Evaluating Adaptive Management Options for Black Ash Forests in the Face of Emerald Ash Borer Invasion


Abstract
1. Introduction
2. Ecological Context of Black Ash Wetlands
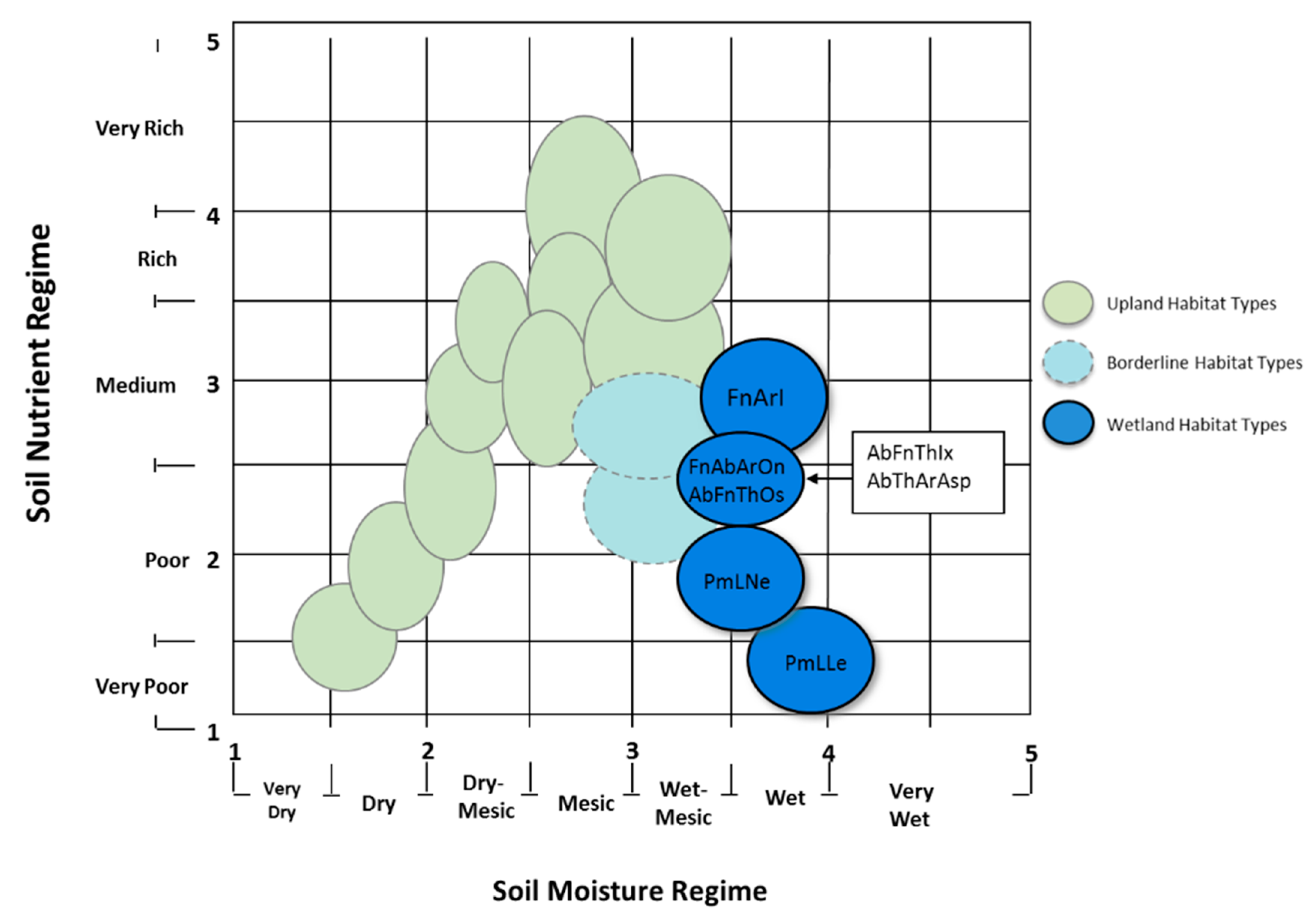
2.1. Black Ash Forest Habitat Types
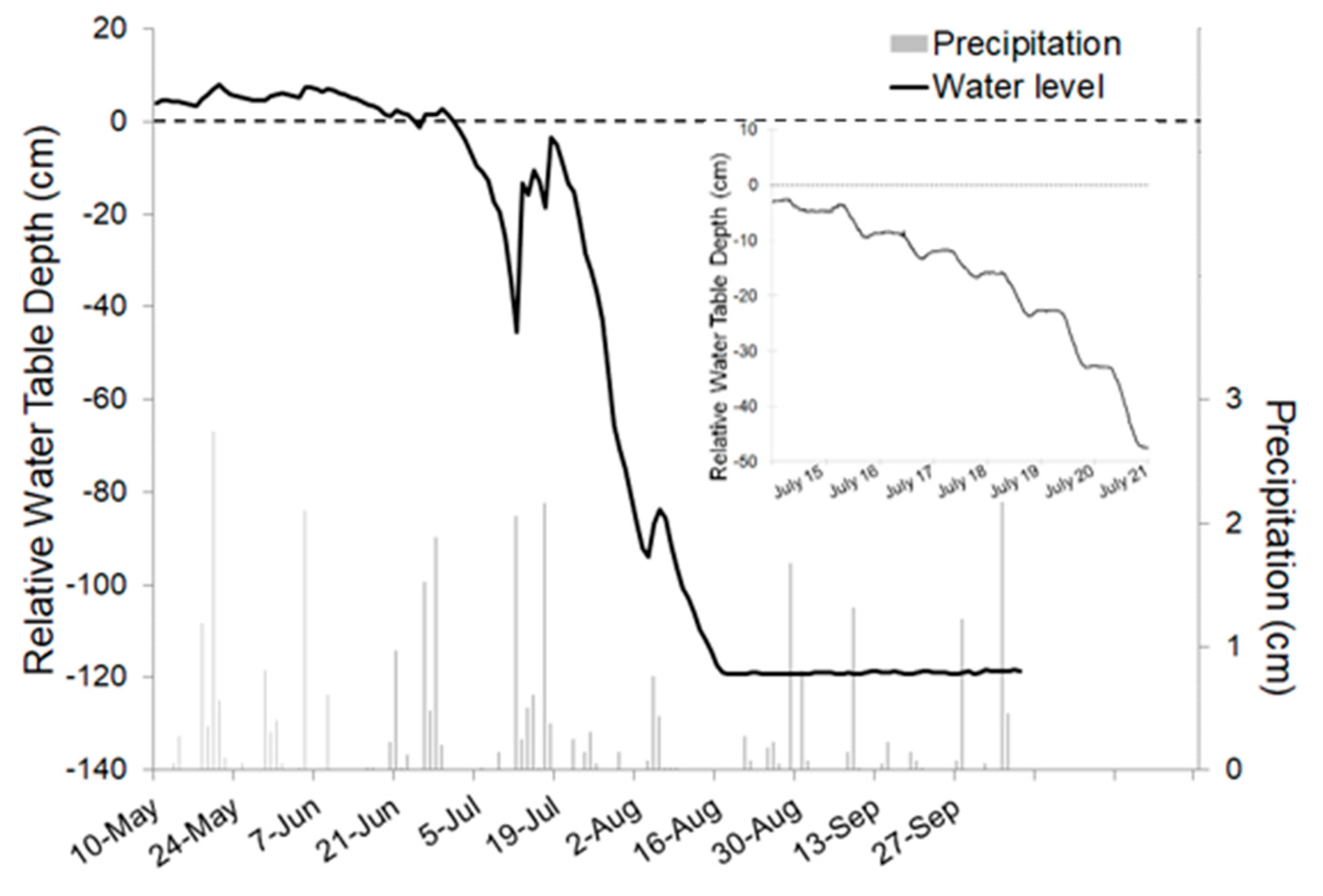
2.2. Hydrologic Setting of Black Ash Forests
2.3. Black Ash Decline and Natural Regeneration Potential for Non-Ash Species
3. Management Experience with Strategies for Increasing Resilience of Black Ash Wetlands to EAB
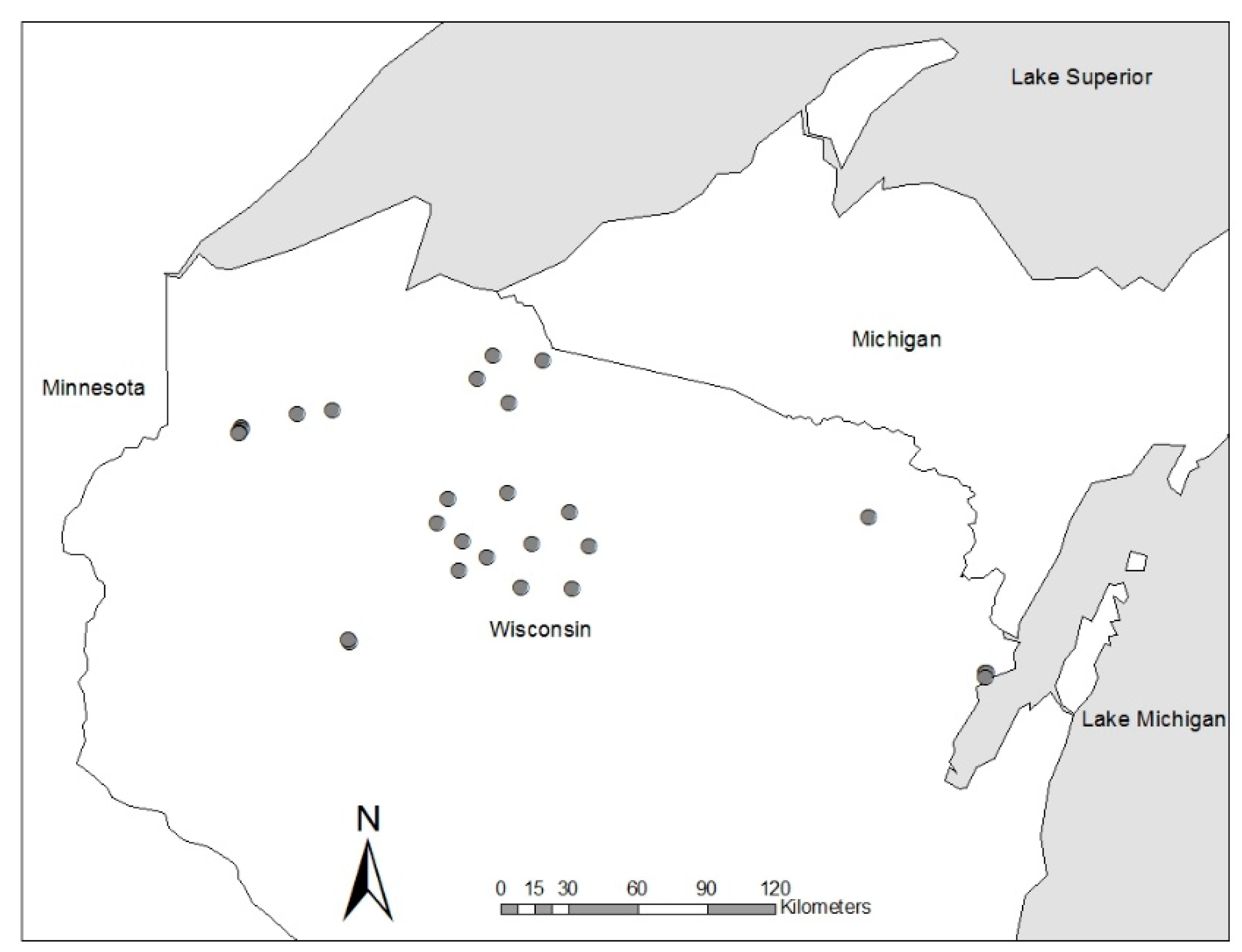
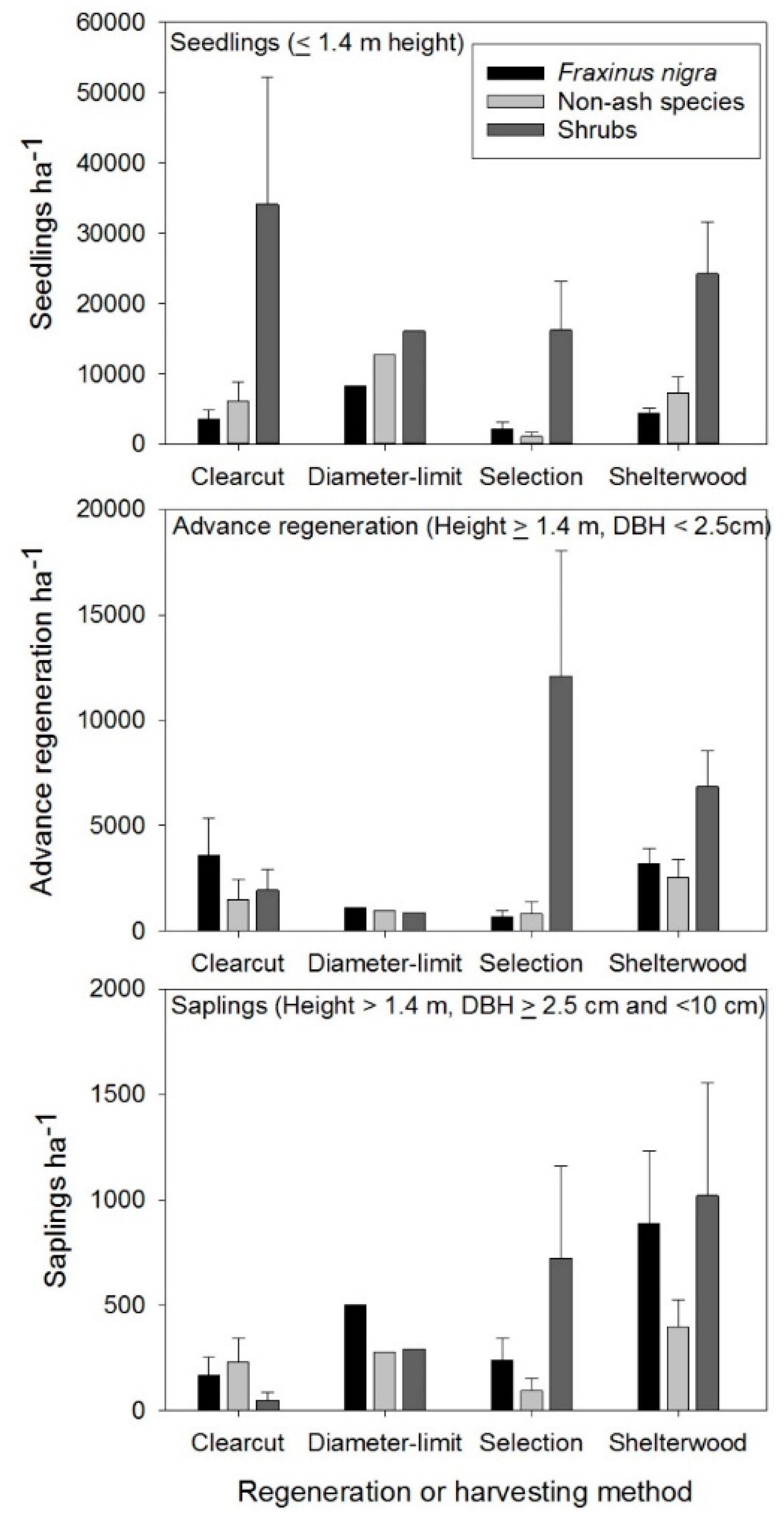
3.1. Research Trials Examining Artificial Regeneration Options to Increase Non-Ash Components
3.2. Encouraging Natural Regeneration of Non-Ash Species
3.3. Decision Support Tools for Guiding Adaptive Management in Black Ash: The Wisconsin DNR Checklist for Evaluating Lowland Ash Stands
3.4. Restoring Forest Cover to Areas Impacted by EAB
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Ayres, M.P.; Lombardero, M.J. Forest pests and their management in the Anthropocene. Can. J. For. Res. 2017, 48, 292–301. [Google Scholar]
- Lovett, G.M.; Weiss, M.; Liebhold, A.M.; Holmes, T.P.; Leung, B.; Lambert, K.F.; Orwig, D.A.; Campbell, F.T.; Rosenthal, J.; McCullough, D.G.; et al. Nonnative forest insects and pathogens in the United States: Impacts and policy options. Ecol. Appl. 2016, 26, 1437–1455. [Google Scholar] [CrossRef] [PubMed]
- Ellison, A.M.; Bank, M.S.; Clinton, B.D.; Colburn, E.A.; Elliott, K.; Ford, C.R.; Foster, D.R.; Kloeppel, B.D.; Knoepp, J.D.; Lovett, G.M.; et al. Loss of foundation species: Consequences for the structure and Dynamics of forested ecosystems. Front. Ecol. Environ. 2005, 3, 479–486. [Google Scholar]
- Waring, K.M.; O’Hara, K.L. Silvicultural strategies in forest ecosystems affected by introduced pests. For. Ecol. Manag. 2005, 209, 27–41. [Google Scholar] [CrossRef]
- Youngquist, M.B.; Eggert, S.L.; D’Amato, A.W.; Palik, B.J.; Slesak, R.A. Potential effects of foundation species loss on Wetland Communities: A case study of black ash wetlands threatened by emerald ash borer. Wetlands 2017, 37, 787–799. [Google Scholar]
- Valachovic, Y.; Lee, C.; Marshall, J.; Scanlon, H. Wildland management of Phytophthora ramorum in northern California forests. In Proceedings of the Sudden Oak Death Science Symposium, Santa Rosa, CA, USA, 5–9 March 2007; Forest Service: Albany, CA, USA, 2008; pp. 305–312. [Google Scholar]
- Smith, M.T.; Turgeon, J.J.; de Groot, P.; Gasman, B. Asian longhorned beetle Anoplophora glabripennis (Motschulsky): Lessons learned and opportunities to improve the process of eradication and management. Am. Entomol. 2009, 55, 21–25. [Google Scholar] [CrossRef]
- McCullough, D.G.; Siegert, N.W. Estimating potential emerald ash borer (Coleoptera: Buprestidae) populations using ash inventory data. J. Econ. Entomol. 2007, 100, 1577–1586. [Google Scholar] [PubMed]
- McCullough, D.G.; Siegert, N.W.; Bedford, J. Slowing ash mortality: A potential strategy to slam emerald ash borer in outlier sites. In Proceedings of the 20th U.S. Department of Agriculture Interagency Research Forum on Invasive Species, Annapolis, MD, USA, 13–16 January 2009; McManus, K.A., Gottschalk, K.W., Eds.; General Technical Report NRS-P-51. U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2009; pp. 44–46. [Google Scholar]
- Kizlinski, M.L.; Orwig, D.A.; Cobb, R.C.; Foster, D.R. Direct and indirect ecosystem consequences of an invasive pest on forests dominated by eastern hemlock. J. Biogeogr. 2002, 29, 1489–1503. [Google Scholar] [CrossRef]
- MacFarlane, D.W.; Meyer, S.P. Characteristics and distribution of potential ash tree hosts for emerald ash borer. For. Ecol. Manag. 2005, 213, 15–24. [Google Scholar]
- Knight, K.S.; Brown, J.P.; Long, R.P. Factors affecting the survival of ash (Fraxinus spp.) trees infested by emerald ash borer (Agrilus planipennis). Biol. Invasions 2013, 15, 371–383. [Google Scholar]
- Erdmann, G.G.; Crow, T.R.; Peterson, R.M., Jr.; Wilson, C.D. Managing Black Ash in the Lake States; General Technical Report, NC-115; USDA Forest Service: Washington, DC, USA, 1987; 12p.
- Slesak, R.A.; Lenhart, C.F.; Brooks, K.N.; D’Amato, A.W.; Palik, B.J. Water table response to harvesting and simulated emerald ash borer mortality in black ash wetlands in Minnesota, USA. Can. J. For. Res. 2014, 44, 961–968. [Google Scholar] [CrossRef]
- Herms, D.A.; McCullough, D.G. Emerald Ash Borer Invasion of North America: History, Biology, Ecology, Impacts, and Management. Ann. Rev. Entomol. 2014, 59, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Kashian, D.M. Sprouting and seed production may promote persistence of Green ash in the presence of the emerald ash borer. Ecosphere 2016, 7, e01332. [Google Scholar] [CrossRef]
- Iverson, L.; Knight, K.S.; Prasad, A.; Herms, D.A.; Matthews, S.; Peters, M.; Smith, A.; Hartzler, D.M.; Long, R.; Almendinger, J. Potential Species Replacements for Black Ash (Fraxinus nigra) at the Confluence of Two Threats: Emerald Ash Borer and a Changing Climate. Ecosystems 2016, 19, 248–270. [Google Scholar] [CrossRef]
- USDA Forest Service Forest Inventory and Analysis (USDA FIA). Forest Inventory and Analysis Database; U.S. Department of Agriculture, Forest Service, Northern Research Station: St. Paul, MN, USA, 2018. Available online: http://apps.fs.fed.us/fiadb-downloads/datamart.html (accessed on 6 April 2018).
- Weber, M. Factors Affecting Natural Tree Reproduction in Black Ash Communities in Northern Minnesota; MS Plan B Paper; College of Forestry, University of Minnesota: St. Paul, MN, USA, 1985; 58p. [Google Scholar]
- Kurmis, V.; Kim, J.H. Black Ash Stand Composition and Structure in Carlton County, Minnesota; Staff Paper Series Number 69; University of Minnesota Agricultural Experiment Station: St. Paul, MN, USA, 1989; 25p. [Google Scholar]
- Sterrett, W.D. The Ashes: Their Characteristics and Management; USDA Bulletin Number 299; US Department of Agriculture: Washington, DC, USA, 1915; 88p.
- Eyre, F.H. Forest Cover Types of the United States and Canada; Society of American Foresters: Washington, DC, USA, 1980; 148p. [Google Scholar]
- Michigan Department of Natural Resources (MIDNR). Forest Management Guide for Lowland Hardwoods Cover Type; Department of Natural Resources, Forest Management Division: Lansing, MI, USA, 1993; 4p.
- Kotar, J.; Kovach, J.A.; Burger, T.L. A Guide to Forest Communities and Habitat Types of Northern Wisconsin; Department of Forest Ecology and Management, University of Wisconsin-Madison: Madison, WI, USA, 2002. [Google Scholar]
- Aaseng, N.E. Field Guide to the Native Plant Communities of Minnesota: The Laurentian Mixed Forest Province; Minnesota Department of Natural Resources: St. Paul, MN, USA, 2003.
- Tardif, J.; Bergeron, Y. Population dynamics of Fraxinus nigra in response to flood-level variations, in Northwestern Quebec. Ecol. Monogr. 1999, 69, 107–125. [Google Scholar] [CrossRef]
- Kotar, J.; Burger, T.L. Wetland Forest Habitat Type Classification System for Northern Wisconsin; PUB-FR-627; Wisconsin Department of Natural Resources: Madison, WI, USA, 2017; 251p.
- Wisconsin Department of Natural Resources (WDNR). Silviculture Handbook: Chapter 46-Swamp Hardwood Cover Type; State of Wisconsin, Department of Natural Resources: Madison, WI, USA, 2013.
- Looney, C.E.; D’Amato, A.W.; Fraver, S.; Palik, B.J.; Reinikainen, M.R. Examining the influences of tree-to-tree competition and climate on size-growth relationships in hydric, multi-aged Fraxinus nigra stands. For. Ecol. Manag. 2016, 375, 238–248. [Google Scholar] [CrossRef]
- Davis, J.C.; Shannon, J.P.; Bolton, N.W.; Kolka, R.K.; Pypker, T.G. Vegetation responses to simulated emerald ash borer infestation in Fraxinus nigra dominated wetlands of Upper Michigan, USA. Can. J. For. Res. 2016, 47, 319–330. [Google Scholar] [CrossRef]
- Bowen, A.K.M.; Stevens, M.H.H. Predicting the effects of emerald ash borer (Agrilus planipennis, Buprestidae) on hardwood swamp forest structure and composition in southern Michigan. J. Torrey Bot. Soc. 2018, 145, 41–54. [Google Scholar] [CrossRef]
- Wright, J.W.; Rauscher, H.M. Black ash. In Silvics of North America, Volume 2, Hardwoods; Burns, R.M., Honkala, B.G., Eds.; Agricultural Handbook 654; United States Department of Agriculture (USDA): Washington, DC, USA, 1990; pp. 344–347. [Google Scholar]
- Palik, B.J.; Batzer, D.P.; Kern, C. Upland forest linkages to seasonal wetlands: Litter flux, processing, and food quality. Ecosystems 2006, 9, 142–151. [Google Scholar] [CrossRef]
- Van Grinsven, M.J.; Shannon, J.P.; Davis, J.C.; Bolton, N.W.; Wagenbrenner, J.W.; Kolka, R.K.; Pypker, T.G. Source water contributions and hydrologic responses to simulated emerald ash borer infestations in depressional black ash wetlands. Ecohydrology 2017, 10, e1862. [Google Scholar] [CrossRef]
- Lenhart, C.; Brooks, K.; Davidson, M.; Slesak, R.; D’Amato, A. Hydrologic source characterization of black ash wetlands: Implications for EAB response. In Proceedings of the American Water Resources Association Summer Specialty Conference Riparian Ecosystems IV: Advancing Science, Economics and Policy, Denver, CO, USA, 27–29 June 2012. [Google Scholar]
- Telander, A.C.; Slesak, R.A.; D’Amato, A.W.; Palik, B.J.; Brooks, K.N.; Lenhart, C.F. Sap flow of black ash in wetland forests of northern Minnesota, USA: Hydrologic implications of tree mortality due to emerald ash borer. Agric. For. Meteorol. 2015, 206, 4–11. [Google Scholar] [CrossRef]
- Shannon, J.; van Grinsven, M.; Davis, J.; Bolton, N.; Noh, N.; Pypker, T.; Kolka, R. Water Level Controls on Sap Flux of Canopy Species in Black Ash Wetlands. Forests 2018, 9, 147. [Google Scholar] [CrossRef]
- Kolka, R.; D’Amato, A.W.; Wagenbrenner, J.; Slesak, R.; Pypker, T.; Youngquist, M.; Grinde, A.; Palik, B. Review of ecosystem level impacts of emerald ash borer on black ash wetlands: What does the future hold? Forests 2018, 9, 179. [Google Scholar] [CrossRef]
- Diamond, J.S.; McLaughlin, D.; Slesak, R.A.; D’Amato, A.W.; Palik, B.J. Forested vs. Herbaceous Wetlands: Can management mitigate ecohydrologic regime shifts from invasive EAB? J. Environ. Manag. 2018, in press. [Google Scholar]
- Croxton, R.J. Detection and Classification of Ash Dieback on Large-Scale Color Aerial Photographs; Research Paper PSW-RP-35; USDA Forest Service: Washington, DC, USA, 1966.
- Livingston, W.H.; Hager, A.; White, A.S.; Hobbins, D. Drought associated with brown ash dieback in Maine. Phytopathology 1995, 85, 1554–1561. [Google Scholar]
- Palik, B.J.; Ostry, M.E.; Venette, R.C.; Abdela, E. Fraxinus nigra (black ash) dieback in Minnesota: Regional variation and potential contributing factors. For. Ecol. Manag. 2011, 261, 128–135. [Google Scholar] [CrossRef]
- Livingston, W.H.; White, A.S. May drought confirmed as likely cause of brown ash dieback in Maine. Phytopathology 1997, 87, S59. [Google Scholar]
- Trial, H., Jr.; Devine, M.E. Forest Health Monitoring Evaluation: Brown Ash (Fraxinus nigra) in Maine. A Survey of Occurrence and Health; Technical Report No. 33; Insect and Disease Management Division: August, ME, USA, 1994. [Google Scholar]
- Peterson, C.E. Natural Regeneration after Logging of Black Ash Stands in Central Minnesota. Master’s Thesis, College of Natural Resources, University of Minnesota, St. Paul, MN, USA, 1989; 58p. [Google Scholar]
- Kashian, D.M.; Witter, J.A. Assessing the potential for ash canopy tree replacement via current regeneration following emerald ash borer-caused mortality on southeastern Michigan landscapes. For. Ecol. Manag. 2011, 261, 480–488. [Google Scholar] [CrossRef]
- Looney, C.E.; D’Amato, A.W.; Palik, B.J.; Slesak, R.A.; Slater, M.A. The response of Fraxinus nigra forest ground-layer vegetation to emulated emerald ash borer mortality and management strategies in Northern Minnesota, USA. For. Ecol. Manag. 2017, 389, 352–363. [Google Scholar] [CrossRef]
- Palik, B.J.; Ostry, M.E.; Venette, R.C.; Abdela, E. Tree regeneration in black ash (Fraxinus nigra) stands exhibiting crown dieback in Minnesota. For. Ecol. Manag. 2012, 269, 26–30. [Google Scholar] [CrossRef]
- Looney, C.E.; D’Amato, A.W.; Palik, B.J.; Slesak, R.A. Overstory treatment and planting season affect survival of replacement tree species in emerald ash borer threatened Fraxinus nigra forests in Minnesota, USA. Can. J. For. Res. 2015, 45, 1728–1738. [Google Scholar] [CrossRef]
- Bolton, N.; Shannon, J.; Davis, J.; Grinsven, M.; Noh, N.; Schooler, S.; Kolka, R.; Pypker, T.; Wagenbrenner, J. Methods to improve survival and growth of planted alternative species seedlings in black ash ecosystems threatened by emerald ash borer. Forests 2018, 9, 146. [Google Scholar] [CrossRef]
- Kabrick, J.M.; Dey, D.C.; van Sambeek, J.W.; Coggeshall, M.V.; Jacobs, D.F. Quantifying flooding effects on hardwood seedling survival and growth for bottomland restoration. New For. 2012, 43, 695–710. [Google Scholar] [CrossRef]
- Knight, K.S.; Slavicek, J.M.; Kappler, R.; Pisarczyk, E.; Wiggin, B.; Menard, K. Using Dutch elm disease-tolerant elm to restore floodplains impacted by emerald ash borer. In Proceedings of the 4th International Workshop on Genetics of Host–Parasite Interactions in Forestry: Disease and Insect Resistance in Forest Trees, Eugene, OR, USA, 31 July–5 August 2011; General Technical Report PSW-GTR-240. USDA Forest Service, Pacific Southwest Research Station: Albany, CA, USA, 2012; pp. 317–323. [Google Scholar]
- WDNR. Silviculture Trials Database: Swamp Hardwood; State of Wisconsin, Department of Natural Resources: Madison, WI, USA, 2015. Available online: https://dnr.wi.gov/topic/forestmanagement/silviculturetrials.html (accessed on 18 April 2018).
- WDNR. Checklist for Evaluating Lowland Ash Stands; State of Wisconsin, Department of Natural Resources: Madison, WI, USA, 2017; 8p.
- University of Minnesota (UMN) Extension. Ash Management Guidelines for Private Forest Landowners; University of Minnesota: St. Paul, MN, USA, 2011; p. 76. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Stock Type | MN | MI | WI | Notes |
|---|---|---|---|---|---|
| Acer rubrum | Bare-root (2+0), containerized (90 cm3) | NR | NR | R | Success observed with bare-root stock in riparian black ash forest |
| Acer saccharinum | Bare-root (4+0) | - | R | - | Success observed with large bare-root stock |
| Abies balsamea | Bare-root (2+0, 3+0) | - | NR | NR | Very low survival regardless of cultural treatment |
| Betula alleghaniensis | Bare-root (2+0), Containerized (90 cm3) | NR | NR | NR | Very low survival with containerized stock. Marginal survival (36%) for bare-root stock planted in clearcut prepared by fecon mower |
| Celtis occidentalis | Bare-root (2+0), Containerized (336 cm3) | R | - | R | Greatest success when planted in group selection harvests in depressional wetlands |
| Fraxinus mandschurica | Bare-root (3+0) | NR | - | - | Limited cold tolerance, despite high survival of sprouts |
| Larix laricina | Bare-root (2+0), containerized (60 cm3) | NR | NR | R | Success observed with bare-root and containerized stock in clearcut prepared with fecon mower |
| Picea glauca | Bare-root (3+0) | - | - | R | Success observed with bare-root stock in clearcut prepared with fecon mower |
| Picea mariana | Containerized (90 cm3) | NR | NR | R | Success observed with bare-root and containerized stock in clearcut prepared with fecon mower |
| Pinus strobus | Bare-root (3+0) | - | - | R | Success observed with bare-root stock in clearcut prepared with fecon mower |
| Platanus occidentalis | Bare-root (1+0) | - | - | R | Success observed with bare-root stock in riparian black ash forest |
| Populus balsamifera | Containerized (164 cm3) | R | - | - | Greatest success when planted in clearcut and group selection harvests in depressional wetlands |
| Populus deltoides | Bare-root (1+0) | NR | - | - | Very low survival regardless of cultural treatment |
| Populus tremuloides | Containerized (90 cm3) | NR | - | NR | Marginal success observed with containerized stock in clearcut prepared with fecon mower |
| Quercus bicolor | Bare-root (1+0) | R | - | R | Greatest success in group selection harvests in depressional wetlands; successful under intact canopy of riparian black ash forests |
| Quercus macrocarpa | Bare-root (3+0) | - | NR | - | Greatest success when planted on hummocks |
| Thuja occidentalis | Bare-root (2+0), Containerized (60 cm3) | R | R | R | Greatest success in intact canopies and group selection harvests; high incidence of browse on seedlings |
| Tilia americana | Bare-root (3+0) | - | R | - | Greatest success when planted on hummocks |
| Tsuga canadensis | Bare-root (3+0) | - | - | NR | |
| Ulmus americana | Containerized (1890 cm3) | R | R | - | Greatest success when planted in group selection harvests and on hummocks using DED1 tolerant cultivars |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Amato, A.W.; Palik, B.J.; Slesak, R.A.; Edge, G.; Matula, C.; Bronson, D.R. Evaluating Adaptive Management Options for Black Ash Forests in the Face of Emerald Ash Borer Invasion. Forests 2018, 9, 348. https://doi.org/10.3390/f9060348
D’Amato AW, Palik BJ, Slesak RA, Edge G, Matula C, Bronson DR. Evaluating Adaptive Management Options for Black Ash Forests in the Face of Emerald Ash Borer Invasion. Forests. 2018; 9(6):348. https://doi.org/10.3390/f9060348
Chicago/Turabian StyleD’Amato, Anthony W., Brian J. Palik, Robert A. Slesak, Greg Edge, Colleen Matula, and Dustin R. Bronson. 2018. "Evaluating Adaptive Management Options for Black Ash Forests in the Face of Emerald Ash Borer Invasion" Forests 9, no. 6: 348. https://doi.org/10.3390/f9060348
APA StyleD’Amato, A. W., Palik, B. J., Slesak, R. A., Edge, G., Matula, C., & Bronson, D. R. (2018). Evaluating Adaptive Management Options for Black Ash Forests in the Face of Emerald Ash Borer Invasion. Forests, 9(6), 348. https://doi.org/10.3390/f9060348