Over- and Underyielding in Time and Space in Experiments with Mixed Stands of Scots Pine and Norway Spruce

 ,
,
Abstract
1. Introduction
- Are Scots pine–Norway spruce mixed stands more productive than the most productive monoculture of either tree species (transgressive overyielding)?
- Are Scots pine–Norway spruce mixtures more productive than the average of both corresponding monocultures (overyielding)?
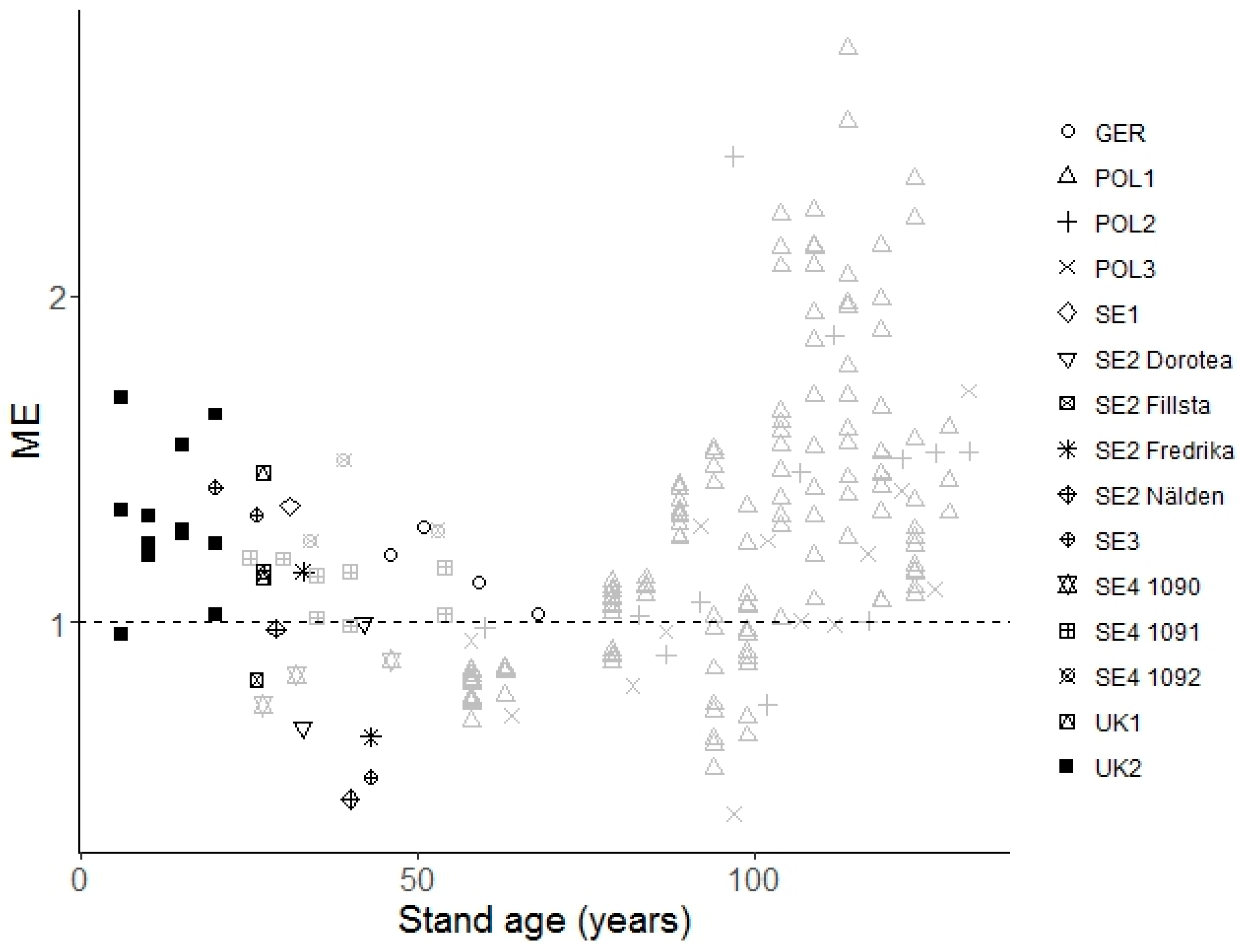
- Does the mixing effect change significantly over stand age or with site conditions?
2. Materials and Methods
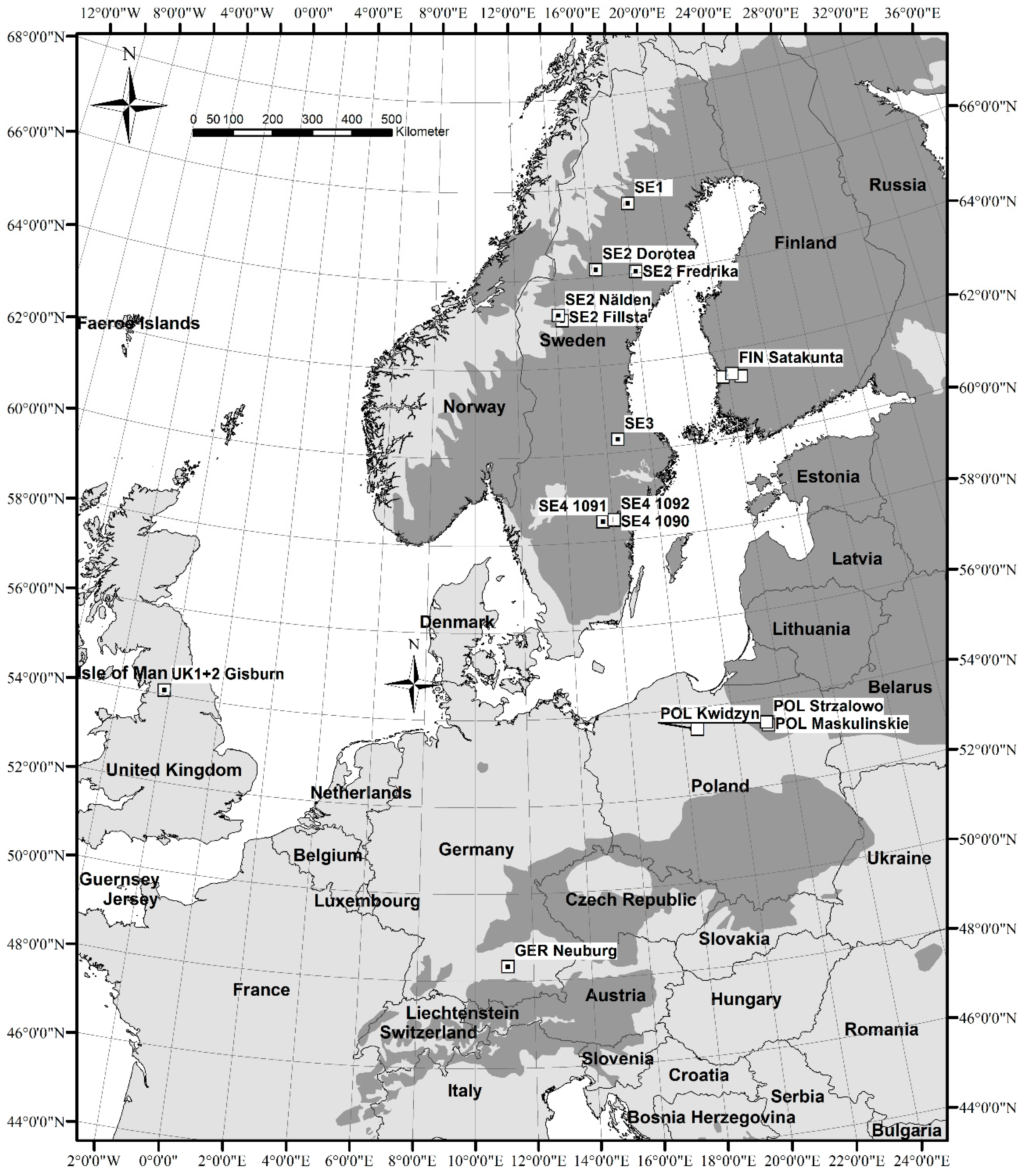
2.1. Material: Study Sites
2.2. Methods
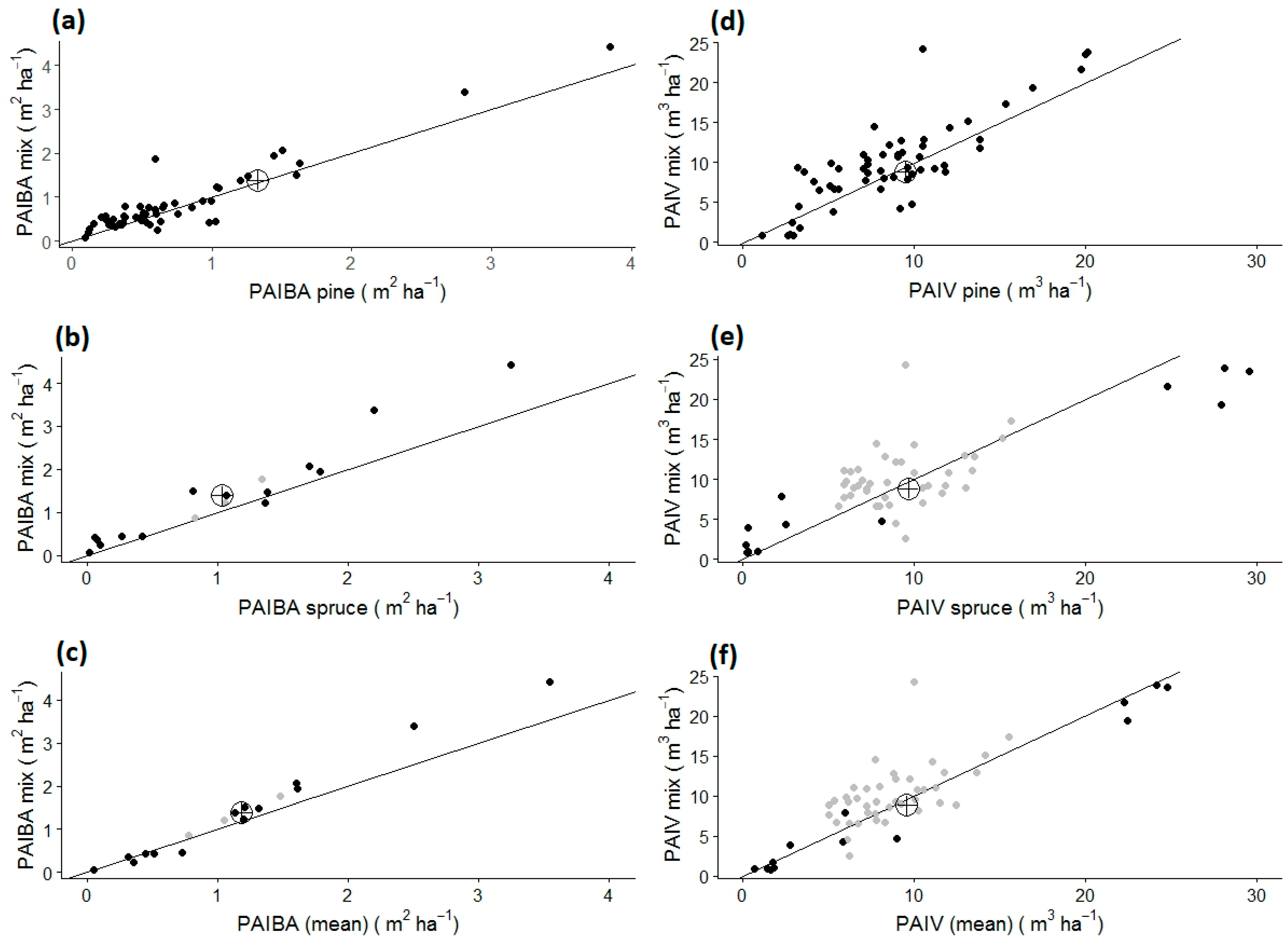
2.2.1. Quantifying of Mixing Effects
2.2.2. Statistical Analysis and Models
3. Results
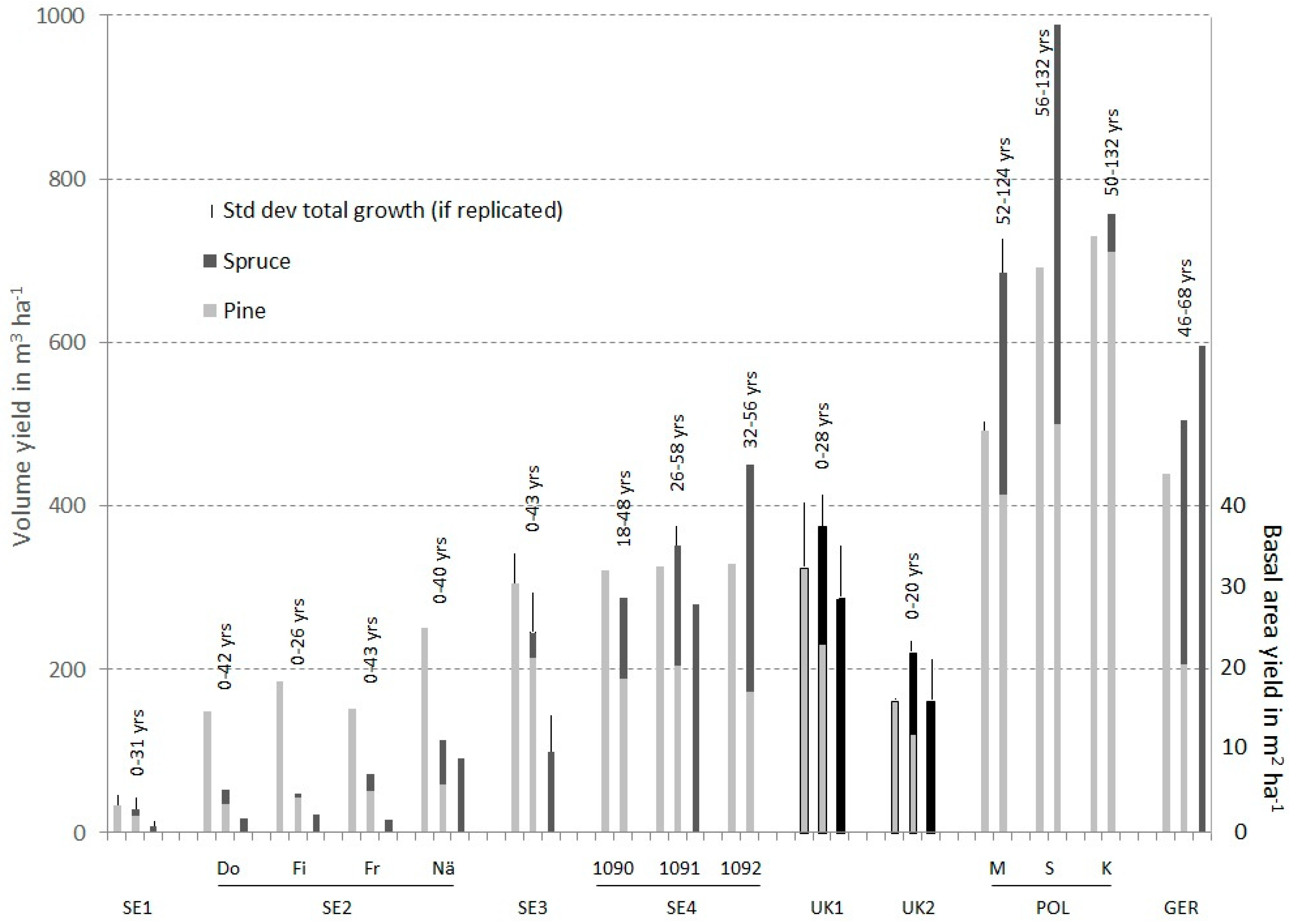
3.1. Over- and Underyielding in Single Experiments (Analysis A)
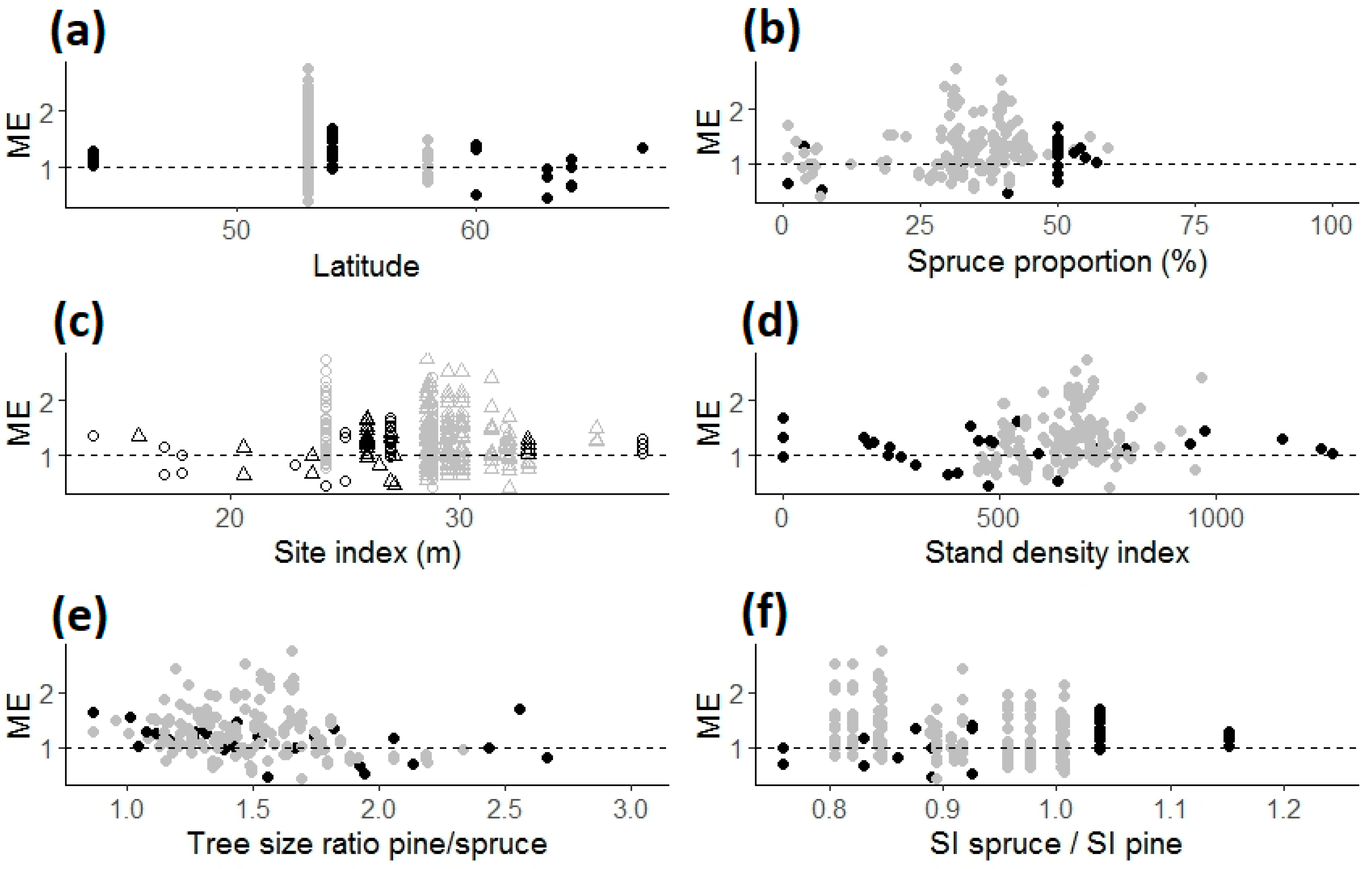
3.2. Influence of Stand Age, Tree Species Proportion, Climate, and Other Factors on the Mixture Effects
4. Discussion
4.1. The Mixing Effect of Observed Increment without Spruce Growth Estimates
4.2. The Mixing Effect of Studied Stands When Complemented by Simulated Growth Estimates for Lacking Treatments
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Stokland, J.N.; Eriksen, R.; Stein, M.T.; Korhonen, K.; Tomppo, E.; Rajaniemi, S.; Söderberg, U.; Toet, H.; Riis-Nielsen, T. Forest Biodiversity Indicators in the Nordic Countries; Nordic Council of Ministers: Copenhagen, Denmark, 2003; 108p. [Google Scholar]
- Karvinen, S.; Välkky, E.; Torniainen, T.; Gerasimov, Y. Northwest Russian Forestry in a Nutshell; Finnish Forest Research Institute: Helsinki, Finland, 2006; ISBN-13: 978-951-40-2007-0. [Google Scholar]
- Engelmark, O.; Hytteborn, H. Coniferous forest. Acta Phytogeogr. Suec. 1999, 84, 55–74. [Google Scholar]
- Drozdowski, S.; Andrzejczyk, T.; Bielak, K.; Buraczyk, W.; Gawro, N.L. Silvicultural planning in spruce mire forests by the means of the BDq method. Sylwan 2014, 158, 733–742. [Google Scholar]
- EUFORGEN 2013 GIS Database. Available online: http://www.euforgen.org (accessed on 2 May 2018).
- Gamfeldt, L.; Snäll, T.; Bagchi, R.; Jonsson, M.; Gustafsson, L.; Kjellander, P.; Ruiz-Jaen, M.C.; Froberg, M.; Stendahl, J.; Philipson, C.D.; et al. Higher levels of multiple ecosystem services are found in forests with more tree species. Nat. Commun. 2013, 4, 1340. [Google Scholar] [CrossRef] [PubMed]
- Bravo-Oviedo, A.; Pretzsch, H.; Ammer, C.; Andenmatten, E.; Barbati, A.; Barreiro, S.; Brang, P.; Bravo, F.; Coll, L.; Corona, P.; et al. European Mixed Forests: Definition and research perspectives. For. Syst. 2014, 23, 518–533. [Google Scholar] [CrossRef]
- Felton, A.; Felton, A.; Holmström, E.; Laudon, H.; Björkman, C.; Nilsson, U.; Sonesson, J.; Fahlvik, N.; Stens, A.; Keskitalo, C.; et al. Replacing monocultures with mixed-species stands: Ecosystem service implications of two production forest mixture alternatives in Sweden. Ambio 2016, 45, 124–139. [Google Scholar] [CrossRef] [PubMed]
- Piri, T.; Korhonen, K.; Sairanen, A. Occurrence of Heterobasidion annosum in monospecific and mixed spruce stands in southern Finland. Scand. J. For. Res. 1990, 5, 113–125. [Google Scholar]
- Milligan, H.T.; Koricheva, J. Effects of tree species richness and composition on moose winter browsing damage and foraging selectivity: An experimental study. J. Anim. Ecol. 2013, 82, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Björkman, C.; Bylund, H.; Nilsson, U.; Nordlander, G.; Schroeder, M. Forest management to mitigate insect damage in a changing climate: Possibilities and uncertainties. In Climate Change and Insect Pests; Björkman, C., Niemelä, J., Eds.; CABI: New York, NY, USA, 2015. [Google Scholar]
- Forrester, D.I. The spatial and temporal dynamics of species interactions in mixed-species forests: From pattern to process. For. Ecol. Manag. 2014, 312, 282–292. [Google Scholar] [CrossRef]
- Kalliokoski, T. Root System Traits of Norway Spruce, Scots Pine, and Silver Birch in Mixed Boreal Forests: Analysis of Root Architecture, Morphology and Anatomy. Ph.D. Thesis, University of Helsinki, Helsinki, Finland, 2011. [Google Scholar]
- Brown, A.H.F. Functioning of mixed-species stands at Gisburn, N.W. England. In The Ecology of Mixed-Species Stands of Trees; Cannell, M.G.R., Malcolm, D.C., Robertson, P.A., Eds.; Blackwell: Boston, MA, USA, 1992; pp. 125–150. [Google Scholar]
- Forrester, D.I.; Bonal, D.; Dawud, S.; Gessler, A.; Granier, A.; Pollastrini, M.; Grossiord, C. Drought responses by individual tree species are not often correlated with tree species diversity in European forests. J. Appl. Ecol. 2016, 53, 1725–1734. [Google Scholar] [CrossRef]
- Valinger, E.; Fridman, J. Factors affecting the probability of windthrow at stand level as a result of Gudrun winter storm in southern Sweden. For. Ecol. Manag. 2011, 262, 398–403. [Google Scholar] [CrossRef]
- Jonsson, B. Volume Yield to Mid-Rotation in Monospecific and Mixed Sown Stands of Pinus Sylvestris and Picea Abies in Sweden; Studia Forestalia Suecica, Swedish University of Agricultural Sciences: Umea, Sweden, 2001; ISSN 91-576-6137-5. Available online: https://pub.epsilon.slu.se/3336/1/SFS211.pdf (accessed on 1 August 2018).
- Jonsson, B. Dynamics of Field-Layer Vegetation and Tree Growth in Young Pinus Sylvestris and Picea Abies Stands on Microsites in Swedish Lapland; Report for Skog & Trä: 2010:5 SLU; Unit for Field-based Forest research, Swedish University of Agricultural Sciences: Vindeln, Sweden, 2010; ISBN 978-91-977896-6-0. [Google Scholar]
- Lindén, M.; Agestam, E. Increment and yield in mixed and monoculture stands of Pinus sylvestris and Picea abies based on an experiment in southern Sweden. Scand. J. For. Res. 2003, 17, 155–162. [Google Scholar] [CrossRef]
- Ekö, P.M. A Growth Simulator for Swedish Forests, Based on Data from the National Forest Survey; Department of Silviculture, Swedish University of Agriculture Science: Umea, Finland, 1985; 224p., ISBN 91-576-2386-4. (In Swedish with English summary). [Google Scholar]
- Hynynen, J.; Ojansuu, R.; Hökkä, H.; Salminen, H.; Siipilehto, J.; Haapala, P. Models for Predicting Stand Development in MELA System; Research Papers; The Institute Finnish Forest Research: Vantaa, Finland, 2002. [Google Scholar]
- Pretzsch, H.; Schütze, G. Transgressive overyielding in mixed compared with monospecific stands of Norway spruce and European beech in Central Europe: Evidence on stand level and explanation on individual tree level. Eur. J. For. Res. 2009, 128, 183–204. [Google Scholar] [CrossRef]
- Mason, W.; Connolly, T. The development of species mixtures in British spruce forests: Guidance from the Gisburn experiment. Forestry 2014, 87, 1–9. [Google Scholar] [CrossRef]
- Bielak, K.; Dudzinska, M.; Pretzsch, H. Mixed stands of Scots pine (Pinus sylvestris L.) and Norway spruce [Picea abies (L.) Karst] can be more productive than monocultures. Evidence from over 100 years of observation of long-term experiments. For. Syst. 2014, 23, 573–589. [Google Scholar] [CrossRef]
- Forrester, D.I.; Pretzsch, H. Tamm Review: On the strength of evidence when comparing ecosystem functions of mixtures with monocultures. For. Ecol. Manag. 2015, 356, 41–53. [Google Scholar] [CrossRef]
- Pretzsch, H.; del Río, M.; Ammer, C.; Avdagic, A.; Barbeito, I.; Bielak, K.; Brazaitis, G.; Coll, L.; Dirnberger, G.; Drössler, L.; et al. Growth and yield of mixed versus monospecific stands of Scots pine (Pinus sylvestris L.) and European beech (Fagus sylvatica L.) analysed along a productivity gradient through Europe. Eur. J. For. Res. 2015, 134, 927–947. [Google Scholar] [CrossRef]
- Toïgo, M.; Vallet, P.; Perot, T.; Bontemps, J.D.; Piedally, C.; Courbaud, B. Overyielding in mixed forests decreases with site productivity. J. Ecol. 2015, 103, 502–512. [Google Scholar] [CrossRef]
- Liang, J.; Crowther, T.W.; Picard, N.; Wiser, S.; Zhou, M. Positive biodiversity-productivity relationship predominant in global forests. Science 2016, 354, aaf8957. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.E.; McNamara, N.; Mason, W.L. Functioning of mixed species stands: Evidence from a long-term forest experiment. In Ecological Studies 176. Forest Diversity and Function: Temperate and Boreal Systems; Springer: Berlin, Germany, 2005; pp. 111–130. [Google Scholar]
- Holm, S.; Jonsson, B.; Kallur, H. Department of Forest Biometry and Management; SLU; Status Report; Umea, Sweden, 1984. [Google Scholar]
- Scherer-Lorenzen, M.; Körner, C.; Schulze, E.-D. Forest Diversity and Function: Temperate and Boreal Systems; Ecological Studies, 176; Springer: Berlin, Germany, 2005; pp. 347–376. [Google Scholar]
- Wellhausen, K. Produktivitat und Klimasensitivitat von Kiefer (Pinus sylvestris L.) und Fichte (Picea abies (L.) Karst.) im Rein-und Misch bestand-Untersuchungen entlang eines okologischen Gradient en. In Proceedings of the Sektion Ertragskunde DVFFA, Kammerforst, Germany, 2015; Available online: http://sektionertragskunde.fvabw.de/2015/Beitrag_12.pdf (accessed on 4 May 2018). (In German).
- Pretzsch, H. Forest Dynamics, Growth and Yield: From Measurement to Model; Springer: Berlin, Germany, 2009. [Google Scholar]
- Hägglund, B.; Lundmark, J.-E. Part 2 Diagrams and tables. In Guide for Site Productivity Assessments in Swedish Forests; Skogsstyrelsen: Jönköping, Sweden, 1982; ISBN 91-85748-7. (In Swedish) [Google Scholar]
- Edwards, P.N.; Christie, J.M. Yield tables for forest management. In Forestry Commission Booklet 48; HMSO: London, UK, 1981. [Google Scholar]
- Assmann, E.; Franz, F. Vorläufige Fichtenertragstafeln für Bayern (Preliminary spruce yield tables for Bavaria). In Bayerisches Staatsministerium für Ernährung; Landwirtschaft, Forsten, Ed.; Hilfstafeln für die Forsteinrichtung, 1963; 334p, Available online: http://www.stmelf.bayern.de/ (accessed on 2 August 2018).
- Wiedemann, E. Ertragstafel für die Baumart Kiefer, mäßige Durchforstung (Yield Tables for Pine, Moderate Thinning). In Bayerisches Staatsministerium für Ernährung; Landwirtschaft, Forsten, Ed.; Hilfstafeln für die Forsteinrichtung, 1943; 334p, Available online: http://www.stmelf.bayern.de/ (accessed on 2 August 2018).
- Bruchwald, A. Change in top height of pine forest stands with age. Bull. Acad. Pol. Sci. Ser. Biol. 1977, 5, 335–342. [Google Scholar]
- Hector, A.; Bazeley-White, E.; Loreau, M.; Otway, S.; Schmid, B. Overyielding in grassland communities: Testing the sampling effect hypothesis with replicated biodiversity experiments. Ecol. Lett. 2002, 5, 502–511. [Google Scholar] [CrossRef]
- Martonne, D.E. La Météorologie 21. In Une Nouvelle Fonction Climatologique: L’indice D’aridité; Gauthier-Villars: Paris, France, 1926; pp. 449–458. (In French) [Google Scholar]
- Reineke, L.H. Perfecting a stand-density index for even-aged forests. J. Agric. Res. 1933, 46, 627–638. [Google Scholar]
- Szymkiewicz, B. Tablice Zasobnosci i Przyrostu Drzewostanów. Polish Yield Tables. Tables for Norway Spruce by Schwappach (1943): I-V Site Classes; PWRiL: Warszawa, Poland, 1966. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; Heisterkamp, S.; Van Willigen, B. Nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1–122. 2015. Available online: http://CRAN.R-project.org/package=nlme (accessed on 2 March 2018).
- Spathelf, P.; Ammer, C. Forest management of Scots pine (Pinus sylvestris L.) in northern Germany—A brief review of the history and current trends. Arch. For. Sci. 2015, 86, 59–66. [Google Scholar]
- Drössler, L.; Fahlvik, N.; Elfving, B. Application and limitations of growth models for silvicultural purposes in heterogeneously structured forest in Sweden. J. For. Sci. 2013, 59, 458–473. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, H.Y.H.; Reich, P.B. Forest productivity increases with evenness, species richness and trait variation: A global meta-analysis. J. Ecol. 2012, 100, 742–749. [Google Scholar] [CrossRef]
- Jactel, H.; Gritti, E.S.; Drössler, L.; Forrester, D.I.; Mason, W.L.; Morin, X.; Pretzsch, H.; Castagneyrol, B. Positive biodiversity-productivity relationships in forests: Climate matters. Biol. Lett. 2018, 14, 20170747. [Google Scholar] [CrossRef] [PubMed]
- Haase, J.; Castagneyrol, B.; Cornelissen, J.H.C.; Ghazoul, J.; Kattge, J.; Koricheva, J.; Scherer-Lorenzen, M.; Morath, S.; Jactel, H. Contrasting effects of tree diversity on young tree growth and resistance to insect herbivores across three biodiversity experiments. Oikos 2015, 124, 1674–1685. [Google Scholar] [CrossRef]
- Vallet, P.; Perot, T. Tree diversity effect on dominant height in temperate forest. For. Ecol. Manag. 2016, 381, 106–114. [Google Scholar] [CrossRef]
- Pretzsch, H.; Dieler, J. The dependency of the size-growth relationship of Norway spruce (Picea abies [L.] Karst.) and European beech (Fagus sylvatica [L.]) in forest stands on long-term site conditions, drought events, and ozone stress. Trees 2011, 25, 355–369. [Google Scholar] [CrossRef]
- Pukkala, T.; Vettenranta, J.; Kolström, T.; Miina, J. Productivity of mixed stands of Pinus sylvestris and Picea abies. Scand. J. For. Res. 1994, 9, 143–153. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Possible Species Interactions | Interactions Found in Mixed Scots Pine–Norway Spruce Forests |
|---|---|
| With influence on nutrient availability | Mostly overlapping fine-roots [13], increased soil nitrogen when spruce was grown in mixture [14] |
| With influence on water availability | Overlapping fine-roots [13], higher water stress in boreal mixed forest, and reduced water stress in hemiboreal mixed forest [15] |
| Influencing light absorption and use | |
| Temporal effects and interactions between resources | Lower incidence of Heterobasidion annosum [9], higher or similar browsing damage [10] |
| Stand structure | Less storm damage [16] |
| Study | SE1 | SE2 | SE3 | SE4 | UK1 | UK2 | POL | GER | FIN |
|---|---|---|---|---|---|---|---|---|---|
| Observed stand age [years] | 0–31 | 0–42 | 5–43 | 14–46 | 0–28 | 0–20 | 52–124 | 46–68 | 0–10 |
| 0–26 | 22–54 | 56–132 | 0–10 | ||||||
| 0–43 | 29–53 | 50–132 | 0–10 | ||||||
| 0–40 | |||||||||
| N of study sites | 1 | 4 | 1 | 3 | 1 | 1 | 3 | 1 | 3 |
| N of plots with pure pine | 10 | 4 | 10 | 3 | 3 | 3 | 6 | 1 | 6 |
| N of plots with pure spruce | 10 | 4 | 10 | 1 | 3 | 3 | - | 1 | 6 |
| N of plots with mixture | 10 | 4 | 10 | 4 | 3 | 3 | 6 | 1 | 6 |
| Plot size [m2] | 1200 | 300–2200 | 1400 | 1000 | 1000 | 1000 | 2500 | 600 | 400 |
| Initial spruce proportion of trees (or basal area BA) [%] | 50 | 50–90 | 50 | 50 | 50 | 50 | 3–33 (BA) | 47 (BA) | 50 |
| Stand establishment | planted | planted | seeded | planted | planted | planted | planted or seeded | planted | planted |
| Management after establishment | no | no | no | equal tree number after removals | no | no | A- and B-grade thinnings | A- and B-grade thinnings | no |
| Study | SE1 | SE2 | SE3 | SE4 | FIN | UK1 | UK2 | POL | GER | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Study site | SE1 | Dorotea | Fillsta | Fredrika | Nälden | SE3 | 1090 | 1091 | 1092 | 1 | 2 | 3 | UK1 | UK2 | Maskulinskie | Strzałowo | Kwidzyn | GER |
| Latitude (° N) | 67 | 64 | 63 | 64 | 63 | 60 | 58 | 58 | 58 | 61 | 61 | 61 | 54 | 54 | 53 | 53 | 53 | 44 |
| Martonne index | 55 | 64 | 59 | 59 | 59 | 48 | 44 | 44 | 44 | 43 | 43 | 43 | 78 | 78 | 35 | 36 | 36 | 42 |
| Growing season (days) | 130 | 150 | 150 | 150 | 150 | 175 | 180 | 180 | 180 | 165 | 165 | 165 | >200 | >200 | 190 | 190 | 190 | 200 |
| SI (Hdom at age 100) for pine | 16 | 24 | 27 | 21 | 27 | 27 | 30 | 32 | 36 | 29 | 28 | 29 | 26 | 26 | 29 | 31 | 32 | 33 (Hmean) |
| SI (Hdom at age 100) for spruce | 14 | 18 | 22 | 17 | 24 | 25 | 33 | 29 | 32 | 35 | 35 | 35 | 27 | 27 | 27 | 29 | 29 | 38 |
| Vegetation type | Thin-leaved grasses/V. myrtillus | V. myrtillus | Low herbs | V. myrtillus | V. myrtillus | n.s. | V. myrtillus/V.vitis-idea | V. myrtillus/V.vitis-idea | V. myrtillus/V.vitis-idea | V. myrtillus | V. myrtillus | V. myrtillus | Festuca-Agrostis before planting | Mosses and coarse grasses | Oxalis acet./Impatiens parviflora | Calama-grostis/V. Myrtillus | Oxalis acet./Impatiens parviflora | Thin-leaved grasses/V. myrtillus |
| Soil type | Podzol on glacial till | Podzol | Podzol | Podzol | Podzol | n.s. | Podzol on glacial till | Podzol on glacial till | Podzol | Podzol | Podzol | Podzol | Stagnogley /gley on glayey glacial till | Stagnogley /gley on glayey glacial till | Cambic arenosols (rusty) | Cambic arenosols (rusty) | Cambic arenosols (rusty) | Brown earth on loamy sand |
| Soil moisture class | Mesic | Mesic | Mesic | Mesic | Mesic | n.s. | Mesic | Mesic | Moist | Mesic–moist | Mesic–moist | Mesic–moist | Very moist | Very moist | Mesic | Mesic | Mesic | Mesic–moist |
| Mean annual temperature (°C) | −0.5 | 1 | 1 | 1 | 1 | 5 | 6 | 6 | 6 | 4 | 4 | 4 | 8 | 8 | 7 | 8 | 8 | 8 |
| Mean annual precipitation (mm) | 520 | 700 | 650 | 650 | 650 | 725 | 700 | 700 | 700 | 600 | 600 | 600 | 1400 | 1400 | 600 | 650 | 650 | 760 |
| Study | SE1 | SE2 | SE3 | SE4 | UK1 | UK2 | POL | GER | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Growth Variable | Volume Growth | Volume Growth | Volume Growth | Volume Growth | Basal Area Growth | Basal Area Growth | Volume Growth | Volume Growth | |||||||
| Unit | m3 ha−1 | m3 ha−1 | m3 ha−1 | m3 ha−1 | m2 ha−1 | m2 ha−1 | m3 ha−1 | m3 ha−1 | |||||||
| Site | Dorotea | Fillsta | Fredrika | Nälden | 1090 | 1091 | 1092 | Maskulinskie | Strzałowo | Kwidzyn | |||||
| Period (stand age) | 0–31 | 0–42 | 0–26 | 0–43 | 0–40 | 5–43 | 18–48 | 26–57 | 32–56 | 0–28 | 0–20 | 52–124 | 56–132 | 50–132 | 46–68 |
| Productivity pure pine | 34.1 (±1.2) | 148 | 185 | 151 | 251 | 304 (±28) | 389 | 326 | 329 | 32.5 (±7.9) | 40.2 (±4.2) | 476 (±11) | 608 | 730 | 439 |
| Productivity pure spruce | 8.1 (± 0.5) | 18 | 21 | 16 | 90 | 98 (±9) | 346 | 279 | 351 | 28.8 (±6.3) | 30.6 (±10.4) | 642 | 710 | 791 | 596 |
| Productivity mixed pine-spruce | 28.6 (±1.2) | 52 | 48 | 71 | 113 | 244 (±22) | 350 | 351 (±23) | 451 | 37.5 (±3.9) | 45.5 (±1.7) | 671 (±29) | 836 | 757 | 504 |
| ME max | 0.84 (±0.07) | 0.35 | 0.26 | 0.47 | 0.45 | 0.80 (±0.16) | 0.90 | 1.08 | 1.28 | 1.15 (±0.23) | 1.13 (±0.10) | 1.41 | 1.38 | 1.04 | 0.85 |
| ME | 1.36 (±0.93) | 0.63 | 0.47 | 0.85 | 0.66 | 1.21 (±0.87) | 0.95 | 1.16 | 1.33 | 1.25 (±0.22) | 1.29 (±0.22) | 1.20 | 1.27 | 1.00 | 0.97 |
| MEpine | 0.84 (±0.07) | 0.35 | 0.26 | 0.47 | 0.45 | 0.80 (±0.16) | 0.90 | 1.08 | 1.37 | 1.15 (±0.23) | 1.13 (±0.10) | 1.41 (±0.03) | 1.38 | 1.04 | 1.15 |
| MEspruce | 3.53 (±0.02) | 2.89 | 2.29 | 4.44 | 1.26 | 2.49 (±0.05) | 1.01 | 1.26 | 1.28 | 1.30 (±0.19) | 1.49 (±0.23) | 1.05 | 1.18 | 0.96 | 0.85 |
| Model | Parameter | Value | Std. Error | DF | t-Value | p-Value |
|---|---|---|---|---|---|---|
| 1 | (Intercept) | 3.0720 | 0.5999 | 18 | 5.1209 | 0.0001 |
| Latitude | −0.0307 | 0.0102 | 9 | −2.9977 | 0.0150 | |
| Stand age | −0.0074 | 0.0030 | 18 | −2.4273 | 0.0259 | |
| 2 | (Intercept) | −1.5503 | 0.5442 | 184 | −2.8486 | 0.0049 |
| Stand age | 0.0147 | 0.0017 | 184 | 8.4765 | <0.0001 | |
| Precipitation | 0.0015 | 0.0004 | 24 | 35.489 | 0.0016 | |
| Tree size ratio | 0.3740 | 0.1373 | 184 | 27.238 | 0.0071 | |
| 3 | (Intercept) | −1.3732 | 0.6573 | 181 | −2.0891 | 0.0381 |
| Stand age | 0.0148 | 0.0020 | 181 | 7.2532 | <0.0001 | |
| Precipitation | 0.0013 | 0.0006 | 22 | 2.1692 | 0.0412 | |
| Tree size ratio | 0.3584 | 0.1626 | 181 | 2.2046 | 0.0287 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drössler, L.; Agestam, E.; Bielak, K.; Dudzinska, M.; Koricheva, J.; Liziniewicz, M.; Löf, M.; Mason, B.; Pretzsch, H.; Valkonen, S.; et al. Over- and Underyielding in Time and Space in Experiments with Mixed Stands of Scots Pine and Norway Spruce. Forests 2018, 9, 495. https://doi.org/10.3390/f9080495
Drössler L, Agestam E, Bielak K, Dudzinska M, Koricheva J, Liziniewicz M, Löf M, Mason B, Pretzsch H, Valkonen S, et al. Over- and Underyielding in Time and Space in Experiments with Mixed Stands of Scots Pine and Norway Spruce. Forests. 2018; 9(8):495. https://doi.org/10.3390/f9080495
Chicago/Turabian StyleDrössler, Lars, Eric Agestam, Kamil Bielak, Małgorzata Dudzinska, Julia Koricheva, Mateusz Liziniewicz, Magnus Löf, Bill Mason, Hans Pretzsch, Sauli Valkonen, and et al. 2018. "Over- and Underyielding in Time and Space in Experiments with Mixed Stands of Scots Pine and Norway Spruce" Forests 9, no. 8: 495. https://doi.org/10.3390/f9080495
APA StyleDrössler, L., Agestam, E., Bielak, K., Dudzinska, M., Koricheva, J., Liziniewicz, M., Löf, M., Mason, B., Pretzsch, H., Valkonen, S., & Wellhausen, K. (2018). Over- and Underyielding in Time and Space in Experiments with Mixed Stands of Scots Pine and Norway Spruce. Forests, 9(8), 495. https://doi.org/10.3390/f9080495