The Performance of Five Willow Cultivars under Different Pedoclimatic Conditions and Rotation Cycles


Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Sites
2.2. Willow Varieties and Experimental Setup
2.3. Measurement and Sampling
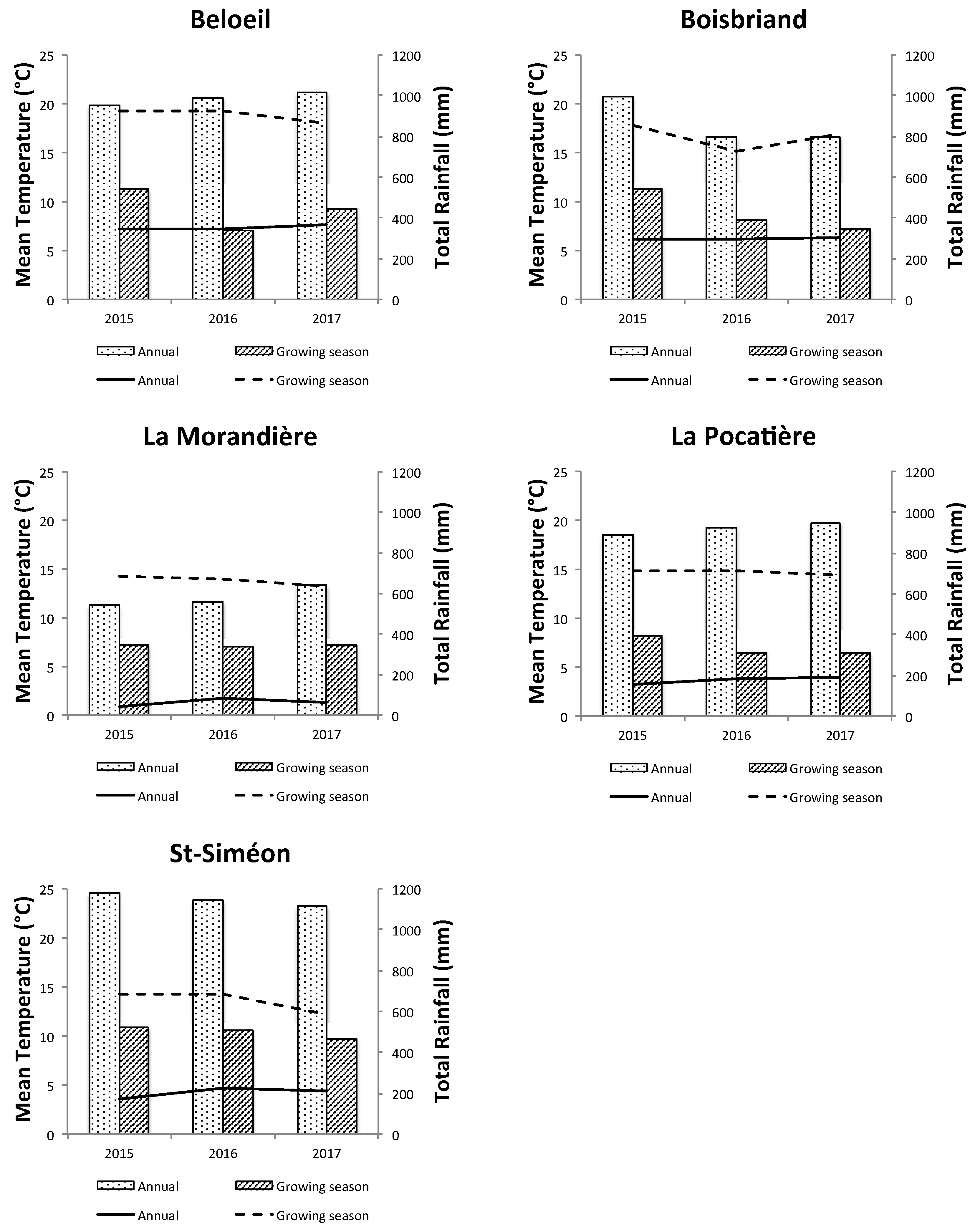
2.4. Meteorological Conditions
2.5. Data Analysis
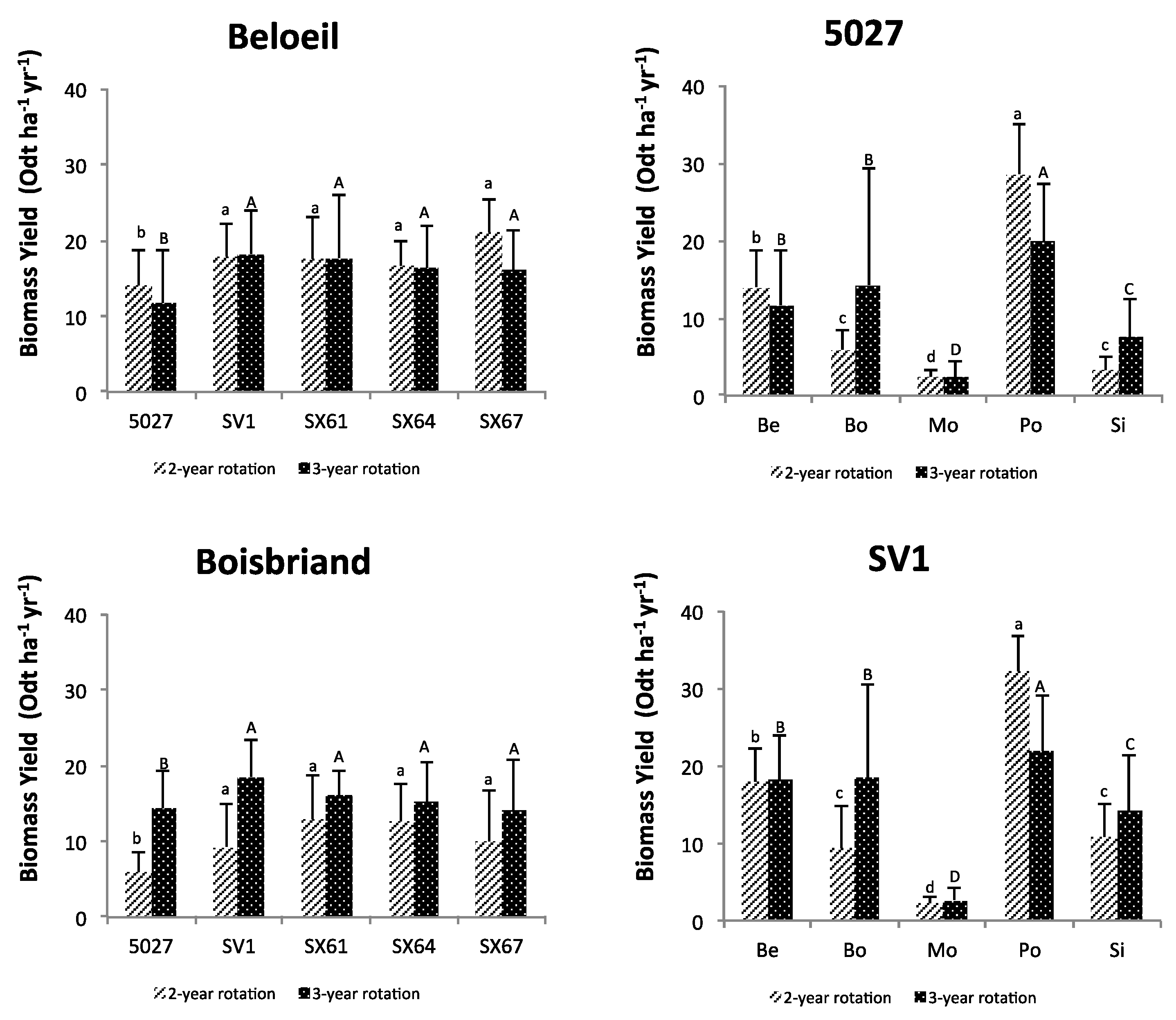
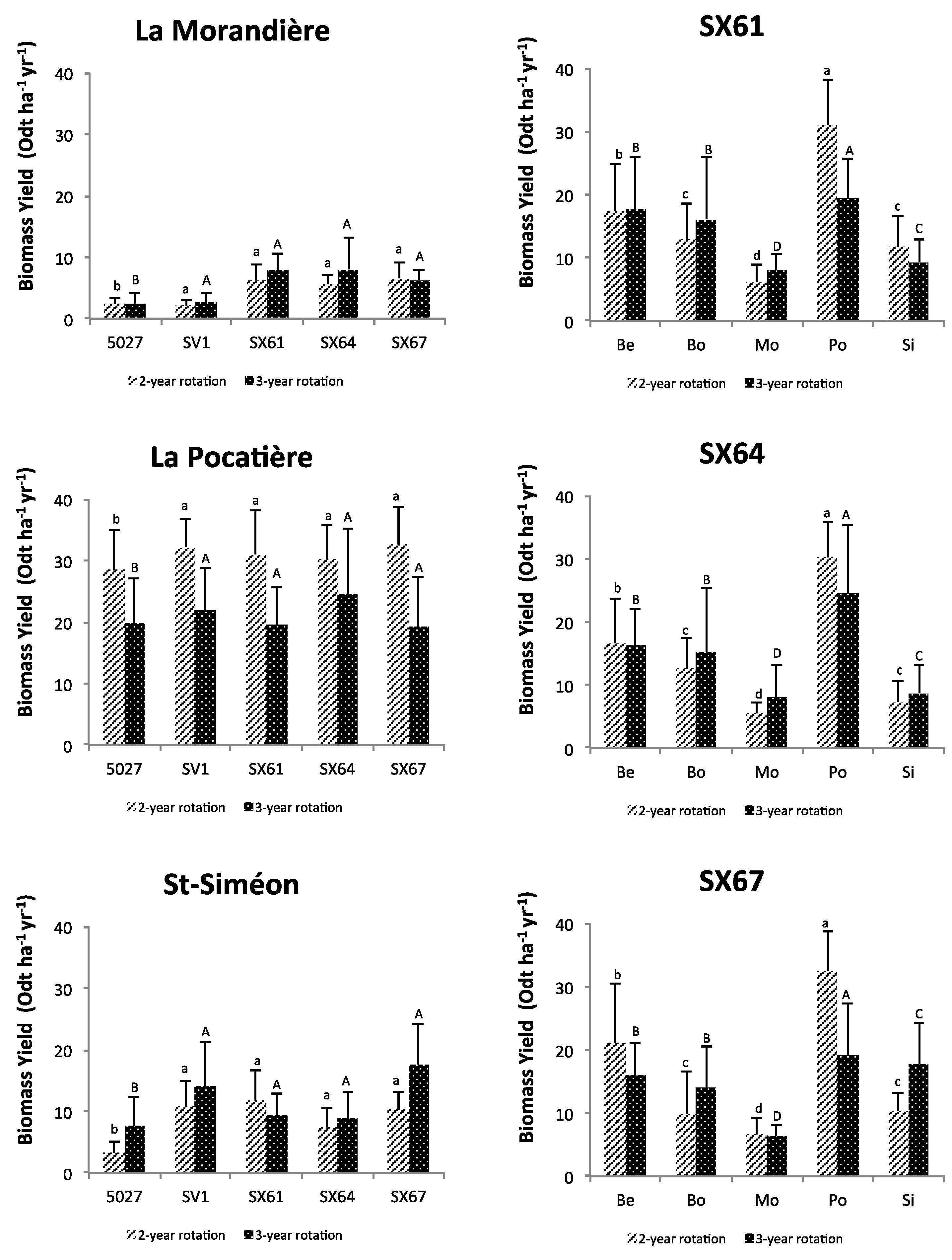
3. Results
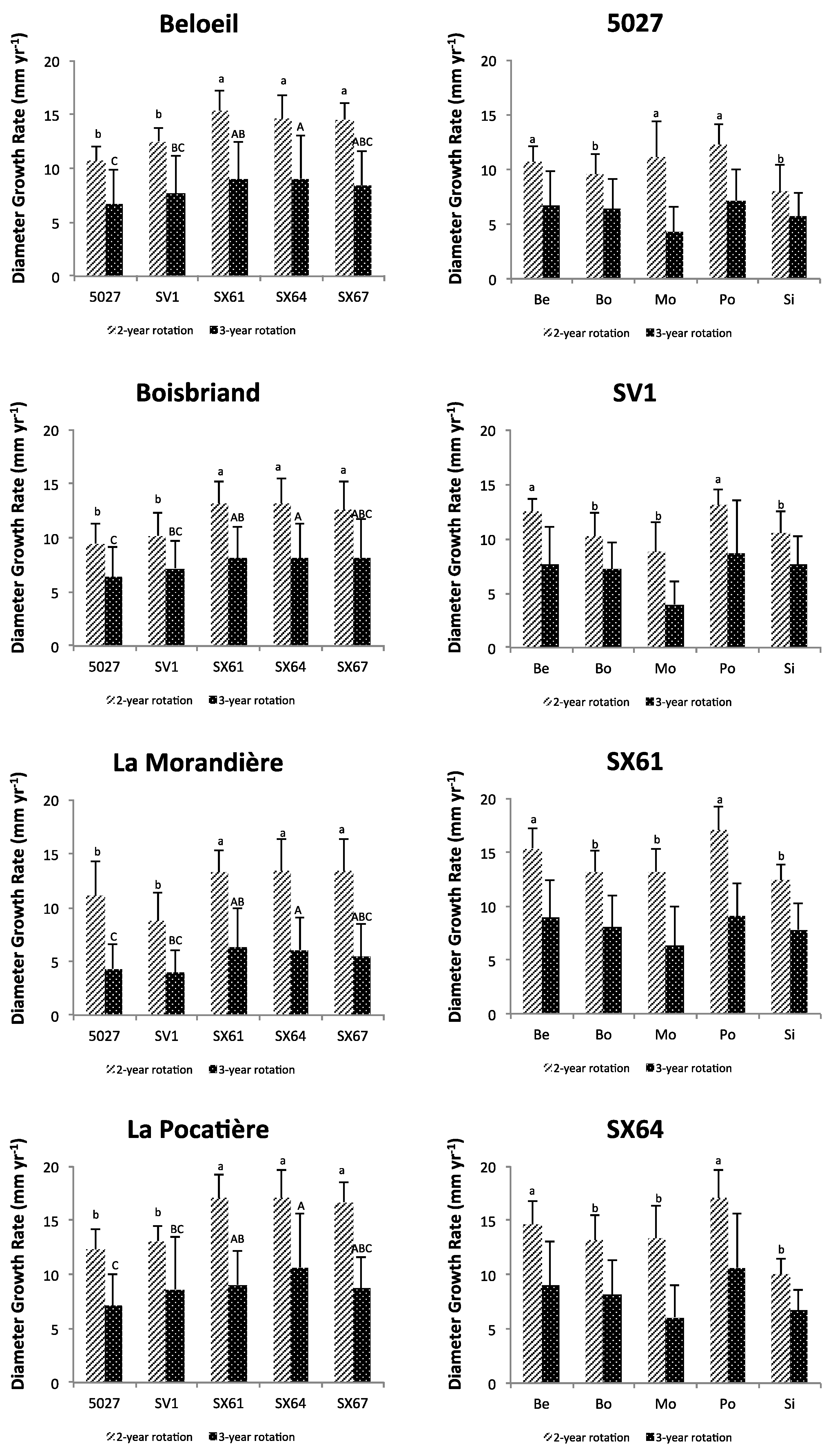
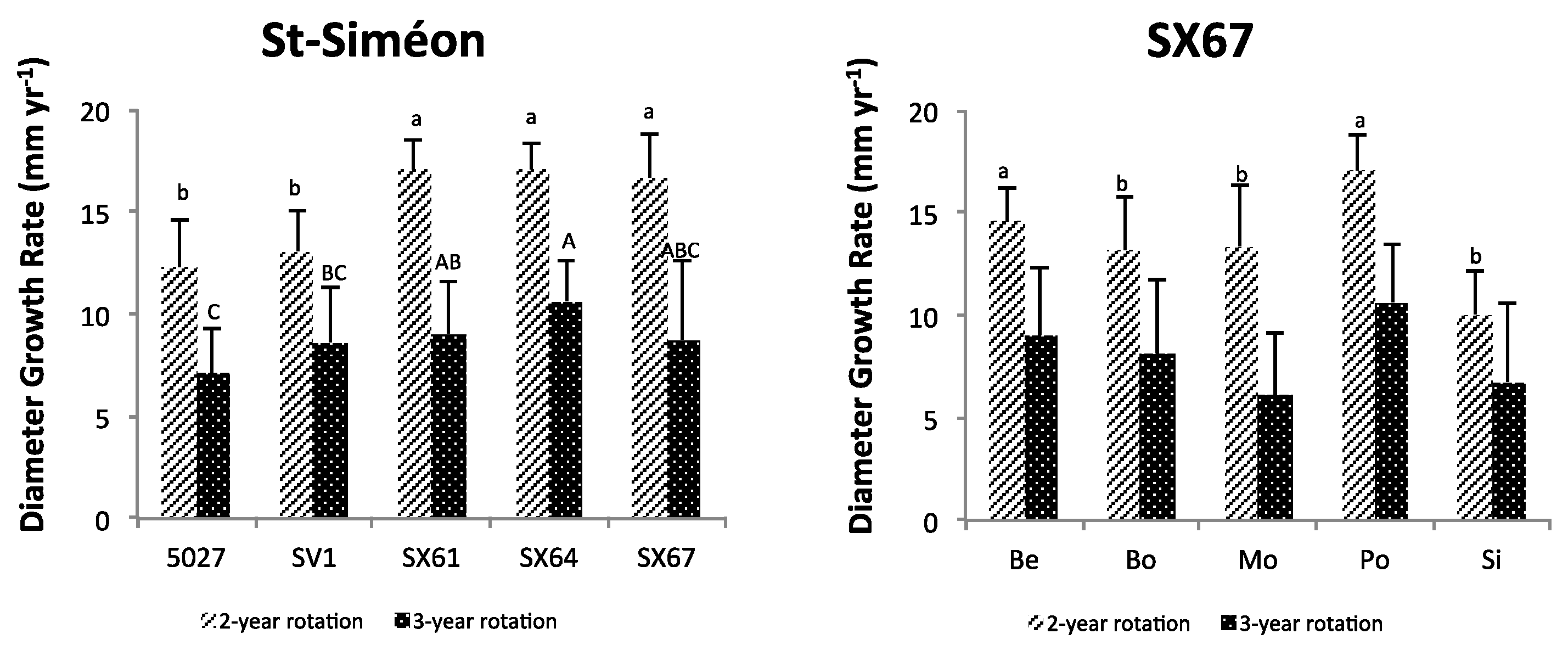
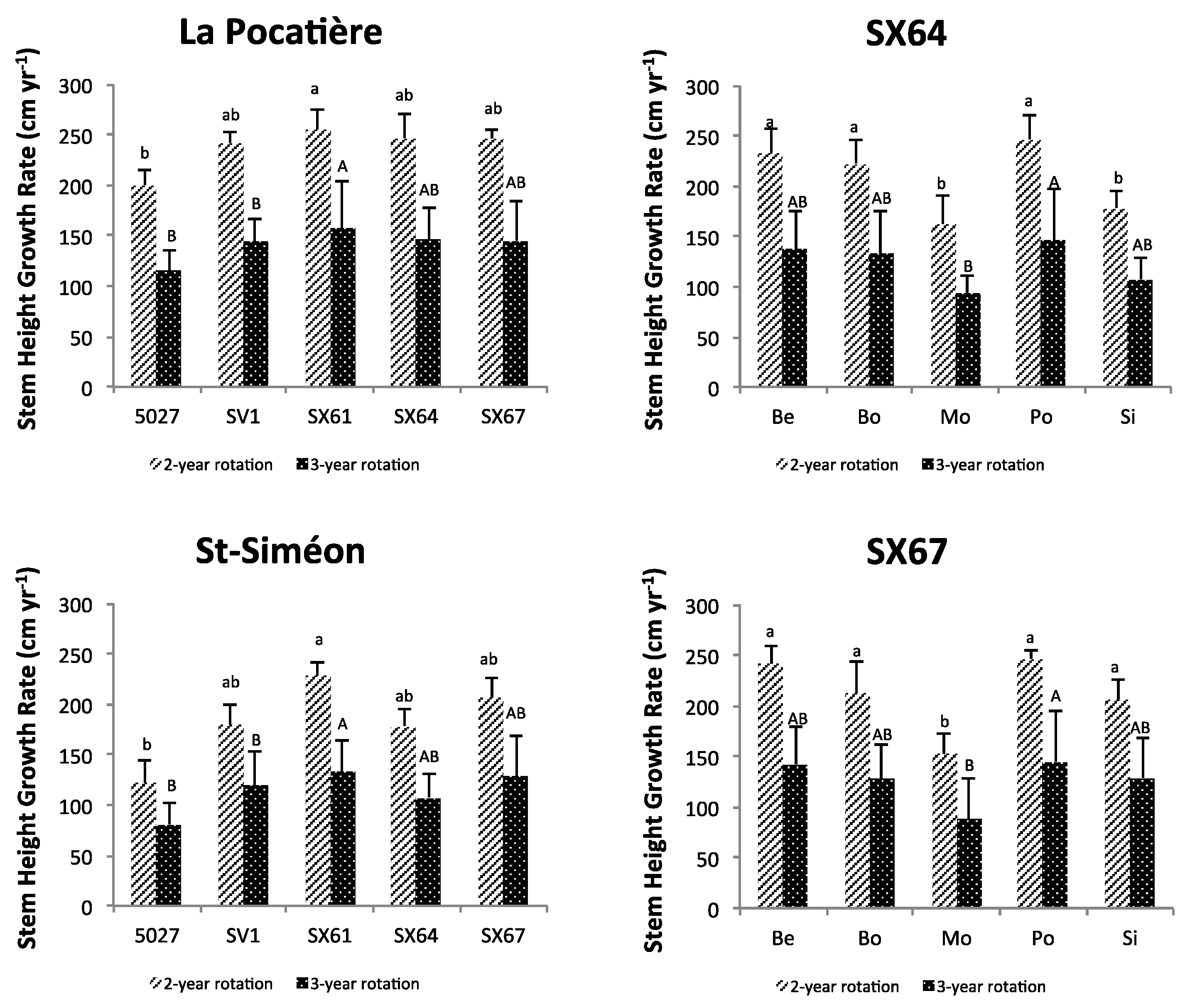
3.1. Plant Growth and Biomass Yield
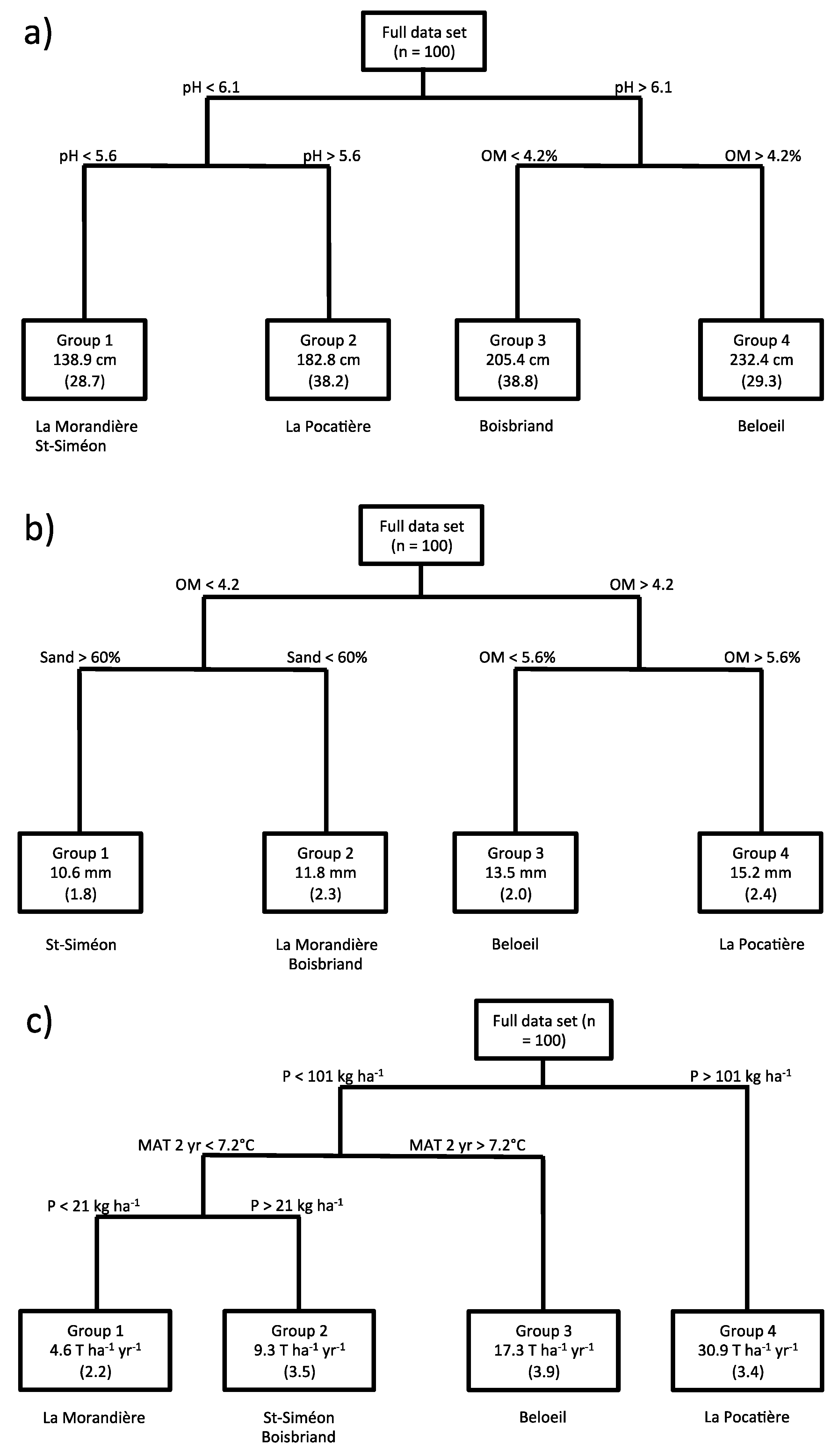
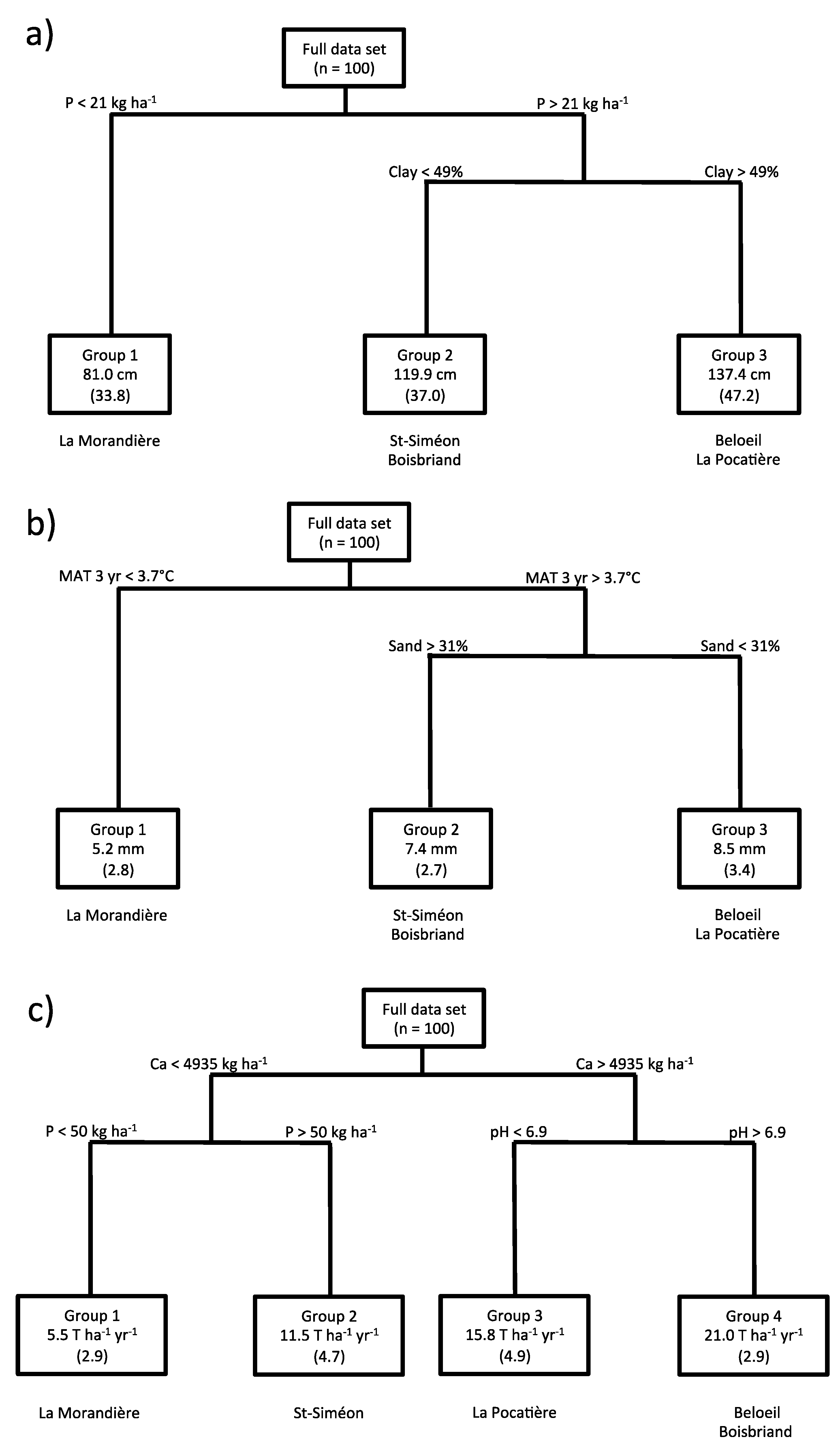
3.2. Relating Plant Height and Diameter Annual Increment and Annual Yield to Climate and Soil Variables
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Hinchee, M.; Rottmann, W.; Mullinax, L.; Zhang, C.; Chang, S.; Cunningham, M.; Pearson, L.; Nehra, N. Short-rotation woody crops for bioenergy and biofuels applications. In Vitro Cell. Dev. Biol. Plant 2009, 45, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Drew, A.P.; Zsuffa, L.; Mitchell, C.P. Terminology relating to woody plant biomass and its production. Biomass 1987, 12, 79–82. [Google Scholar] [CrossRef]
- Auclair, D.; Bouvarel, L. Intensive or extensive cultivation of short rotation hybrid poplar coppice on forest land. Bioresour. Technol. 1992, 42, 53–59. [Google Scholar] [CrossRef]
- Kenney, W.A.; Sennerby-Forsse, L.; Layton, P. A review of biomass quality research relevant to the use of poplar and willow for energy conversion. Biomass 1990, 21, 163–188. [Google Scholar] [CrossRef]
- Sims, R.E.H.; Senelwa, K.; Maiava, T.; Bullock, T. Eucalyptus species for biomass energy in Nex Zealand-I: Growth screening trials at first harvest. Biomass Bioenergy 1999, 16, 199–205. [Google Scholar] [CrossRef]
- Grünewald, H.; Böhm, C.; Quinkenstein, A.; Grundmann, P.; Jörg Eberts, J.; von Wühlisch, G. Robinia pseudoacacia L.: A Lesser Known Tree Species for Biomass Production. Bioenergy Res. 2009, 2, 123–133. [Google Scholar] [CrossRef]
- Rytter, L.; Šlapokas, T.; Granhall, U. Woody biomass and litter production of fertilized grey alder plantations on a low-humified peat bog. For. Ecol. Manag. 1989, 28, 161–176. [Google Scholar] [CrossRef]
- Lopez, F.; Garcia, M.M.; Ynez, R.; Tapias, R.; Fernàndez, M.; Diaz, M.J. Leucaena species valoration for biomass and paper production in 1 and 2 year harvest. Bioresour. Technol. 2008, 99, 4846–4853. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.P. Ecophysiology of short rotation forest crops. Biomass Bioenergy 1992, 2, 25–37. [Google Scholar] [CrossRef]
- Dimitriou, I.; Baum, C.; Baum, S.; Busch, G.; Schulz, U.; Köhn, J.; Lamersdorf, N.; Leinweber, P.; Aronsson, P.; Weih, M.; et al. The impact of Short Rotation Coppice (SRC) cultivation on the environment. vTI Agric. For. Res. 2009, 59, 159–162. [Google Scholar]
- Park, B.B.; Yanai, R.D.; Sahm, J.M.; Ballard, B.D.; Abrahamson, L.P. Wood Ash Effects on Soil Solution and Nutrient Budgets in A Willow Bioenergy Plantation. Water Air Soil Pollut. 2004, 159, 209–224. [Google Scholar] [CrossRef]
- Labrecque, M.; Teodorescu, T.I.; Daigle, S. Biomass productivity and wood energy of salix species after 2 years growth in SRIC fertilized with wastewater sludge. Biomass Bioenergy 1997, 12, 409–417. [Google Scholar] [CrossRef]
- Guidi Nissim, W.; Cincinelli, A.; Martellini, T.; Alvisi, L.; Palm, E.; Mancuso, S.; Azzarello, E. Phytoremediation of sewage sludge contaminated by trace elements and organic compounds. Environ. Res. 2018, 164, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, A.; Gasser, M.O.; Labrecque, M. Pig slurry as fertilizer on willow plantation. Biomass Bioenergy 2011, 35, 4165–4173. [Google Scholar] [CrossRef]
- Guidi Nissim, W.; Voicu, A.; Labrecque, M. Willow short-rotation coppice for treatment of polluted groundwater. Ecol. Eng. 2014, 62, 102–114. [Google Scholar] [CrossRef]
- Dimitriou, I.; Aronsson, P. Wastewater and sewage sludge application to willows and poplars grown in lysimeters—Plant response and treatment efficiency. Biomass Bioenergy 2011, 35, 161–170. [Google Scholar] [CrossRef]
- Börjesson, P.; Berndes, G. The prospects for willow plantations for wastewater treatment in Sweden. Biomass Bioenergy 2006, 30, 428–438. [Google Scholar] [CrossRef]
- Guidi Nissim, W.; Pitre, F.E.; Teodorescu, T.I.; Labrecque, M. Long-term biomass productivity of willow bioenergy plantations maintained in southern Quebec, Canada. Biomass Bioenergy 2013, 56, 361–369. [Google Scholar] [CrossRef]
- Amichev, B.Y.; Hangs, R.D.; Konecsni, S.M.; Stadnyk, C.N.; Volk, T.A.; Bélanger, N.; Vujanovic, V.; Schoenau, J.J.; Moukoumi, J.; Van Rees, K.C.J. Willow Short-Rotation Production Systems in Canada and Northern United States: A Review. Soil Sci. Soc. Am. J. 2014, 78, S168–S181. [Google Scholar] [CrossRef]
- Lafleur, B.; Lalonde, O.; Labrecque, M. First-Rotation Performance of Five Short-Rotation Willow Cultivars on Different Soil Types and Along a Large Climate Gradient. BioEnergy Res. 2017, 10, 158–166. [Google Scholar] [CrossRef]
- Grime, J.P. Plant Strategies and Vegetation Processes; John Wiley & Sons, Ltd.: Chichester, UK; New York, NY, USA; Brisbane, Australia; Toronto, ON, Cadana, 1979. [Google Scholar]
- Fontana, M.; Labrecque, M.; Messier, C.; Courchesne, F.; Bélanger, N. Quantifying the effects of soil and climate on aboveground biomass production of Salix miyabeana SX67 in Quebec. New For. 2017, 48, 817–835. [Google Scholar] [CrossRef]
- Fontana, M.; Labrecque, M.; Messier, C.; Bélanger, N. Permanent site characteristics exert a larger influence than atmospheric conditions on leaf mass, foliar nutrients and ultimately aboveground biomass productivity of Salix miyabeana ‘SX67’. For. Ecol. Manag 2018, in press. [Google Scholar] [CrossRef]
- Willebrand, E.; Ledin, S.; Verwijst, T. Willow coppice systems in short rotation forestry; effects of plant spacing, rotation length and clonal disposition on biomass production. Biomass Bioenergy 1993, 4, 323–331. [Google Scholar] [CrossRef]
- Breiman, L.; Friedman, J.H.; Olshen, R.A.; Stone, C.J. Classification and Regression Trees; Chapman and Hall/CRC: Boca Raton, FL, USA, 1998. [Google Scholar]
- Labrecque, M.; Teodorescu, T.I. Field performances and biomass production of 12 willow and poplar clones in short-rotation coppice in southern Quebec (Canada). Biomass Bioenergy 2005, 29, 1–9. [Google Scholar] [CrossRef]
- Guidi, W.; Pitre, F.E.; Labrecque, M. Short-Rotation Coppice of Willows for the Production of Biomass in Eastern Canada. In Biomass Now—Sustainable Growth and Use; Matovic, M.D., Ed.; InTech: London, UK, 2013; Chapter 17; pp. 421–448. [Google Scholar]
- Larsen, S.; Jørgensen, U.; Lærke, P.E. Willow Yield Is Highly Dependent on Clone and Site. BioEnergy Res. 2014, 7, 1280–1292. [Google Scholar] [CrossRef]
- Mosseler, A.; Major, J.E.; Labrecque, M. Genetic by environment interactions of two North American Salix species assessed for coppice yield and components of growth on three sites of varying quality. Trees 2014, 28, 1401–1411. [Google Scholar] [CrossRef]
- Fabio, E.S.; Volk, T.A.; Miller, R.O.; Serapiglia, M.J.; Gauch, H.G.; Van Rees, K.C.J.; Hangs, R.D.; Amichev, B.Y.; Kuzovkina, Y.A.; Labrecque, M.; et al. Genotype × environment interaction analysis of North American shrub willow yield trials confirms superior performance of triploid hybrids. GCB Bioenergy 2016, 9, 445–459. [Google Scholar] [CrossRef]
- Aylott, M.J.; Casella, E.; Tubby, I.; Street, N.R.; Smith, P.; Taylor, G. Yield and spatial supply of bioenergy poplar and willow short‚ rotation coppice in the UK. New Phytol. 2008, 178, 358–370. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, J.M.; Evans, E.J.; Bilsborrow, P.E.; Wright, C.; Hewison, W.O.; Pilbeam, D.J. Yield of willow cultivars at different planting densities in a commercial short rotation coppice in the north of England. Biomass Bioenergy 2007, 31, 469–474. [Google Scholar] [CrossRef]
- Grenier, V.; Pitre, F.E.; Guidi Nissim, W.; Labrecque, M. Genotypic differences explain most of the response of willow cultivars to petroleum-contaminated soil. Trees 2015, 29, 871–881. [Google Scholar] [CrossRef]
- Mosseler, A.; Major, J.E. Coppice growth responses of two North American willows in acidic clay soils on coal mine overburden. Can. J. Plant Sci. 2014, 94, 1269–1279. [Google Scholar] [CrossRef]
- Stolarski, M.; Szczukowski, S.; Tworkowski, J.; Klasa, A. Productivity of seven clones of willow coppice in annual and quadrennial cutting cycles. Biomass Bioenergy 2008, 32, 1227–1234. [Google Scholar] [CrossRef]
- Thevathasan, N.; Coleman, B.; Zabek, L.; Ward, T.; Gordon, A. Agroforestry in Canada and its Role in Farming Systems. In Temperate Agroforestery Systems, 2nd ed.; Gordon, A., Newman, S.M., Coleman, B., Eds.; CABI: London, UK, 2018; Chapter 2; pp. 7–49. [Google Scholar]
- Kopp, R.F.; Abrahamson, L.P.; White, E.H.; Burns, K.F.; Nowak, C.A. Cutting cycle and spacing effects on biomass production by a willow clone in New York. Biomass Bioenergy 1997, 12, 313–319. [Google Scholar] [CrossRef]
- Coyle, D.R.; Aubrey, D.P.; Coleman, M.D. Growth responses of narrow or broad site adapted tree species to a range of resource availability treatments after a full harvest rotation. For. Ecol. Manag. 2016, 362, 107–119. [Google Scholar] [CrossRef]
- Hénault-Ethier, L.; Gomes, M.P.; Lucotte, M.; Smedbol, É.; Maccario, S.; Lepage, L.; Juneau, P.; Labrecque, M. High yields of riparian buffer strips planted with Salix miyabena ‘SX64’ along field crops in Québec, Canada. Biomass Bioenergy 2017, 105, 219–229. [Google Scholar] [CrossRef]
- Adnan, A.; Mavinic, D.S.; Koch, F.A. Pilot-scale study of phosphorus recovery through struvite crystallization-examining to process feasibility. J. Environ. Eng. Sci. 2003, 2, 315–324. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | pH | OM | P | K | Ca | Mg | Sand | Silt | Clay | Texture | Soil Type |
|---|---|---|---|---|---|---|---|---|---|---|---|
| % | (kg ha−1) | % | |||||||||
| Beloeil | 7.3 | 4.2 | 31 | 729 | 7570 | 2490 | 10 | 40 | 50 | Silty clay | Gleysols |
| Boisbriand | 6.9 | 4 | 21 | 166 | 4935 | 1120 | 31 | 33 | 36 | Clayey loam | Brunisols |
| La Morandière | 5.6 | 2.9 | 20 | 410 | 2796 | 1261 | 1 | 36 | 63 | Clay | Luvisols |
| La Pocatière | 6.1 | 5.6 | 101 | 475 | 5889 | 1138 | 20 | 31 | 49 | Clay | Gleysols |
| St-Siméon | 5.2 | 2.1 | 50 | 155 | 1662 | 72 | 60 | 20 | 20 | Sandy loam | Brunisols |
| Source | Nparm | DF | DFDen | F Ratio | Prob. > F |
| Site | 4 | 4 | 50 | 99.0831 | <0.0001 |
| Cultivar | 4 | 4 | 50 | 7.8034 | <0.0001 |
| Site × Cultivar | 16 | 16 | 50 | 1.752 | 0.0668 |
| Rotation | 1 | 1 | 50 | 0.0422 | 0.838 |
| Site × Rotation | 4 | 4 | 50 | 9.9017 | <0.0001 |
| Cultivar × Rotation | 4 | 4 | 50 | 0.3704 | 0.8286 |
| Site × Cultivar × Rotation | 16 | 16 | 50 | 0.6305 | 0.8437 |
| Least Square Mean | |||||
| Site | Rotation | ||||
| Two-year | Three-year | ||||
| Beloeil | 16.729656 | A | 15.294729 | A | |
| Boisbriand | 9.270192 | B | 14.119425 | A | |
| La Morandière | 4.1636872 | B | 4.8795649 | A | |
| La Pocatière | 30.626091 | A | 20.151016 | B | |
| La Pocatière | 10.681216 | A | 7.98445 | B | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guidi Nissim, W.; Lafleur, B.; Labrecque, M. The Performance of Five Willow Cultivars under Different Pedoclimatic Conditions and Rotation Cycles. Forests 2018, 9, 349. https://doi.org/10.3390/f9060349
Guidi Nissim W, Lafleur B, Labrecque M. The Performance of Five Willow Cultivars under Different Pedoclimatic Conditions and Rotation Cycles. Forests. 2018; 9(6):349. https://doi.org/10.3390/f9060349
Chicago/Turabian StyleGuidi Nissim, Werther, Benoit Lafleur, and Michel Labrecque. 2018. "The Performance of Five Willow Cultivars under Different Pedoclimatic Conditions and Rotation Cycles" Forests 9, no. 6: 349. https://doi.org/10.3390/f9060349
APA StyleGuidi Nissim, W., Lafleur, B., & Labrecque, M. (2018). The Performance of Five Willow Cultivars under Different Pedoclimatic Conditions and Rotation Cycles. Forests, 9(6), 349. https://doi.org/10.3390/f9060349