Ungulate Browsing Limits Bird Diversity of the Central European Hardwood Floodplain Forests



Abstract
1. Introduction
2. Materials and Methods
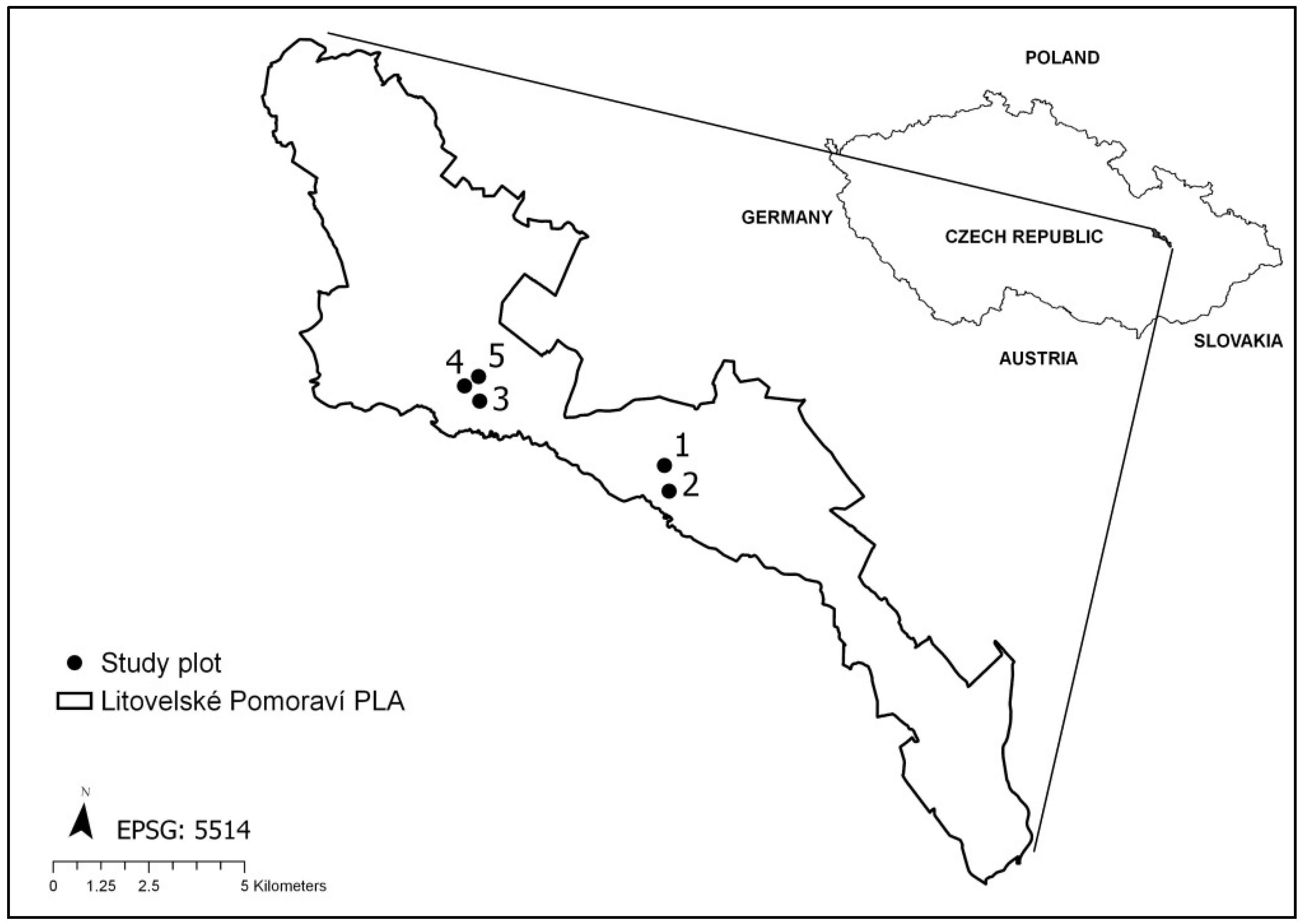
2.1. Study Area
2.2. Bird Census Data
2.3. Ungulate Browsing Research
2.4. Statistical Analysis and Control Plots
3. Results
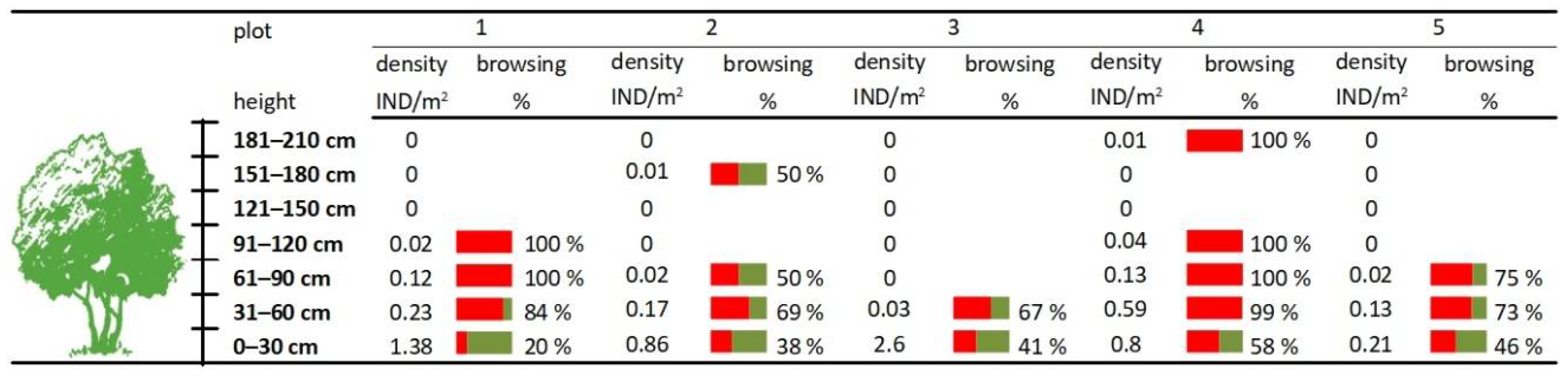
3.1. Ungulate Browsing and Woody Plants in Herb and Shrub Layers
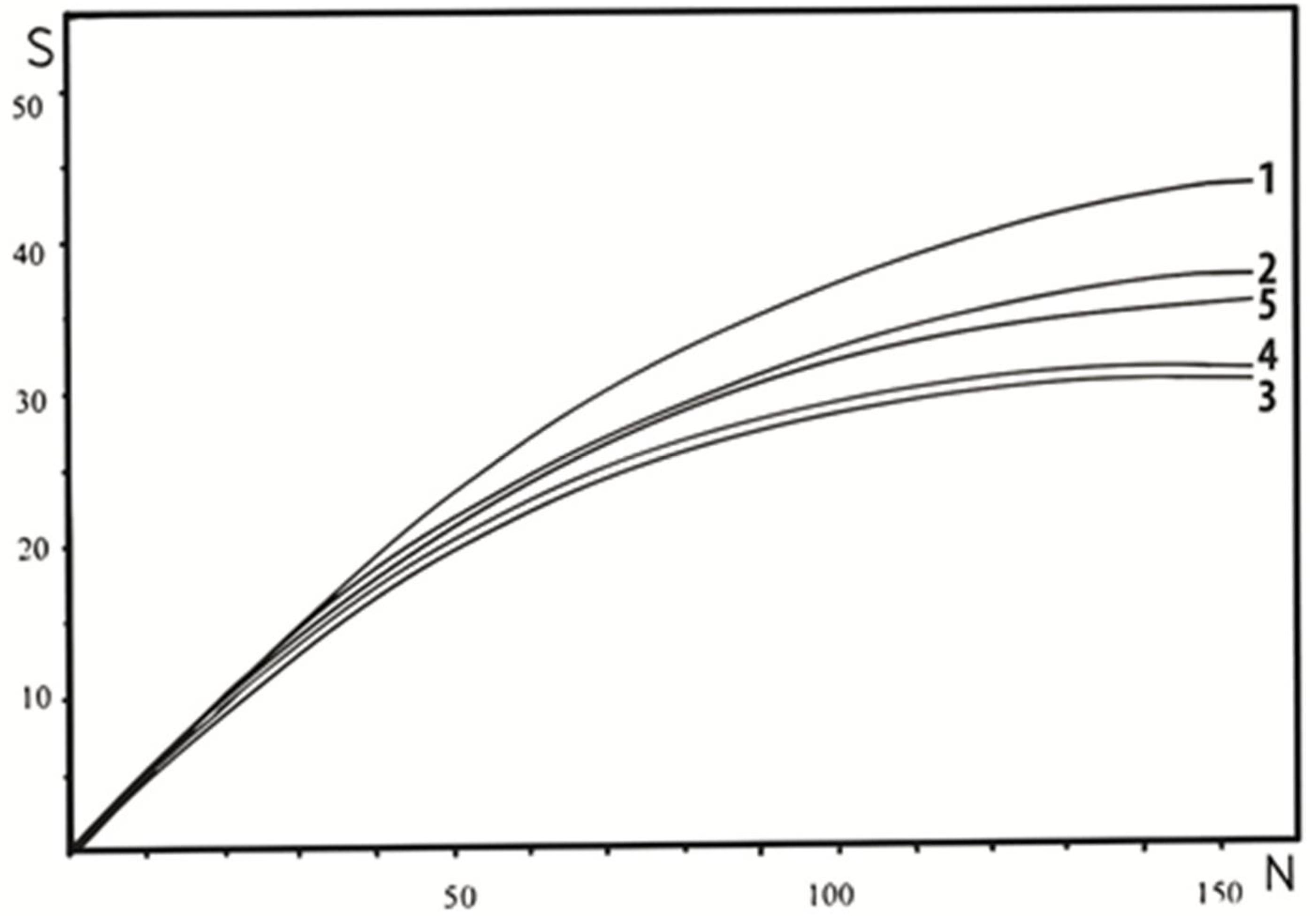
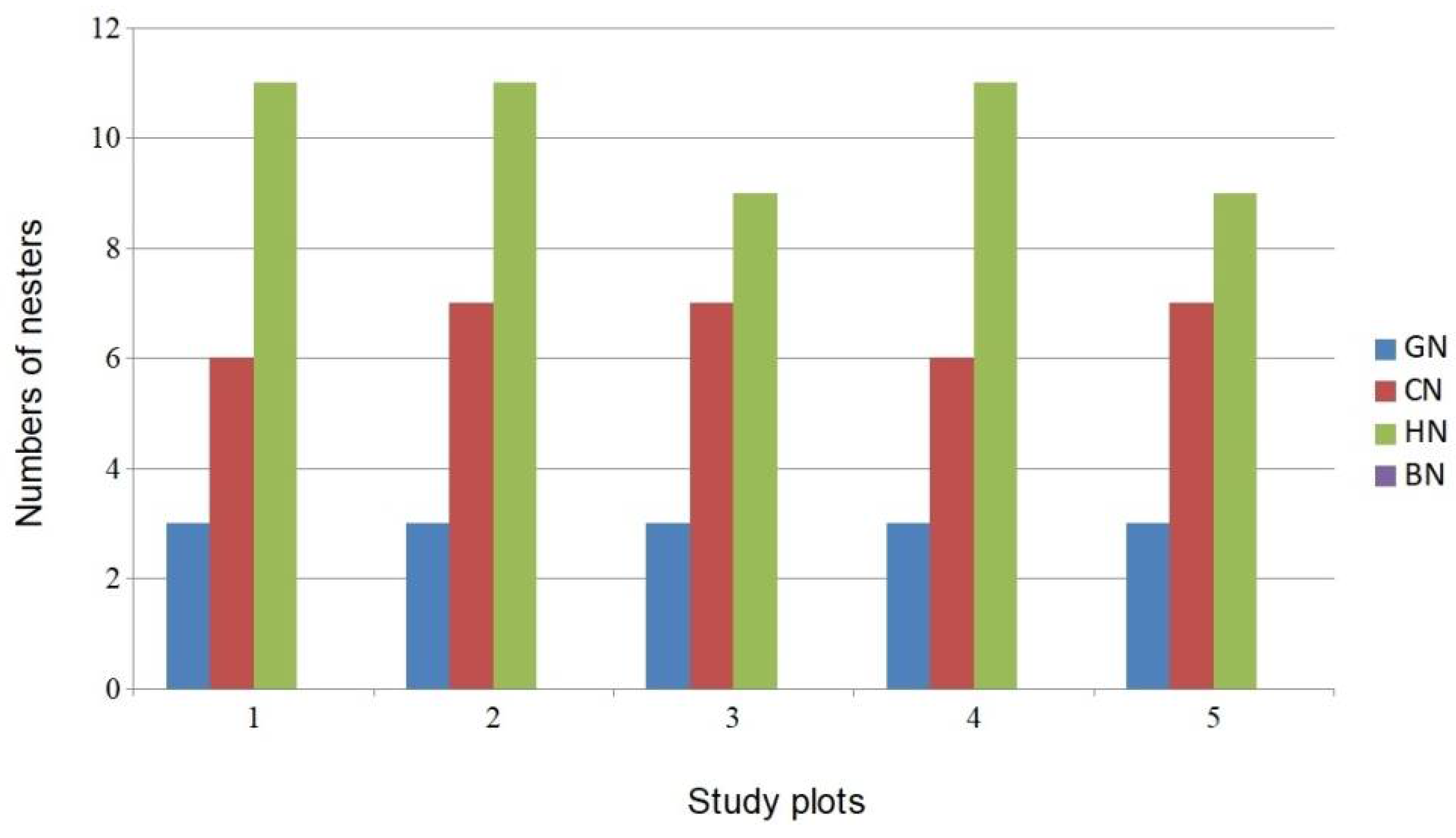
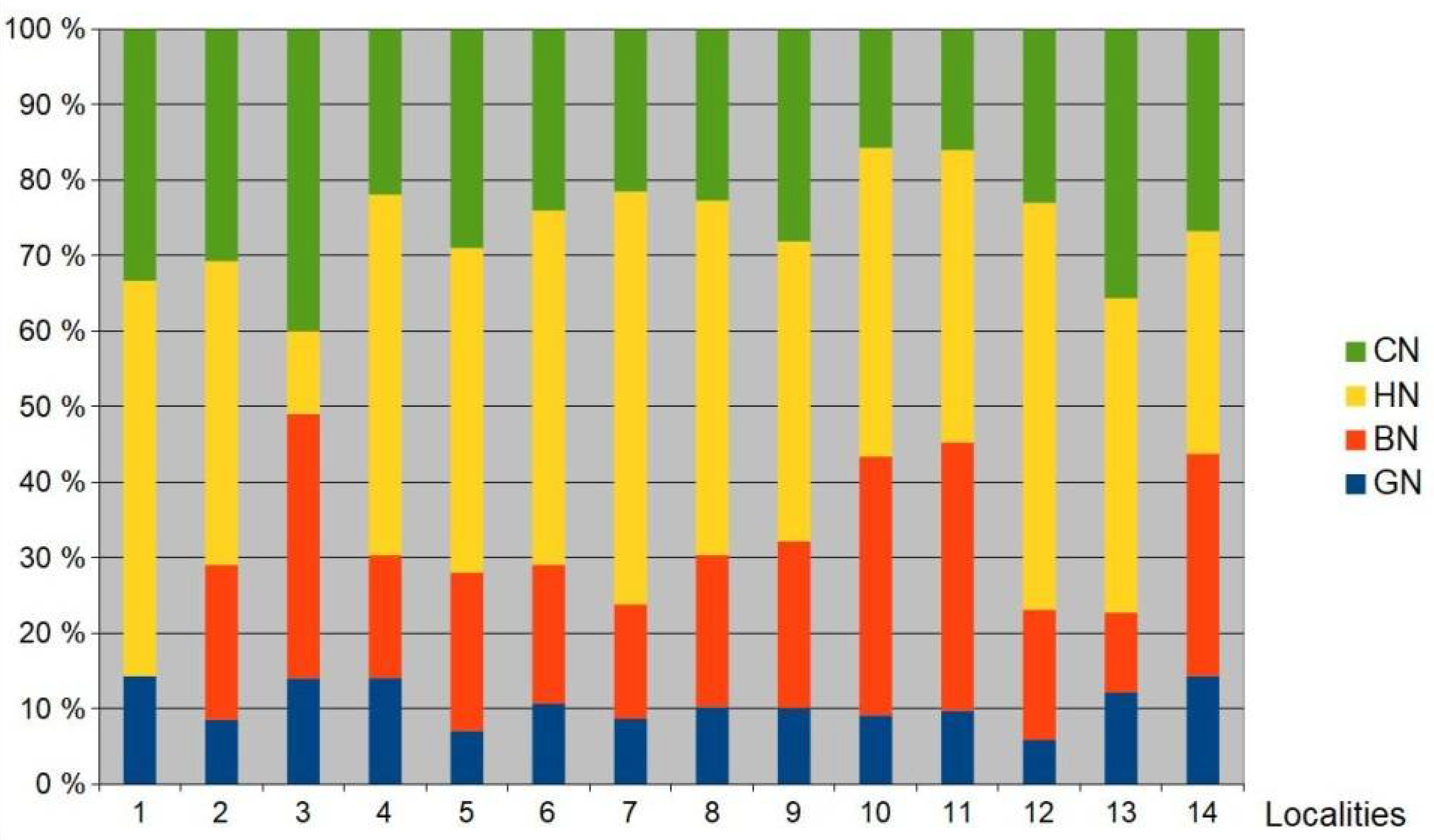
3.2. Bird Community
4. Discussion
4.1. Impact of Ungulate Browsing on the Forest Understorey of Floodplain Forests
4.2. Bird Communities in Floodplain Forests
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Doyon, F.; Giroux, J.F.; Gagnon, D. Effects of different types of management on the diversity of birds and small mammals in a hardwood forest. In Landscape Ecology in Land use Planning Methods and Practice, Proceedings of the 4th Workshop of the Canadian-Society-for-Landscape-Ecology-and-Management, Laval, QC, Canada, 5–7 June 1995; Université Laval: Sanite Foy, QC, Canada; pp. 211–219.
- Laiolo, P. Effects of habitat structure, floral composition and diverzity on a forest bird community in north-western Italy. Folia Zool. 2002, 51, 121–128. [Google Scholar]
- Lindbladh, M.; Lindstrom, A.; Hedwall, P.O.; Felton, A. Avian diversity in Norway spruce production forests—How variation in structure and composition reveals pathways for improving habitat quality. For. Ecol. Manag. 2017, 397, 48–56. [Google Scholar] [CrossRef]
- Perry, R.W.; Jenkins, J.M.A.; Thill, R.E.; Thompson, F.R. Long-term effects of different forest regeneration methods on mature forest birds. For. Ecol. Manag. 2018, 408, 183–194. [Google Scholar] [CrossRef]
- Sallabanks, R.; Haufler, J.B.; Mehl, C.A. Influence of forest vegetation structure on avian community composition in west-central Idaho. Wildl. Soc. B 2006, 34, 1079–1093. [Google Scholar] [CrossRef]
- Pierce, A.R.; King, S.L. A comparison of avian communities and habitat characteristics in floodplain forests associated with valley plugs and unchannelized streams. River Res. Appl. 2011, 27, 1315–1324. [Google Scholar] [CrossRef]
- Zenzal, T.J.; Smith, R.J.; Ewert, D.N.; Diehl, R.H.; Buler, J.J. Fine-scale heterogeneity drives forest use by spring migrant landbirds across a broad, contiguous forest matrix. Condor 2018, 120, 166–184. [Google Scholar] [CrossRef]
- Knutson, M.G.; McColl, L.E.; Suarez, S.A. Breeding bird assemblages associated with stages of forest succession in large river floodplains. Nat. Areas J. 2005, 25, 55–70. [Google Scholar]
- Vacek, Z.; Vacek, S.; Bilek, L.; Kral, J.; Remes, J.; Bulusek, D.; Kralicek, I. Ungulate impact on natural regeneration in spruce-beech-fir stands in Cerny Dul nature reserve in the Orlicke Hory mountains, Case study from Central Sudetes. Forests 2014, 5, 2929–2946. [Google Scholar] [CrossRef]
- Chapron, G.; Kaczensky, P.; Linnell, J.D.C.; von Arx, M.; Huber, D.; Andren, H.; Lopez-Bao, J.V.; Adamec, M.; Alvares, F.; Anders, O.; et al. Recovery of large carnivores in Europe’s modern human-dominated landscapes. Science 2014, 346, 1517–1519. [Google Scholar] [CrossRef] [PubMed]
- De Jager, N.R.; Cogger, B.J.; Thomsen, M.A. Interactive effects of flooding and deer (Odocoileus virginianus) browsing on floodplain forest recruitment. For. Ecol. Manag. 2013, 303, 11–19. [Google Scholar] [CrossRef]
- Holt, C.A.; Fuller, R.J.; Dolman, P.M. Exclusion of deer affects responses of birds to woodland regeneration in winter and summer. IBIS 2014, 156, 116–131. [Google Scholar] [CrossRef]
- Newson, S.E.; Johnston, A.; Renwick, A.R.; Baillie, S.R.; Fuller, R.J. Modelling large-scale relationship between changes in woodland deer and bird populations. J. Anim. Ecol. 2012, 49, 278–286. [Google Scholar] [CrossRef]
- Hinsley, S.A.; Hill, R.A.; Fuller, R.J.; Bellamy, P.E.; Rothery, P. Bird species distributions across woodland canopy structure gradients. Community Ecol. 2009, 10, 99–110. [Google Scholar] [CrossRef]
- Charchuk, C.; Bayne, E.M. Avian community response to understory protection harvesting in the boreal forest of Alberta, Canada. For. Ecol. Manag. 2018, 407, 9–15. [Google Scholar] [CrossRef]
- Eichhorn, M.P.; Ryding, J.; Smith, M.J.; Gill, R.M.A.; Siriwardena, G.M.; Fuller, R.J. Effects of deer on woodland structure revealed through terrestrial laser scanning. J. Appl. Ecol. 2017. [Google Scholar] [CrossRef]
- Garabedian, J.E.; Moorman, C.E.; Peterson, M.N.; Kilgo, J.C. Use of LiDAR to define habitat thresholds for forest bird conservation. For. Ecol. Manag. 2017, 399, 24–36. [Google Scholar] [CrossRef]
- Fuller, R.J. Searching for biodiversity gains through woodfuel and forest management. J. Appl. Ecol. 2013, 50, 1295–1300. [Google Scholar] [CrossRef]
- Petty, S.J.; Avery, M.I. Forest Bird Communities. A Review of the Ecology and Management of Forest Bird Communities in Relation to Silvicultural Practices in the British Uplands; Forestry Commission Paper; Forestry Commission: Edinburgh, UK, 1990; Volume 26, pp. 1–41. [Google Scholar]
- Fuller, R.J.; Smith, K.W.; Hinsley, S.A. Temperate western European woodland as a dynamic environments for birds: A resource-based review. In Birds and Habitat: Relationship in Changing Landscapes; Fuller, R.J., Ed.; Cambridge University Press: Cambridge, UK, 2012; pp. 352–380. [Google Scholar]
- Machar, I. Conservation and Management of Floodplain Forests in the Protected Landscape Area Litovelske Pomoravi (Czech Republic) Introduction. In Conservation and Management of Floodplain Forests in the Protected Landscape Area Litovelske Pomoravi (Czech Republic); Accession Number: WOS:000331015800001; Machar, I., Ed.; Palacky University: Olomouc, Czech Republic, 2009; pp. 7–108. ISBN 978-80-244-2355-5. [Google Scholar]
- Kilianova, H.; Pechanec, V.; Svobodova, J.; Machar, I. Analysis of the evolution of the floodplain forests in the aluvium of the Morava river. In Proceedings of the 12th International Multidisciplinary Scientific Geoconference (SGEM 2012), Albena, Bulgaria, 17–23 June 2012; SGEM: Albena, Bulgaria48535300001, 2012. Accession Number: WOS:000348535300001. Volume IV, pp. 1–8, ISSN 1314-2704. [Google Scholar]
- Simon, J.; Machar, I.; Bucek, A. Linking the historical research with the growth simulation model of hardwood floodplain forests. Pol. J. Ecol. 2014, 62, 273–288. [Google Scholar] [CrossRef]
- Kusbach, A.; Friedl, M.; Zouhar, V.; Mikita, T.; Šebesta, J. Assessing Forest Classification in a Landscape-Level Framework: An Example from Central European Forests. Forests 2017, 8, 461. [Google Scholar] [CrossRef]
- Miko, L. Nature and landscape protection in the European context. In Ochrana prirody a krajiny v Ceske Republice, Vols I and II; Accession Number: WOS:000334387900004; Machar, I., Drobilova, L., Eds.; Palacky University: Olomouc, Czech Republic, 2012; pp. 43–49. ISBN 978-80-244-3041-6. [Google Scholar]
- Hanzelka, J.; Reif, J. Responses to the black locust (Robinia pseudoacacia) invasion differ between habitat specialist and generalist in central European forest birds. J. Ornithol. 2015, 156, 1015–1024. [Google Scholar] [CrossRef]
- Kroftova, M.; Reif, J. Management implications of bird responses to variation in non-native/native tree ratios within central European forest stands. For. Ecol. Manag. 2017, 391, 330–337. [Google Scholar] [CrossRef]
- Bibby, C.J.; Burges, N.D.; Hill, D.A.; Mustoe, S. Bird Census Techniques; Academic Press: London, UK, 2007; pp. 42–64. ISBN 978-0-12-095831-3. [Google Scholar]
- Poprach, K.; Vrbkova, J.; Machar, I. Detectability as an important factor influencing the knowledge of bird diversity in a floodplain forest ecosystem. J. For. Sci. 2015, 61, 89–97. [Google Scholar] [CrossRef]
- Wiens, J.A. The Ecology of Bird Communities, Vol. 1, Foundation and Patterns; Cambridge University Press: Cambridge, UK, 1989; pp. 1–539. ISBN 0-521-26030. [Google Scholar]
- Machar, I. Changes in ecological stability and biodiversity in a floodplain landscape. In Applying Landscape Ecology in Conservation and Management of the Floodplain Forest (Czech Republic); Accession Number: WOS:000325436900004; Machar, I., Ed.; Palacky University: Olomouc, Czech Republic, 2012; pp. 73–87. ISBN 978-80-244-2997-7. [Google Scholar]
- Stastny, K.; Hudec, K. Fauna of the Czech Republic. Birds 2 and 3, 2nd ed.; Academia: Prague, Czech Republic, 2011; pp. 1–1178. ISBN 80-200-1113-7. [Google Scholar]
- Aulak, W. Small mammal communities of the Białowieża National Park. Acta Theriol. 1970, 15, 465–515. [Google Scholar] [CrossRef]
- Cermak, P.; Horsak, P.; Spirik, M.; Mrkva, R. Relationships between browsing damage and woody species dominance. J. For. Sci. (Prague) 2009, 55, 23–31. [Google Scholar]
- Cermak, P.; Beranova, P.; Oralkova, J.; Horsak, P.; Plsek, J. Relationships between browsing damage and the species dominance by the highly food-attractive and less food-attractive trees. Acta Univ. Agric. Mendel. Brun. 2011, 59, 29–36. [Google Scholar] [CrossRef]
- Cermak, P.; Mrkva, R. Browsing damage to broadleaves in some national nature reserves (Czech Republic) in 2000–2001. Ekológia (Bratislava) 2003, 22, 132–141. [Google Scholar]
- StatSoft, s.r.o. Statistica [software, CD-ROM]. Ver. 12. Praha, 2013. [Google Scholar]
- Jaccard, P. Étude comparative de la distribution florale dans une portion des Alpes et des Jura. Bull. Soc. Vaud. Sci. Nat. 1901, 37, 547–579. [Google Scholar]
- James, F.C.; Rathbun, S. Rarefaction, relative abundance and diversity of avian communities. Auk 1981, 98, 785–800. [Google Scholar]
- Author Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples. Version 8.0.0. 2005. Available online: http://viceroy.eeb.uconn.edu/estimates (accessed on 12 January 2018).
- Bures, S.; Maton, K. Ptaci slozka segmentu skupiny typu geobiocenu Ulmi-fraxineta populi v navrhovane CHKO Pomoravi [Birds of hardwood floodplain forest in proclaimed PLA Pomoravi—In Czech]. Sylvia 1984, 23–24, 37–46. [Google Scholar]
- Bures, S. Analyza ptaci slozky navrhovane SPR Sargoun [Birds of harwood floodplain forest in Sargoun locality—In Czech]. Unpublished Work. 1996. [Google Scholar]
- Horak, Z. Ptactvo okolí Starého Labe u Cihelny u Pardubic [Birds of Stare Labe u Cihelny locality—In Czech]. Panurus 1988, 9, 53–61. [Google Scholar]
- Chytil, J. Srovnání Produkce Savců a ptáků v Lužním lese [Comparison of the Production of Birds and Mammals in Floodplain Forest—In Czech]. Ph.D. Thesis, University JE Purkyne, Brno, Czech Republic, 1991. [Google Scholar]
- Kubecka, D. Avifauna lužního lesa na lokalitě Horní Záseky v CHKO Litovelské Pomoraví [Birds of floodplain forest in Horni Zaseky locality—In Czech]. Ph.D. Thesis, Palacky University, Olomouc, Czech Republic, 2003. [Google Scholar]
- Lemberk, V. Srovnání ornitocenóz čtyř lužních lesů ve východních Čechách [Comparison of avian communities in floodplain forests—In Czech]. Panurus 2001, 11, 69–79. [Google Scholar]
- Machar, I. The effect of floodplain forest fragmentation on bird community. J. For. Sci. 2012, 58, 213–224. [Google Scholar] [CrossRef]
- Machar, I. The impact of floodplain forest habitat conservation on the structure of bird breeding communities. Ekológia (Bratislava) 2011, 30, 36–50. [Google Scholar] [CrossRef]
- Pavelka, J. Hnízdní ornitocenóza v lužním lese u řeky Odry [Birds in floodplain forest along Odra river—In Czech]. Zprávy MOS (Přerov) 1987, 46, 115–118. [Google Scholar]
- Polasek, V. Výzkum Ornitocenózy lužního lesa v Litovelském Pomoraví [Birds of Floodplain Forest in Litovelske Pomoravi—In Czech]. Ph.D. Thesis, Palacky University, Olomouc, Czech Republic, 2001. [Google Scholar]
- Pykal, J. Ornitocenosy různých typů přirozených lesních společenstev v pahorkatině jihozápadních Čech [Avian community in hardwood forests in Czechia—In Czech]. Panurus 1991, 3, 67–76. [Google Scholar]
- Ruzicka, I. Ornitologický výzkum lokality Chrbovský les u Záříčí [Birds of Floodplain Forest Near Zarici—In Czech]. Ph.D. Thesis, Palacky University, Olomouc, Czech Republic, 2005. [Google Scholar]
- Storch, D. Densities and territory of birds in two different lowland communities in eastern Bohemia. Folia Zool. 1998, 47, 181–188. [Google Scholar]
- Toman, A. Avifauna SPR Zastudanci [Avian Community in Floodplain Forest Zastudanci Locality]. Ph.D. Thesis, Palacky University, Olomouc, Czech Republic, 2004. [Google Scholar]
- Nascher, F.A. Zur Waldbaulichen Bedeutung des Rothirschverbisses in der Waldgesellschaft des subalpinen Fichtenwalds in der Umgebung des Schweizerischen Nationalparks. Ph.D. Thesis, ETH Zürich, Rämistr, Zürich Switzerland, 1979. Diss. Nr. 6373. [Google Scholar]
- Heuze, P.; Schnitzler, A.; Klein, F. Is browsing the major factor of silver fir decline in the Vosges Mountains of France? For. Ecol. Manag. 2005, 217, 219–228. [Google Scholar] [CrossRef]
- Barancekova, M.; Krojerova-Prokesova, J.; Homolka, M. Impact of deer browsing on natural and artificial regeneration in floodplain forest. Folia Zool. 2007, 56, 354–364. [Google Scholar]
- Kubicek, F.; Simonovic, V.; Kollar, J.; Kanka, R. Herb layer biomass of the Morava river floodplain forests. Ekológia (Bratislava) 2008, 27, 23–30. [Google Scholar]
- Cogger, B.J.; De Jager, N.R.; Thomsen, M.; Adams, C.R. Winter Browse Selection by White-Tailed Deer and Implications for Bottomland Forest Restoration in the Upper Mississippi River Valley, USA. Nat. Areas J. 2014, 34, 144–153. [Google Scholar] [CrossRef]
- Cermak, P.; Mrkva, R.; Horsak, P.; Spirik, M.; Beranova, P.; Oralkova, J.; Plsek, J.; Kadlec, M.; Zarybnicky, O.; Svatos, M. Impact of Ungulate Browsing on Forest Dynamics, 1st ed.; Lesnická Práce, Folia Forestalia Bohemica: Kostelec nad Černými lesy, Czech Republic, 2011; pp. 61–66. ISBN 978-80-87154-94-6. [Google Scholar]
- Frerker, K.; Sonnier, G.; Waller, D.M. Browsing rates and ratios provide reliable indices of ungulate impacts on forest plant communities. For. Ecol. Manag. 2013, 291, 55–64. [Google Scholar] [CrossRef]
- Ammer, C. Impact of ungulates on structure and dynamics of natural regeneration of mixed mountain forests in the Bavarian Alps. For. Ecol. Manag. 1996, 88, 45–53. [Google Scholar] [CrossRef]
- Stergar, M. Objedenost Mladja Drevesnih Vrstv Odvisnosti od Zgradbe Sestoja. Ph.D. Thesis, Biotehniška fakulteta, Univerza v Ljubljani, Ljubljana, Slovenia, 2005; p. 70. [Google Scholar]
- Hubalek, Z. Seasonal variation of forest habitat preferences by birds in a lowland riverine ecosystem. Folia Zool. 2001, 50, 281–289. [Google Scholar]
- Schlaghamersky, J.; Hudec, K. The fauna of temperate European floodplain forests. In Floodplain Forests of the Temperate Zone of Europe; Klimo, E., Hager, H., Matic, S., Anic, I., Kulhavy, J., Eds.; Lesnicka Prace: Kostelec, Czech Republic, 2008; pp. 160–230. ISBN 978-80-87154-16-8. [Google Scholar]
- Walther, B.A.; Martin, J.L. Species richness estimation of bird communities: How to control for sampling effort? IBIS 2001, 143, 413–419. [Google Scholar] [CrossRef]
- Kolecek, J.; Paclik, M.; Weidinger, K.; Reif, J. Abundance and species richness of birds in two lowland riverine forests in Central Moravia—Possibilities for analyses of point-count data. Sylvia 2010, 46, 71–85. [Google Scholar]
- Freemark, K.E.; Collins, B. Landscape ecology of birds breeding in temperate forest fragments. In Ecology ans Conservation of Neotropical Migrant Landbird; Hagan, J.M., Johnston, D.W., Eds.; Smithsomian Institution Press: Washington, DC, USA, 1992; pp. 443–454. [Google Scholar]
- Pechanec, V.; Brus, J.; Kilianova, H.; Machar, I. Decision support tool for the evaluation of landscapes. Ecol. Inform. 2015, 30, 305–308. [Google Scholar] [CrossRef]
- Kornan, M. Comparison of bird assemblage structure between forest ecotone and interior of an alder swamp. Sylvia 2009, 45, 151–176. [Google Scholar]
- Truax, B.; Gagnon, D.; Lambert, F.; Fortier, J. Multiple-Use Zoning Model for Private Forest Owners in Agricultural Landscapes: A Case Study. Forests 2015, 6, 3614–3664. [Google Scholar] [CrossRef]
- Anic, I.; Mestrovic, S.; Matic, S. Important events in the history of forestry in Croatia. Sumar. List 2012, 136, 169–177. [Google Scholar]
- Kilianova, H.; Pechanec, V.; Brus, J.; Kirchner, K.; Machar, I. Analysis of the development of land use in the Morava River floodplain, with special emphasis on the landscape matrix. Morav. Geogr. Rep. 2017, 25. [Google Scholar] [CrossRef]
- Leso, P. Breeding bird communities of two succession stages of young oak forests. Sylvia 2003, 39, 67–78. [Google Scholar]
- Fulller, R.J. Bird Life of Woodland and Forest, 1st ed.; Cambridge University Press: Cambridge, UK, 2003; pp. 126–127. [Google Scholar]
- Glowacinski, Z. Succession of bird communities in the Niepolomice Forest (Southern Poland). Ekol. Polska 1975, 23, 231–263. [Google Scholar]
- Kropil, R. Struktura a Produkcia Ornitocenoz Vybranych Prirodnych Lesov Slovenska [Structure and Production of Bird Communities in Natural Forests in Slovakia]. Ph.D. Thesis, Technical University, Zvolen, Zvolen, Slovakia, 1993. [Google Scholar]
- Waliczky, Z. Bird community changes in different-aged oak forest stands in the Buda-hills. Ornis Hung. 1991, 1, 1–9. [Google Scholar]
- Wesolovski, T.; Rowinski, P.; Mitrus, C.; Czeszczewik, D. Breeding bird community of a primeval temperate forest (Bialowieza National Park, Poland) at the beginning of the 21st century. Acta Ornithol. 2006, 41, 55–70. [Google Scholar] [CrossRef]
- Tucker, G.M.; Evans, M.I. Habitats for Birds in Europe; Birdlife International: Cambridge, UK, 1997; pp. 1–464. ISBN 978-0-94688-8320. [Google Scholar]
- Stastny, K.; Bejcek, V.; Hudec, K. Atlas of Breeding Bird Distribution in the Czech Republic; Academia: Prague, Czech Republic, 2006; pp. 1–463. ISBN 80-86858-19-7. [Google Scholar]
- Reif, J.; Jiguet, F.; Šťastný, K. Habitat specialization of birds in the Czech Republic: Comparison of objective measures with expert opinion. Bird Study 2010, 57, 197–212. [Google Scholar] [CrossRef]
- Reif, J.; Storch, D.; Šímová, I. The effect of scale-dependent habitat gradients on the structure of bird assemblages in the Czech Republic. Acta Ornithol. 2008, 43, 197–206. [Google Scholar] [CrossRef]
- Machar, I. Attempt to summarize the problems: Is a sustainable management of floodplain forest geobiocenoses possible? In Biodiversity and Target Management of Floodplain Forests in the Morava River Basin (Czech Republic); Accession Number: WOS:000328003200016; Machar, I., Ed.; Palacky University: Olomouc, Czech Republic, 2010; pp. 189–226. ISBN 978-80-244-2530-6. [Google Scholar]
- Kovarik, P.; Kutal, M.; Machar, I. Sheep and wolves: Is the occurrence of large predators a limiting factor for sheep grazing in the Czech Carpathians? J. Nat. Conserv. 2014, 22, 5. [Google Scholar] [CrossRef]
- Machar, I.; Harmacek, J.; Vrublova, K.; Filippovova, J.; Brus, J. Biocontrol of common vole populations by avian predators versus rodenticide application. Pol. J. Ecol. 2017, 65, 434–444. [Google Scholar] [CrossRef]
- Janik, D.; Adam, D.; Vrska, T.; Hort, L.; Unar, P.; Kral, K.; Samonil, P.; Horal, D. Tree layer dynamics of the Cahnov-Soutok near-natural floodplain forest after 33 years (1973–2006). Eur. J. For. Res. 2008, 127, 337–345. [Google Scholar] [CrossRef]
- Machar, I. Protection of nature and landscapes in the Czech Republic Selected current issues and possibilities of their solution. In Ochrana Prirody a Krajiny v Ceske Republice, Vols I and II; Accession Number: WOS:000334387900001; Machar, I., Drobilova, L., Eds.; Palacky University: Olomouc, Czech Republic, 2012; p. 9-+. ISBN 978-80-244-3041-6. [Google Scholar]
- Machar, I.; Vlckova, V.; Bucek, A.; Vozenilek, V.; Salek, L.; Jerabkova, L. Modelling of Climate Conditions in Forest Vegetation Zones as a Support Tool for Forest Management Strategy in European Beech Dominated Forests. Forests 2017, 8, 82. [Google Scholar] [CrossRef]
- Machar, I.; Simon, J.; Rejsek, K.; Pechanec, V.; Brus, J.; Kilianova, H. Assessment of Forest Management in Protected Areas Based on Multidisciplinary Research. Forests 2016, 7, 285. [Google Scholar] [CrossRef]
- Spathelf, P. Sustainable Forest Management as a Model for Sustainbale Development: Conclusions Toward a Concrete Vision. Sustain. For. Manag. Chang. World Manag. For. Ecosyst. 2009, 19, 237–240. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Height | Density | Browsing % | |
|---|---|---|---|
| Height | - | −0.5653 1 | 0.4271 |
| Density | - | −0.5318 1 | |
| Browsing % | - |
| Bird Species | Nesting Guilds 1 | Study Plot | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |||||||
| DE 2 | DO 3 | DE | DO | DE | DO | DE | DO | DE | DO | ||
| Anthus trivialis | GN | 0.5 | 0.8 | 0.4 | 0.6 | 0.7 | 1.2 | 0.3 | 0.5 | 0.5 | 0.8 |
| Certhia brachydactyla | HN | 2.1 | 3.4 | 2.7 | 3.9 | 0.4 | 0.6 | 2.9 | 5.1 | 0.0 | 0.0 |
| Columba palumbus | CN | 1.6 | 2.6 | 2.2 | 3.2 | 2.6 | 4.3 | 0.6 | 1.1 | 2.0 | 3.2 |
| Cyanistes caeruleus | HN | 3.2 | 4.9 | 0.9 | 1.3 | 2.7 | 4.5 | 3.9 | 6.9 | 3.0 | 4.7 |
| Dendrocopos major | HN | 2.8 | 4.3 | 3.9 | 5.6 | 1.5 | 2.5 | 2.8 | 4.9 | 3.1 | 4.9 |
| Dendrocopos medius | HN | 0.0 | 0.0 | 0.7 | 1.0 | 0.0 | 0.0 | 0.5 | 0.9 | 0.6 | 0.9 |
| Erithacus rubecula | GN | 2.4 | 3.8 | 2.7 | 3.9 | 2.1 | 3.5 | 1.3 | 2.3 | 2.9 | 4.6 |
| Ficedula albicollis | HN | 7.0 | 10.8 | 4.3 | 6.2 | 7.6 | 12.7 | 7.9 | 13.9 | 3.8 | 6.0 |
| Fringilla coelebs | CN | 4.3 | 6.6 | 3.2 | 4.6 | 3.9 | 6.5 | 1.9 | 3.4 | 2.7 | 4.2 |
| Garrulus glandarius | CN | 0.6 | 0.9 | 0.9 | 1.3 | 0.5 | 0.8 | 1.2 | 2.0 | 0.7 | 1.1 |
| Muscicapa striata | HN | 0.6 | 0.9 | 1.1 | 1.7 | 1.3 | 2.3 | 0.9 | 1.6 | 0.5 | 0.8 |
| Oriolus oriolus | CN | 1.8 | 2.8 | 1.1 | 1.7 | 0.7 | 1.1 | 1.1 | 1.9 | 1.6 | 2.5 |
| Parus major | HN | 10.3 | 16.1 | 7.1 | 10.2 | 8.2 | 13.7 | 8.9 | 15.7 | 9.6 | 15.1 |
| Phylloscopus collybita | GN | 2.2 | 3.4 | 2.9 | 4.2 | 2.2 | 3.7 | 3.7 | 6.5 | 1.8 | 2.8 |
| Picus viridis | HN | 0.3 | 0.4 | 0.9 | 1.3 | 0.0 | 0.0 | 1.0 | 1.8 | 0.0 | 0.0 |
| Poecile palustris | HN | 0.9 | 1.4 | 0.2 | 0.3 | 0.7 | 1.2 | 0.8 | 1.4 | 0.6 | 0.9 |
| Sitta europaea | HN | 5.3 | 8.2 | 6.7 | 9.7 | 5.1 | 8.5 | 6.9 | 12.2 | 5.0 | 7.9 |
| Steptopelia turtur | CN | 0.8 | 1.2 | 1.5 | 2.2 | 0.6 | 1.0 | 0.6 | 1.1 | 0.9 | 1.4 |
| Sturnus vulgaris | HN | 17.2 | 26.6 | 19.0 | 27.5 | 15.4 | 25.7 | 9.1 | 16.1 | 18.3 | 28.8 |
| Turdus philomelos | CN | 0.6 | 0.9 | 0.5 | 0.7 | 0.6 | 0.6 | 0.4 | 0.7 | 0.7 | 1.1 |
| Turdus pilaris | CN | 0.0 | 0.0 | 6.2 | 8.9 | 3.1 | 5.2 | 0.0 | 0.0 | 5.3 | 8.3 |
| Total of DE and DO | 64.5 | 100 | 69.1 | 100 | 59.9 | 100 | 56.7 | 100 | 63.6 | 100 | |
| Total of bird species | 19 | 21 | 19 | 20 | 19 | ||||||
| Source 1 | Size of Study Plot (ha) | Deer Density (Individuals/1000 ha) | Species Richness of Nesting Birds in Study Plot | Density of Birds (Nesting Pairs/10 ha) | |
|---|---|---|---|---|---|
| Capreolus capreolus | Dama dama | ||||
| Results of this study in LPPLA (study plot 1) | 1 | 63 | 11 | 19 | 64.5 |
| Results of this study in LPPLA (study plot 2) | 1 | 63 | 11 | 21 | 69.1 |
| Results of this study in LPPLA (study plot 3) | 1 | 98 | 63 | 19 | 59.9 |
| Results of this study in LPPLA (study plot 4) | 1 | 98 | 63 | 20 | 56.7 |
| Results of this study in LPPLA (study plot 5) | 1 | 98 | 63 | 19 | 63.6 |
| [41] | 15 | 21 | 5 | 39 | 161 |
| [42] | 5 | 21 | 5 | 48 | 79 |
| [43] | 9 | 39 | - | 44 | 39 |
| [44] | 10 | 18 | 15 | 37 | 177 |
| [45] | 12 | 44 | - | 28 | 93 |
| [46] locality Bosin | 32 | 32 | 3 | 44 | 103 |
| [46] locality Dubno | 51 | 29 | 3 | 40 | 101 |
| [46] locality Choltice | 52 | 37 | 3 | 42 | 109 |
| [46] locality Zbytka | 43 | 40 | 3 | 41 | 80.5 |
| [47] locality Panensky les I | 9 | 35 | - | 31 | 139 |
| [47] locality Panensky les II | 9 | 35 | - | 36 | 118 |
| [48] locality Sargoun | 12 | 21 | 5 | 33 | 135 |
| [48] locality Vrapac | 13 | 35 | 11 | 31 | 126 |
| [49] | 10 | 20 | 3 | 25 | 113 |
| [50] | 10 | 50 | 11 | 23 | 92 |
| [51] | 22 | 41 | 5 | 36 | 112 |
| [52] | 10 | 36 | - | 23 | 102 |
| [53] | 12 | ? | - | 30 | 270 |
| [54] | 5 | 33 | 3 | 30 | 174 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Machar, I.; Cermak, P.; Pechanec, V. Ungulate Browsing Limits Bird Diversity of the Central European Hardwood Floodplain Forests. Forests 2018, 9, 373. https://doi.org/10.3390/f9070373
Machar I, Cermak P, Pechanec V. Ungulate Browsing Limits Bird Diversity of the Central European Hardwood Floodplain Forests. Forests. 2018; 9(7):373. https://doi.org/10.3390/f9070373
Chicago/Turabian StyleMachar, Ivo, Petr Cermak, and Vilem Pechanec. 2018. "Ungulate Browsing Limits Bird Diversity of the Central European Hardwood Floodplain Forests" Forests 9, no. 7: 373. https://doi.org/10.3390/f9070373
APA StyleMachar, I., Cermak, P., & Pechanec, V. (2018). Ungulate Browsing Limits Bird Diversity of the Central European Hardwood Floodplain Forests. Forests, 9(7), 373. https://doi.org/10.3390/f9070373