White Pine Data
2.5.2. Bitternut Hickory and Black Walnut Data
3. Results
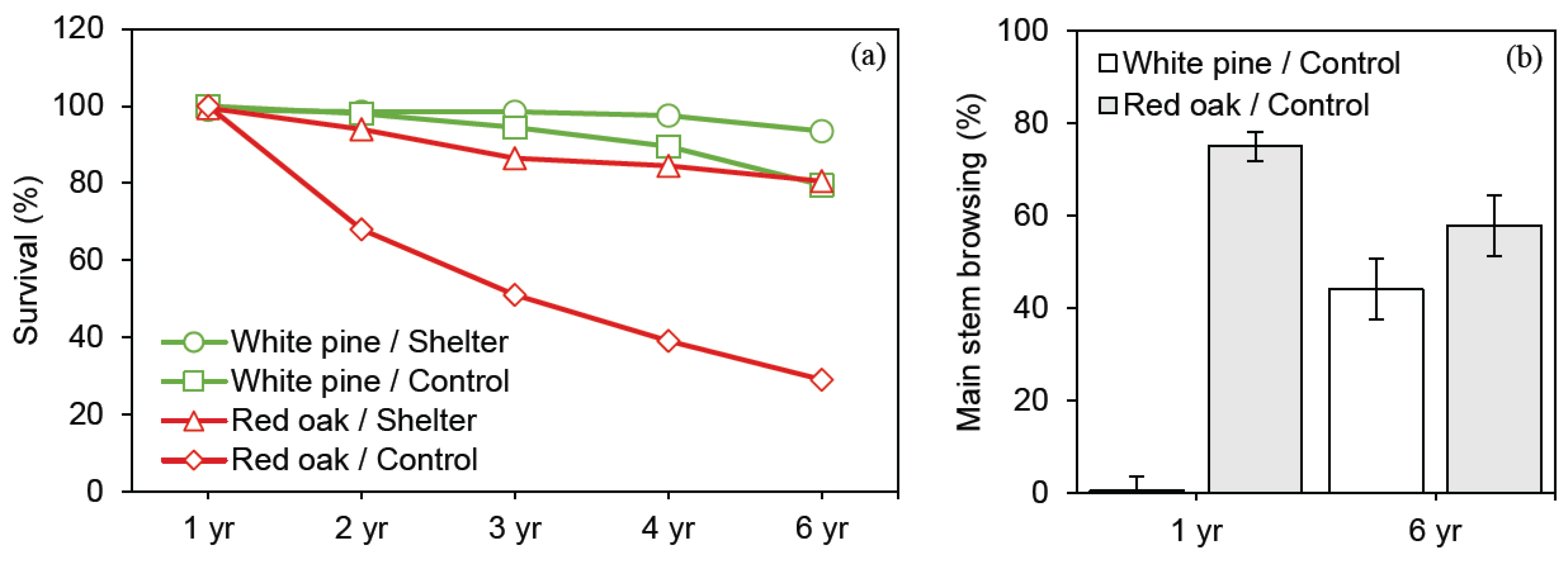
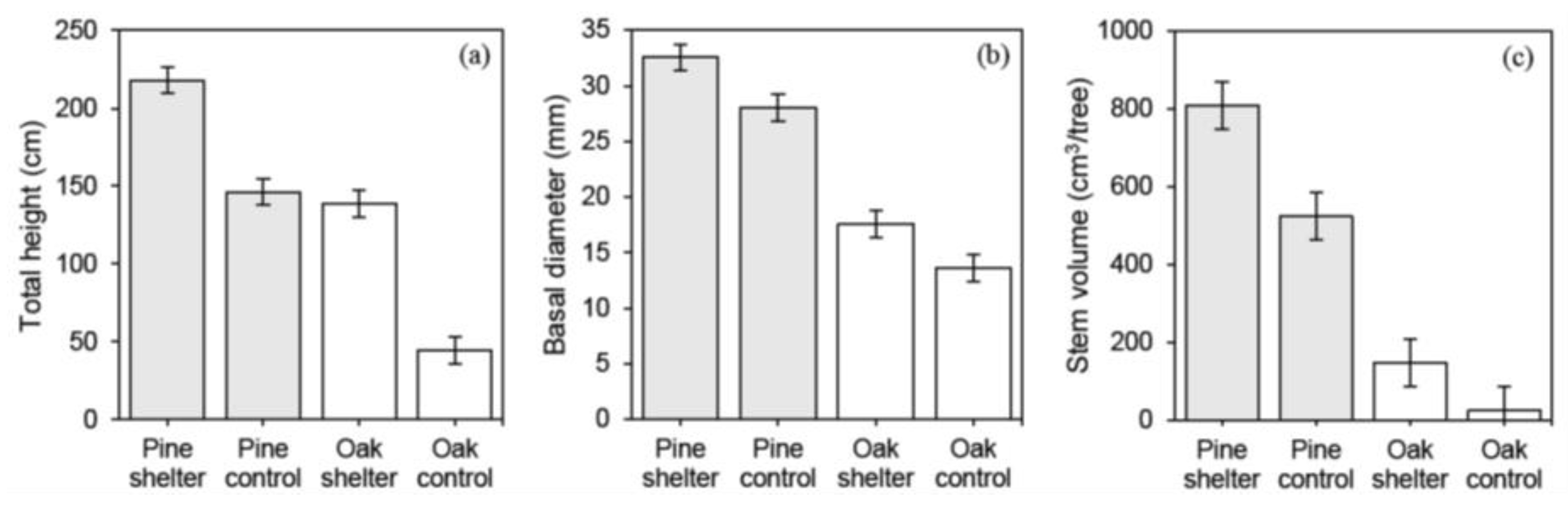
3.1. Effect of Tree Species and Deer Protection Treatments on Survival and Growth
3.2. Effect of the Forest Community Type on Ecological Variables, and on Tree Survival and Growth
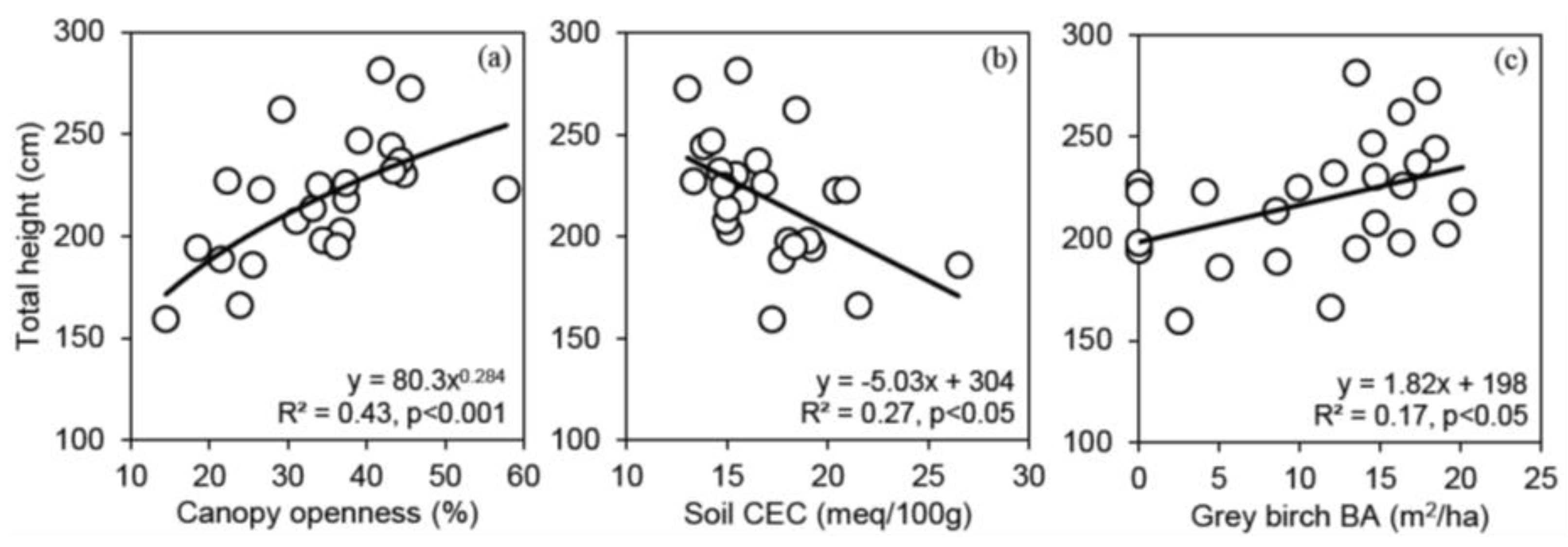
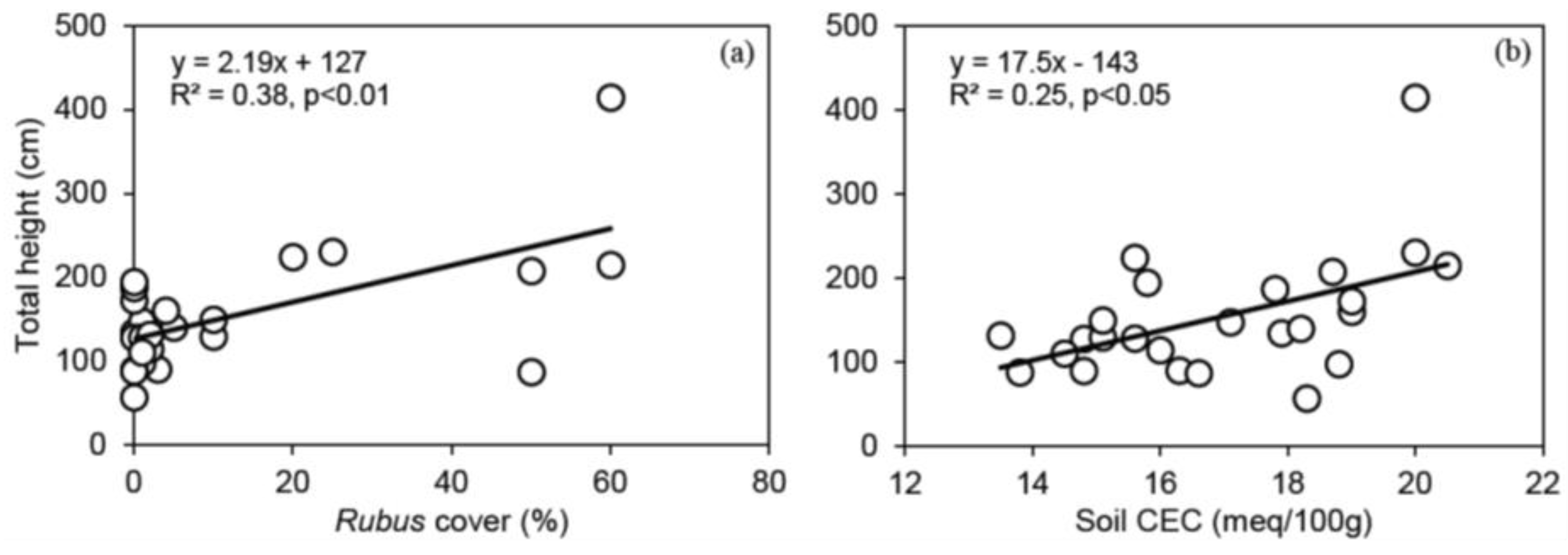
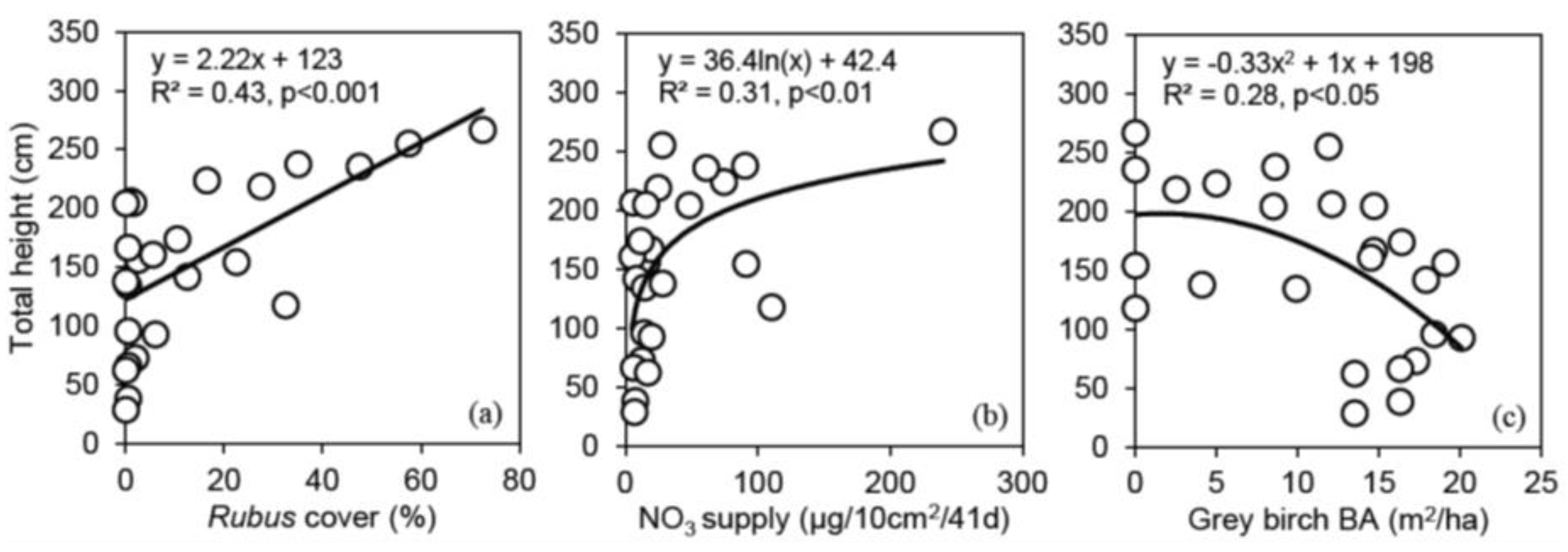
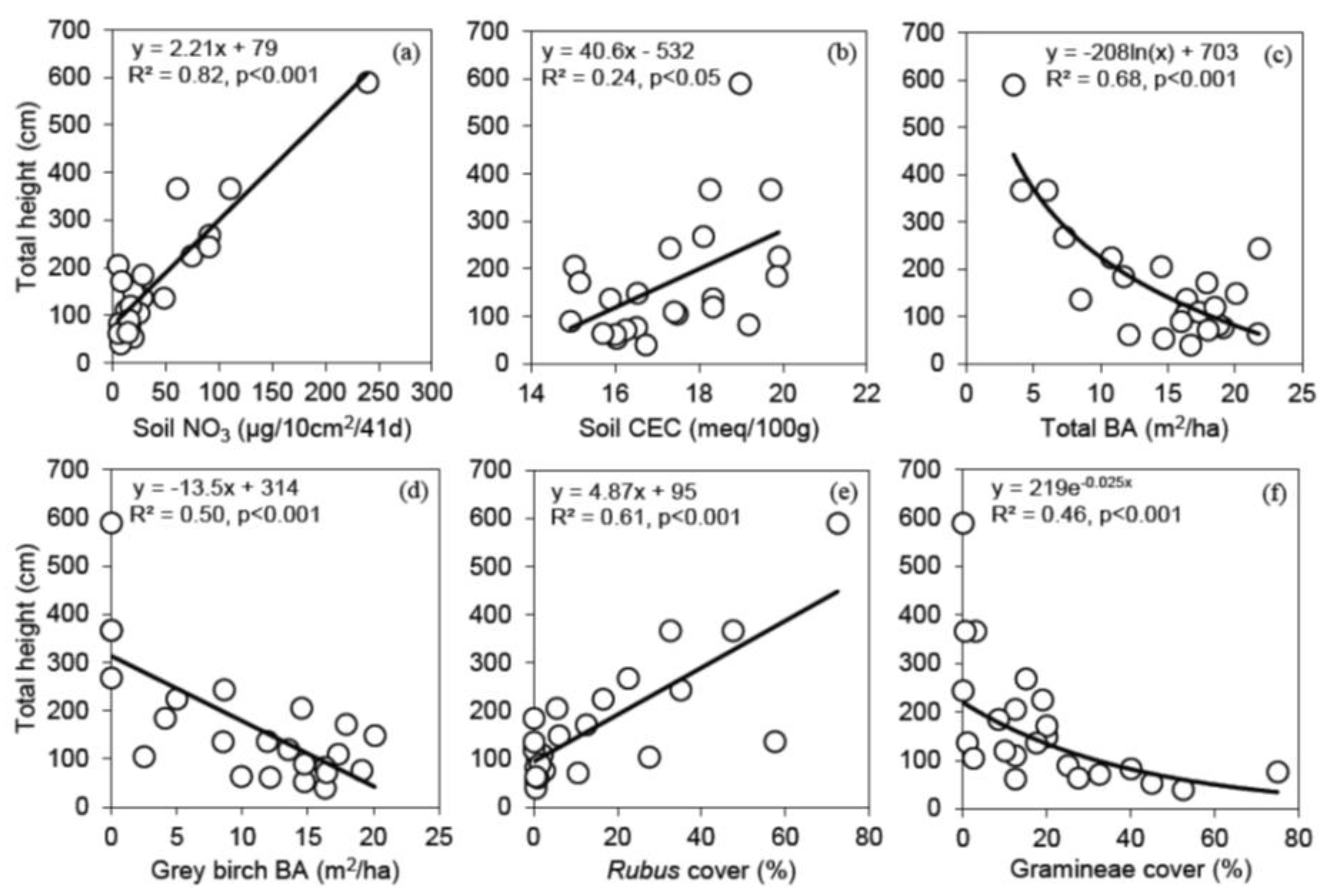
3.3. Relationships between Shelterwood Characteristics and Total Height after 6 Years for Red Oak, White Pine, Bitternut Hickory and Black Walnut Protected with Tree Shelters
4. Discussion
4.1. Deer Impact on Red Oak and White Pine Survival and Growth
4.2. The Effect of Shelterwood Characteristics on Underplanted Tree Growth
4.3. Management Implications for Forest Restoration and Hardwood Species Migration
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Variable 1 | Min. | Max. | Median | Mean | Std. dev. |
|---|---|---|---|---|---|---|
| Soil | pH | 4.7 | 5.6 | 5.3 | 5.3 | 0.2 |
| CEC (meq/100 g) | 14.3 | 19.9 | 16.7 | 17.1 | 1.7 | |
| Base saturation (%) | 31.0 | 58.7 | 45.4 | 45.3 | 8.1 | |
| NO3 (µg/10 cm2/41d) | 4.9 | 239.6 | 16.7 | 38.8 | 51.8 | |
| NH4 (µg/10 cm2/41d) | 3.0 | 39.0 | 4.4 | 5.9 | 7.0 | |
| P (µg/10 cm2/41d) | 1.1 | 8.7 | 2.8 | 3.1 | 1.7 | |
| K (µg/10 cm2/41d) | 7.4 | 92.4 | 26.4 | 34.1 | 22.3 | |
| Ca (µg/10 cm2/41d) | 1561 | 2384 | 2095 | 2040 | 228 | |
| Mg (µg/10 cm2/41d) | 286 | 567 | 432 | 430 | 64 | |
| S (µg/10 cm2/41d) | 17 | 128 | 55 | 58 | 27 | |
| Organic matter (%) | 6.9 | 13.4 | 10.6 | 10.3 | 1.6 | |
| C/N | 9.4 | 12.0 | 10.8 | 10.8 | 0.7 | |
| Clay (%) | 0 | 62 | 26 | 30 | 13 | |
| Silt (%) | 18 | 83 | 44 | 44 | 11 | |
| Sand (%) | 0 | 58 | 29 | 26 | 11 | |
| Stand | Canopy openness (%) | 20.8 | 46.2 | 34.0 | 32.7 | 6.9 |
| structure and | Total BA (m2/ha) | 3.5 | 21.8 | 16.3 | 14.5 | 5.3 |
| composition | Betula pop. BA (m2/ha) | 0 | 20.1 | 13.5 | 11.0 | 6.7 |
| Ulmus am. BA (m2/ha) | 0 | 11.8 | 0.0 | 1.2 | 3.1 | |
| Fraxinus am. BA (m2/ha) | 0 | 10.8 | 0.0 | 1.0 | 2.6 | |
| Understory | Rubus spp. (% cover) | 0 | 73 | 3 | 14 | 20 |
| plant cover | Gramineae spp. (% cover) | 0 | 75 | 15 | 19 | 18 |
| Solidago spp. (% cover) | 8 | 80 | 38 | 38 | 21 | |
| Fragaria virginiana (% cover) | 1 | 45 | 18 | 20 | 13 | |
| Carex spp. (% cover) | 0 | 28 | 2 | 5 | 7 | |
| Phalaris arundinacea (% cover) | 0 | 33 | 5 | 9 | 10 | |
| Onoclea sensibilis (% cover) | 0 | 16 | 0 | 3 | 4 |
| Red oak | Total height | Rubus cover | Soil CEC | ||||
| Total height | 1.00 | 0.61 | 0.50 | ||||
| Rubus cover | 0.61 | 1.00 | 0.49 | ||||
| Soil CEC | 0.50 | 0.49 | 1.00 | ||||
| White pine | Total height | Canopy openness | Soil CEC | Grey birch BA | |||
| Total height | 1.00 | 0.60 | −0.52 | 0.41 | |||
| Canopy openness | 0.60 | 1.00 | −0.29 | 0.49 | |||
| Soil CEC | −0.52 | −0.29 | 1.00 | −0.40 | |||
| Grey birch BA | 0.41 | 0.49 | −0.40 | 1.00 | |||
| Bitternut hickory | Total height | Rubus cover | Soil NO3 | Grey birch BA | |||
| Total height | 1.00 | 0.65 | 0.48 | −0.50 | |||
| Rubus cover | 0.65 | 1.00 | 0.76 | −0.62 | |||
| Soil NO3 | 0.48 | 0.76 | 1.00 | −0.70 | |||
| Grey birch BA | −0.50 | −0.62 | −0.70 | 1.00 | |||
| Black walnut | Total height | Soil NO3 | Soil CEC | Total BA | Grey birch BA | Rubus cover | Gramineae cover |
| Total height | 1.00 | 0.91 | 0.49 | −0.72 | −0.71 | 0.78 | −0.57 |
| Soil NO3 | 0.91 | 1.00 | 0.45 | −0.67 | −0.69 | 0.75 | −0.43 |
| Soil CEC | 0.49 | 0.45 | 1.00 | −0.42 | −0.59 | 0.46 | −0.34 |
| Total BA | −0.72 | −0.67 | −0.42 | 1.00 | 0.76 | −0.48 | 0.40 |
| Grey birch BA | −0.71 | −0.69 | −0.59 | 0.76 | 1.00 | −0.60 | 0.61 |
| Rubus cover | 0.78 | 0.75 | 0.46 | −0.48 | −0.60 | 1.00 | −0.57 |
| Gramineae cover | −0.57 | −0.43 | −0.34 | 0.40 | 0.61 | −0.57 | 1.00 |
References
- Burns, R.M.; Honkala, B.H. Silvics of North America: Vol. 1 Conifers; Forest Service Agriculture (USDA): Washington, DC, USA, 1990; Agriculture Handbook; p. 654.
- Fralish, J.S. The Keystone Role of Oak and Hickory in the Central Hardwood Forest; Gen. Tech. Rep. SRS-73; U.S. Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2004; pp. 78–87.
- Ward, J.S.; Mervosh, T.L. Strategies to reduce browse damage on eastern white pine (Pinus strobus) in southern New England, USA. For. Ecol. Manag. 2008, 255, 1559–1567. [Google Scholar] [CrossRef]
- Booth, J.D. Timber utilization on the agricultural frontier in southern Québec. J. East. Townsh. Stud. 1994, 4, 15–30. [Google Scholar]
- Simard, H.; Bouchard, A. The precolonial 19th century forest of the Upper St. Lawrence Region of Quebec; a record of its exploitation and transformation through notary deeds of wood sales. Can. J. For. Res. 1996, 26, 1670–1676. [Google Scholar] [CrossRef]
- Tanguay, C. Distribution, Abondance et état de Santé du Noyer cEndré (Juglans cinerea) en Relation avec les Gradients écologiques dans les Cantons-de-l’Est. Master’s Thesis, Université du Québec à Montréal, Montréal, QC, Canada, 2011. [Google Scholar]
- Abrams, M.D. Fire and the development of oak forests. BioSc. 1992, 42, 346–353. [Google Scholar] [CrossRef]
- Weyenberg, S.A.; Frelich, L.E.; Reich, P.B. Logging versus fire: How does disturbance type influence the abundance of Pinus strobus regeneration? Silva Fenn. 2004, 38, 179–194. [Google Scholar] [CrossRef]
- Lefland, A.B.; Duguid, M.C.; Morin, R.S.; Ashton, M.S. The demographics and regeneration dynamic of hickory in second-growth temperate forest. For. Ecol. Manag. 2018, 419–420, 187–196. [Google Scholar] [CrossRef]
- Côté, S.D.; Rooney, T.P.; Tremblay, J.-P.; Dussault, C.; Waller, D.M. Ecological impacts of deer overabundance. Ann. Rev. Ecol. Evol. Syst. 2004, 35, 113–147. [Google Scholar] [CrossRef]
- Kittredge, D.B.; Ashton, P.M.S. Impact of deer browsing on regeneration in mixed stands in southern New England. North. J. Appl. For. 1995, 12, 115–120. [Google Scholar]
- White, M.A. Long-term effects of deer browsing: Composition, structure and productivity in a northeastern Minnesota old-growth forest. For. Ecol. Manag. 2012, 269, 222–228. [Google Scholar] [CrossRef]
- Horsley, S.B.; Stout, S.L.; deCalesta, D.S. White-tailed deer impact on the vegetation dynamics of a northern hardwood forest. Ecol. Appl. 2003, 13, 98–118. [Google Scholar] [CrossRef]
- Ouranos. Vers l’adaptation. Synthèse des connaissances sur les changements climatiques au Québec. Partie 1: Évolution climatique au Québec; Ouranos: Montréal, QC, Canada, 2015. [Google Scholar]
- Niinemets, Ü.; Valladares, F. Tolerance to shade, drought and waterlogging of temperate northern hemisphere trees and shrubs. Ecol. Monogr. 2006, 76, 521–547. [Google Scholar] [CrossRef]
- Lafleur, B.; Paré, D.; Munson, A.D.; Bergeron, Y. Response of northeastern North American forests to climate change: Will soil conditions constrain tree species migration? Environ. Rev. 2010, 18, 279–289. [Google Scholar] [CrossRef]
- Cogliastro, A.; Gagnon, D.; Bouchard, A. Experimental determination of soil characteristics optimal for the growth of ten hardwoods planted on abandoned farmland. For. Ecol. Manag. 1997, 96, 49–63. [Google Scholar] [CrossRef]
- von Althen, F.W. Afforestation of former farmland with high-value hardwoods. For. Chron. 1991, 67, 209–212. [Google Scholar] [CrossRef]
- Peichl, M.; Arain, M.A. Above- and belowground ecosystem biomass and carbon pools in an age-sequence of temperate pine plantation forests. Agric. For. Meteorol. 2006, 140, 51–63. [Google Scholar] [CrossRef]
- Neumann, P.D.; Krahn, H.J.; Krogman, N.T.; Thomas, B.R. ‘My grandfather would roll over in his grave’: Family farming and tree plantation on farmland. Rural Sociol. 2007, 72, 111–135. [Google Scholar] [CrossRef]
- Hatch, A.B. The role of mycorrhizae in afforestation. J. For. 1936, 34, 22–29. [Google Scholar]
- Ostfeld, R.S.; Canham, C.D. Effects of meadow vole population density on tree seedling survival in old fields. Ecol. 1993, 74, 1792–1801. [Google Scholar] [CrossRef]
- Margolis, H.A.; Brand, D.G. An ecophysiological basis for understanding plantation establishment. Can. J. For. Res. 1990, 20, 375–390. [Google Scholar] [CrossRef]
- Laungani, R.; Knops, J.M.H. Species-driven changes in nitrogen cycling can provide a mechanism for plant invasions. PNAS 2009, 106, 12400–12405. [Google Scholar] [CrossRef] [PubMed]
- Ostry, M.E.; Laflamme, G.; Katovich, S.A. Silvicultural approaches for management of eastern white pine to minimize impacts of damaging agents. For. Pathol. 2010, 40, 332–346. [Google Scholar] [CrossRef]
- Gardiner, E.S.; Stanturf, J.A.; Schweitzer, C.J. An afforestation system for restoring bottomland hardwood forests: Biomass accumulation of nuttall oak seedlings interplanted beneath eastern cottonwood. Rest. Ecol. 2004, 12, 525–532. [Google Scholar] [CrossRef]
- Truax, B.; Lambert, F.; Gagnon, D. Herbicide-free plantations of oaks and ashes along a gradient of open to forested mesic environments. For. Ecol. Manag. 2000, 137, 155–169. [Google Scholar] [CrossRef]
- Williston, H.L.; Huckenpahler, B.J. Hardwood underplanting in North Mississippi. J. For. 1957, 55, 287–290. [Google Scholar]
- Johnson, P.S. Responses of planted northern red oak to three overstory treatments. Can. J. For. Res. 1984, 14, 536–542. [Google Scholar] [CrossRef]
- Tworkoski, T.J.; Smith, D.W.; Parrish, D.J. Regeneration of red oak, white oak, and white pine by underplanting prior to canopy removal in the Virginia Piedmont. South. J. Appl. Ecol. 1986, 10, 206–210. [Google Scholar]
- Craig, J.M.; Lhotka, J.M.; Stringer, J.W. Evaluating initial responses of natural and underplanted oak reproduction and a shade-tolerant competitor to midstory removal. For. Sci. 2014, 60, 1164–1171. [Google Scholar] [CrossRef]
- Truax, B.; Gagnon, D.; Chevrier, N. Nitrate reductase activity in relation to growth and soil N-forms in red oak and red ash planted in three different environments: Forest, clear-cut and field. For. Ecol. Manag. 1994, 64, 71–82. [Google Scholar] [CrossRef]
- Balandier, P.; Collet, C.; Miller, J.H.; Reynolds, P.E.; Zedaker, S.M. Designing forest vegetation management strategies based on the mechanisms and dynamics of crop tree competition by neighbouring vegetation. Forestry 2006, 79, 3–27. [Google Scholar] [CrossRef]
- Dey, D.C.; Gardiner, E.S.; Schweitzer, C.J.; Kabrick, J.M.; Jacobs, D.F. Underplanting to sustain future stocking of oak (Quercus) in temperate deciduous forests. New For. 2012, 43, 955–978. [Google Scholar] [CrossRef]
- Pretzsch, H.; Rais, A. Wood quality in complex forests versus even-aged monocultures: Review and perspectives. Wood Sci. Technol. 2016, 50, 845–880. [Google Scholar] [CrossRef]
- Crow, T.R. Reproductive mode and mechanisms for self-replacement of northern red oak (Quercus rubra)—A review. For. Sci. 1988, 34, 19–40. [Google Scholar]
- Wendel, G.W.; Clay Smith, H. Eastern white pine. In Silvics of North America: 1. Conifers. Agriculture Handbook 654; Burns, R.M., Honkala, B.H., Eds.; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1990; Volume 2, pp. 972–999. [Google Scholar]
- Parker, W.C.; Dey, D.C.; Newmaster, S.G.; Elliott, K.A.; Boysen, E. Managing succession in conifer plantations: Converting young red pine (Pinus resinosa Ait.) plantations to native forest types by thinning and underplanting. For. Chron. 2001, 77, 721–734. [Google Scholar] [CrossRef]
- Götmark, F.; Schott, K.M.; Jensen, A.M. Factors influencing presence–absence of oak (Quercus spp.) seedlings after conservation-oriented partial cutting of high forests in Sweden. Scand. J. For. Res. 2011, 26, 136–145. [Google Scholar] [CrossRef]
- Paquette, A.; Bouchard, A.; Cogliastro, A. Successful under-planting of red oak and black cherry in early-successional deciduous shelterwoods of North America. Ann. For. Sc. 2006, 673, 823–831. [Google Scholar] [CrossRef]
- Truax, B.; Gagnon, D.; Lambert, F.; Fortier, J. Multiple-use zoning model for private forest owners in agricultural landscapes: A case study. Forests 2015, 6, 3614–3664. [Google Scholar] [CrossRef]
- Frey, B.R.; Ashton, M.S. Growth, survival and sunfleck response of underplanted red oaks (Quercus spp., section Erythrobalanus) along a topographic gradient in southern New England. For. Ecol. Manag. 2018, 419–420, 179–186. [Google Scholar] [CrossRef]
- Farrar, J.L. Les arbres du Canada; Fides et le Service Canadien des Forêts, Ressources Naturelles Canada: St-Laurent, QC, Canada, 2006.
- Sander, I.L. Northern red oak. In Silvics of North America: 2. Hardwoods. Agriculture Handbook 654; Burns, R.M., Honkala, B.H., Eds.; Department of Agriculture, Forest Service, U.S.: Washington, DC, USA, 1990; Volume 2, pp. 1401–1414. [Google Scholar]
- Clements, J.R. Development of a white pine underplantation in thinned and unthinned aspen. For. Chron. 1966, 42, 244–250. [Google Scholar] [CrossRef]
- Smidt, M.F.; Puettmann, K.J. Overstory and understory competition affect underplanted eastern white pine. For. Ecol. Manag. 1998, 105, 137–150. [Google Scholar] [CrossRef]
- Oliver, L.B.; Jeremy, P.S.; Comer, C.E.; Williams, H.M.; Symmank, M.E. Weed control and overstory reduction improve survival and growth of under-planted oak and hickory seedlings. Rest. Ecol. 2018, 1–12. [Google Scholar] [CrossRef]
- Doyon, F.; Bouchard, A.; Gagnon, D. Tree productivity and successional status in Québec northern hardwoods. Écoscience 1998, 5, 222–231. [Google Scholar] [CrossRef]
- St-Jacques, C.; Gagnon, D. La végétation forestière du secteur nord-ouest de la vallée du Saint-Laurent, Québec. Can. J. Bot. 1988, 66, 793–804. [Google Scholar] [CrossRef]
- Von Althen, F.W. Sowing and Planting Shagbark and Bitternut Hickories on Former Farmland in Southern Ontario; Information Report O-X-403; Forestry Canada, Ontario Region: Sault Ste. Marie, ON, Canada, 1990; p. 11.
- Smith, H.C. Bitternut hickory. In Silvics of North America: 2. Hardwoods. Agriculture Handbook 654; Burns, R.M., Honkala, B.H., Eds.; Department of Agriculture, Forest Service, U.S.: Washington, DC, USA, 1990; Volume 2, pp. 389–417. [Google Scholar]
- Marie-Victorin, F.; Rouleau, E.; Brouillet, L.; Hay, S.G.; Goulet, I. Flore Laurentienne—3e edition; Gaëtan Morin éditeur ltée: Montréal, QC, Canada, 2002; p. 1093. [Google Scholar]
- Gagnon, D.; Bouchard, A. La végétation de l’escarpement d’Eardley, parc de la Gatineau, Québec. Can. J. Bot. 1981, 59, 2667–2691. [Google Scholar] [CrossRef]
- Gauthier, S.; Gagnon, D. La végétation des contreforts des Laurentides: Une analyse des gradients écologiques et du niveau successionnel des communautés. Can. J. Bot. 1990, 68, 391–401. [Google Scholar] [CrossRef]
- The Committee on the Status of Endangered Wildlife in Canada (COSEWIC). Butternut Juglans cinerea. Available online: http://www.cosewic.gc.ca/eng/sct1/searchdetail_e.cfm?id=793&StartRow=1&boxStatus=All&boxTaxonomic=All&location=All&change=All&board=All&commonName=butternut&scienceName=&returnFlag=0&Page=1 (accessed on 22 May 2015).
- Williams, R.D. Black walnut. In Silvics of North America: 2. Hardwoods. Agriculture Handbook 654; Burns, R.M., Honkala, B.H., Eds.; Department of Agriculture, Forest Service, U.S.: Washington, DC, USA, 1990; pp. 771–789. [Google Scholar]
- Cogliastro, A.; Gagnon, D.; Daigle, S.; Bouchard, A. Improving hardwood afforestation success: An analysis of the effects of soil properties in southwestern Quebec. For. Ecol. Manag. 2003, 177, 347–359. [Google Scholar] [CrossRef]
- Van Sambeek, J.W.; Schlesinger, R.C.; Roth, P.L.; Bocoum, I. Revitalizing slow-growth black walnut plantings. In Proceedings of the Seventh Central Hardwood Forest Conference; Rink, G., Budelsky, C.A., Eds.; USDA Forest Service; North Central Forest Experiment Station: Carbondale, IL, USA, 1989; pp. 108–114. [Google Scholar]
- Von Althen, F.W. Revitalizing a black walnut plantation through weed control and fertilization. For. Chron. 1985, 61, 71–74. [Google Scholar] [CrossRef]
- Huot, M.; Lebel, F. Le plan de gestion du cerf de Virginie au Québec 2010–2017; Direction de l’expertise sur la faune et ses habitats, Ministère des Ressources naturelles et de la Faune du Québec (MRNF): Québec, QC, Canada, 2010.
- Rooney, T.P.; Waller, D.M. Direct and indirect effects of white-tailed deer in forest ecosystems. For. Ecol. Manag. 2003, 181, 165–176. [Google Scholar] [CrossRef]
- Association Maritime du Québec. Lac Memphrémagog. Available online: http://www.navigationquebec.com/fiche_lac.php?l_id=46 (accessed on 14 June 2015).
- Robitaille, A.; Saucier, J.-P. Paysages régionaux du Québec méridional; Les publications du Québec: Ste-Foy, QC, Canada, 1998; p. 213. [Google Scholar]
- Westveld, M. Natural forest vegetation zones of New England. J. For. 1956, 54, 332–338. [Google Scholar]
- Cogbill, C.V.; Burk, J.; Motzkin, G. The forests of presettlement New England, USA: Spatial and compositional patterns based on town proprietor surveys. J. Bio geogr. 2002, 29, 1279–1304. [Google Scholar] [CrossRef]
- Government of Canada. Station results-1981–2010 climate normals and averages. Available online: http://climate.weather.gc.ca/climate_normals/station_select_1981_2010_e.html?searchType=stnProv&lstProvince=QC (accessed on 16 February 2017).
- Bardon, R.E.; Countryman, D.W.; Hall, R.B. Tree shelters reduced growth and survival of underplanted red oak seedlings in southern Iowa. North. J. Appl. For. 1999, 16, 103–107. [Google Scholar]
- West, P. Tree and Forest Measurement; Springer-Verglag: Berlin, Germany, 2009; p. 190. [Google Scholar]
- Perron, J.-Y. Inventaire forestier. In Manuel de foresterie; Ordre des ingénieurs forestiers du Québec, Ed. Les Presses de l’Université Laval: Ste-Foy, QC, Canada, 1996; pp. 390–473. [Google Scholar]
- Conseil des Productions Végétales du Québec. Méthodes D’analyse des Sols, des Fumiers et des Tissus végétaux. Available online: https://www.agrireseau.net/documents/96351/methode-d_analyse-des-sols-des-fumiers-et-des-tissus-vegetaux-agdex-533-mai-1988 (accessed on 15 August 2018).
- Bouyoucos, G.J. Hydrometer method improved for making particle size analysis of soils. Agron. J. 1962, 54, 464–465. [Google Scholar] [CrossRef]
- Centre de référence en agriculture et agroalimentaire du Québec (CRAAQ). Guide de référence en fertilization, 1re ed.; Centre de référence en agriculture et agroalimentaire du Québec: Ste-Foy, QC, Canada, 2003; p. 40. [Google Scholar]
- Tran, T.S.; Simard, R.R. Mehlich III-Extractable elements. In Soil Sampling and Methods of Analysis; Carter, M.R., Ed.; Lewis Publishers and CRC Press: Boca Raton, FL, USA, 1993; pp. 43–49. [Google Scholar]
- Association of Official Agricultural Chemists (AOAC). Official Methods of Analysis. Method 984.27: Calcium, Copper, Iron, Magnesium, Manganese, Phosphorus, Potassium, Sodium and Zinc in Infant Formula—Inductively Coupled Plasma Emission Spectroscopic, 16th ed.; AOAC International: Rockville, MD, USA, 1999; p. 1200. [Google Scholar]
- Qian, P.; Schoenau, J.J.; Huang, W.Z. Use of ion exchange membranes in routine soil testing. Comm. Soil Sci. Plant Anal. 1992, 23, 1791–1804. [Google Scholar] [CrossRef]
- Petersen, R.G. Design and Analysis of Experiments; Marcel-Dekker: New York, NY, USA, 1985; p. 429. [Google Scholar]
- Warton, D.I.; Hui, F.K.C. The arcsine is asinine: the analysis of proportions in ecology. Ecol. 2011, 92, 3–10. [Google Scholar] [CrossRef]
- Bowersox, T.W.; McCormick, L.W. Herbaceous communities reduce the juvenile growth of northern red oak, white ash, yellow poplar, but not white pine. In Proceedings of the Central Hardwood Forest Conference VI; Hoy, R.L., Woods, F.W., DeSelm, H., Eds.; University of Tennessee: Knoxville, TN, USA, 1987; pp. 39–43. [Google Scholar]
- Saunders, M.R.; Puettmann, K.J. Effects of overstory and understory competition and simulated herbivory on growth and survival of white pine seedlings. Can. J. For. Res. 1999, 29, 536–546. [Google Scholar] [CrossRef]
- Boucher, J.F.; Bernier, P.Y.; Munson, A.D. Radiation and soil temperature interactions on the growth and physiology of eastern white pine (Pinus strobus L.) seedlings. Plant Soil 2001, 236, 165–174. [Google Scholar] [CrossRef]
- Kern, C.C.; Reich, P.B.; Montgomery, R.A.; Strong, T.F. Do deer and shrubs override canopy gap size effects on growth and survival of yellow birch, northern red oak, eastern white pine, and eastern hemlock seedlings? For. Ecol. Manag. 2012, 267, 134–143. [Google Scholar] [CrossRef]
- Meilleur, A.; Véronneau, H.; Bouchard, A. Shrub communities as inhibitors of plant succession in southern Quebec. Environ. Manag. 1994, 18, 907–921. [Google Scholar] [CrossRef]
- Truax, B.; Gagnon, D.; Lambert, F.; Chevrier, N. Nitrate assimilation of raspberry and pin cherry in a recent clearcut. Can. J. Bot. 1994, 72, 1343–1348. [Google Scholar] [CrossRef]
- Kim, T.; Mills, H.A.; Wetzstein, H.Y. Studies on effects of nitrogen form on growth, development, and nutrient uptake in pecan. J. Plant Nutr. 2002, 25, 497–508. [Google Scholar] [CrossRef]
- Nicodemus, M.; Salifu, K.; Jacobs, D. Nitrate reductase activity and nitrogen compounds in xylem exudate of Juglans nigra seedlings: Relation to nitrogen source and supply. Tree Struct. Funct. 2008, 22, 685–695. [Google Scholar] [CrossRef]
- Truax, B.; Lambert, F.; Gagnon, D.; Chevrier, N. Nitrate reductase and glutamine synthetase activities in relation to growth and nitrogen assimilation in red oak and red ash seedlings: Effects of N-forms, N concentration and light intensity. Tree Struct. Funct. 1994, 9, 12–18. [Google Scholar] [CrossRef]
- Truax, B.; Fortier, J.; Gagnon, D.; Lambert, F. Black plastic mulch or herbicide to accelerate bur oak, black walnut, and white pine growth in agricultural riparian buffers? Forests 2018, 9, 258. [Google Scholar] [CrossRef]
- Truax, B.; Gagnon, D.; Lambert, F.; Fortier, J. Riparian buffer growth and soil nitrate supply are affected by tree species selection and black plastic mulching. Ecol. Eng. 2017, 106, 82–93. [Google Scholar] [CrossRef]
- Boucher, S.; Crête, M.; Ouellet, J.-P.; Daigle, C.; Lesage, L. Large-scale trophic interactions: White-tailed deer growth and forest understory. Écoscience 2004, 11, 286–295. [Google Scholar] [CrossRef]
- Burns, R.M.; Honkala, B.H. Silvics of North America; USDA, Forest Service: Washington, DC, USA, 1990; Volume 2, Harwoods, Agriculture Handbook 654.
- Davis, V.; Burger, J.A.; Rathfon, R.; Zipper, C.E.; Miller, C.R. Chapter 7: Selecting tree species for reforestation of Appalachian mined lands. In The Forestry Reclamation Approach: Guide to Successful Reforestation of Mined Lands; Adams, M.B., Ed.; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2017; Gen. Tech. Rep. NRS-169; pp. 1–10. [Google Scholar]
- Krajicek, J.E. Planted Black Walnut Does Well on Cleared Forest Sites—If Competition Is Controlled; USDA, Forest Service, North Central Forest Experiment Station: St. Paul, MN, USA, 1975; pp. 1–4.
- Daigle, C.; Crête, M.; Lesage, L.; Ouellet, J.-P.; Huot, J. Summer diet of two white-tailed deer, Odocoileus virginianus, populations living at low and high density in southern Québec. Can. Fld.-Nat. 2004, 118, 360–367. [Google Scholar] [CrossRef]
- D’Orangeville, L.; Bouchard, A.; Cogliastro, A. Post-agricultural forests: Landscape patterns add to stand-scale factors in causing insufficient hardwood regeneration. For. Ecol. Manag. 2008, 255, 1637–1646. [Google Scholar] [CrossRef]
- Lorimer, C.G.; Chapman, J.W.; Lambert, W.D. Tall understorey vegetation as a factor in the poor development of oak seedlings beneath mature stands. J. Ecol. 1994, 82, 227–237. [Google Scholar] [CrossRef]
- Lucas, R.W.; Roberto, S.-G.; Cobb, D.B.; Waring, B.G.; Anderson, F.; McShea, W.J.; Casper, B.B. White-tailed deer (Odocoileus virginianus) positively affect the growth of mature northern red oak (Quercus rubra) trees. Ecosphere 2013, 4, 1–15. [Google Scholar] [CrossRef]
- Ward, J.S.; Gent, M.P.N.; Stephens, G.R. Effects of planting stock quality and browse protection-type on height growth of northern red oak and eastern white pine. For. Ecol. Manag. 2000, 127, 205–216. [Google Scholar] [CrossRef]










| Species | Survival (%) | Total Height (cm) | Basal Diameter (mm) | Stem Volume (cm3/tree) |
|---|---|---|---|---|
| Bitternut hickory | 90 | 162 | 16.4 | 158 |
| Black walnut | 64 | 171 | 27.6 | 862 |
| SE | 3.6 | 17 | 2.1 | 316 |
| p-value | <0.001 | 0.74 | <0.001 | <0.05 |
| Community Type | Stand Structure | Soil | Understory Plants | |||||
|---|---|---|---|---|---|---|---|---|
| Total Basal Area (m2/ha) | Canopy Openness (%) | CEC (meq/100 g) | NO3 Supply (µg/10 cm2/41d) | Gramineae spp. (%) | Rubus spp. (%) | Solidago spp. (%) | Carex spp. (%) | |
| White ash | 10.8 ± 1.7 | 25.9 ± 1.8 | 18.5 ± 0.5 | 89.8 ± 13.6 | 5 ± 6 | 39 ± 4 | 25 ± 7 | 11 ± 2 |
| Grey birch | 16.3 ± 1.1 | 35.9 ± 1.2 | 16.4 ± 0.3 | 14.9 ± 9.3 | 26 ± 4 | 3 ± 3 | 45 ± 5 | 2 ± 1 |
| Community Type | Total Height of Sheltered Trees (cm) | |||
|---|---|---|---|---|
| Red Oak | White Pine | Bitternut Hickory | Black Walnut | |
| White ash | 200.5 ± 23.1 | 192.8 ± 8.7 | 214.0 ± 20.0 | 287.9 ± 34.8 |
| Grey birch | 131.1 ± 15.9 | 230.4 ± 6.0 | 126.3 ± 13.7 | 108.0 ± 25.4 |
| Community Type | Unprotected White Pine | ||
|---|---|---|---|
| Total Height (cm) | Survival (%) | Stem Browsing (%) | |
| White ash | 87.5 ± 14.1 | 59.4 ± 7.0 | 81.5 ± 6.7 |
| Grey birch | 163.5 ± 9.6 | 89.0 ± 4.8 | 34.5 ± 4.6 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Truax, B.; Gagnon, D.; Fortier, J.; Lambert, F.; Pétrin, M.-A. Ecological Factors Affecting White Pine, Red Oak, Bitternut Hickory and Black Walnut Underplanting Success in a Northern Temperate Post-Agricultural Forest. Forests 2018, 9, 499. https://doi.org/10.3390/f9080499
Truax B, Gagnon D, Fortier J, Lambert F, Pétrin M-A. Ecological Factors Affecting White Pine, Red Oak, Bitternut Hickory and Black Walnut Underplanting Success in a Northern Temperate Post-Agricultural Forest. Forests. 2018; 9(8):499. https://doi.org/10.3390/f9080499
Chicago/Turabian StyleTruax, Benoit, Daniel Gagnon, Julien Fortier, France Lambert, and Marc-Antoine Pétrin. 2018. "Ecological Factors Affecting White Pine, Red Oak, Bitternut Hickory and Black Walnut Underplanting Success in a Northern Temperate Post-Agricultural Forest" Forests 9, no. 8: 499. https://doi.org/10.3390/f9080499
APA StyleTruax, B., Gagnon, D., Fortier, J., Lambert, F., & Pétrin, M.-A. (2018). Ecological Factors Affecting White Pine, Red Oak, Bitternut Hickory and Black Walnut Underplanting Success in a Northern Temperate Post-Agricultural Forest. Forests, 9(8), 499. https://doi.org/10.3390/f9080499