Contribution of Ecological and Socioeconomic Factors to the Presence and Abundance of Invasive Tree Species in Mississippi, USA

Abstract
:1. Introduction
2. Materials and Methods
2.1. Target Invasive Tree Species
2.2. Data Sources
2.3. Estimation Methods
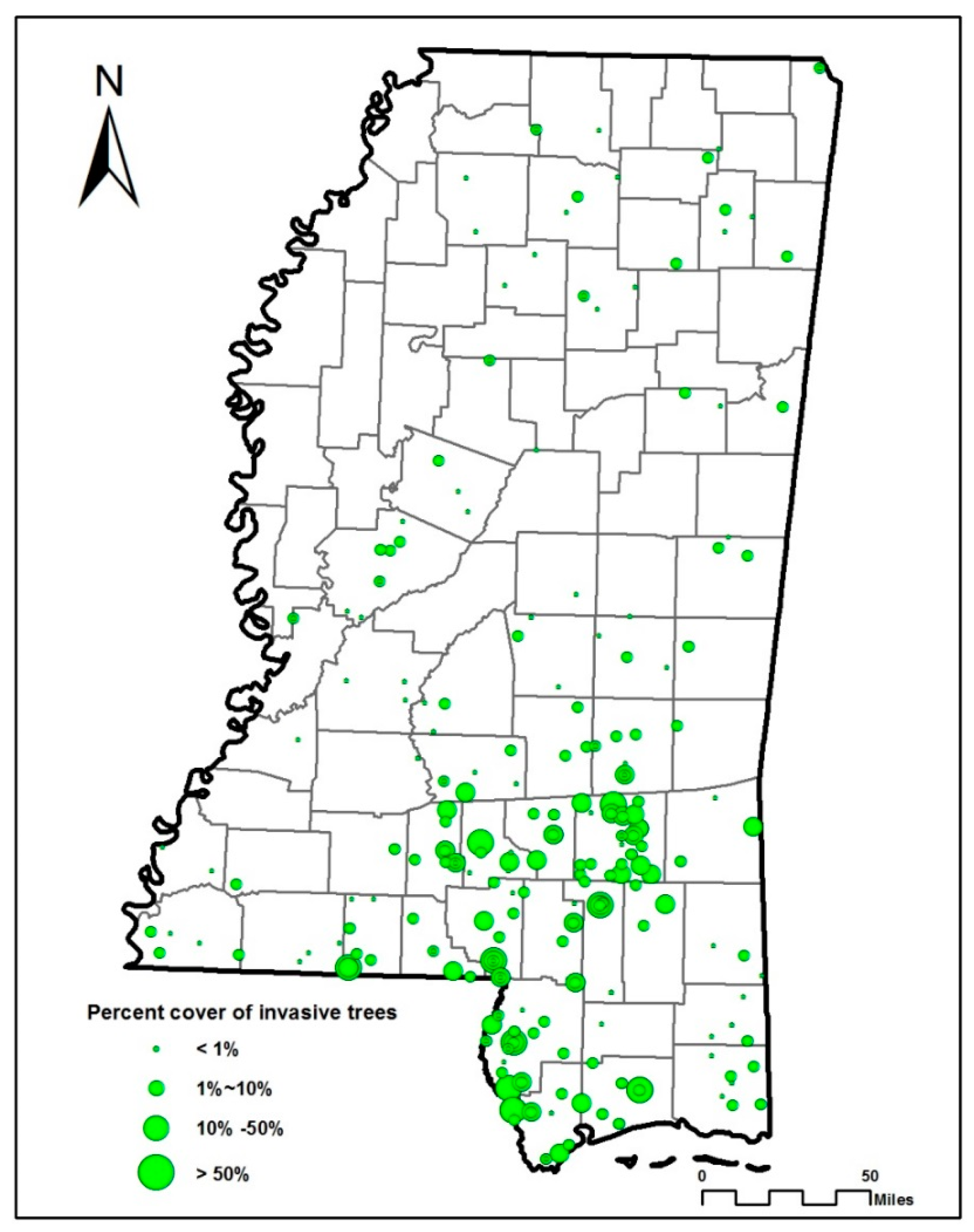
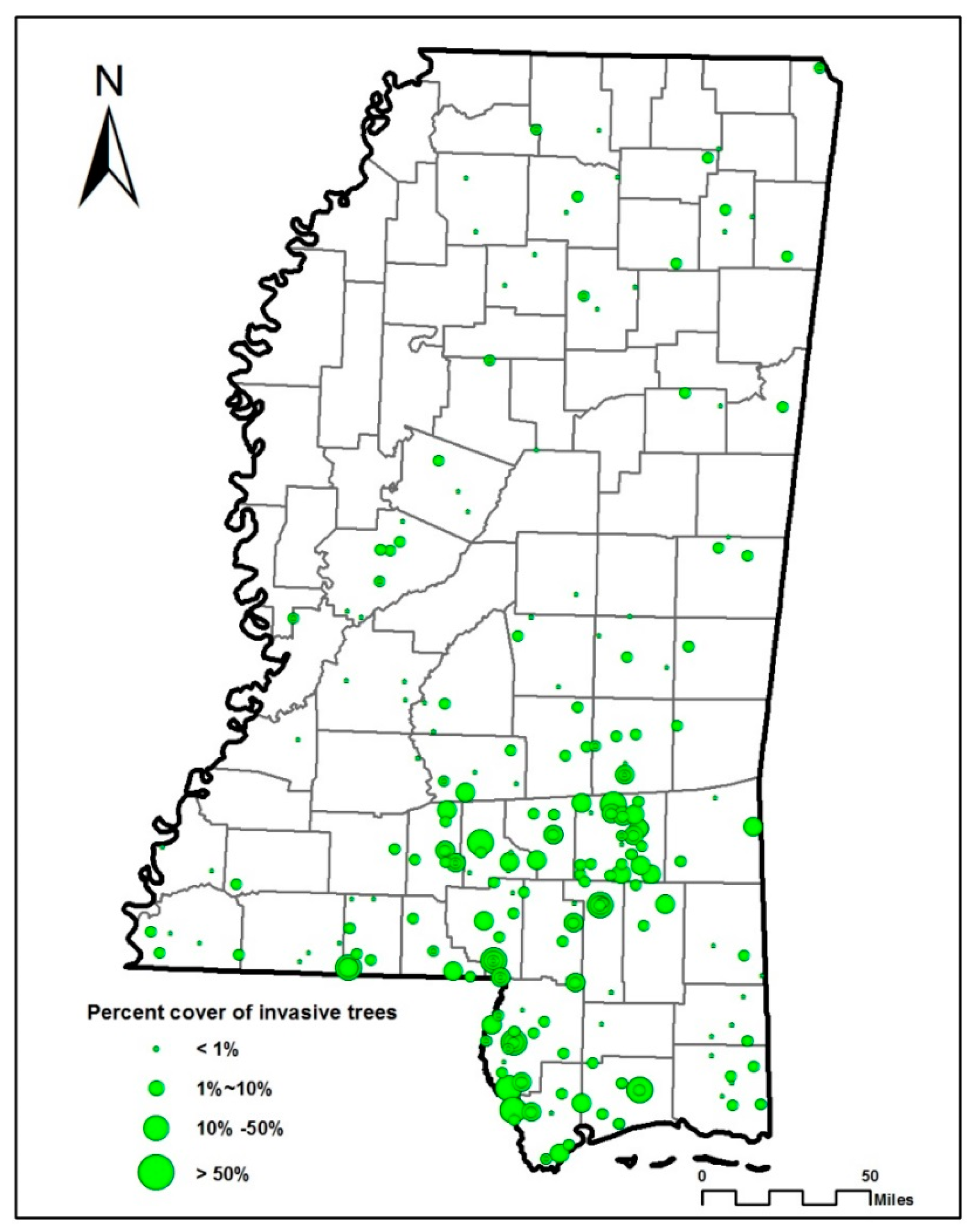
3. Results
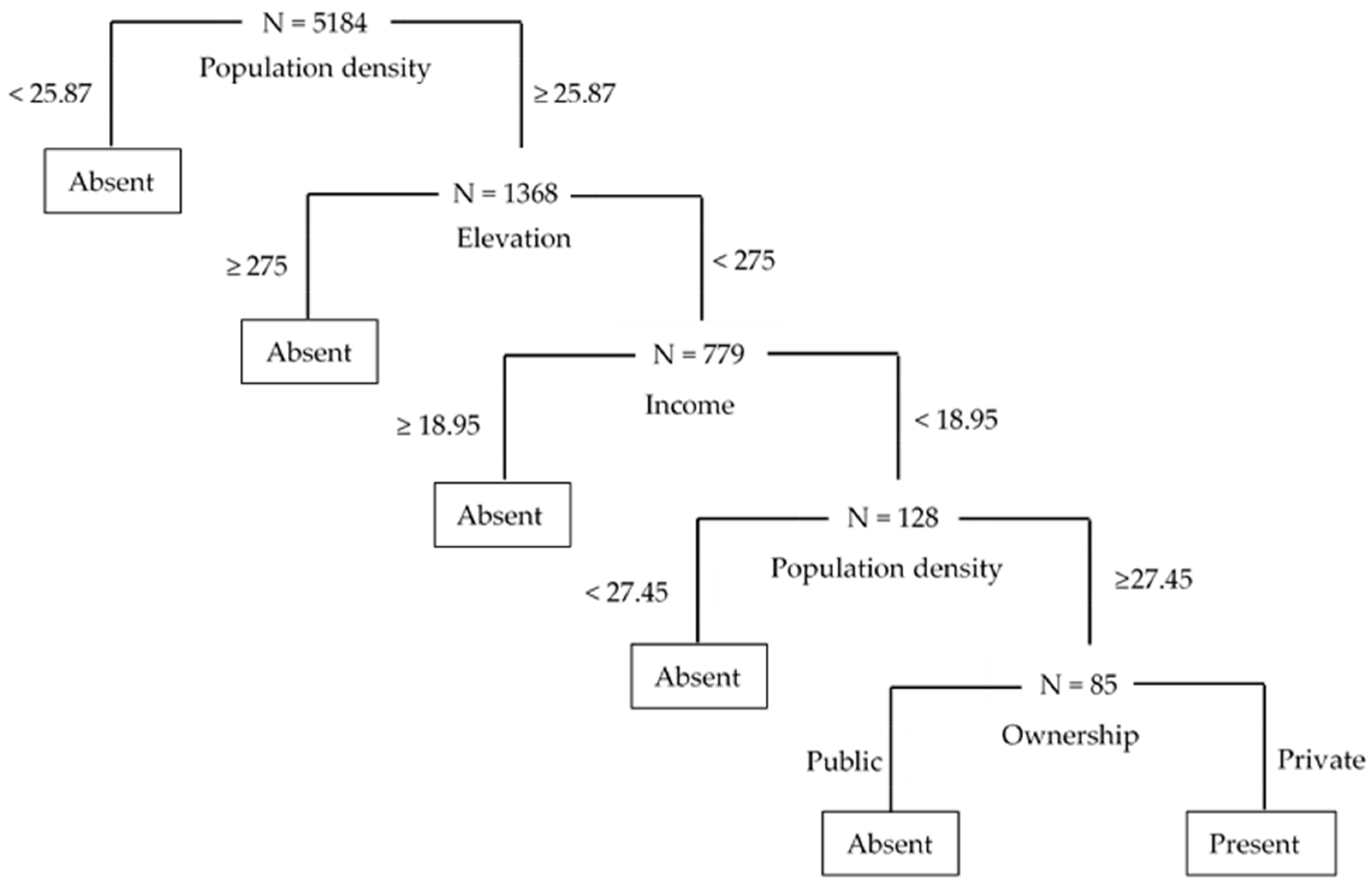
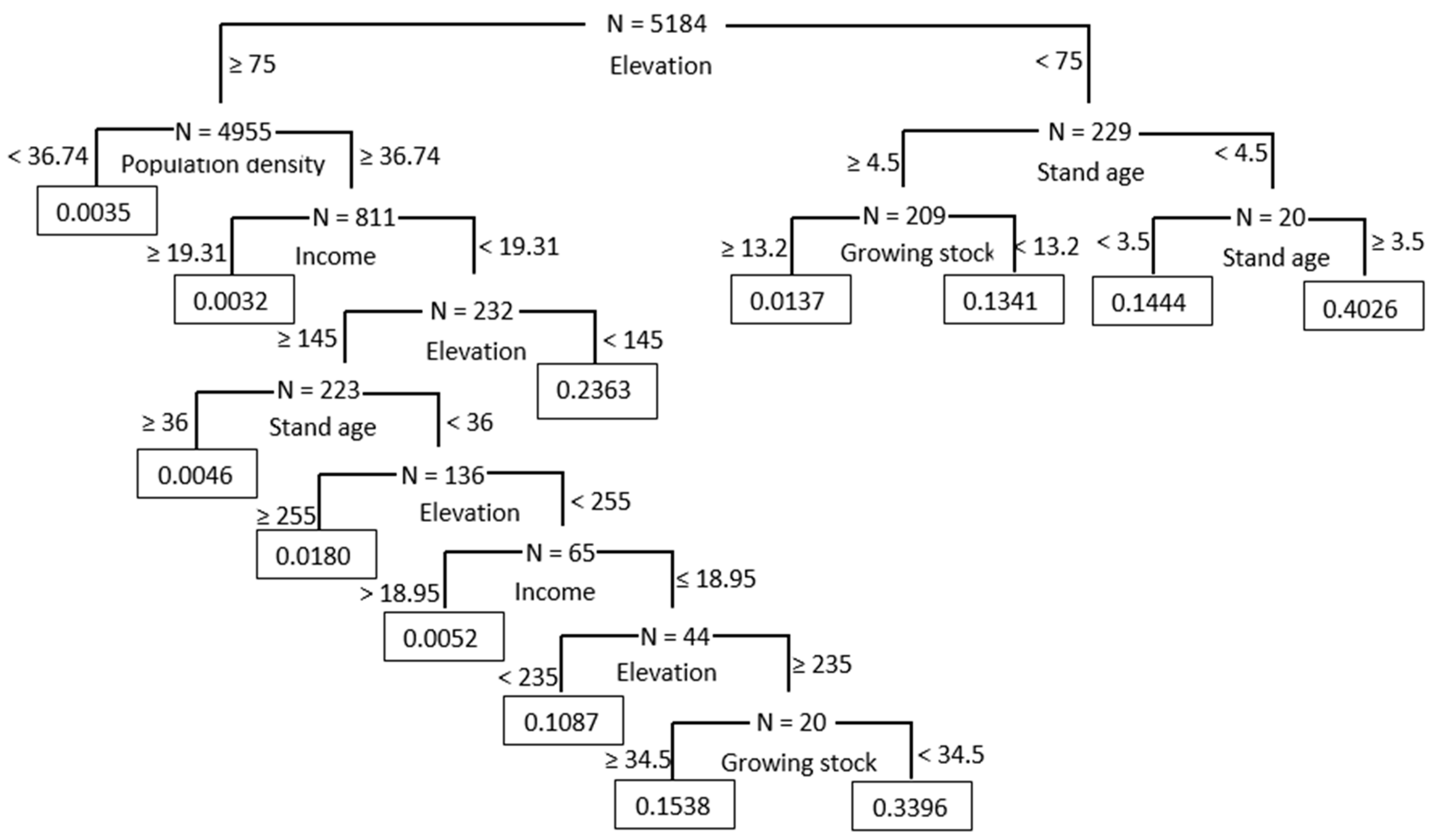
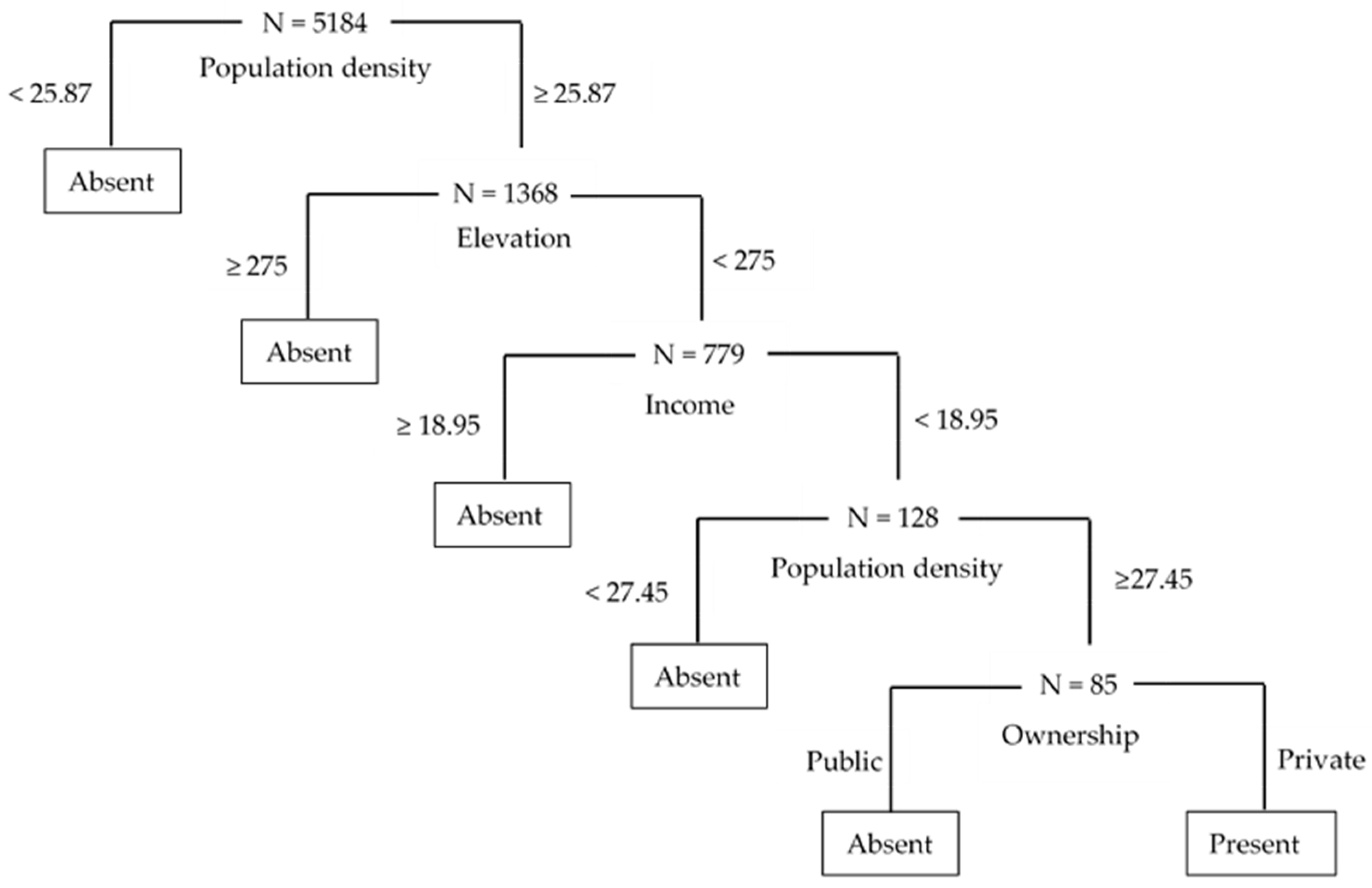
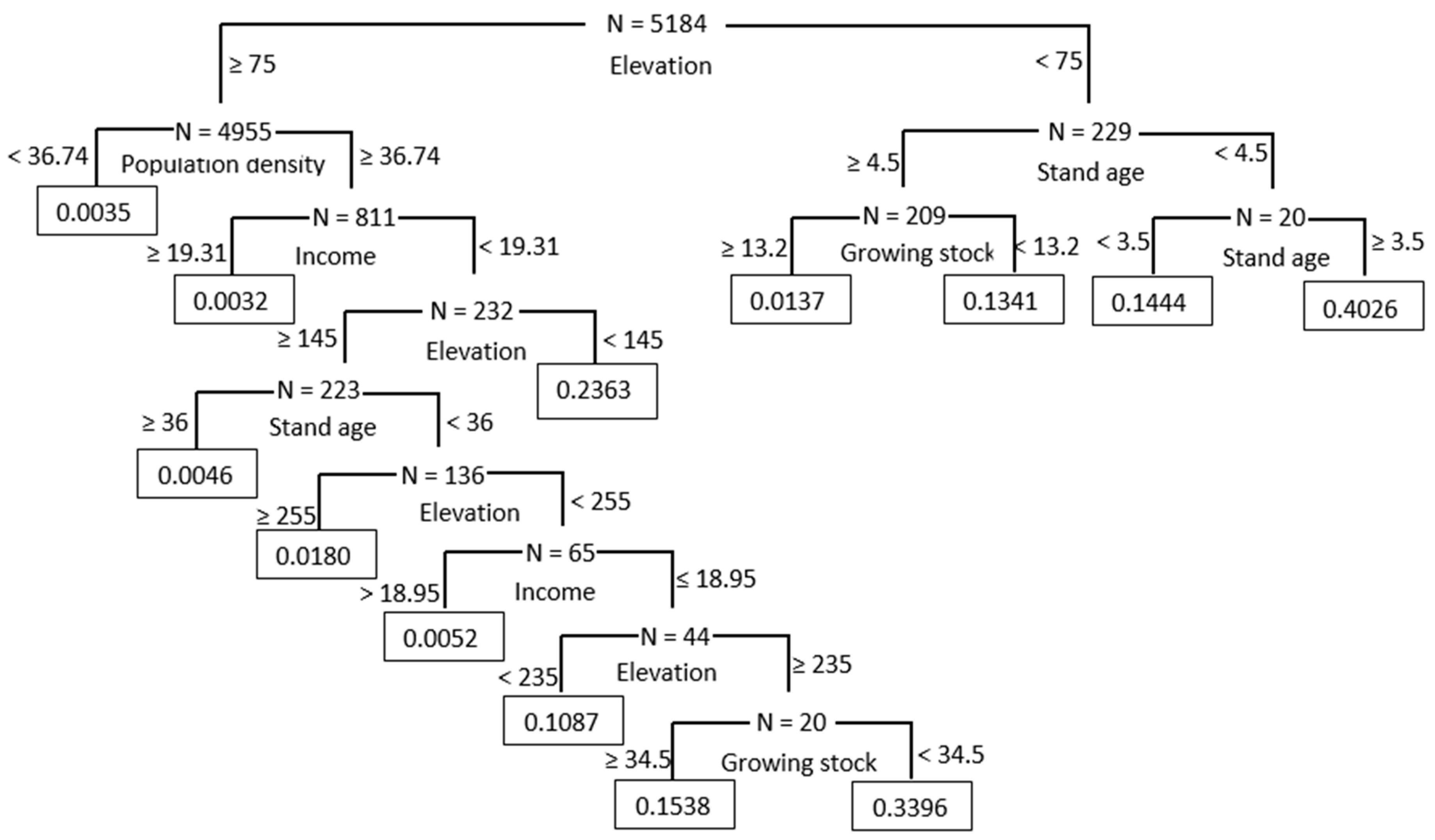
3.1. Results from CART Analysis
3.2. Results from the Spatial Lag Model
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Novak, S.J.; Mack, R.N. Tracing plant introduction and spread: Genetic evidence from Bromus tectorum (cheatgrass). BioScience 2001, 51, 114–122. [Google Scholar] [CrossRef]
- Pimentel, D.; Zuniga, R.; Morrison, D. Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol. Econ. 2005, 52, 273–288. [Google Scholar] [CrossRef]
- Gavier-Pizarro, G.I.; Radeloff, V.C.; Stewart, S.I.; Huebner, C.D.; Keuler, N.S. Housing is positively associated with invasive exotic plant species richness in New England, USA. Ecol. Appl. 2010, 20, 1913–1925. [Google Scholar] [CrossRef] [PubMed]
- Mississippi State University Division of Agriculture, Forestry, and Veterinary Medicine. Mississippi Agriculture, Forestry and Natural Resources; Mississippi State University: Starkville, MS, USA, 2016; p. 7. [Google Scholar]
- Ridley, T.; Oswalt, C.M. Southern Nonnative Invasive Plant data Extraction Tool; version 4.0; U.S. Department of Agriculture, Forest Service, Southern Research Station, Forest Inventory and Analysis Program: Knoxville, TN, USA. Available online: http://srsfia2.fs.fed.us/nonnative_invasive/Southern_Nonnative_Invasives.htm (accessed on 7 March 2017).
- Miller, J.H. Nonnative Invasive Plants of Southern Forests: A Field Guide for Identification and Control; General Technical Reports SRS-62; U.S. Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2003; 93p.
- Dark, S.J. The biogeography of invasive alien plants in California: An application of GIS and spatial regression analysis. Divers. Distrib. 2004, 10, 1–9. [Google Scholar] [CrossRef]
- Gan, J.; Miller, J.H.; Wang, H.; Taylor, J.W. Invasion of tallow tree into Southern US forests: Influencing factors and implications for mitigation. Can. J. For. Res. 2009, 39, 1346–1356. [Google Scholar] [CrossRef]
- Flory, S.L.; Clay, K. Invasive shrub distribution varies with distance to roads and stand age in eastern deciduous forests in Indiana, USA. Plant Ecol. 2006, 184, 131–141. [Google Scholar] [CrossRef]
- Levine, M.T.; Feller, I.C. Effects of forest age and disturbance on population persistence in the understory herb, Arisaema triphyllum (araceae). Plant Ecol. 2004, 172, 73–82. [Google Scholar] [CrossRef]
- Leuschner, C. Forest succession and water resources: Soil hydrology and ecosystem water turnover in early, mid and late stages of a 300-yr-long chronosequence on sandy soil. In Forest Development; Dohrenbusch, A., Bartsch, N., Eds.; Springer: Berlin, Germany, 2002; pp. 1–68. [Google Scholar]
- Brothers, T.S.; Spingarn, A. Forest fragmentation and alien plant invasion of central Indiana old-growth forests. Conserv. Biol. 1992, 6, 91–100. [Google Scholar] [CrossRef]
- Hussain, A.; Sun, C.; Zhou, X.; Munn, I.A. Ecological and economic determinants of invasive tree species on Alabama forestland. For. Sci. 2008, 54, 339–348. [Google Scholar]
- Fan, Z.; Tan, Y.; Crosby, M.K. Factors associated with the spread of Chinese tallow in east Texas forestlands. Open J. Ecol. 2012, 2, 121–130. [Google Scholar] [CrossRef]
- Bellingham, P.J.; Tanner, E.V.; Healey, J.R. Hurricane disturbance accelerates invasion by the alien tree Pittosporum undulatum in Jamaican montane rain forests. J. Veg. Sci. 2005, 16, 675–684. [Google Scholar] [CrossRef]
- Tan, Y. Predicting the Potential Distributions of Major Invasive Species Using Geospatial Models in Southern Forest Lands; Ph.D. Dissertation, Mississippi State University: Starkville, MS, USA, 2011. [Google Scholar]
- Parendes, L.A.; Jones, J.A. Role of light availability and dispersal in exotic plant invasion along roads and streams in the HJ Andrews Experimental forest, Oregon. Conserv. Biol. 2000, 14, 64–75. [Google Scholar] [CrossRef]
- Watkins, R.Z.; Chen, J.; Pickens, J.; Brosofske, K.D. Effects of forest roads on understory plants in a managed hardwood landscape. Conserv. Biol. 2003, 17, 411–419. [Google Scholar] [CrossRef]
- Gelbard, J.L.; Harrison, S. Roadless habitats as refuges for native grasslands: Interactions with soil, aspect, and grazing. Ecol. Appl. 2003, 13, 404–415. [Google Scholar] [CrossRef]
- Smith, W.B.; Miles, P.D.; Vissage, J.S.; Pugh, S.A. Forest Resources of the United States, 2002; USDA Forest Service General Technical Reports; United States Department of Agriculture: Washington, DC, USA, 2004; p. NC-241.
- Colton, T.F.; Alpert, P. Lack of public awareness of biological invasions by plants. Nat. Area J. 1998, 18, 262–266. [Google Scholar]
- McKinney, M.L. Do human activities raise species richness? Contrasting patterns in United States plants and fishes. Glob. Ecol. Biogeogr. 2002, 11, 343–348. [Google Scholar] [CrossRef]
- McKinney, M.L. Measuring floristic homogenization by non-native plants in North America. Glob. Ecol. Biogeogr. 2004, 13, 47–53. [Google Scholar] [CrossRef]
- Qian, H.; Ricklefs, R.E. The role of exotic species in homogenizing the North American flora. Ecol. Lett. 2006, 9, 1293–1298. [Google Scholar] [CrossRef] [PubMed]
- Hope, D.; Gries, C.; Zhu, W.; Fagan, W.F.; Redman, C.L.; Grimm, N.B.; Nelson, A.L.; Martin, C.; Kinzig, A. Socioeconomics drive urban plant diversity. Proc. Natl. Acad. Sci. USA 2003, 100, 8788–8792. [Google Scholar] [CrossRef] [PubMed]
- Stewart, S.I.; Radeloff, V.C.; Hammer, R.B. Characteristics and location of the wildland-urban interface in the United States. In Proceedings of the 2nd International Wildland Wildfire Ecology and Wildfire Management Congress, Orlando, FL, USA, 16–20 November 2003. [Google Scholar]
- McKinney, M.L. Urbanization, biodiversity, and conservation the impacts of urbanization on native species are poorly studied, but educating a highly urbanized human population about these impacts can greatly improve species conservation in all ecosystems. BioScience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- US Geological Survey. The National Map Small Scale; US Department of the Interior: Washington, DC, USA. Available online: https://nationalmap.gov/small_scale/mld/roadtrl.html (accessed on 7 March 2017).
- US Census Bureau. In Population and Household Estimates; US Department of Commerce: Washington, DC, USA. Available online: www.census.gov (accessed on 7 March 2017).
- Spatial Analysis for Conservation and Sustainability. Wildland Urban Interface (WUI) Project. Forest and Wildlife Ecology Russell Laboratories (Map). Available online: http://silvis.forest.wisc.edu/ (accessed on 7 March 2017).
- Yohannes, Y.; Hoddinott, J. Classification and Regression Trees: An Introduction; International Food Policy Research Institute: Washington, DC, USA, 1999. [Google Scholar]
- Lewis, R.J. An introduction to classification and regression tree (cart) analysis. In Proceedings of the Annual Meeting of the Society for Academic Emergency Medicine, San Francisco, CA, USA, 22–25 May 2000; pp. 1–14. [Google Scholar]
- Fan, Z.; Kabrick, J.M.; Shifley, S.R. Classification and regression tree based survival analysis in oak-dominated forests of Missouri’s Ozark highlands. Can. J. For. Res. 2006, 36, 1740–1748. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria; Available online: http://www.R-project.org (accessed on 7 March 2017).
- Anselin, L. Spacestat Tutorial: A Workbook for Using Spacestat in the Analysis of Spatial Data; Center for Geographic Information and Analysis: Santa Barbara, CA, USA, 1992. [Google Scholar]
- Sax, D.F. Native and naturalized plant diversity are positively correlated in scrub communities of California and Chile. Divers. Distrib. 2002, 8, 193–210. [Google Scholar] [CrossRef]
- Rejmánek, M.; Randall, J.M. Invasive alien plants in California: 1993 summary and comparison with other areas in North America. Madrono 1994, 41, 161–177. [Google Scholar]
- Inouye, R.S.; Huntly, N.J.; Tilman, D.; Tester, J.R.; Stillwell, M.; Zinnel, K.C. Old-field succession on a Minnesota Sand Plain. Ecology 1987, 68, 12–26. [Google Scholar] [CrossRef]
- Norcini, J.; Aldrich, J. Native wildflowers: Mimosa strigillosa Torr. and A. Gray; Florida Cooperative Extension Service Publication ENH: Gainesville, FL, USA, 2009; p. 1075. [Google Scholar]
- Grime, J.P. Shade avoidance and shade tolerance in flowering plants. Symp. Br. Ecol. Soc. 1966, 7, 187–207. [Google Scholar]
- Longbrake, A.C.W. Ecology and Invasive Potential of Paulownia tomentosa (Scrulariaceae) in a Hardwood Forest Landscape; Ohio University: Athens, OH, USA, 2001. [Google Scholar]
- Williams, C.E. The exotic empress tree, Paulownia tomentosa: An invasive pest of forests? Nat. Areas J. 1993, 13, 221–222. [Google Scholar]
- Lin, J.; Harcombe, P.A.; Fulton, M.R.; Hall, R.W. Sapling growth and survivorship as affected by light and flooding in a river floodplain forest of southeast Texas. Oecologia 2004, 139, 399–407. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| No. | Common Name | Scientific Name | Year Introduced to the U.S./Native Range |
|---|---|---|---|
| 1 | Tree of heaven | Ailanthus altissima | 1784/northeast and central China |
| 2 | Silktree/Mimosa | Albizia julibrissin | 1745/Iran to Korea |
| 3 | Princesstree | Paulownia tomentosa | 1900s/central and western China |
| 4 | Chinaberry | Melia azedarach | 1830s/Indomalaya and Australasia |
| 5 | Chinese tallow | Triadica sebifera | 1970s/eastern Asia |
| Category | Classification | Definition |
|---|---|---|
| Ecological factors | Elevation | The average elevation of all plots in county i |
| Successional age | Average age of all plots in county i | |
| Forest type | ||
| loblolly pine | The number of plots with loblolly pine in county i/total number of plots in county i | |
| white oak/red oak/hickory | The number of plots with white oak/red oak/hickory in county i/total number of plots in county i | |
| loblolly pine/hardwood | The number of plots with loblolly pine/hardwood in county i/total number of plots in county i | |
| Natural disturbance | ||
| ground fire | The number of plots with ground fire records in county i/total number of plots in county i | |
| hurricane, tornado | The number of plots with hurricane or tornado records in county i/total number of plots in county i | |
| Site productivity | Average site productivity of all plots in county i | |
| Growing stock | Average growing stock of all plots in county i | |
| Stand origin (artificial) | The number of plots with artificial origin in count i/total number of plots in county i | |
| Socio-economic factors | Road density | The length of major roads/the area of county i |
| Ownership (private) | The number of plots with private ownership in county i/total number of plots in county i | |
| Population density | Number of inhabitants per county divided by area of county i | |
| Per capita mean annual income | Per capita mean annual income by county (US$) | |
| Wildland urban interface | Area of wildland urban interface/area of county i (km2/km2) | |
| Factors | Coefficient | SE | OR | p Value |
|---|---|---|---|---|
| Ecological variables | ||||
| Elevation | −0.0001 | 0.00006 | 0.9999 | 0.0310 ** |
| Socioeconomic variables | ||||
| Ownership (public) | −0.0645 | 0.0370 | 0.9375 | 0.0810 * |
| Population density | 0.0000 | 0.0003 | 1.0000 | 0.9967 |
| Per capita mean annual income | 0.6700 | 0.3264 | 1.9542 | 0.0401 ** |
| Factors | Coefficient | SE | Z Value | p Value |
|---|---|---|---|---|
| Ecological variables | ||||
| Stand age | −1.5629 | 0.6470 | −2.4156 | 0.0157 ** |
| Elevation | −0.0962 | 0.0457 | −2.1054 | 0.0353 ** |
| Growing stock | 0.0248 | 0.6438 | 0.0385 | 0.9693 |
| Socioeconomic variables | ||||
| Population density | −0.0014 | 0.2008 | −0.0069 | 0.9945 |
| Per capita mean annual income | 294.5935 | 217.1067 | 1.3569 | 0.17481 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhai, J.; Grebner, D.L.; Grala, R.K.; Fan, Z.; Munn, I.A. Contribution of Ecological and Socioeconomic Factors to the Presence and Abundance of Invasive Tree Species in Mississippi, USA. Forests 2018, 9, 38. https://doi.org/10.3390/f9010038
Zhai J, Grebner DL, Grala RK, Fan Z, Munn IA. Contribution of Ecological and Socioeconomic Factors to the Presence and Abundance of Invasive Tree Species in Mississippi, USA. Forests. 2018; 9(1):38. https://doi.org/10.3390/f9010038
Chicago/Turabian StyleZhai, Jun, Donald L. Grebner, Robert K. Grala, Zhaofei Fan, and Ian A. Munn. 2018. "Contribution of Ecological and Socioeconomic Factors to the Presence and Abundance of Invasive Tree Species in Mississippi, USA" Forests 9, no. 1: 38. https://doi.org/10.3390/f9010038
APA StyleZhai, J., Grebner, D. L., Grala, R. K., Fan, Z., & Munn, I. A. (2018). Contribution of Ecological and Socioeconomic Factors to the Presence and Abundance of Invasive Tree Species in Mississippi, USA. Forests, 9(1), 38. https://doi.org/10.3390/f9010038