4.1. Implications and Conservation Strategies
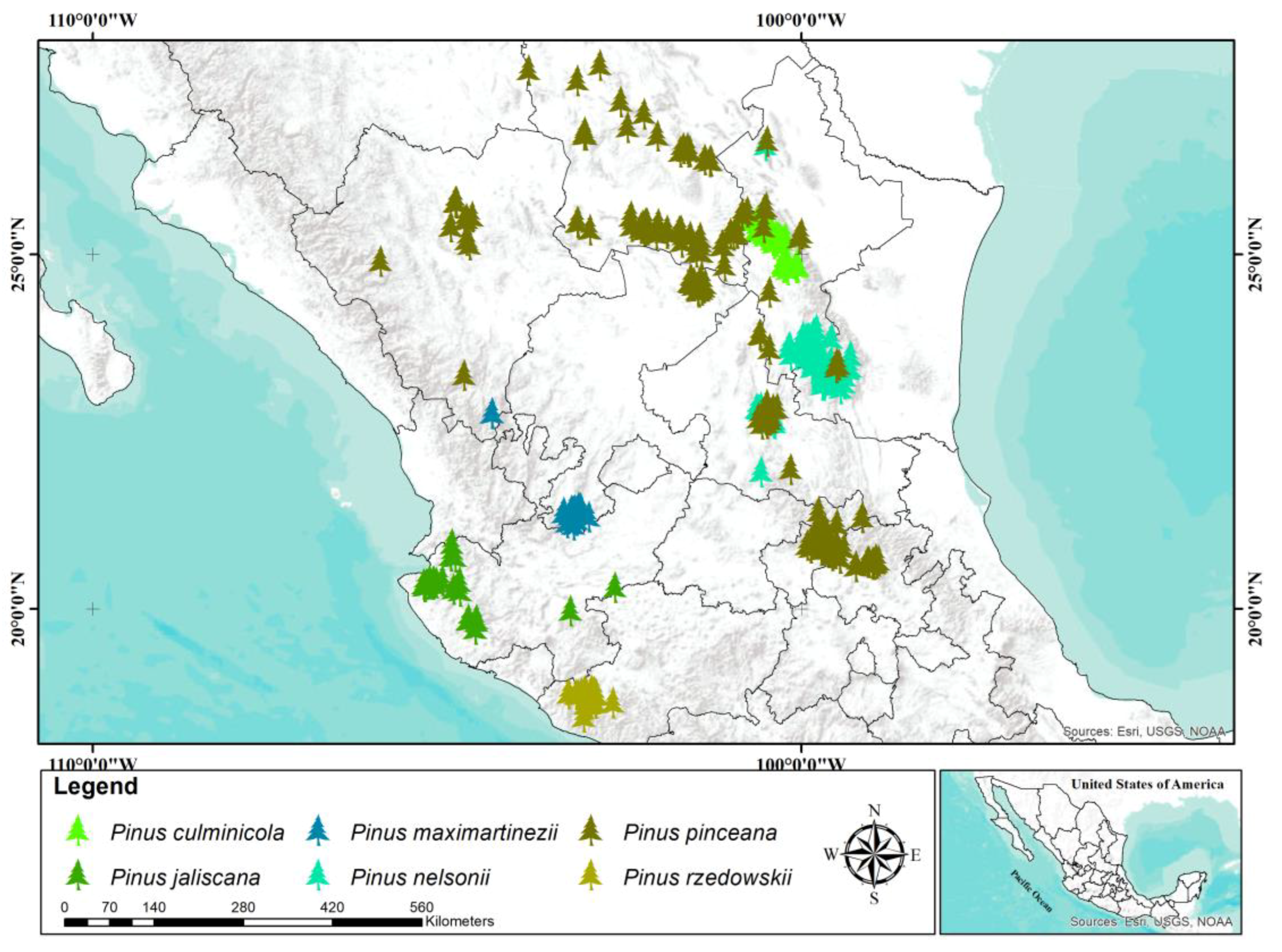
The modeled distribution of
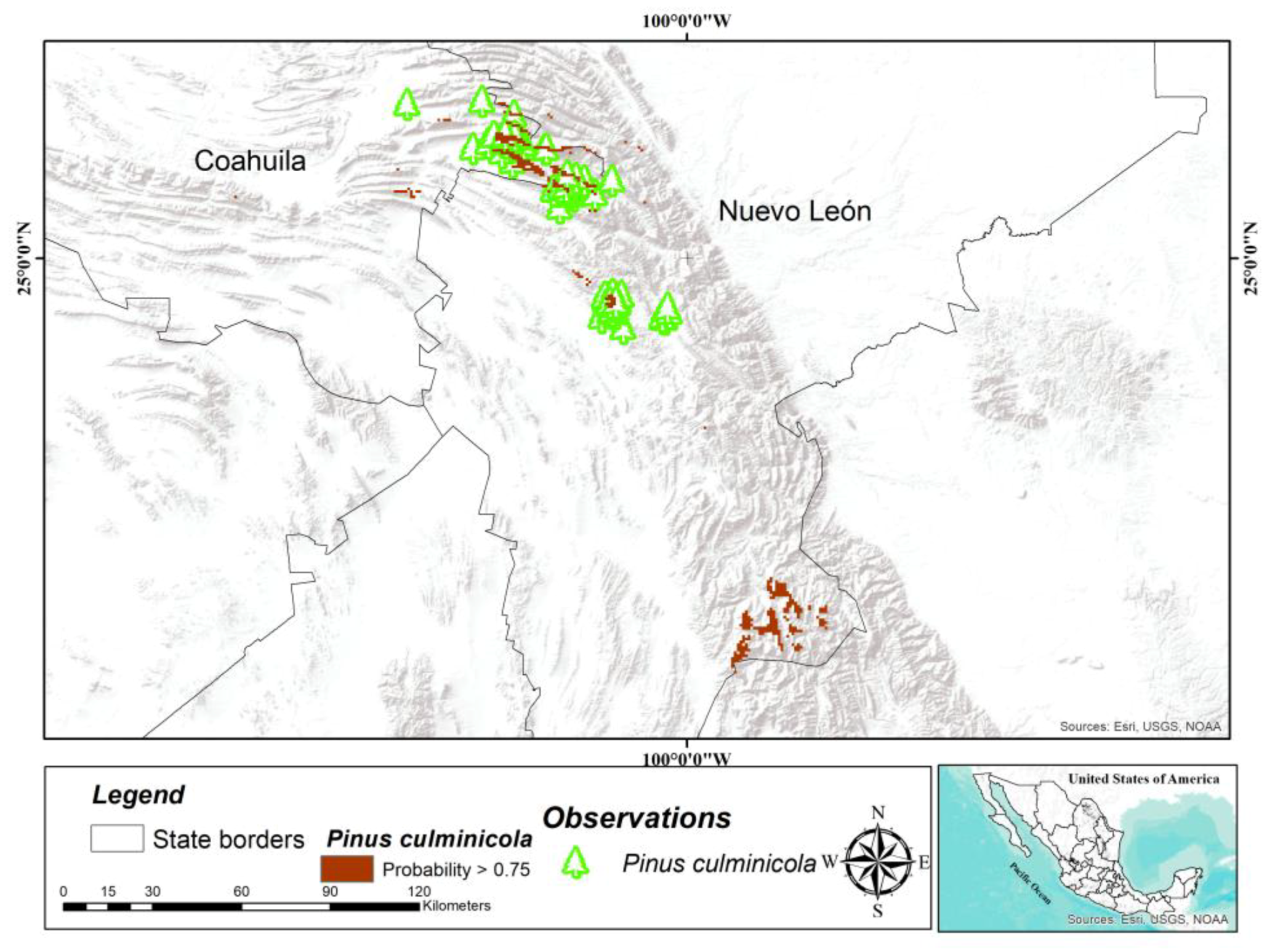
P. culminicola coincided with previous records at Cerro Potosí, Sierra La Viga, and Sierra La Marta in the states of Coahuila and Nuevo León [
32,
33]. The estimated potential distribution areas reaffirm the restricted distribution of this species, which is strongly determined by precipitation in the coldest quarter (Bio19); however, it has been found that the minimum temperature of the coldest month, precipitation in the wettest quarter, and total precipitation also determine the distribution of this species [
33]. Even with the protection of Mexican and international laws, the populations of this species are affected by land use change, road construction, cattle grazing, and forest wildfires [
10], causing the population of this species to undergo drastic reductions. For example, in the Cerro Potosí from 1960 to 1970 the population decreased 66% [
34]. According to some reports [
33,
35,
36],
P. culminicola covers between 10 and 20 km
2; however, the potential area determined in this study is higher (421 km
2), which suggests that there are large areas with adequate conditions for ex situ conservation of this species. Therefore, the specific in situ conservation activities for
P. culminicola should be aimed at reducing the effect of fire and livestock, since no seedling of this species survived the grazing and trampling of cattle after four years [
10].
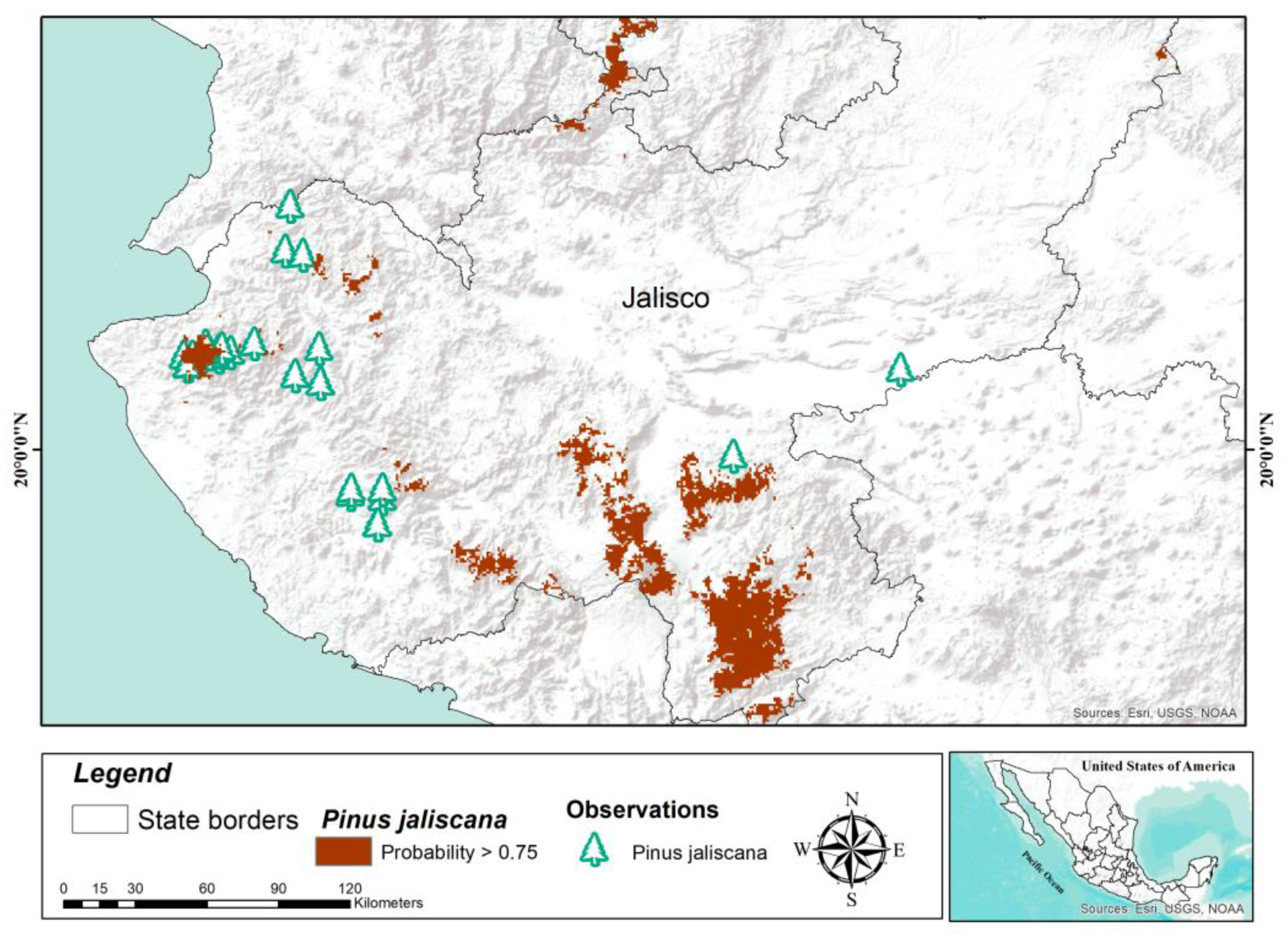
Pinus jaliscana is distributed in nine populations in the west of the state of Jalisco [
37,
38]. Agricultural activities, livestock, and wildfires put natural regeneration at risk in some populations [
38]. It is estimated that this species covers more than 4185 ha [
37], which is lower than what was estimated in this study (5502 km
2). The above suggests a vast area with potential for ex situ conservation of this species, including areas of the north of the state of Jalisco where there are no reports of
P. jaliscana. On the other hand, the seed efficiency rate is 4% [
38], which indicates reproductive problems associated with self-fertilization due to the isolation and small size of the populations. Therefore, in situ conservation activities for this species should be oriented towards increasing the genetic variability of the populations to avoid inbreeding problems.
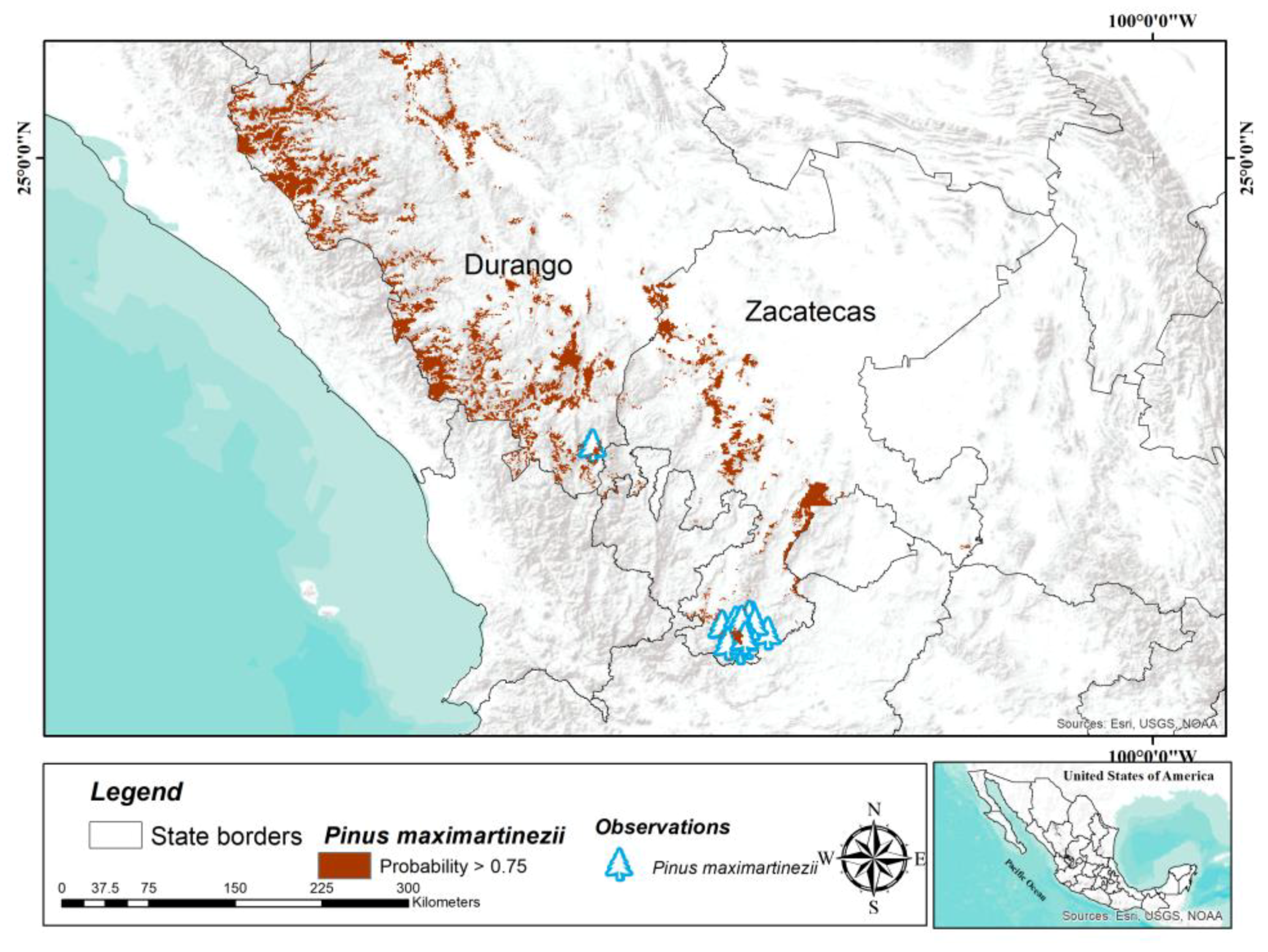
Pinus maximartinezii has only two natural populations, one in the southern state of Zacatecas and another in southern Durango [
39]. The population located in the state of Durango has an area of 110 ha, while the population of southern Zacatecas can have up to 925 ha [
39]; however, the potential distribution area determined in this study is considerably larger (18,355 km
2) and extends north of its distribution, on the Sierra Madre Oriental, in the state of Durango and Zacatecas. Our results suggest the possible existence of other populations that have not yet been located, but also large areas with conditions suitable to establish plantations for ex situ conservation strategies. The seeds of this species are larger and more nutritious than other pine species [
40], so they are used as food; however, even with the use of seeds, the population growth rate of the southern Zacatecas population is 1117, which suggests that the use of seeds does not put this population at risk [
41]. Still, the population of southern Zacatecas has a low percentage of polymorphic loci (30.3%) and only two alleles per polymorphic locus, suggesting a bottleneck effect less than 100 years ago [
42]. The in situ conservation activities for this species should be oriented towards favoring the survival of young and adult individuals during the first reproductive events, protecting the habitat of the species [
41], and increasing genetic diversity through the exchange of genetic material of both populations.
Pinus nelsonii is endemic and restricted in distribution to the Sierra Madre Oriental, in the states of Tamaulipas, San Luis Potosí, Nuevo Leon, and Coahuila [
43,
44]. Due to the decline in its populations, it is categorized as an endangered species by NOM-059-SEMARNAT-2010 (SEMARNAT 2010) and IUCN [
45]. Although, among the pines of the Sierra Madre Oriental,
P. nelsonii is the second most important species [
12], there is little information on the number and size of its populations. It is estimated that it covers only 84 km
2 [
45], smaller than the potential distribution area (814 km
2) defined in this study, but partially coinciding with the current distribution; if we know the zones we can carry out ex situ conservation programs such as protection plantations. The natural populations of this species are severely affected by pests and forest fires. For example, in a population of Tamaulipas, 98% of the plants were affected by the pinyon pitch nodule moth
(Retinia arizonensis) [
44], and 30% of the area was affected by droughts, which reduced their rate of population growth (from 1.083 to 0.990) in a single year [
12]. On the other hand, the variation and genetic differentiation of several populations of this species are low to moderate [
43]. Therefore, the in situ conservation activities of
P. nelsonii should be oriented to avoid forest fires to favor natural regeneration, monitoring and control of sanitation, as well as increasing the genetic diversity of the populations.
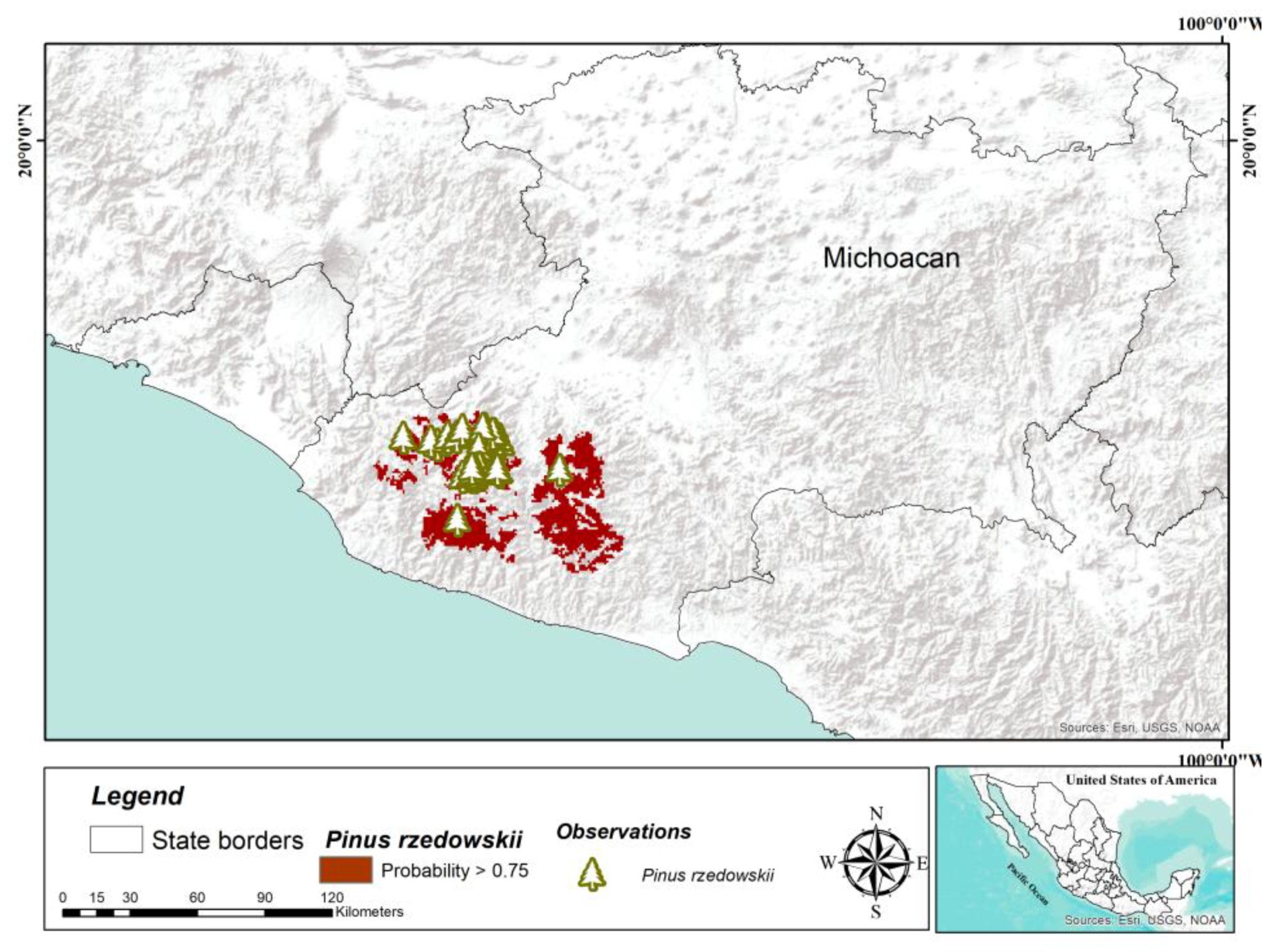
Among the species studied in this work,
P. rzedowskii has a more restricted distribution; it is only distributed in the state of Michoacán [
5]. Only one population with 12 localities is recognized, which in total hold approximately 6500 individuals [
46]. Although the genetic structure is marked, and the genetic diversity is moderate to high [
46,
47], because the populations are tiny, the risk of extinction due to significant forest fires is high [
48]. Therefore, in situ conservation activities for this species should be oriented towards forest fire prevention measures, as well as avoiding activities related to agriculture and grazing. Of the six species studied,
P. rzedowskii has a big potential area of modeled distribution (2936 km
2), which can be explained by its restricted distribution; this area is enough to establish plantations for ex situ conservation.
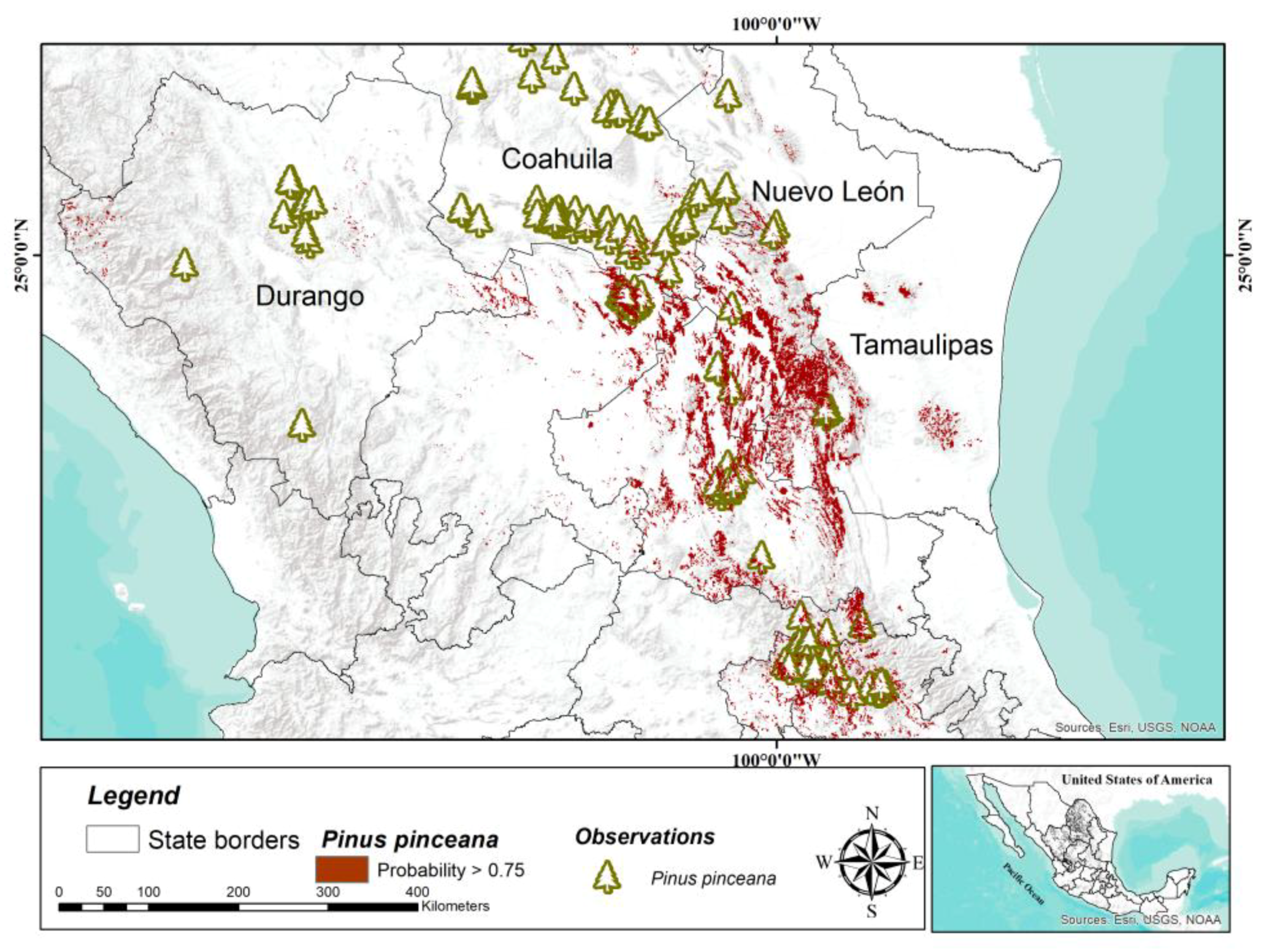
Among the species studied,
P. pinceana has the widest distribution; its distribution extends 750 km along the Sierra Madre Oriental [
49] in the states of Hidalgo, Querétaro, San Luis Potosí, Zacatecas, and Coahuila [
50,
51]. Despite its wide distribution, its populations are fragmented; there is low natural regeneration and potential threats due to the collection of firewood and seeds, but there are no signs of significant recent or current decline, which is why IUCN considers it to be of less concern [
49]. Also, the differentiation and genetic diversity of the populations are high [
11]. Within the natural distribution of
P. pinceana, according to the composition and coefficient of floristic similarity, they differ among three sets of communities [
50], which coincides partially with genetic isolation between populations from the north, center, and south [
11]. The exact area covered by this species is unknown, but it is estimated at 2000 km
2 [
49], which is lower than the potential area modeled in the present study (30,745 km
2). On the other hand, the northeast of Mexico presents conditions of high aptitude for this species, with 10,268 km
2 suitable for the distribution of the species [
52]. These results suggest a large potential area for the establishment of conservation plantations of this species as an ex situ conservation strategy.
For target species, there are seed dispersal limitations due to low tree density, wildfires, grazing, small animals, and extreme climatic conditions: e.g.,
P. culminicola seedlings were not detected in Cerro El Potosí for four years due to them being consumed by rodents [
10], and the survival of the species was strongly affected by low temperatures, strong winds, scarce rainfall, low relative humidity, little soil depth, and loss of edaphic material [
53]. Seedlings of
P. maximartinezi have growth problems in natural stands because of soil degradation due to grazing, fires, agricultural activities, and desiccation [
41].
P. nelsonii populations have fewer than 10,000 mature trees and face two main problems: increased cattle and incidence of destructive fires [
54], both of which reduce seed germination and seedling growth.
In Mexico, the primary strategy of in situ conservation is the establishment of Protected Natural Areas (ANPs). In this sense, the studied species can be conserved in the existing ANPs, and, in the case of
P. pinceana,
P. culminicola, and
P. nelsonii, also in proposed ANPs [
55]. In the present study, although the percentage of the potential distribution area corresponding to ANPs was not determined, between 1.4% (
P. rzedowskii) and 17.0% (
P. maximartinezii) of this surface could encompass ANPs [
31]. However, the network of ANPs in Mexico does not adequately protect the
Pinus species [
31], so the in situ conservation activities discussed for each species are necessary, as is the primary strategy of in situ conservation. Special attention should be given to strategies related to the use of seeds for human consumption; the establishment of Management Units for the Conservation of Wildlife (UMAs) could be proposed as an option because the owners can make sustainable use of the habitat, and at the same time work towards the conservation of the species [
56]. Although in situ conservation is essential to renew genetic diversity and face future environmental changes, ex situ conservation is operationally convenient for short-term results [
57]. It is suggested that the main ex situ conservation strategy should be the establishment of conservation plantations in the areas determined in this study.
The establishment of conservation plantations exclusively in the areas determined for each species should enhance the success and adaptation of the plantations. Also, ex situ conservation strategies should include the collection, storage, and conservation of germplasm in botanical gardens, germplasm banks, and plant tissue culture laboratories in Mexico [
58]. Due to the genetic problems that several of these species present, the germplasm collection for ex situ conservation must be carried out in all the localities or populations of each species. Although Mexico has made significant progress in regulating the use of plant species [
56], legislation must be revised and adapted to favor in situ and ex situ conservation activities with the species studied in this work, as well as others that are at risk.
4.2. Final Considerations
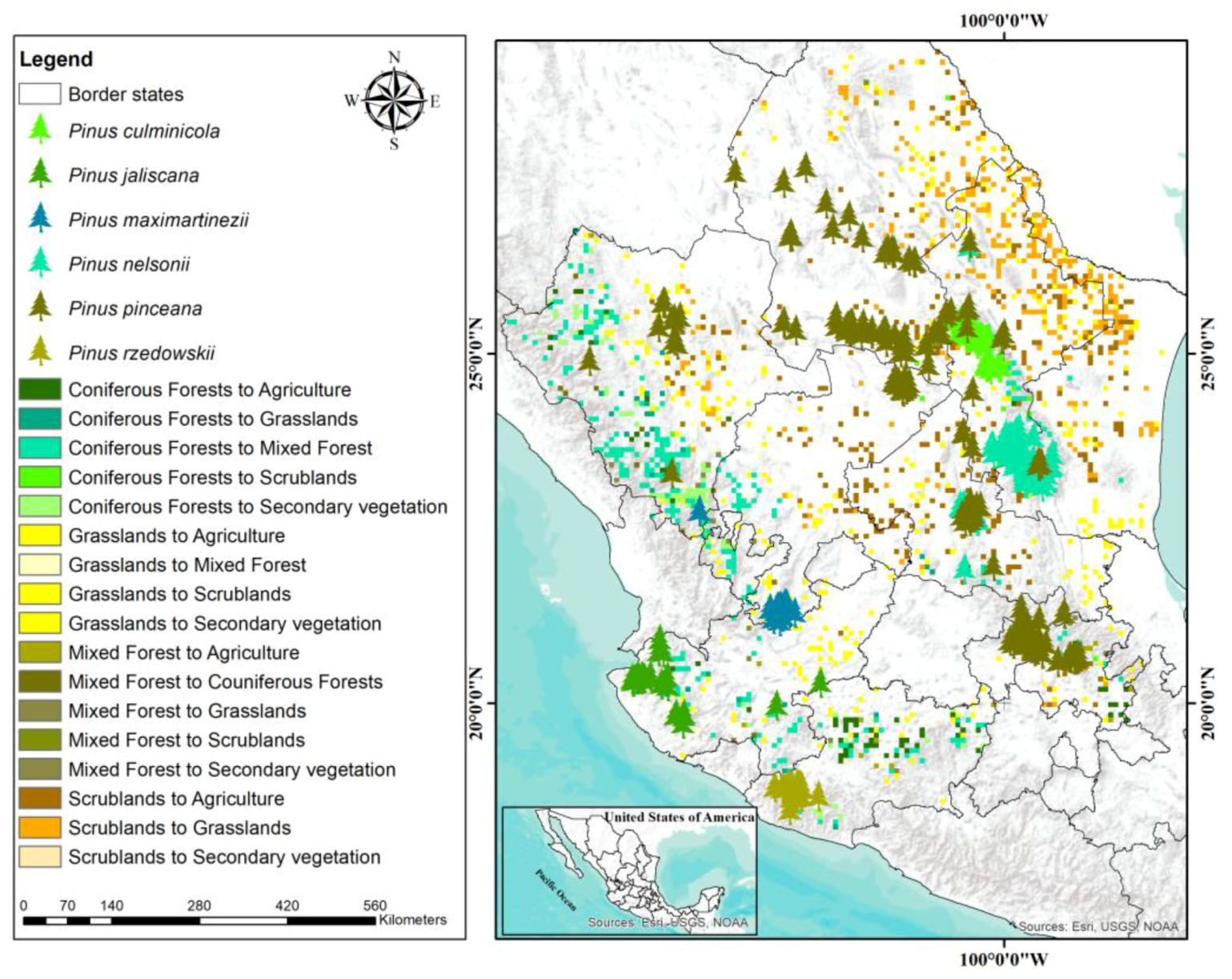
In this study, the primary potential distribution of the target species was located within the temperate coniferous forest from the central part of Mexico (part of the “Sierra Madre Occidental” and “Sierra Madre Oriental” systems), but also within other vegetation communities such as mixed forests and scrubland. Our results are in agreement with some previous reports on the distribution of endangered Mexican pines [
5,
31,
33,
39,
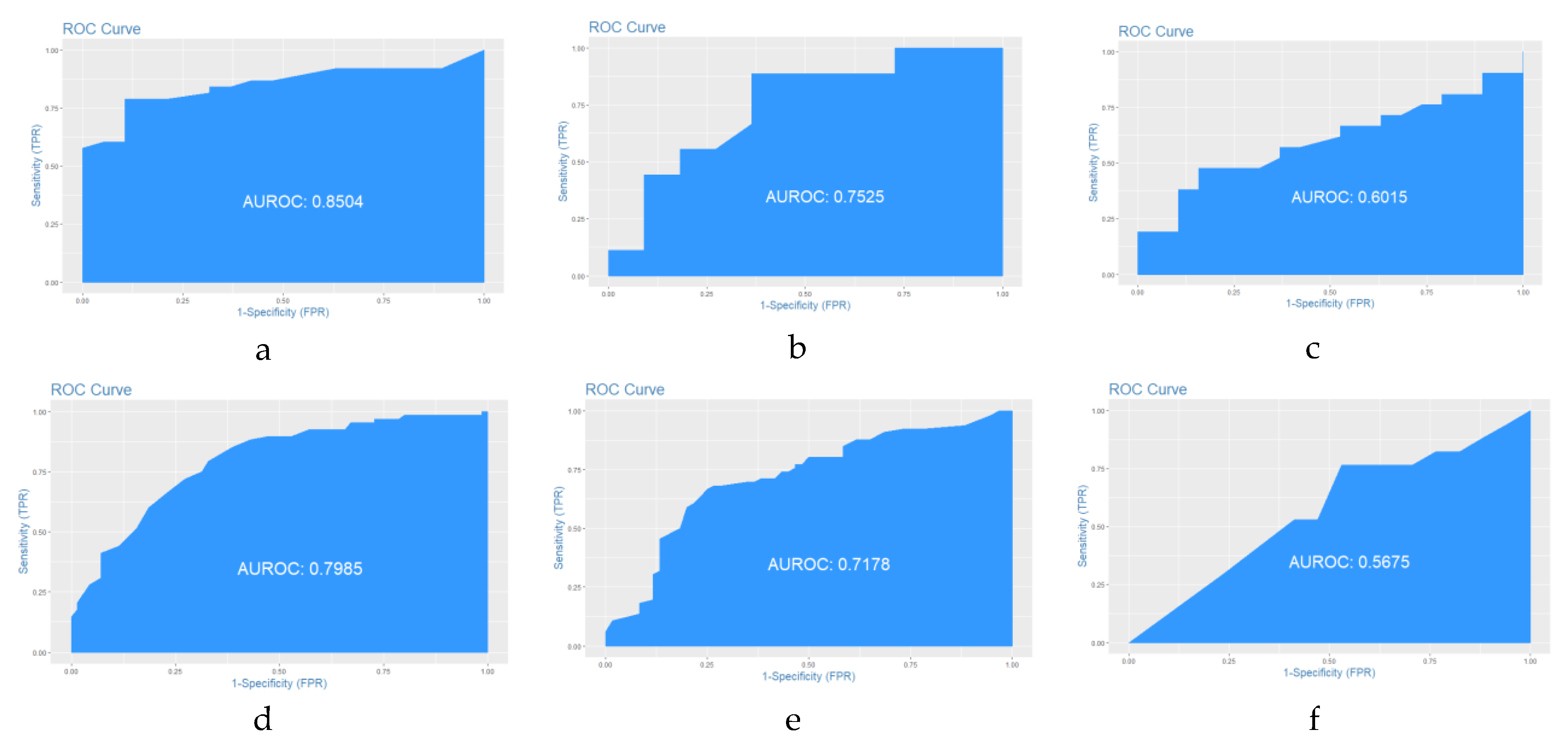
44]. It is important to highlight that the spatial distribution and number of observations collected and retrieved from the diverse databases were an essential factor in the performance of all models and the ability to predict the potential spatial distribution of Mexican pines accurately.
Although it was not part of this study, it would be worth performing an analysis by region including other crucial variables such as distance to roads or population density, as they are considered factors in the vulnerability of Mexican forests [
59]. It is also important to highlight that land cover change is affecting potential suitable areas of distribution of the species. Forest net loss was about 25%. It is imperative to consider land use change and land cover dynamics to promote conservation strategies for each species. The Mexican territory suffers fragmentation, disturbances, and some degradation processes [
59,
60,
61], causing changes in the structure, function, composition, productivity, and extent of forests that may contribute to the diminishing of forest species [
62,
63].
In this paper we used spatial modeling and niche ecological models combined with other analytical tools (e.g., GIS) to evaluate non-inventoried sites as well as model the distribution of species [
14]. The selection of methods used accomplished a double function: first, to provide knowledge about the potential distribution of the species and use said predictions in the choice of sites of particular interest as biological conservation zones [
19,
20]. Although species distribution models (e.g., MaxEnt algorithm) can fit with limited presence-only data [
64], logistic regression was considered a better option as it allows better control of modeling compared to MaxEnt [
16,
65]. Still some issues need to be considered before selecting the appropriate modeling method: the clarification of the niche concept, sampled data, parametrization strategies, model selection and predictor contribution, and evaluation strategies [
66].
Our approach, as we stated before, took advantage of freely available data and, although there was quality control during the modeling, we may have carried over some errors from the data collection, identification of the species, and registration of the geographic data. Still, our results showed good agreement with the performance of all models according to the validation process. As far as we know, there is no maximum number of observations suitable for spatial modeling; however, species distributions models (SDMs) provide the most useful performance using a minimum of 14 to 25 observations for narrow-ranged and widespread species, respectively [
67]. In this study, we had a variety in the number of observations from the field, and assumed that the spatial distribution and number for each pine species were important to the performance of each of the models developed in this study. Our results corroborated the importance and usefulness of diagnosis data for occurrence prediction and their contribution to a better understanding of a species’ reaction to climatic variables [
68].
Finally, we would like to highlight, as future research endeavors, the inclusion of the effects of disturbances, landscape fragmentation and land use dynamics, and climate change on species distribution modeling to provide more comprehensive and accurate results for the implementation of conservation strategies.

 ,
, 












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}