Fog Water Is Important in Maintaining the Water Budgets of Vascular Epiphytes in an Asian Tropical Karst Forests during the Dry Season

 ,
,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
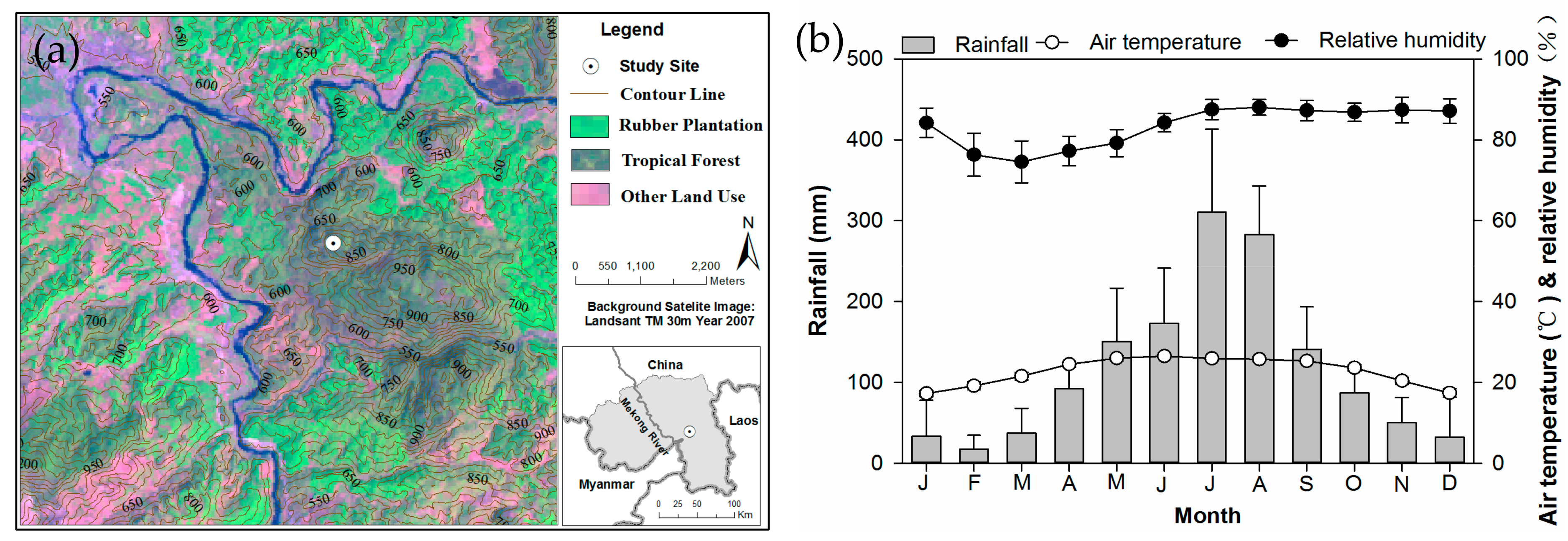
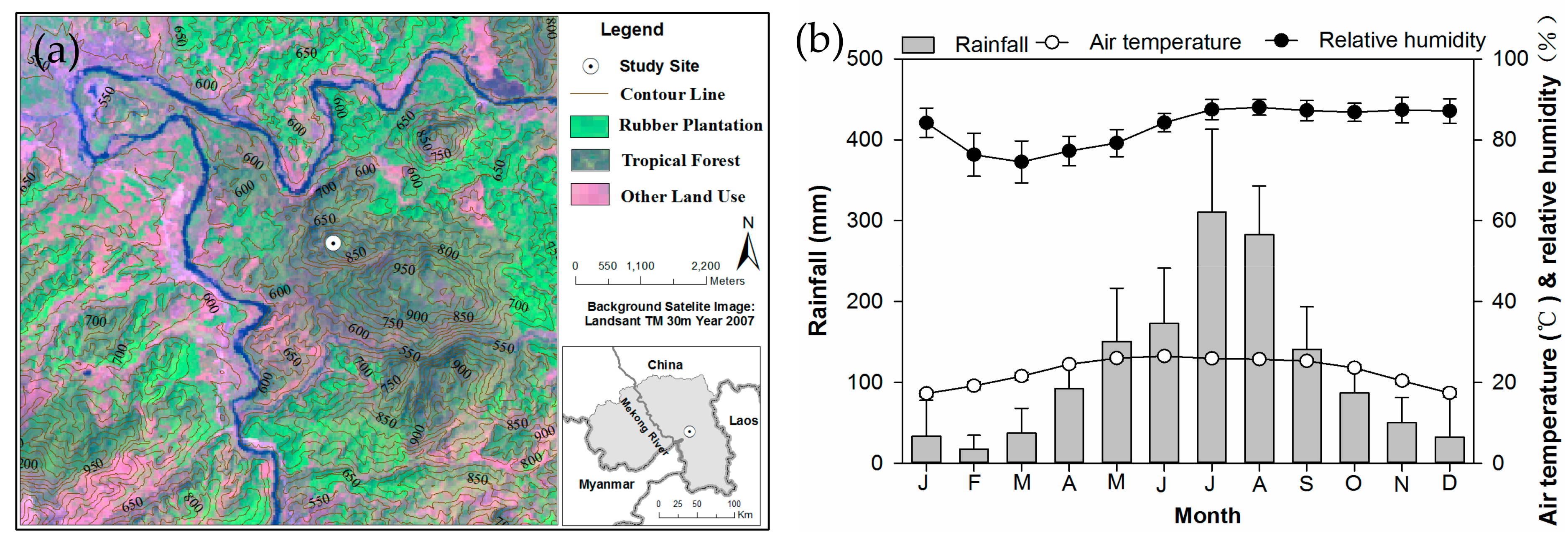
2.1. Study Site and Species
2.2. Sampling
2.3. Stable Isotope Analysis
2.4. Foliar Water Uptake
2.5. Statistical Analyses
3. Results
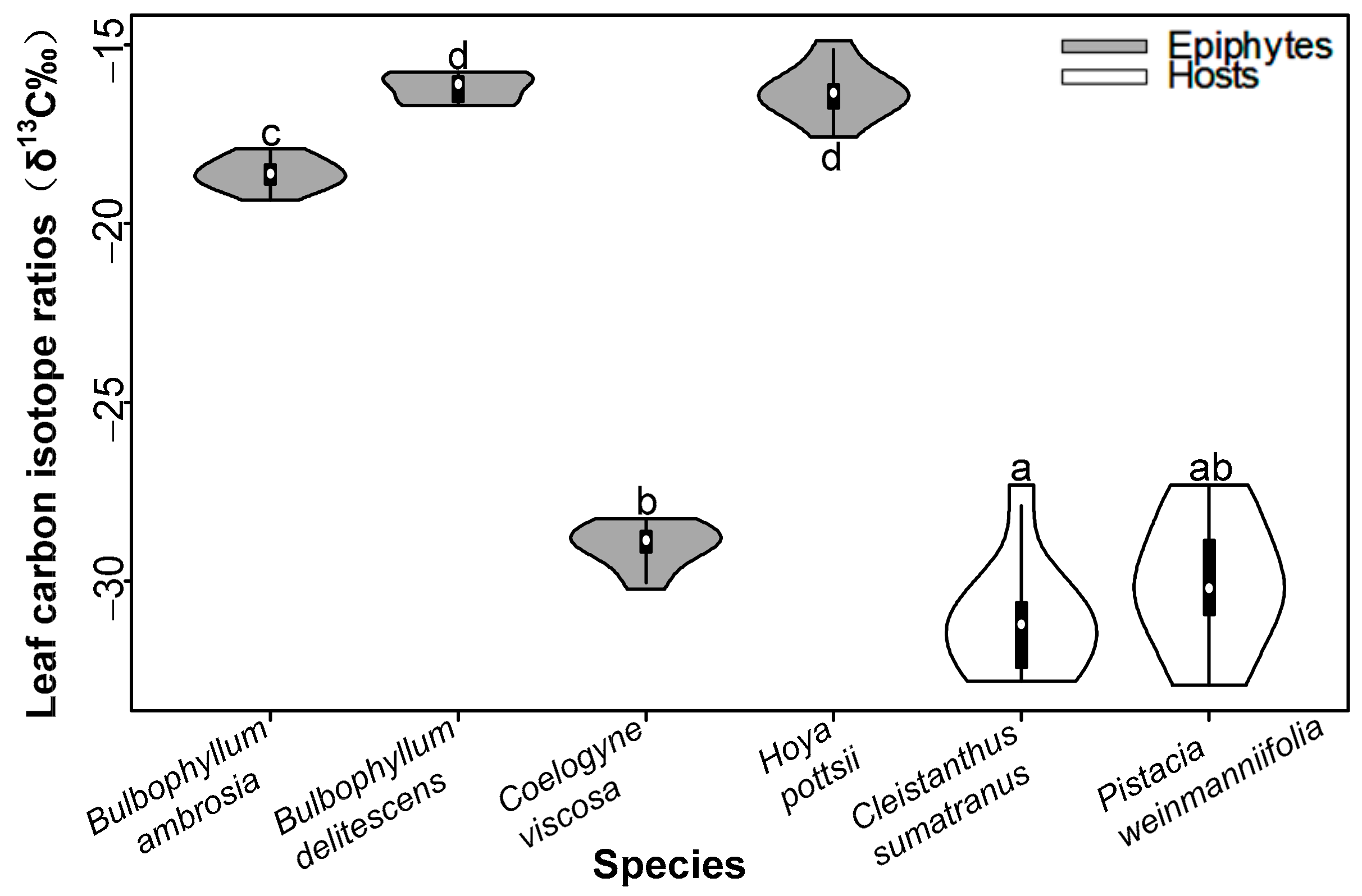
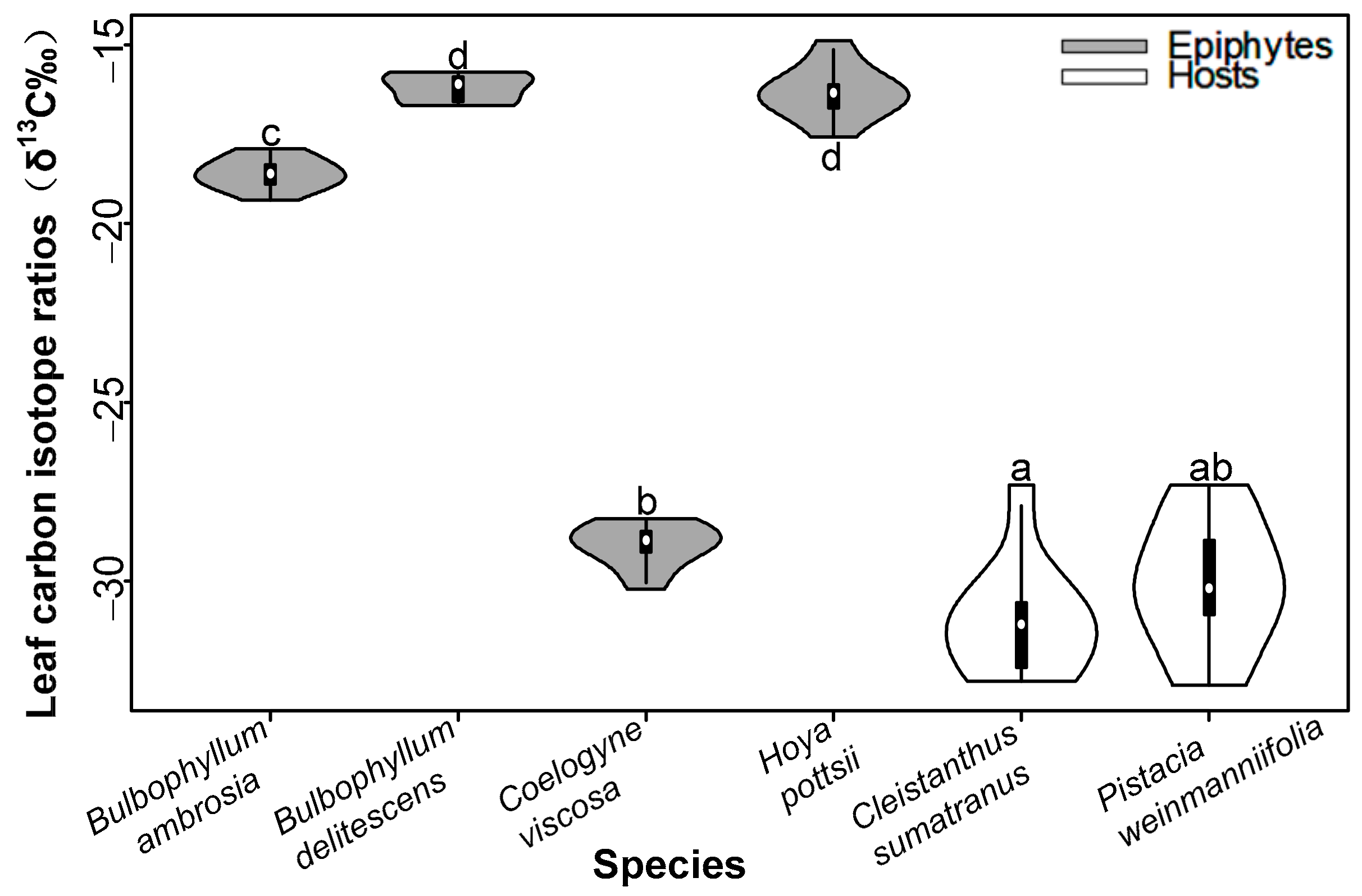
3.1. Water Use Efficiency
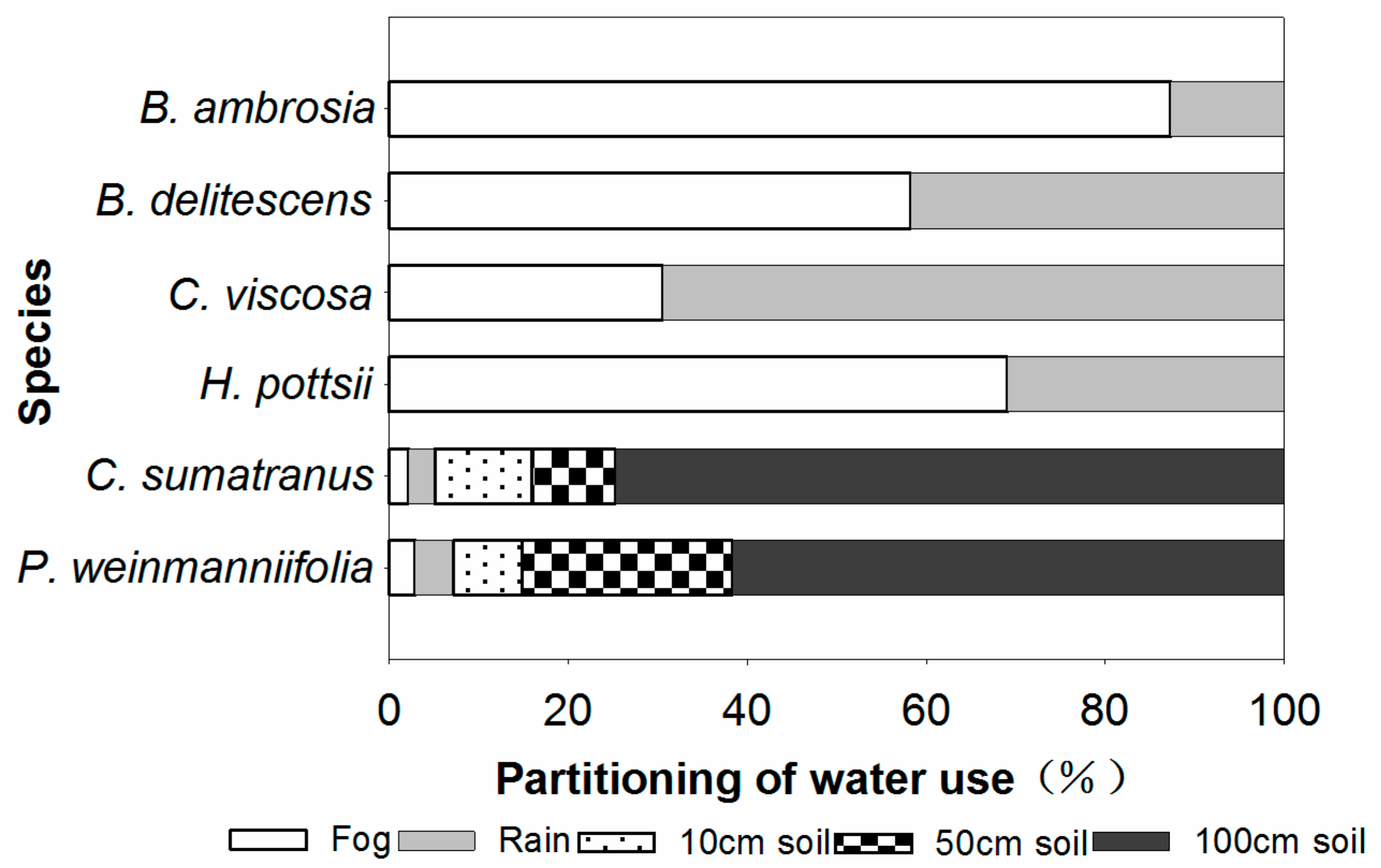
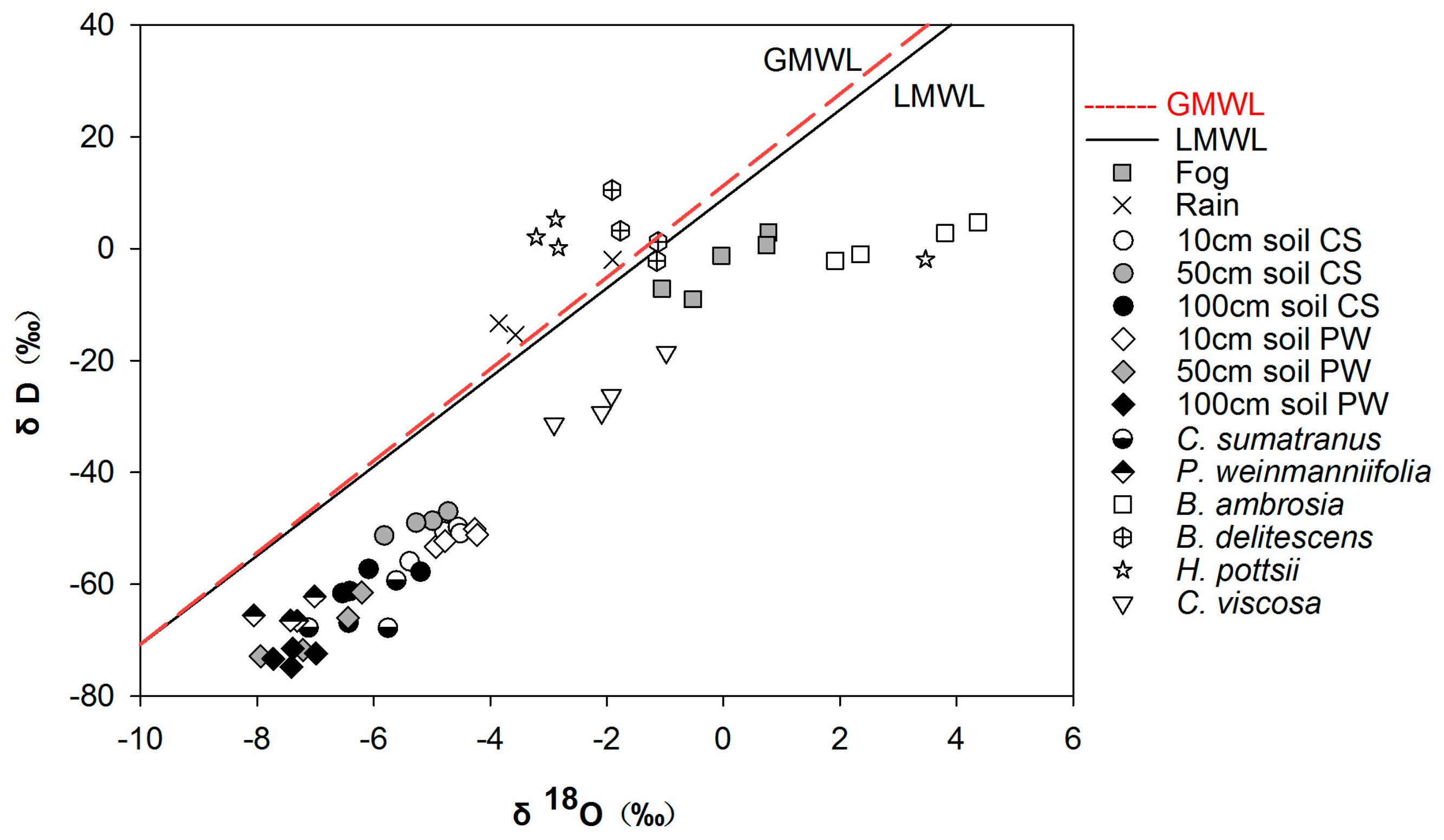
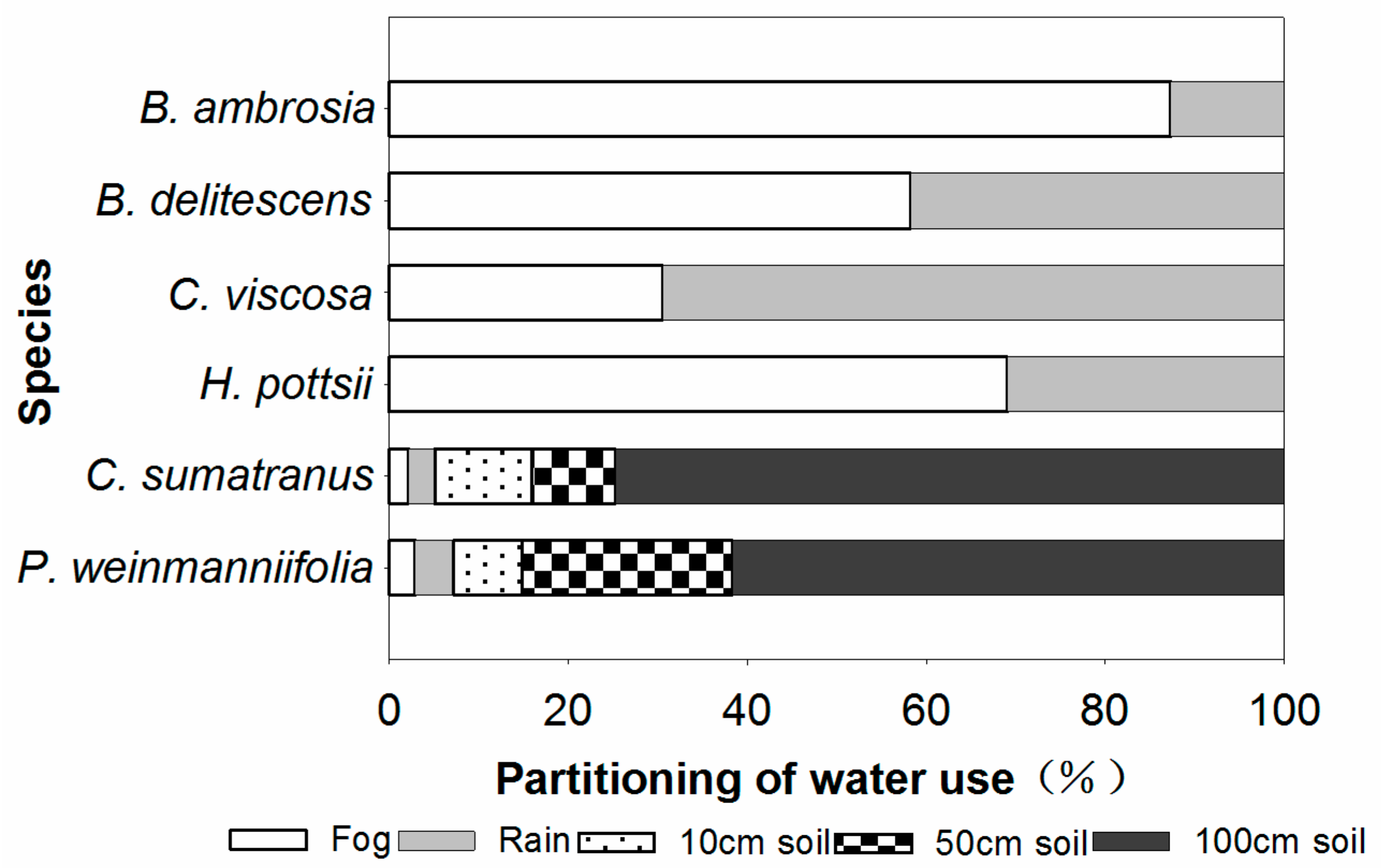
3.2. Partitioning of Water Use
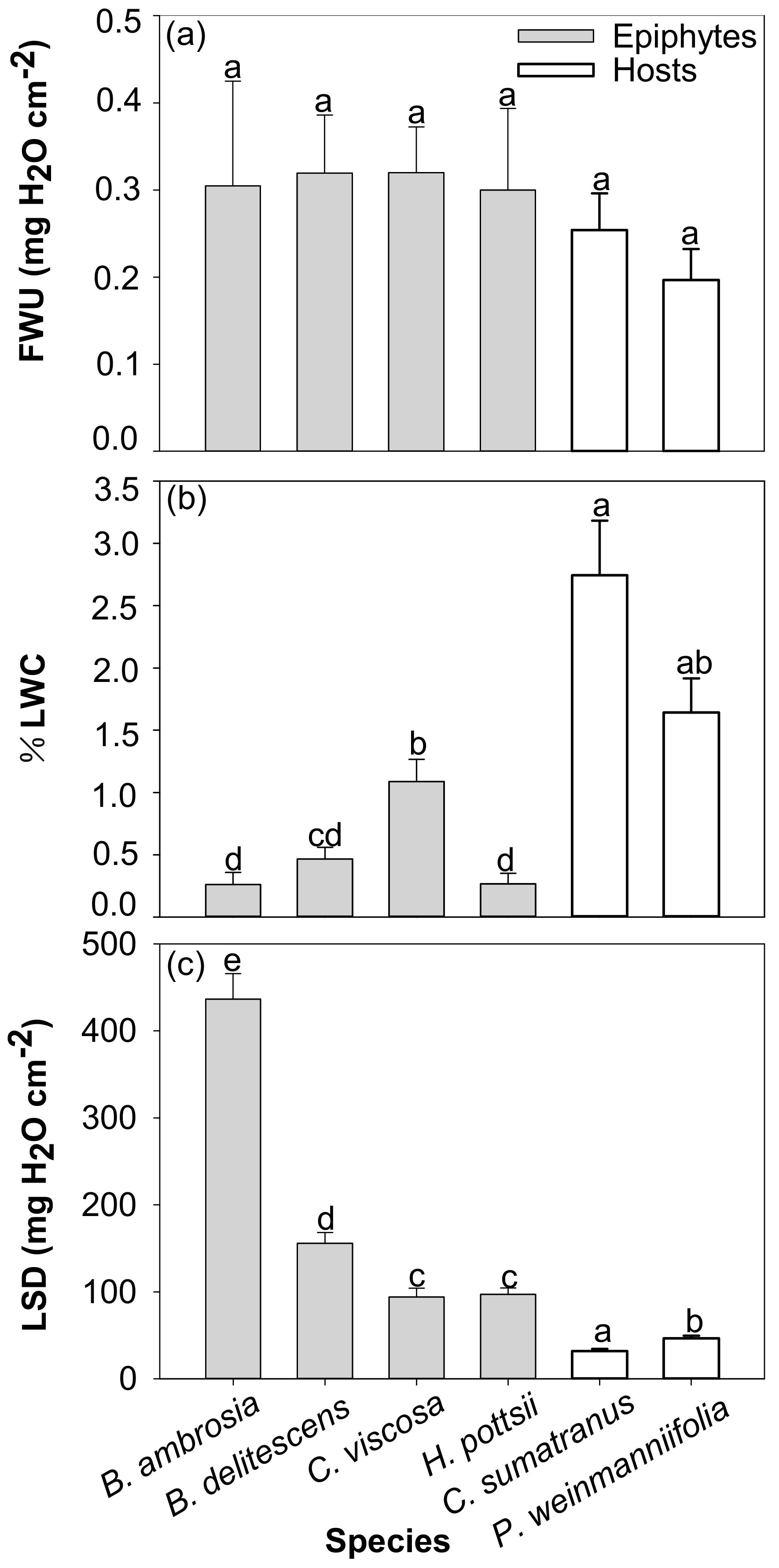
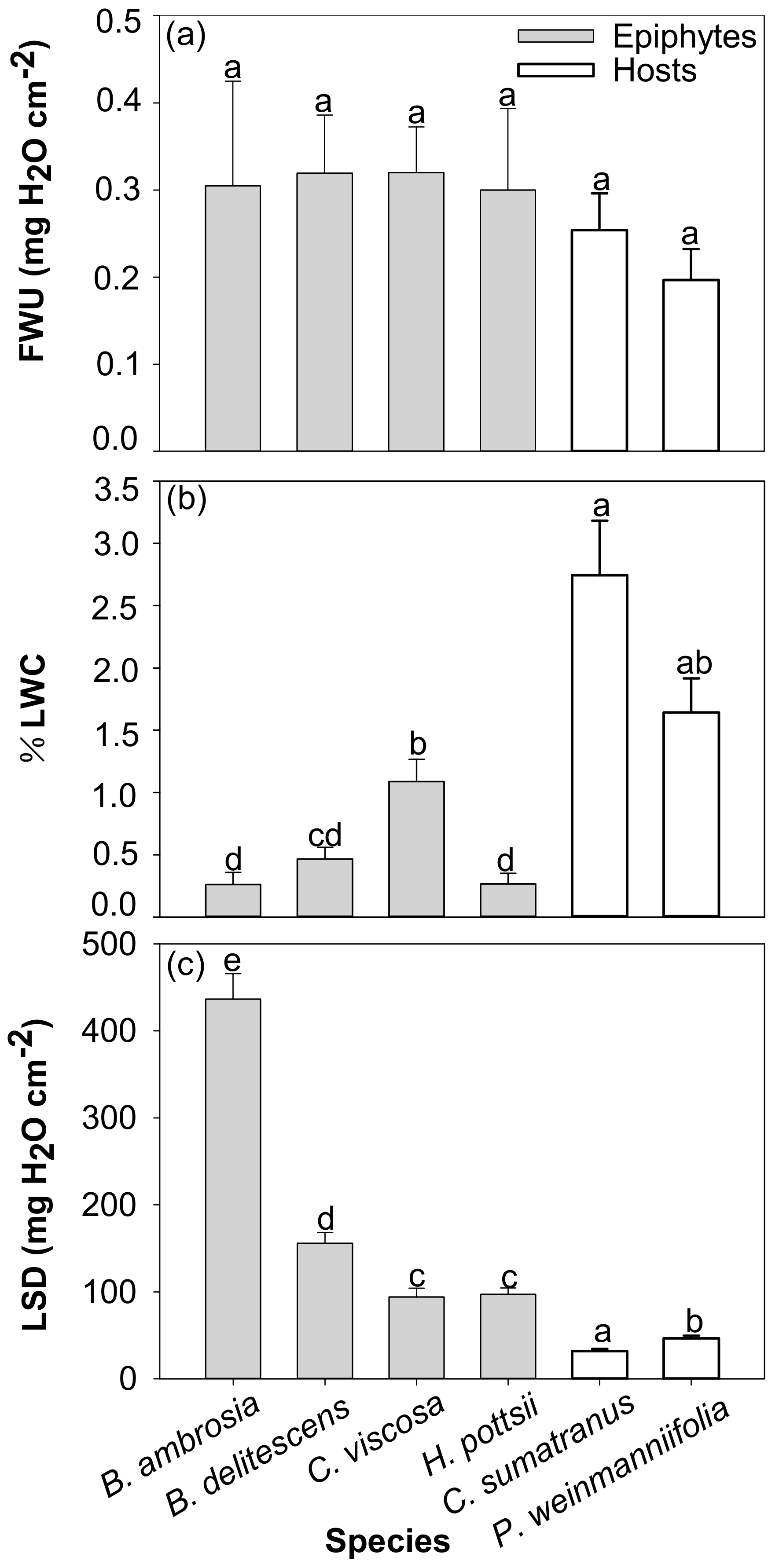
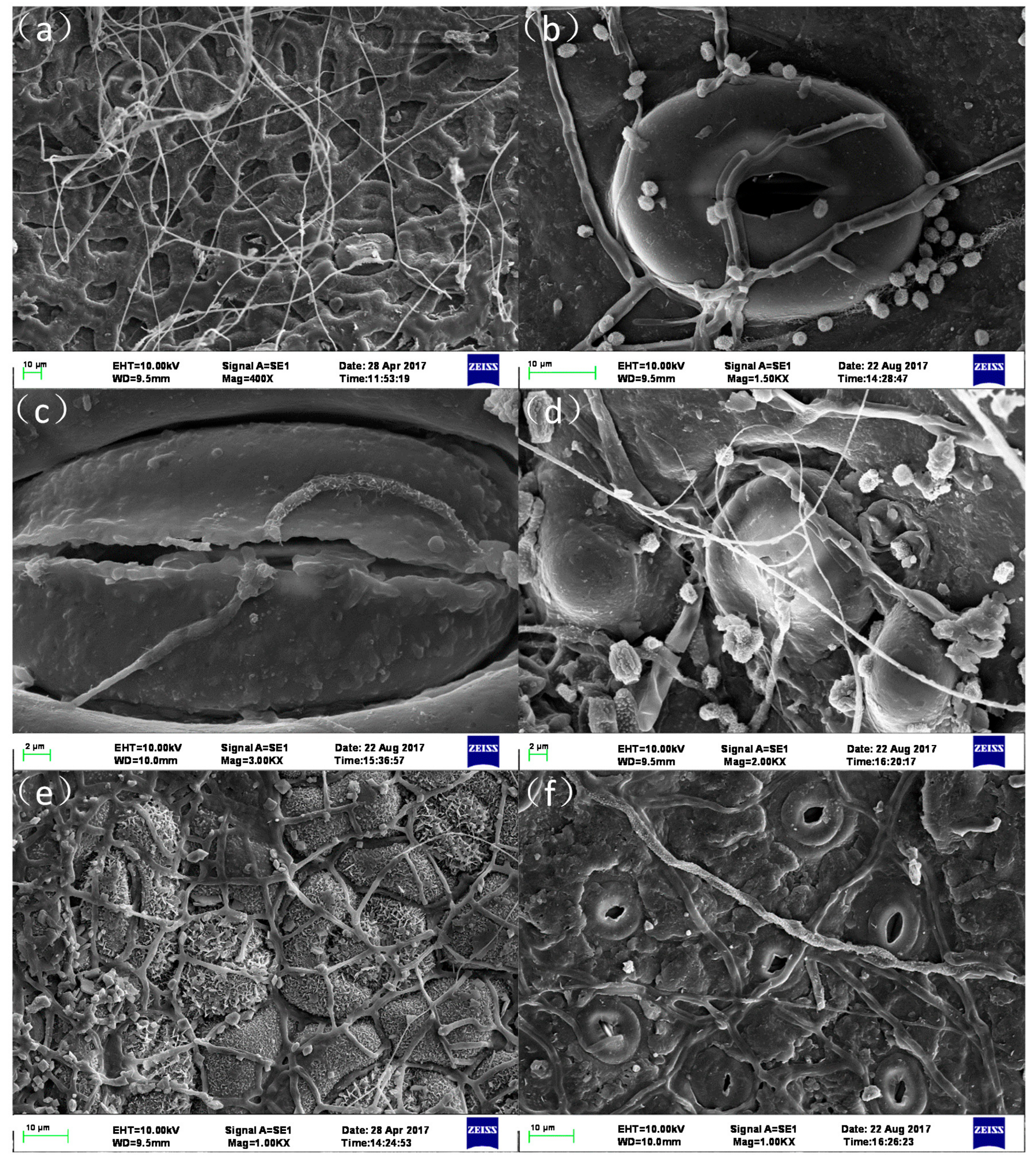
3.3. Foliar Water Uptake
4. Discussion
4.1. Water Use Efficiency
4.2. Partitioning of Water Use
4.3. Foliar Water Uptake
4.4. Ecological Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gotsch, S.G.; Nadkarni, N.; Darby, A.; Glunk, A.; Dix, M.; Davidson, K.; Dawson, T.E. Life in the treetops: Ecophysiological strategies of canopy epiphytes in a tropical montane cloud forest. Ecol. Monogr. 2015, 85, 393–412. [Google Scholar] [CrossRef]
- Bruijnzeel, L.; Mulligan, M.; Scatena, F.N. Hydrometeorology of tropical montane cloud forests: Emerging patterns. Hydrol. Process. 2011, 25, 465–498. [Google Scholar] [CrossRef]
- Liu, W.; Liu, W.; Li, P.; Duan, W.; Li, H. Dry season water uptake by two dominant canopy tree species in a tropical seasonal rainforest of Xishuangbanna, SW China. Agric. For. Meteorol. 2010, 150, 380–388. [Google Scholar] [CrossRef]
- Darby, A.; Draguljić, D.; Glunk, A.; Gotsch, S.G. Habitat moisture is an important driver of patterns of sap flow and water balance in tropical montane cloud forest epiphytes. Oecologia 2016, 182, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Gotsch, S.G.; Dawson, T.E.; Draguljic, D. Variation in the resilience of cloud forest vascular epiphytes to severe drought. New Phytol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Gehrig-Downie, C.; Obregon, A.; Bendix, J.; Gradstein, S.R. Epiphyte biomass and canopy microclimate in the tropical lowland cloud forest of French Guiana. Biotropica 2011, 43, 591–596. [Google Scholar] [CrossRef]
- Cavelier, J.; Goldstein, G. Mist and fog interception in elfin cloud forests in Colombia and Venezuela. J. Trop. Ecol. 1989, 5, 309–322. [Google Scholar] [CrossRef]
- Grubb, P.J.; Whitmore, T.C. A comparison of montane and lowland rain forest in Ecuador: II. the climate and its effects on the distribution and physiognomy of the forests. J. Ecol. 1966, 54, 303–333. [Google Scholar] [CrossRef]
- Gotsch, S.G.; Nadkarni, N.; Amici, A. The functional roles of epiphytes and arboreal soils in tropical montane cloud forests. J. Trop. Ecol. 2016, 32, 455–468. [Google Scholar] [CrossRef]
- Hietz, P. Ecology and ecophysiology of epiphytes in tropical montane cloud forests. In Tropical Montane Cloud Forests Science for Conservation and Management; Bruijnzeel, L.A., Scatena, F.N., Hamilton, L.S., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 67–76. [Google Scholar]
- Watkins, J.E., Jr.; Rundel, P.W.; Cardelús, C.L. The influence of life form on carbon and nitrogen relationships in tropical rainforest ferns. Oecologia 2007, 153, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.N.; Liu, W.J.; Chen, C.F. Can intercropping with the world's three major beverage plants help improve the water use of rubber trees? J. Appl. Ecol. 2016, 53, 1787–1799. [Google Scholar] [CrossRef]
- Winter, K.; Aranda, J.; Holtum, J.A.M. Carbon isotope composition and water-use efficiency in plants with crassulacean acid metabolism. Funct. Plant Biol. 2005, 32, 381–388. [Google Scholar] [CrossRef]
- Crayn, D.M.; Winter, K.; Schulte, K.; Smith, J.A.C. Photosynthetic pathways in Bromeliaceae: Phylogenetic and ecological significance of CAM and C-3 based on carbon isotope ratios for 1893 species. Bot. J. Linn. Soc. 2015, 178, 169–221. [Google Scholar] [CrossRef]
- Silvera, K.; Santiago, L.S.; Cushman, J.C.; Winter, K. The incidence of crassulacean acid metabolism in Orchidaceae derived from carbon isotope ratios: A checklist of the flora of Panama and Costa Rica. Bot. J. Linn. Soc. 2010, 163, 194–222. [Google Scholar] [CrossRef]
- Givnish, T.J.; Spalink, D.; Ames, M.; Lyon, S.P.; Hunter, S.J.; Zuluaga, A.; Iles, W.J.; Clements, M.A.; Arroyo, M.T.; Leebens-Mack, J.; et al. Orchid phylogenomics and multiple drivers of their extraordinary diversification. Proceedings. Biol. Sci. 2015, 282, 2108–2111. [Google Scholar] [CrossRef] [PubMed]
- Silvera, K.; Santiago, L.S.; Cushman, J.C.; Winter, K. Crassulacean acid metabolism and epiphytism linked to adaptive radiations in the Orchidaceae. Plant Physiol. 2009, 149, 1838–1847. [Google Scholar] [CrossRef] [PubMed]
- Cardelús, C.L.; Mack, M.C. The nutrient status of epiphytes and their host trees along an elevational gradient in Costa Rica. Plant Ecol. 2010, 207, 25–37. [Google Scholar] [CrossRef]
- Burgess, S.S.O.; Dawson, T.E. The contribution of fog to the water relations of Sequoia sempervirens (D. Don): Foliar uptake and prevention of dehydration. Plant Cell Environ. 2004, 27, 1023–1034. [Google Scholar] [CrossRef]
- Dawson, T.E. Fog in the California redwood forest: Ecosystem inputs and use by plants. Oecologia 1998, 117, 476–485. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.J.; Li, P.J.; Duan, W.P.; Liu, W.Y. Dry-season water utilization by trees growing on thin karst soils in a seasonal tropical rainforest of Xishuangbanna, Southwest China. Ecohydrology 2014, 7, 927–935. [Google Scholar] [CrossRef]
- Fu, P.L.; Liu, W.J.; Fan, Z.X.; Cao, K.F. Is fog an important water source for woody plants in an Asian tropical karst forest during dry season? Ecohydrology 2016, 9, 964–972. [Google Scholar] [CrossRef]
- Feild, T.S.; Dawson, T.E. Water sources used by Didymopanax pittieri at different life stages in a tropical cloud forest. Ecology 1998, 79, 1448–1452. [Google Scholar] [CrossRef]
- Goldsmith, G.R.; Lehmann, M.M.; Cernusak, L.A.; Arend, M.; Siegwolf, R.T.W. Inferring foliar water uptake using stable isotopes of water. Oecologia 2017, 184, 763–766. [Google Scholar] [CrossRef] [PubMed]
- Limm, E.B.; Simonin, K.A.; Bothman, A.G.; Dawson, T.E. Foliar water uptake: A common water acquisition strategy for plants of the redwood forest. Oecologia 2009, 161, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Ponette-Gonzalez, A.G.; Weathers, K.C.; Curran, L.M. Water inputs across a tropical montane landscape in Veracruz, Mexico: Synergistic effects of land cover, rain and fog seasonality, and interannual precipitation variability. Glob. Chang. Biol. 2010, 16, 946–963. [Google Scholar] [CrossRef]
- Van Stan II, J. T.; Pypker, T.G. A review and evaluation of forest canopy epiphyte roles in the partitioning and chemical alteration of precipitation. Sci. Total Environ. 2015, 536, 813–824. [Google Scholar] [CrossRef] [PubMed]
- Helliker, B.R. Reconstructing the δ18O of atmospheric water vapour via the CAM epiphyte Tillandsia usneoides: Seasonal controls on δ18O in the field and large-scale reconstruction of δ18Oa. Plant Cell Environ. 2014, 37, 541–556. [Google Scholar] [CrossRef] [PubMed]
- Eller, C.B.; Lima, A.L.; Oliveira, R.S. Foliar uptake of fog water and transport belowground alleviates drought effects in the cloud forest tree species, Drimys brasiliensis (Winteraceae). New Phytol. 2013, 199, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Pitt, W.G.; Zhang, Y.; Wu, N.; Tao, Y.; Truscott, T.T. The upside-down water collection system of Syntrichia caninervis. Nat. Plants 2016, 2, 16076. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.J.; Wang, P.Y.; Li, J.T.; Li, P.J.; Liu, W.Y. The importance of radiation fog in the tropical seasonal rain forest of Xishuangbanna, south-west China. Hydrol. Res. 2008, 39, 79–87. [Google Scholar] [CrossRef]
- Wu, Y.; Song, L.; Liu, Q.; Zhao, M.; Lu, H.; Tan, Y.; Liu, W. Diversity and floristic characteristics of vascular epiphytes in the tropical forest of Xishuangbanna. Biodivers. Sci. 2016, 24, 271–279. [Google Scholar] [CrossRef]
- Cheng, J.; Xie, M. The analysis of regional climate change features over Yunnan in recent 50 years. Progress Geogr. 2008, 27, 19–26, (In Chinese with English Abstract). [Google Scholar]
- Zhang, Y.J.; Holbrook, N.M.; Cao, K.F. Seasonal dynamics in photosynthesis of woody plants at the northern limit of Asian tropics: Potential role of fog in maintaining tropical rainforests and agriculture in Southwest China. Tree Physiol. 2014, 34, 1069–1078. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Stott, L.; Buckley, B.; Yoshimura, K. 20th century seasonal moisture balance in Southeast Asian montane forests from tree cellulose δ18O. Clim. Chang. 2012, 115, 505–517. [Google Scholar] [CrossRef]
- Liu, J.-J.; Slik, J.W.F. Forest fragment spatial distribution matters for tropical tree conservation. Biol. Conserv. 2014, 171, 99–106. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, Y.; Liu, Y.; Li, H.; Duan, W. Fog throughfall at a seasonal rain forest in Xishuangbanna, Southwest China. Acta Phytoecol. Sin. 2003, 27, 749–756. [Google Scholar]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Clements, R.; Sodhi, N.S.; Schilthuizen, M.; Ng, P.K.L. Limestone karsts of Southeast Asia: Imperiled arks of biodiversity. Bioscience 2006, 56, 733–742. [Google Scholar] [CrossRef]
- Zhu, H.; Wang, H.; Baogui, L.; Sirirugsa, P. Biogeography and floristic affinities of the limestone flora in Southern Yunnan, China. Ann. Mo. Bot. Gard. 2003, 90, 444–465. [Google Scholar] [CrossRef]
- Zhu, H. Forest vegetation of Xishuangbanna, south China. For. Stud. China 2006, 8, 1–58. [Google Scholar]
- Zhu, H.; Wang, H.; Baogui, L. The structure, species composition and diversity of the limestone vegetation in Xishuangbanna, SW China. Gard. Bull. Singap. 1998, 50, 5–30. [Google Scholar]
- Tang, J.W.; Lu, X.T.; Yin, J.X.; Qi, J.F. Diversity, composition and physical structure of tropical forest over limestone in xishuangbanna, south-west China. J. Trop. For. Sci. 2011, 23, 425–433. [Google Scholar]
- Ehleringer, J.R.; Roden, J.; Dawson, T.E. Assessing Ecosystem-Level Water Relations through Stable Isotope Ratio Analyses. In Methods in Ecosystem Science; Springer: New York, NY, USA, 2000; pp. 181–198. [Google Scholar]
- Mohammadian, M.A.; Hill, R.S.; Watling, J.R. Stomatal plugs and their impact on fungal invasion in Agathis robusta. Aust. J. Bot. 2009, 57, 389–395. [Google Scholar] [CrossRef]
- Parnell, A.C.; Phillips, D.L.; Bearhop, S.; Semmens, B.X.; Ward, E.J.; Moore, J.W.; Jackson, A.L.; Grey, J.; Kelly, D.J.; Inger, R. Bayesian stable isotope mixing models. Environmetrics 2013, 24, 387–399. [Google Scholar] [CrossRef]
- R Core Team. The R Foundation for Statistical Computing Platform. R Version 3.4.2. Available online: http://www.r-project.org (accessed on 28 September 2017).
- Liu, W.J.; Liu, W.Y.; Li, P.J.; Gao, L.; Shen, Y.X.; Wang, P.Y.; Zhang, Y.P.; Li, H.M. Using stable isotopes to determine sources of fog drip in a tropical seasonal rain forest of Xishuangbanna, SW China. Agric. For. Meteorol. 2007, 143, 80–91. [Google Scholar] [CrossRef]
- Martin, C.E.; Mas, E.J.; Lu, C.; Ong, B.L. The photosynthetic pathway of the roots of twelve epiphytic orchids with CAM leaves. Photosynthetica 2010, 48, 42–50. [Google Scholar] [CrossRef]
- Yang, X.; Cushman, J.C.; Borland, A.M.; Edwards, E.J.; Wullschleger, S.D.; Tuskan, G.A.; Owen, N.A.; Griffiths, H.; Smith, J.A.C.; De Paoli, H.C.; et al. A roadmap for research on crassulacean acid metabolism (CAM) to enhance sustainable food and bioenergy production in a hotter, drier world. New Phytol. 2015, 207, 491–504. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Norhafis, H.; Qin, L. Responses of green leaves and green pseudobulbs of CAM Orchid Cattleya laeliocattleya Aloha Case to drought stress. J. Bot. 2013, 2013, 710539. [Google Scholar]
- Martorell, C.; Ezcurra, E. The narrow-leaf syndrome: A functional and evolutionary approach to the form of fog-harvesting rosette plants. Oecologia 2007, 151, 561–573. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Wang, P.; Li, J.; Liu, W.; Li, H. Plasticity of source-water acquisition in epiphytic, transitional and terrestrial growth phases of Ficus tinctoria. Ecohydrology 2014, 7, 1524–1533. [Google Scholar] [CrossRef]
- Tobón, C.; Köhler, L.; Frumau, K.; Bruijnzeel, L.; Burkard, R.; Schmid, S. Water dynamics of epiphytic vegetation in a lower montane cloud forest: Fog interception, storage, and evaporation. In Tropical Montane Cloud Forests: Science for Conservation and Management; Cambridge University Press: Cambridge UK, 2011; pp. 261–267. [Google Scholar]
- Reyes-Garcia, C.; Mejia-Chang, M.; Griffiths, H. High but not dry: Diverse epiphytic bromeliad adaptations to exposure within a seasonally dry tropical forest community. New Phytol. 2012, 193, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Orlowski, N.; Breuer, L.; McDonnell, J.J. Critical issues with cryogenic extraction of soil water for stable isotope analysis. Ecohydrology 2016, 9, 1–5. [Google Scholar] [CrossRef]
- Orlowski, N.; Pratt, L.; McDonnell, J.J. Intercomparison of soil pore water extraction methods for stable isotope analysis. Hydrol. Process. 2016, 30, 3434–3449. [Google Scholar] [CrossRef]
- Goldsmith, G.R.; Matzke, N.J.; Dawson, T.E. The incidence and implications of clouds for cloud forest plant water relations. Ecol. Lett. 2013, 16, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.B.; Dai, Y.; Hao, G.Y.; Li, J.W.; Fu, X.W.; Zhang, J.L. Differentiation of water-related traits in terrestrial and epiphytic Cymbidium species. Front. Plant Sci. 2015, 6, 260. [Google Scholar] [CrossRef] [PubMed]
- Hietz, P.; Wanek, W. Size-dependent variation of carbon and nitrogen isotope abundances in epiphytic bromeliads. Plant Biol. 2003, 5, 137–142. [Google Scholar] [CrossRef]
- Hu, J.; Riveros-Iregui, D.A. Life in the clouds: Are tropical montane cloud forests responding to changes in climate? Oecologia 2016, 180, 1061–1073. [Google Scholar] [CrossRef] [PubMed]
- Pariyar, S.; Chang, S.C.; Zinsmeister, D.; Zhou, H.; Grantz, D.A.; Hunsche, M.; Burkhardt, J. Xeromorphic traits help to maintain photosynthesis in the perhumid climate of a Taiwanese cloud forest. Oecologia 2017, 184, 609–621. [Google Scholar] [CrossRef] [PubMed]
- Lawton, R.O.; Nair, U.S.; Pielke, R., Sr.; Welch, R.M. Climatic impact of tropical lowland deforestation on nearby montane cloud forests. Science 2001, 294, 584–587. [Google Scholar] [PubMed]
- Still, C.J.; Foster, P.N.; Schneider, S.H. Simulating the effects of climate change on tropical montane cloud forests. Nature 1999, 398, 608–610. [Google Scholar] [CrossRef]
- Wright, J.S.; Fu, R.; Worden, J.R.; Chakraborty, S.; Clinton, N.E.; Risi, C.; Sun, Y.; Yin, L. Rainforest-initiated wet season onset over the southern Amazon. Proc. Natl. Acad. Sci. USA 2017, 114, 8481–8486. [Google Scholar] [CrossRef] [PubMed]
- Hsu, R.C.C.; Oostermeijer, J.G.B.; Wolf, J.H.D. Adaptation of a widespread epiphytic fern to simulated climate change conditions. Plant Ecol. 2014, 215, 889–897. [Google Scholar] [CrossRef]
- Nadkarni, N.M.; Solano, R. Potential effects of climate change on canopy communities in a tropical cloud forest: An experimental approach. Oecologia 2002, 131, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Liu, W.Y.; Nadkarni, N.M. Response of non-vascular epiphytes to simulated climate change in a montane moist evergreen broad-leaved forest in southwest China. Biol. Conserv. 2012, 152, 127–135. [Google Scholar] [CrossRef]
- Xu, J.C.; Grumbine, R.E.; Beckschafer, P. Landscape transformation through the use of ecological and socioeconomic indicators in Xishuangbanna, Southwest China, Mekong Region. Ecol. Indic. 2014, 36, 749–756. [Google Scholar] [CrossRef]
- Li, H.; Aide, T.M.; Ma, Y.; Liu, W.; Cao, M. Demand for rubber is causing the loss of high diversity rain forest in SW China. Biodivers. Conserv. 2007, 16, 1731–1745. [Google Scholar] [CrossRef]
- Tan, Z.H.; Zhang, Y.P.; Song, Q.H.; Liu, W.J.; Deng, X.B.; Tang, J.W.; Deng, Y.; Zhou, W.J.; Yang, L.Y.; Yu, G.R.; et al. Rubber plantations act as water pumps in tropical China. Geophys. Res. Lett. 2011, 38. [Google Scholar] [CrossRef]
- Fan, Z.X.; Bräuning, A.; Thomas, A.; Li, J.B.; Cao, K.F. Spatial and temporal temperature trends on the Yunnan Plateau (Southwest China) during 1961–2004. Int. J. Climatol. 2011, 31, 2078–2090. [Google Scholar] [CrossRef]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Song, L.; Liu, W.; Liu, W.; Li, S.; Fu, P.; Shen, Y.; Wu, J.; Wang, P.; Chen, Q.; et al. Fog Water Is Important in Maintaining the Water Budgets of Vascular Epiphytes in an Asian Tropical Karst Forests during the Dry Season. Forests 2018, 9, 260. https://doi.org/10.3390/f9050260
Wu Y, Song L, Liu W, Liu W, Li S, Fu P, Shen Y, Wu J, Wang P, Chen Q, et al. Fog Water Is Important in Maintaining the Water Budgets of Vascular Epiphytes in an Asian Tropical Karst Forests during the Dry Season. Forests. 2018; 9(5):260. https://doi.org/10.3390/f9050260
Chicago/Turabian StyleWu, Yi, Liang Song, Wenyao Liu, Wenjie Liu, Su Li, Peili Fu, Youxin Shen, Junen Wu, Pingyuan Wang, Quan Chen, and et al. 2018. "Fog Water Is Important in Maintaining the Water Budgets of Vascular Epiphytes in an Asian Tropical Karst Forests during the Dry Season" Forests 9, no. 5: 260. https://doi.org/10.3390/f9050260
APA StyleWu, Y., Song, L., Liu, W., Liu, W., Li, S., Fu, P., Shen, Y., Wu, J., Wang, P., Chen, Q., & Lu, H. (2018). Fog Water Is Important in Maintaining the Water Budgets of Vascular Epiphytes in an Asian Tropical Karst Forests during the Dry Season. Forests, 9(5), 260. https://doi.org/10.3390/f9050260