Long-Term Abandonment of Forest Management Has a Strong Impact on Tree Morphology and Wood Volume Allocation Pattern of European Beech (Fagus sylvatica L.)

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites
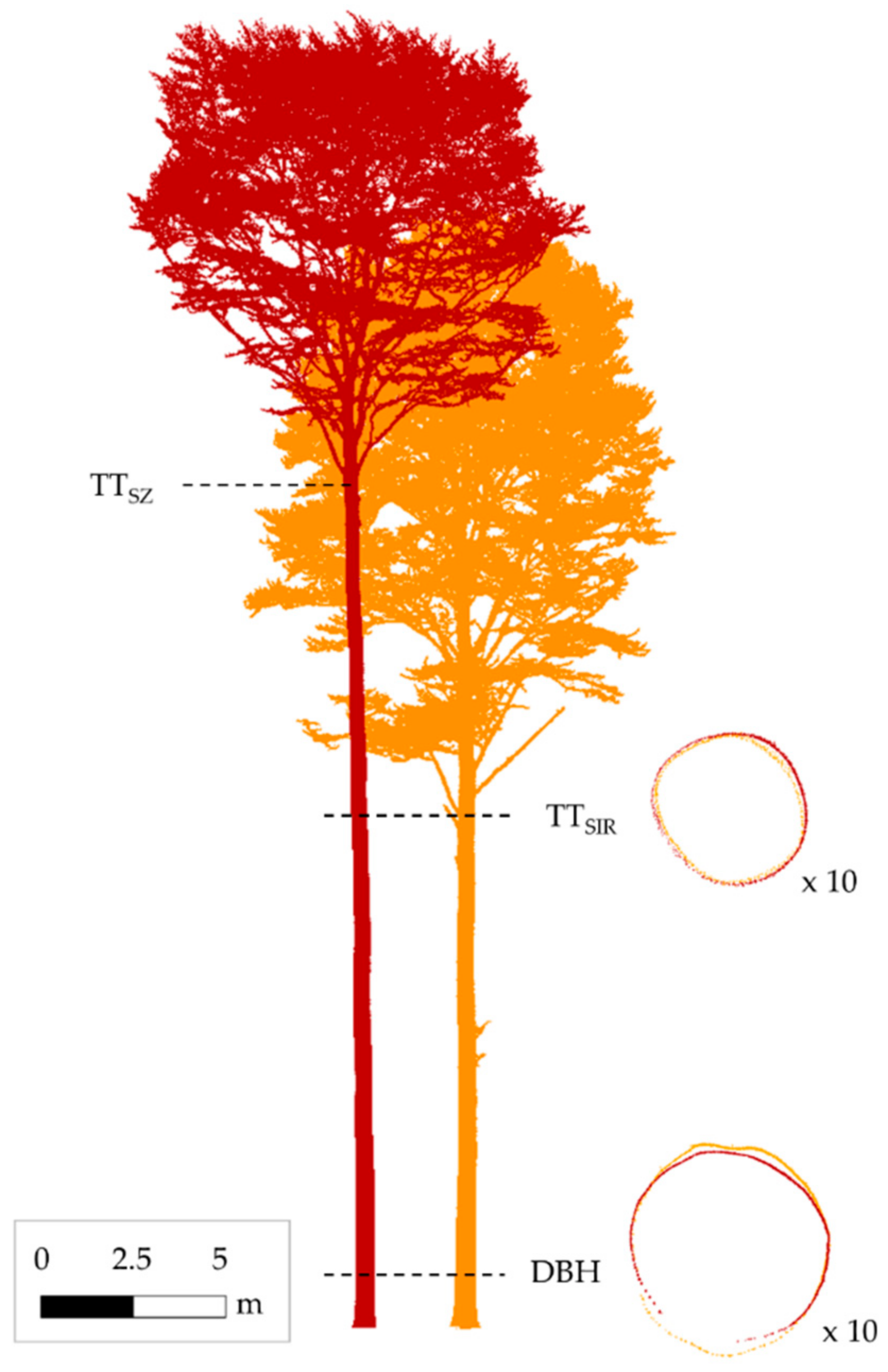
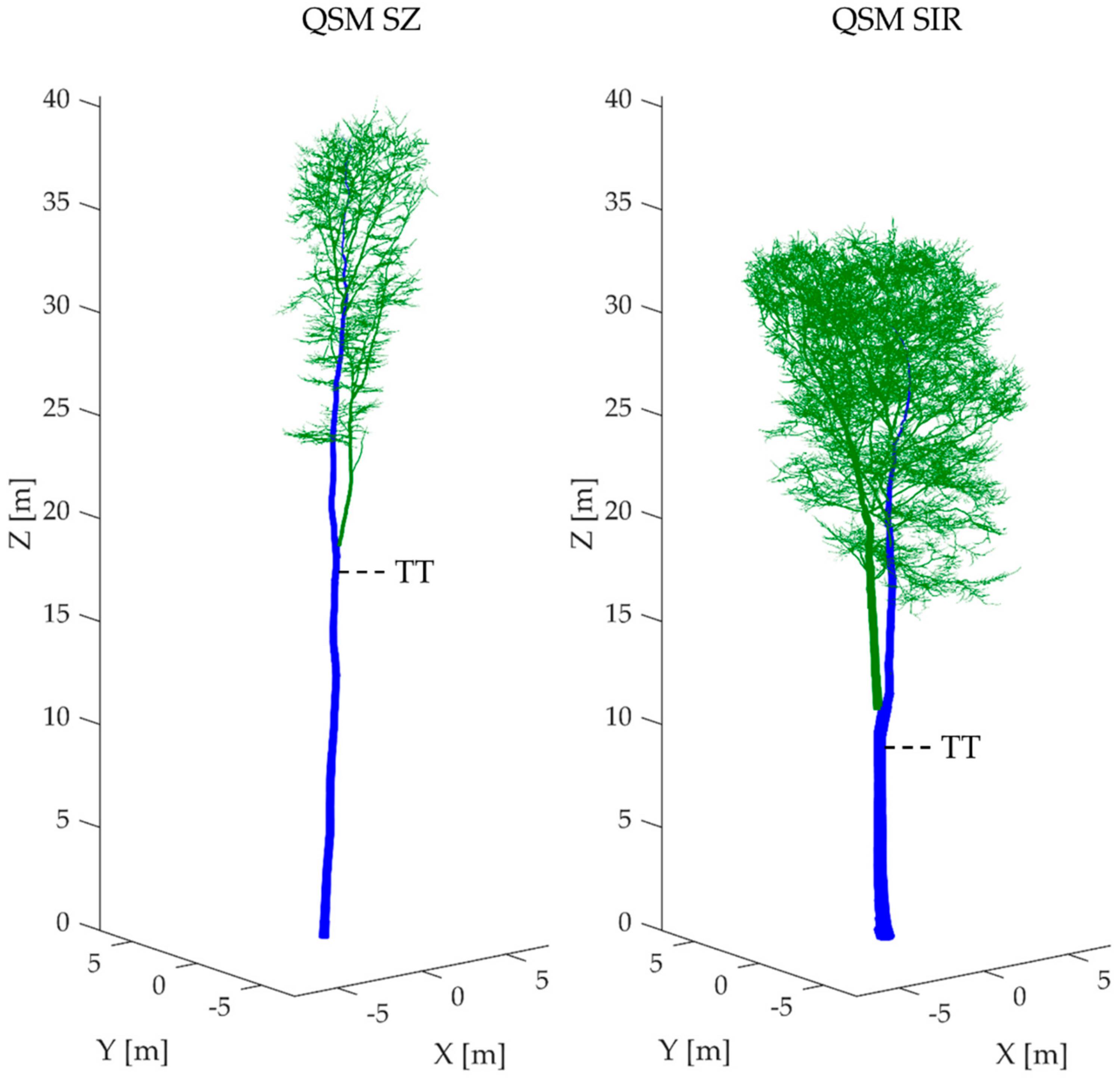
2.2. TLS Data Acquisition and Registration
2.3. TLS Data Post-Processing
2.4. Statistical Analysis
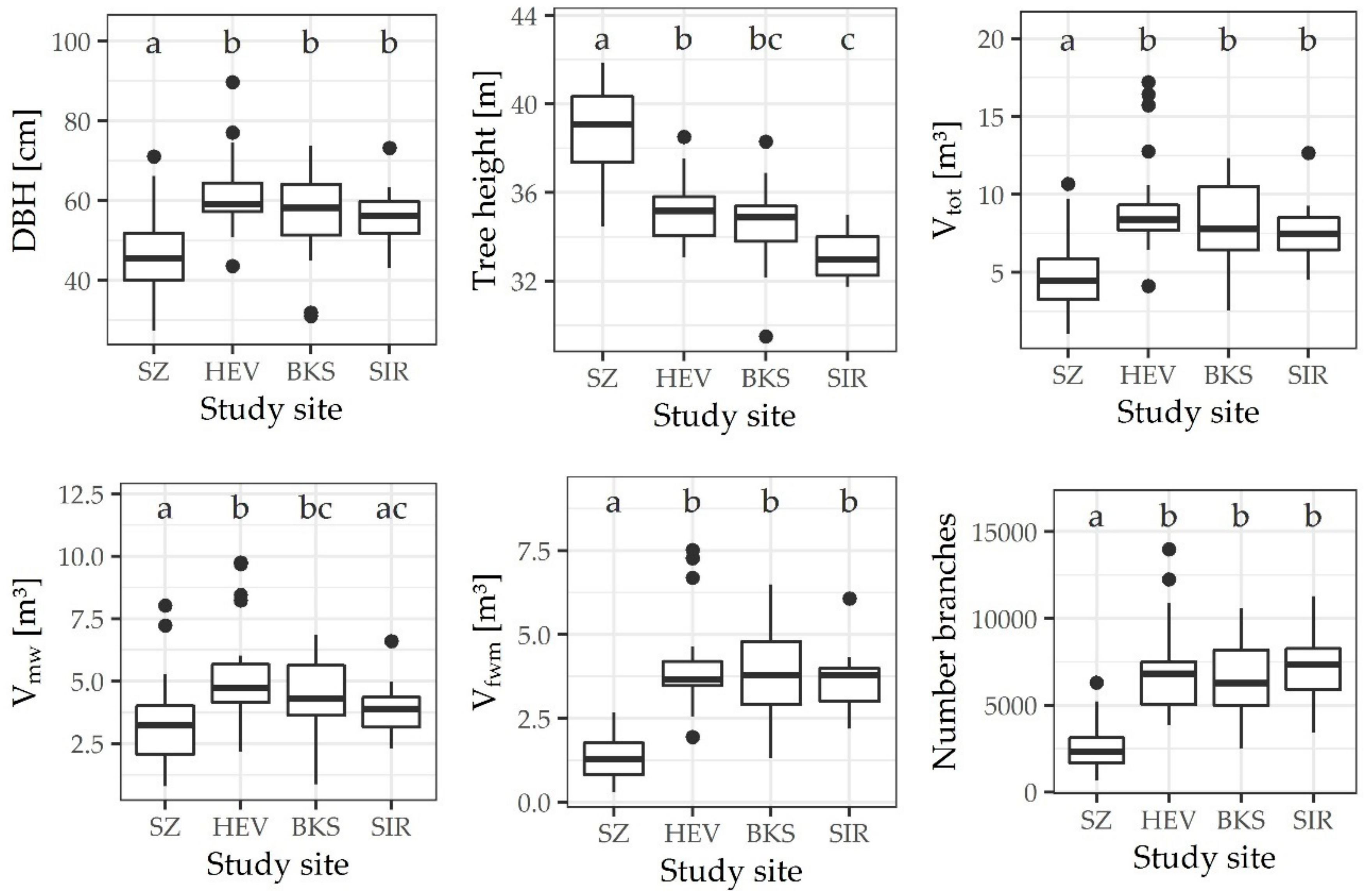
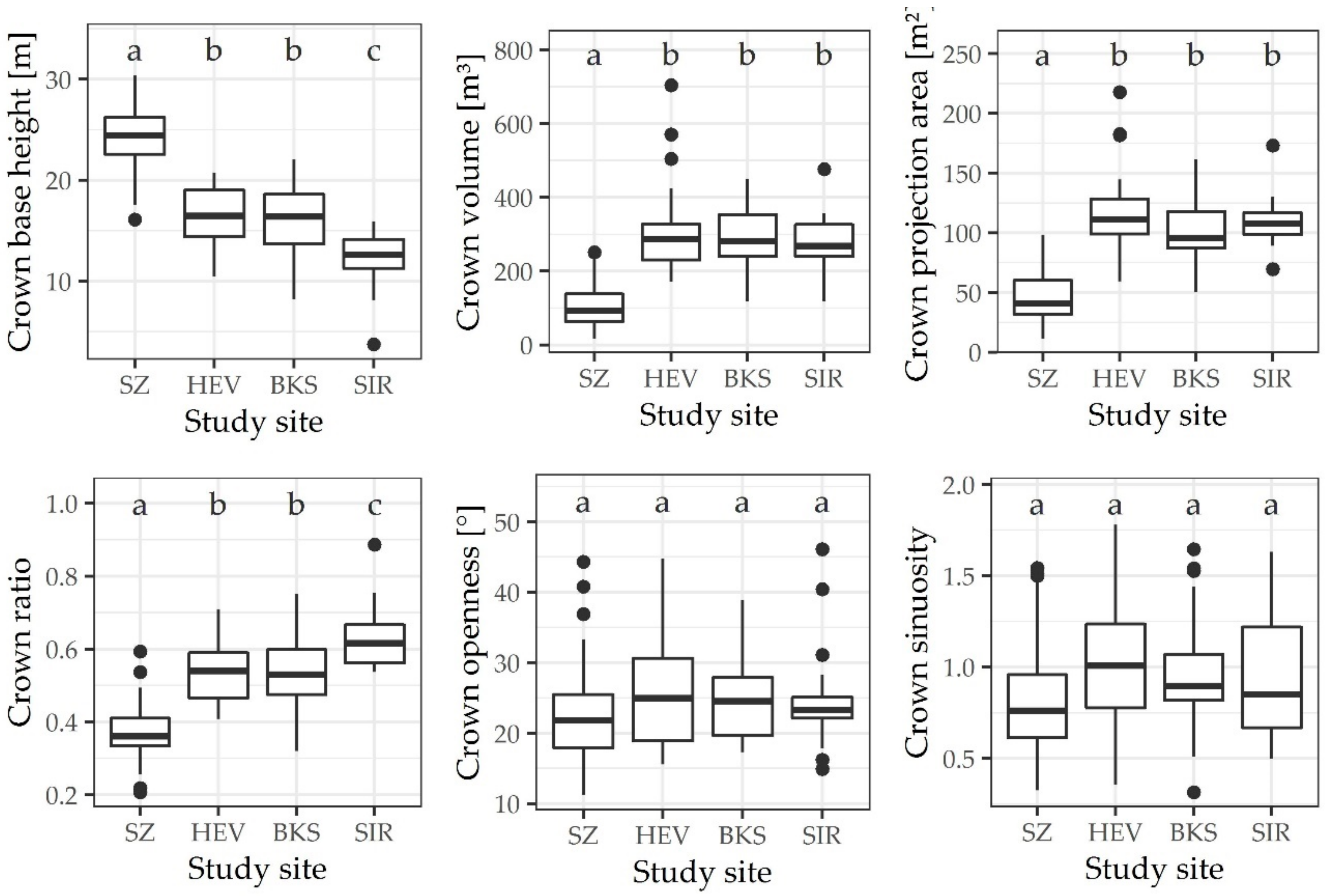
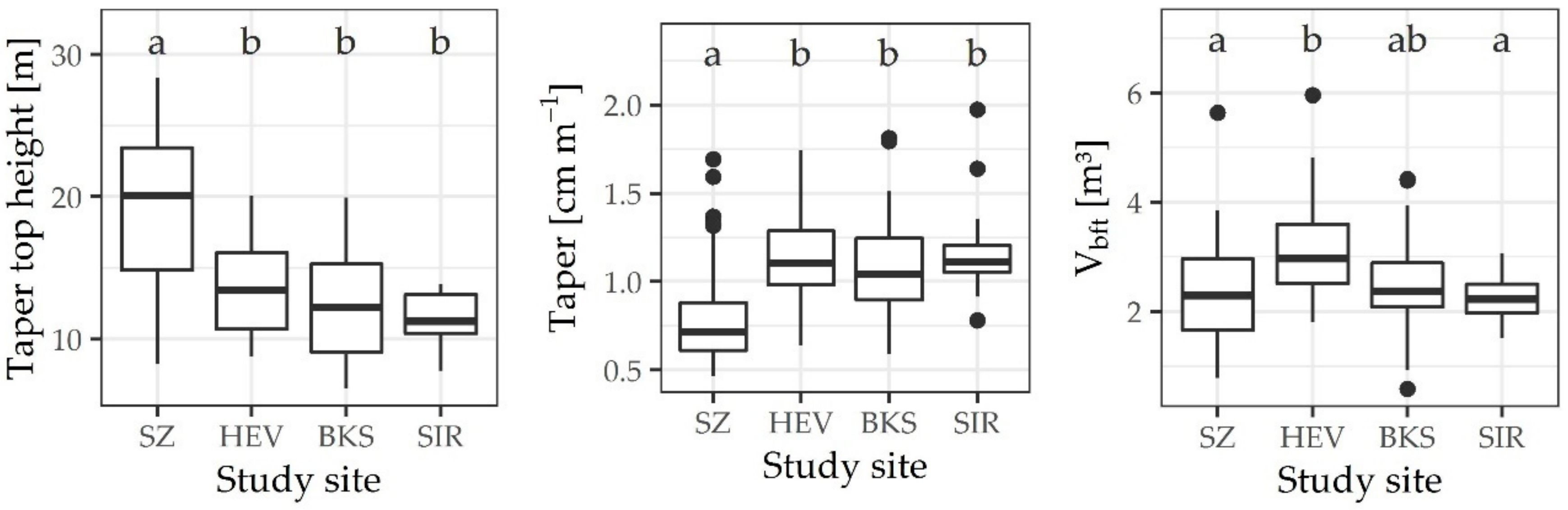
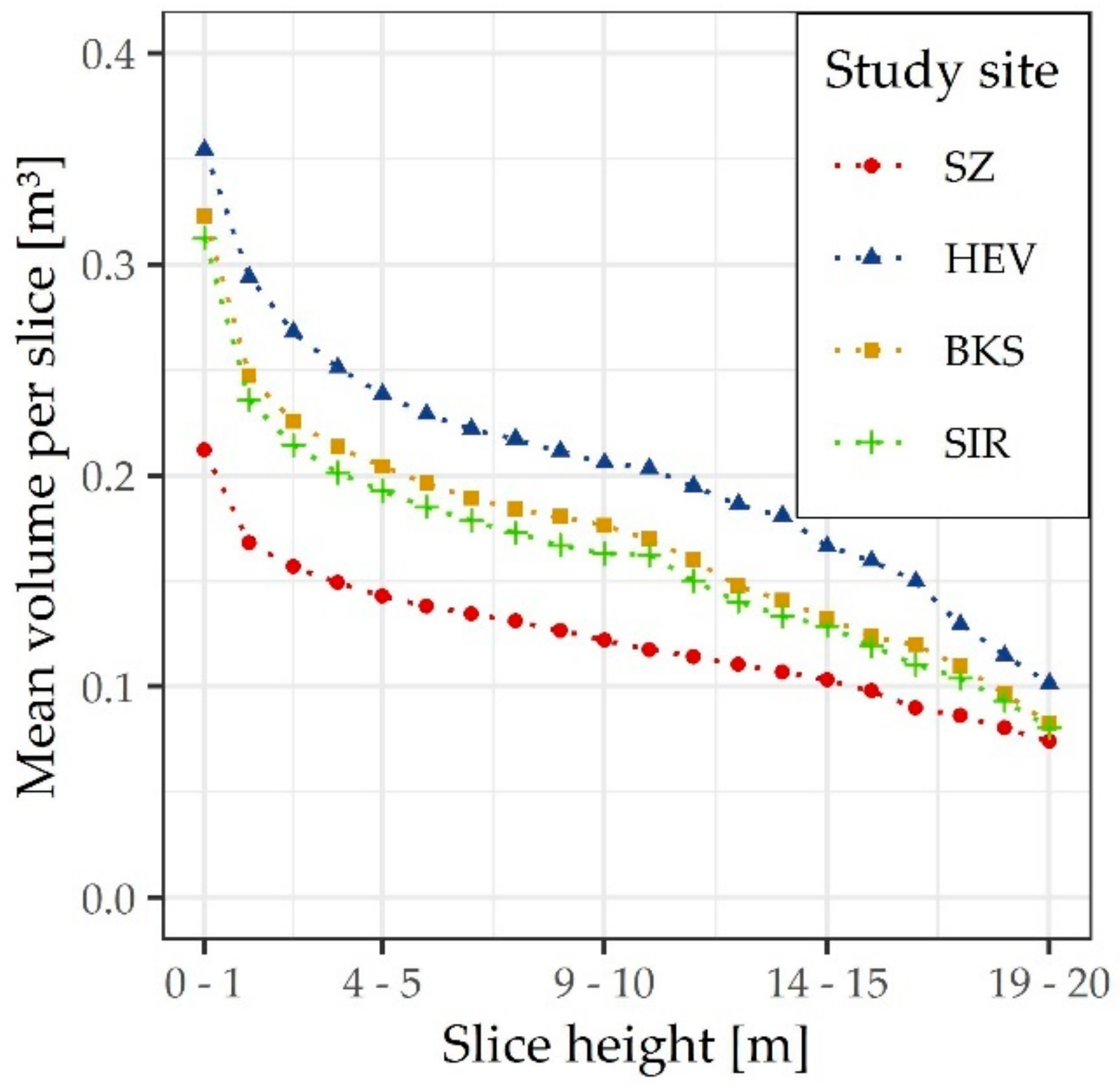
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Site | SZ | HEV | BKS | SIR |
|---|---|---|---|---|
| Height-diameter ratio | 0.86 | 0.58 | 0.63 | 0.60 |
| a | b | b | b | |
| Mean branch length sum [m] | 1515.6 | 4391.9 | 4072.7 | 4479.9 |
| a | b | b | b | |
| Mean maximum branch order | 8.3 | 9.5 | 9.5 | 9.2 |
| a | b | b | b | |
| Mean branch number 1st order | 44.7 | 39.1 | 44.9 | 53.8 |
| ab | a | ab | b | |
| Mean branch number 2nd order | 274.6 | 389.9 | 407.3 | 559.6 |
| a | b | b | c | |
| Branch volume 1st order [m3] | 638.9 | 1213.0 | 948.1 | 898.6 |
| a | b | ab | ab | |
| Branch volume 2nd order [m3] | 392.4 | 949.0 | 902.3 | 901.9 |
| a | b | b | b | |
| Branch length 1st order [m] | 110.8 | 165.0 | 158.2 | 200.9 |
| a | b | b | c | |
| Branch length 2nd order [m] | 302.1 | 586.9 | 569.5 | 729.1 |
| a | b | b | c | |
| Crown surface area [m2] | 574.9 | 1640.3 | 1437.8 | 1669.9 |
| a | b | b | b | |
| Crown length [m] | 14.7 | 19.3 | 19.2 | 21.4 |
| a | b | b | b | |
| Crown length-width ratio | 2.09 | 1.62 | 1.71 | 1.83 |
| a | b | b | ab | |
| Crown roughness | 0.33 | 0.25 | 0.25 | 0.23 |
| a | b | b | b |
References
- West, G.B.; Brown, J.H.; Enquist, B.J. A general model for the origin of allometric scaling laws in biology. Science 1997, 276, 122–126. [Google Scholar] [CrossRef] [PubMed]
- Valladares, F.; Niinemets, U. The architecture of plant crowns: From design rules to light capture and performance. In Functional Plant Ecology; Pugnaire, F., Valladares, F., Eds.; Taylor and Francis: New York, NY, USA, 2007; pp. 101–149. [Google Scholar]
- Malhi, Y.; Jackson, T.; Patrick Bentley, L.; Lau, A.; Shenkin, A.; Herold, M.; Calders, K.; Bartholomeus, H.; Disney, M.I. New perspectives on the ecology of tree structure and tree communities through terrestrial laser scanning. Interface Focus 2018, 8, 20170052. [Google Scholar] [CrossRef] [PubMed]
- Maas, H.G.; Bienert, A.; Scheller, S.; Keane, E. Automatic forest inventory parameter determination from terrestrial laser scanner data. Int. J. Remote Sens. 2008, 29, 1579–1593. [Google Scholar] [CrossRef]
- Hess, C.; Bienert, A.; Härdtle, W.; von Oheimb, G. Does tree architectural complexity influence the accuracy of wood volume estimates of single young trees by terrestrial laser scanning? Forests 2015, 6, 3847–3867. [Google Scholar] [CrossRef]
- Liang, X.; Kankare, V.; Hyyppä, J.; Wang, Y.; Kukko, A.; Haggrén, H.; Yu, X.; Kaartinen, H.; Jaakkola, A.; Guan, F.; et al. Terrestrial laser scanning in forest inventories. J. Photogramm. Remote Sens. 2016, 115, 63–77. [Google Scholar] [CrossRef]
- Leuschner, C.; Ellenberg, H. Ecology of Central European Forests: Vegetation Ecology of Central Europe; Springer: Berlin, Germany, 2017; Volume I, p. 971. [Google Scholar]
- Bayer, D.; Seifert, S.; Pretzsch, H. Structural crown properties of Norway spruce (Picea abies L. Karst.) and European beech (Fagus sylvatica L.) in mixed versus pure stands revealed by terrestrial laser scanning. Trees 2013, 27, 1035–1047. [Google Scholar] [CrossRef]
- Metz, J.; Seidel, D.; Schall, P.; Scheffer, D.; Schulze, E.D.; Ammer, C. Crown modeling by terrestrial laser scanning as an approach to assess the effect of aboveground intra- and interspecific competition on tree growth. For. Ecol. Manag. 2013, 310, 275–288. [Google Scholar] [CrossRef]
- Barbeito, I.; Dassot, M.; Bayer, D.; Collet, C.; Drössler, L.; Löf, M.; Del Rio, M.; Ruiz-Peinado, R.; Forrester, D.I.; Bravo-Oviedo, A.; et al. Terrestrial laser scanning reveals differences in crown structure of Fagus sylvatica in mixed vs pure European forests. For. Ecol. Manag. 2017, 405, 381–390. [Google Scholar] [CrossRef]
- Höwler, K.; Annighöfer, P.; Ammer, C.; Seidel, D. Competition improves quality-related external stem characteristics of Fagus sylvatica. Can. J. For. Res. 2017, 47, 1603–1613. [Google Scholar] [CrossRef]
- Seidel, D.; Leuschner, C.; Müller, A.; Krause, B. Crown plasticity in mixed forests—Quantifying asymmetry as a measure of competition using terrestrial laser scanning. For. Ecol. Manag. 2011, 261, 2123–2132. [Google Scholar] [CrossRef]
- Dieler, J.; Pretzsch, H. Morphological plasticity of European beech (Fagus sylvatica L.) in pure and mixed-species stands. For. Ecol. Manag. 2013, 295, 97–108. [Google Scholar] [CrossRef]
- Schröter, M.; Härdtle, W.; von Oheimb, G. Crown plasticity and neighborhood interactions of European beech (Fagus sylvatica L.) in an old-growth forest. Eur. J. For. Res. 2012, 131, 787–798. [Google Scholar] [CrossRef]
- Engel, M.; Körner, M.; Berger, U. Plastic tree crowns contribute to small-scale heterogeneity in virgin beech forests—An individual-based modeling approach. Ecol. Model. 2018, 376, 28–39. [Google Scholar] [CrossRef]
- Fichtner, A.; Sturm, K.; Rickert, C.; von Oheimb, G.; Härdtle, W. Crown size-growth relationships of European beech (Fagus sylvatica L.) are driven by the interplay of disturbance intensity and inter-specific competition. For. Ecol. Manag. 2013, 302, 178–184. [Google Scholar] [CrossRef]
- Seidel, D.; Schall, P.; Gille, M.; Ammer, C. Relationship between tree growth and physical dimensions of Fagus sylvatica crowns assessed from terrestrial laser scanning. iForest 2015, 8, 735–742. [Google Scholar] [CrossRef]
- Juchheim, J.; Annighöfer, P.; Ammer, C.; Calders, K.; Raumonen, P.; Seidel, D. How management intensity and neighborhood composition affect the structure of beech (Fagus sylvatica L.) trees. Trees 2017, 31, 1723–1735. [Google Scholar] [CrossRef]
- Mausolf, K.; Wilm, P.; Härdtle, W.; Jansen, K.; Schuldt, B.; Sturm, K.; von Oheimb, G.; Hertel, D.; Leuschner, C.; Fichtner, A. Higher drought sensitivity of radial growth of European beech in managed than in unmanaged forests. Sci. Total Environ. 2018, 642, 1201–1208. [Google Scholar] [CrossRef] [PubMed]
- Petzold, J.; Dittrich, S.; Fichtner, A.; Härdtle, W.; Naumann, B.; von Oheimb, G. Effects of forest management intensity on herbaceous plant diversity and composition of deciduous forest communities in Northern Germany. Tuexenia 2018, 38, 79–96. [Google Scholar] [CrossRef]
- Stiers, M.; Willim, K.; Seidel, D.; Ehbrecht, M.; Kabal, M.; Ammer, C.; Annighöfer, P. A quantitative comparison of the structural complexity of managed, lately unmanaged and primary European beech (Fagus sylvatica L.) forests. For. Ecol. Manag. 2018, 430, 357–365. [Google Scholar] [CrossRef]
- Deutscher Wetterdienst. Klimareport Schleswig-Holstein; Deutscher Wetterdienst: Offenbach am Main, Germany, 2017; p. 44.
- Stadtwald Lübeck. Konzept der Naturnahen Waldwirtschaft und Forstplanung im Stadtwald Lübeck. Ergebnisse aus Waldbiotopkartierung, Forstinventur und Betriebsprüfung; Forstamt der Stadt Lübeck: Lübeck, Germany, 2004.
- Sturm, K. Grundlagen und Ziele des integrativen Prozessschutz-Waldbaus. In Natura 2000 im Wald—Lebensraumtypen, Erhaltungszustand, Management; Lehrke, S., Ellwanger, G., Buschmann, A., Frederking, W., Paulsch, C., Schröder, E., Ssymank, A., Eds.; Landwirtschaftsvlg: Münster, Germany, 2013; Volume 131, pp. 219–232. [Google Scholar]
- Pfennigbauer, M.; Ullrich, A. Improving quality of laser scanning data acquisition through calibrated amplitude and pulse deviation measurement. In Proceedings of the SPIE Defense, Security and Sensing, Orlando, FL, USA, 29 April 2010. [Google Scholar] [CrossRef]
- Riegl RIEGL Product Line–Innovations in 3D. Available online: https://user-539731.cld.bz/RIEGL-Online-Catalog-2017-20182 (accessed on 29 August 2018).
- Pretzsch, H. Forests Dynamics, Growth and Yield; Springer: Berlin, Germany, 2009; p. 664. [Google Scholar]
- Hackenberg, J.; Wassenberg, M.; Spiecker, H.; Sun, D. Non-destructive method for biomass prediction combining TLS derived tree volume and wood density. Forests 2015, 6, 1274–1300. [Google Scholar] [CrossRef]
- Raumonen, P.; Kaasalainen, M.; Åkerblom, M.; Kaasalainen, S.; Kaartinen, H.; Vastaranta, M.; Holopainen, M.; Disney, M.; Lewis, P. Fast automatic precision tree models from terrestrial laser scanner data. Remote Sens. 2013, 5, 491–520. [Google Scholar] [CrossRef]
- Calders, K.; Newnham, G.; Burt, A.; Murphy, S.; Raumonen, P.; Herold, M.; Culvenor, D.; Avitabile, V.; Disney, M.; Armston, J.; et al. Nondestructive estimates of above-ground biomass using terrestrial laser scanning. Methods Ecol. Evol. 2015, 6, 198–208. [Google Scholar] [CrossRef]
- Bienert, A.; Georgi, L.; Kunz, M.; Maas, H.G.; von Oheimb, G. Comparison and combination of mobile and terrestrial laser scanning for natural forest inventories. Forests 2018, 9, 395. [Google Scholar] [CrossRef]
- Raumonen, P.; Casella, E.; Calders, K.; Murphy, S.; Åkerblom, M.; Kaasalainen, M. Massive-scale tree modelling from TlS data. ISPRS Ann. Photogramm. Remote Sens. Spat. Inf. Sci. 2015, II-3/W4, 189–196. [Google Scholar] [CrossRef]
- Kunz, M.; Hess, C.; Raumonen, P.; Bienert, A.; Hackenberg, J.; Maas, H.G.; Härdtle, W.; Fichtner, A.; von Oheimb, G. Comparison of wood volume estimates of young trees from terrestrial laser scan data. iForest 2017, 10, 451–458. [Google Scholar] [CrossRef]
- Rusu, R.B.; Cousins, S. 3D is here: Point cloud library (PCL). In Proceedings of the ‘IEEE International Conference on Robotics and Automation (ICRA), Shanghai, China, 9–13 May 2011. [Google Scholar] [CrossRef]
- Kai, T.; Da, F.; Yvinec, M. 3D Alpha Shapes. Available online: http://www.ics.uci.edu/~dock/manuals/ cgal_manual/Alpha_shapes_3/Chapter_main.html (accessed on 29 August 2018).
- Martin-Ducup, O.; Robert, S.; Fournier, R.A. Response of sugar maple (Acer saccharum, Marsh.) tree crown structure to competition in pure versus mixed stands. For. Ecol. Manag. 2016, 374, 20–32. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Lang, A.C.; Härdtle, W.; Bruelheide, H.; Geißler, C.; Nadrowski, K.; Schuldt, A.; von Oheimb, G. Tree morphology responds to neighbourhood competition and slope in species-rich forests of subtropical China. For. Ecol. Manag. 2010, 260, 1708–1715. [Google Scholar] [CrossRef]
- Dassot, M.; Constant, T.; Ningre, F.; Fournier, M. Impact of stand density on tree morphology and growth stress in young beech (Fagus sylvatica L.) stands. Trees 2015, 29, 583–591. [Google Scholar] [CrossRef]
- Von Oheimb, G.; Lang, A.C.; Bruelheide, H.; Forrester, D.I.; Wäsche, I.; Yu, M.; Härdtle, W. Individual-tree radial growth in a subtropical broad-leaved forest: The role of local neighbourhood competition. Forest Ecol. Manag. 2011, 261, 499–507. [Google Scholar] [CrossRef]
- Fichtner, A.; Härdtle, W.; Bruelheide, H.; Kunz, M.; Li, Y.; von Oheimb, G. Neighbourhood interactions drive overyielding in mixed-species tree communities. Nat. Commun. 2018, 9, 1144. [Google Scholar] [CrossRef] [PubMed]
- Pretzsch, H. Canopy space filling and tree crown morphology in mixed-species stands compared with monocultures. For. Ecol. Manag. 2014, 327, 251–264. [Google Scholar] [CrossRef]
- Fichtner, A.; Härdtle, W.; Li, Y.; Bruelheide, H.; Kunz, M.; von Oheimb, G. From competition to facilitation: How tree species respond to neighbourhood diversity. Ecol. Lett. 2017, 20, 892–900. [Google Scholar] [CrossRef] [PubMed]
- Fichtner, A.; Forrester, D.I.; Härdtle, W.; Sturm, K.; von Oheimb, G. Facilitative-competitive interactions in an old-growth forest: The importance of large-diameter trees as benefactors and stimulators for forest community assembly. PLoS ONE 2015, 10, E0120335. [Google Scholar] [CrossRef] [PubMed]







| Schattiner Zuschlag (SZ) | Hevenbruch (HEV) | Berkenstrücken (BKS) | Sirksfelder Zuschlag (SIR) | |
|---|---|---|---|---|
| Management intensity | Unmanaged > 50 years | Unmanaged > 20 years | Extensively managed | Intensively managed |
| Elevation (m a.s.l.) | 58 | 75 | 80 | 65 |
| Soil type | (pseudogleyic) luvisol | (pseudogleyic) luvisol | (pseudogleyic) luvisol | (pseudogleyic) luvisol |
| Age (years) | 109 | 121 | 105 | 110 |
| Stem density (n ha−1) | 369 | 194 | 188 | 104 |
| Stand volume (m3 ha−1) | 903 | 690 | 652 | 392 |
| Stand basal area (m2 ha−1) | 59 | 34 | 39 | 25 |
| Mean diameter at breast height (cm) | 44 | 45 | 47 | 54 |
| Mean tree height (m) | 41 | 36 | 39 | 34 |
| Measure | Abbreviation | Origin | Reference/Calculation |
|---|---|---|---|
| Diameter at breast height (cm) | DBH | QSM | Raumonen et al. (2013) |
| Tree height (m) | TH | Point cloud | Zmax-Zmin |
| Total wood volume (m3) | Vtot | QSM | Raumonen et al. (2013) |
| Total branch number | QSM | Raumonen et al. (2013) | |
| Height-to-diameter ratio | H/D-ratio | Point cloud | TH/DBH 100 |
| Merchantable wood volume (m3) | Vmw | QSM | See Section 2.3. this publication |
| Volume of fine woody material (m3) | Vfwm | QSM | See Section 2.3. this publication |
| Crown base height (m) | CBH | Point cloud | See Section 2.3. this publication |
| Crown volume (m3) | CV | Point cloud | See Section 2.3. this publication |
| Crown projection area (m2) | CPA | Point cloud | See Section 2.3. this publication |
| Crown surface area (m2) | CSA | Point cloud | See Section 2.3. this publication |
| Crown ratio | CR | Point cloud | CL/TH |
| Crown length (m) | CL | Point cloud | TH-CBH |
| Crown openness (°) | Point cloud | Martin-Ducup et al. (2016) | |
| Crown sinuosity | Point cloud | Martin-Ducup et al. (2016) | |
| Taper (cm m−1) | Point cloud | See Section 2.3. this publication | |
| Taper top height (m) | TT | Point cloud | See Section 2.3. this publication |
| Volume of branch-free trunk (m3) | Vbft | QSM | See Section 2.3. this publication |
| Mean branch length sum (m) | QSM | Raumonen et al. (2013) | |
| Mean maximum branch order | QSM | Raumonen et al. (2013) | |
| Mean branch number 1st order | QSM | Raumonen et al. (2013) | |
| Mean branch number 2nd order | QSM | Raumonen et al. (2013) | |
| Branch volume 1st order (m3) | QSM | Raumonen et al. (2013) | |
| Branch volume 2nd order (m3) | QSM | Raumonen et al. (2013) | |
| Branch length 1st order (m) | QSM | Raumonen et al. (2013) | |
| Branch length 2nd order (m) | QSM | Raumonen et al. (2013) | |
| Crown length-width ratio | Point cloud | CL/(sqrt(CPA 4 π−1)) | |
| Crown roughness | Point cloud | Martin-Ducup et al. (2016) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Georgi, L.; Kunz, M.; Fichtner, A.; Härdtle, W.; Reich, K.F.; Sturm, K.; Welle, T.; Oheimb, G.v. Long-Term Abandonment of Forest Management Has a Strong Impact on Tree Morphology and Wood Volume Allocation Pattern of European Beech (Fagus sylvatica L.). Forests 2018, 9, 704. https://doi.org/10.3390/f9110704
Georgi L, Kunz M, Fichtner A, Härdtle W, Reich KF, Sturm K, Welle T, Oheimb Gv. Long-Term Abandonment of Forest Management Has a Strong Impact on Tree Morphology and Wood Volume Allocation Pattern of European Beech (Fagus sylvatica L.). Forests. 2018; 9(11):704. https://doi.org/10.3390/f9110704
Chicago/Turabian StyleGeorgi, Louis, Matthias Kunz, Andreas Fichtner, Werner Härdtle, Karl Friedrich Reich, Knut Sturm, Torsten Welle, and Goddert von Oheimb. 2018. "Long-Term Abandonment of Forest Management Has a Strong Impact on Tree Morphology and Wood Volume Allocation Pattern of European Beech (Fagus sylvatica L.)" Forests 9, no. 11: 704. https://doi.org/10.3390/f9110704
APA StyleGeorgi, L., Kunz, M., Fichtner, A., Härdtle, W., Reich, K. F., Sturm, K., Welle, T., & Oheimb, G. v. (2018). Long-Term Abandonment of Forest Management Has a Strong Impact on Tree Morphology and Wood Volume Allocation Pattern of European Beech (Fagus sylvatica L.). Forests, 9(11), 704. https://doi.org/10.3390/f9110704