Abstract
Some studies indicate that metal nanoparticles can be used in plant cultivation as fungicides and growth stimulators. The aim of this study was to evaluate the effect of silver (AgNPs) and copper nanoparticles (CuNPs) on the growth parameters, on the extent of leaves infected by powdery mildew and on spontaneous ectomycorrhizal colonization of English oak (Quercus robur L.) seedlings growing in containers. Nanoparticles were applied to foliage four times during one vegetation season, at four concentrations: 0, 5, 25 and 50 ppm. The adsorption of NPs to leaves was observed by microscopical imaging (TEM). The tested concentrations of AgNPs and CuNPs did not have any significant effect on the growth parameters of the oak seedlings. TEM results showed disturbances in the shape of plastids, plastoglobules and the starch content of oak leaves treated with 50 ppm Cu- and AgNPs, while no changes in the ultrastructure of stems and roots of oak plants treated with NPs were observed. No significant difference in powdery mildew disease intensity was observed after NP foliar app lication. Four ectomycorrhizal taxa were detected on oak roots (Sphaerosporella brunnea, Thelephora terrestris, Paxillus involutus and Laccaria proxima). Oak seedlings treated (foliar) with CuNPs and AgNPs at 25 ppm were characterised by the highest degree of mycorrhization (respectively, 37.1% and 37.5%) among all treatments including the control treatment. None of the tested NPs manifested phytotoxicity in the examined Q. robur seedlings under container nursery conditions.
1. Introduction
Nanoparticles (nano-scale particles, NP) are atomic or molecular aggregates with dimensions between 1 nm and 100 nm [1,2]. Unique properties of NPs include: very large specific surface area, high surface energy and quantum confinement [3], which creates new opportunities to use NPs as a novel element of the plant protection toolbox. Currently, within the Integrated Pest Management (IPM) program in forest nursery production in Poland, trials are carried out to use non-conventional plant protection methods [4]. In integrated plant protection, physical and biological methods have priority over chemical preparations. Alternative methods are being sought, particularly in forestry, which may offer phytosanitary protection and growth improvement with minimal impact on the environment.
The potential application of nanotechnology in forestry requires research- and evidence-based risk assessment. Some authors [5] suggest that more information on the fate of NPs in water, soil and food webs is needed. Plants as primary producers with huge absorbing surfaces may play a key role as a potential gateway for NPs into the food webs. Moreover, safety assessment of NPs influence on various functional guilds of (micro)organisms, such as decomposers, plant endophytes and mycorrhizal fungi should be taken into account. Nanoparticles can affect plant growth directly or indirectly, through effects on the plants’ microbial associates. Growth effects are often considered as the result of the antimicrobial properties of nanoparticles [6]. The question as to which extent mutualistic symbionts such as ectomycorrhizal fungi are affected by the application of nanoparticles is still a research gap.
Various studies indicate negative, positive or neutral effects of engineered metal nanoparticles on growth and seed germination of higher plants. Obtained results depend on the properties and concentration of nanoparticles and on treated plant species [7]. For example, Lin et al. [8] experimentally demonstrated that nanostructured silicon dioxide can be used as a plant growth stimulator of coniferous seedlings. On the other hand, Lin and Xing [9] showed an inhibition of seed germination and root growth by certain NPs in higher plant species. It seems that effects are specific to the type of nanoparticles applied, their concentrations, as well as to the plants tested (Table 1). Ma et al. [10] argued that the surface area of NPs is a more appropriate estimator of biocidal effects and phytotoxicity than nominal concentration of NPs. Qi et al. [11] showed that copper-carbon core-shell nanoparticles can enhance Cu uptake in 2-year-old bald cypress (Taxodium distichum) seedlings but did not observe significant effects on the growth parameters of the seedlings after 25 weeks of NPs treatment. On the other hand, Lee et al. [12] reported toxicity of copper nanoparticles to two crop species, mung bean (Phaseolus radiatus) and wheat (Triticum aestivum), as demonstrated by the reduced seedling growth rate.

Table 1.
Summary of selected studies on the influence of nanoparticles on woody and non-woody plants.
Management of fungal diseases in plant nurseries is economically important and of environmental concern, to avoid the introduction and spread of diseases. Diseases such as powdery mildew, needle casts and damping-off are among the most destructive foliar and soil-borne infections of forest tree seedlings [21]. It has been shown that nano-silver can cause significant reduction in seedling infection by Fusarium culmorum, an agent of damping-off [22]. However, an analysis of antioxidant enzyme activity characteristic of stress response indicated that the toxicity of nano-silver treatment is comparable to damage caused by Fusarium treatment [23]. Qi et al. [11] demonstrated that wood treated with copper-carbon core-shell nanoparticles is highly resistant to blue stain (Ophiostoma minus) and white rot (Trametes versicolor) fungi, and Formosa subterranean termite (Coptotermes formosanus). Some attempts were made to use AgNPs for controlling powdery mildew diseases [24,25].
According to available literature, NPs can be used to improve the phytosanitary status of seedlings in nursery conditions; however, multiple potential mechanisms affecting plant growth can be expected: direct (plant toxicity, plant nutrition and hormone-like effects) and indirect (suppression on plant pathogens or effects on mutualistic symbionts), as both AgNPs and CuNPs exhibit antifungal, antibacterial, and antiviral properties [7].
Ectomycorrhizal (ECM) symbiosis is a type of mutualistic symbiosis which concerns about 3% of land plant species, while arbuscular mycorrhizal fungi (AMF) are associated with over 90%. Despite the limited number of host plant species, the global importance of ECM plant species is greatly increased by their disproportionate occupancy of the terrestrial land surface and their economic value as major producers of timber. Fagaceae as the dominants or co-dominants of temperate and mediterranean forests are one example of economically important forest tree families with obligate ECM symbiosis [26]. In ECM tree species, the formation of ectomycorrhizae is the natural state, and ECM symbiosis can be considered an extension of their root systems and a factor to take into account when assessing their quality [27]. ECM fungi are beneficial for plant growth because they can actively release nutrients from mineral particles and rock surfaces via weathering [28,29], mobilising nutrients such as N and P from structural and other organic polymers which are otherwise unavailable to plant roots [30]. In addition, ECM fungi play a role in carbon flow [31] and mediate plant responses to metal toxicity, oxidative stress, water stress, and effects of soil acidification [32,33] and plant pathogens [26] as well as possible interactions with groups of other soil microorganisms [34]. Moreover, some authors suggested that differences in above-ground tree vitality are often reflected below-ground, in particular, in the vitality and the mycorrhizal status of the root systems of trees [35,36,37]. Currently available information about the effects of NPs on mycorrhizal symbioses is quite limited. Manceau et al. [38] showed that the common wetlands plants Phragmites australis and Iris pseudoacorus can transform copper into metallic nanoparticles with evidence of assistance by endomycorrhizal fungi, thus limiting Cu toxicity. Some studies have highlighted the negative effect of nanoparticles on mycorrhizal colonization [39,40]. The investigation of effects of silver sulphide nanomaterials on arbuscular mycorrhizal fungal (AMF) colonization of tomato (Solanum lycopersicum), and overall microbial community structure in biosolids-amended soil showed that bacteria, actinomycetes and fungi were inhibited by NP treatments. For example, it was shown that silver sulfide nanomaterials were toxic to mycorrhizal symbiosis, resulting in reduced plant biomass and greatly reduced mycorrhizal colonization [40]. Sweet and Singleton [13] showed that soil contamination with AgNPs reduced root and shoot growth of treated Bishop pine (Pinus muricata) and ectomycorrhizal diversity on pine roots.
Currently, no data on the potential growth stimulating effects of NPs on ECM woody plants in forest nurseries are available. Moreover the mechanisms of how metal NPs may affect tree growth are still not elucidated. Therefore, the aims of this study were to (i) determine the effects of foliar applications of AgNPs and CuNPs on the growth of English oak (Quercus robur L.) seedlings in experimental nursery container production; (ii) determine the effect of NPs on powdery mildew disease; (iii) compare the ultrastructure of leaves of tested plants by transmission electron microscopy and (iv) determine the state and degree of mycorrhizal colonization. We tested the hypothesis that the impact of AgNPs and CuNPs on examined ECM woody plants after foliage application will result in changes in leaf ultrastructure and in effects on powdery mildew disease intensity and on mycorrhizal colonisation.
2. Materials and Methods
2.1. Study Design, Plant Material, Soil Substrate and Sampling Scheme
The study was conducted in the nursery of the Forest Experimental Station (Rogów, 51°40′ N, 19°55′ E, 195 m a.s.l) of Warsaw University of Life Sciences, Poland, in a foil-covered greenhouse with the following dimensions: height 2.8 m, width 6 m and length 30 m. English oak seedlings were grown in plastic container trays V-360 (15 pots per tray with a capacity of 360 cm3) in peat—perlite substrate, purchased from the container nursery in Nędza, Rudy Raciborskie Forest District, Poland. The substrate (pH = 4.5) comprised sphagnum peat from Estonia −85% and coarse-grained perlite (No. 3) −15%, which is considered to improve the aeration of the substrate [41]. Seedlings were fertilized with a mixture of Osmocote Exact Standard controlled release fertilizers with different release characteristics: 3–4M, 5–6M, 8–9M (1:1:1). Seeds of English oak were of local origin from managed forests. Cotyledons of acorns were cut before sowing, resulting in even emergence of the seedlings and uniformity of growth in the first weeks. On 12 May 2014, two oak acorns were seeded per pot, in order to obtain two seedlings in each pot. After germination, one plant per pot was randomly selected and the others removed. The containers were placed in a foil-covered greenhouse and regularly watered, when necessary. The temperature in the greenhouse during the experiment was 25–32 °C. The seedlings were spontaneously infected by oak powdery mildew (Erysiphe alphitoides); the first symptoms were visible between first and second treatment. There were four spray treatments (between June and August) of the seedlings’ aerial parts using 1000 L/ha (=100 mL/m2) aqueous solutions of NPs (AgNPs, CuNPs) at 4 different concentrations: 0 ppm, 5 ppm, 25 ppm, 50 ppm. Previously, the NPs were suspended in deionized water by vigorously shaking for at least 10 min. Nanoparticles were applied to foliage four times: 11, 25 June, 9 July, 6 August 2014. The first treatment was done one month after sowing. The seedlings had formed two leaves that did not yet manifest powdery mildew disease. This is the optimal time for the first protective treatment. Conditions of all the treatments were similar. Sprayings were applied in the early morning with a manual compressed air sprayer (Kisan Kraft, model KK-PS5000). The greenhouse foil protected the oak seedlings from rain, which might have affected the effectiveness of the treatment. The experimental design comprised two factors with four levels each (AgNPs 0 ppm, AgNPs 5 ppm, AgNPs 25 ppm, AgNPs 50 pmm, CuNps 0 ppm, CuNPs 5 ppm, CuNPs 25 ppm and CuNPs 50 ppm). The experiment was performed in a randomised complete block design. At the end of the growing season (October), 40 seedlings (10 plants × 4 blocs) were randomly harvested from each treatment. In total, 280 English oak seedlings were sampled.
2.2. Characteristics of Nano-Silver and Nano-Copper Colloids
In this research project, two frequently studied types of metal nanoparticles (MNPs) were chosen, silver (AgNPs) and copper (CuNPs). Samples of commercially available solutions of silver (Figure 1) and copper nanoparticles were obtained from Nano-Koloid (Nano-Koloid sp. z o.o.) licensee of Nano Technologies Group, Inc. (Naperville, IL, USA), manufactured according to the European patent EP2081672 A2. As the producer noted, these are generated in a physical process, consist of around 100 atoms and are suspended in demineralised water. The concentration of nanoparticles in the commercially available product is 50 ppm.
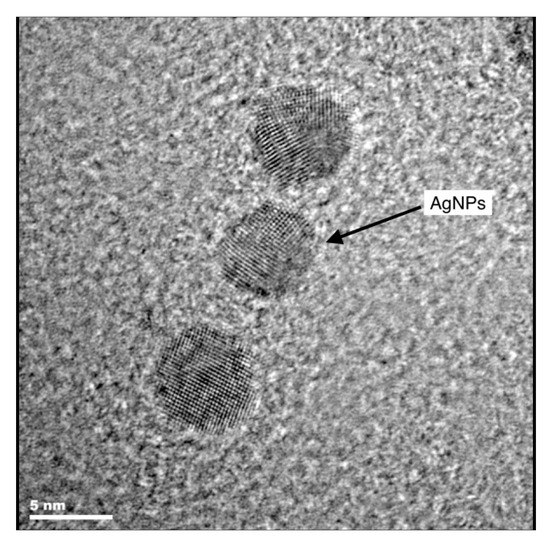
Figure 1.
Transmission electron micrograph of silver nanoparticles employed in the present study [42].
2.3. Biometric Parameters and Powdery Mildew Assessment
The following shoot growth parameters were measured: shoot length (cm) and root collar diameter (mm). To determine the total root length (cm) and the number of root tips (n), the root systems were analysed using an Epson Perfection scanner 4990 Photo integrated with WinRhizo software (Regent Instruments Inc., Quebec City, QC, Canada). At the end of the laboratory work, the dry mass of the shoot and root (g) was determined after drying at 105 °C for 24 h.
The degree of powdery mildew infection of all oak seedlings was assessed based on the fraction of affected leaf area according to the following scale: 0—no visible infection; 1—25% of the surface infected; 2—26–50%; 3—51–75%; 4—>75%. The infestation index (I) was calculated using the following formula:
I = 0 × N0 + 1 × N1 + 2 × N2 + 3 × N3 + 4 × N4/N0+1+2+3+4
I—powdery mildew infestation index; 0, 1, 2, 3, 4—scale of infection of individual leaves; Nx—number of infested leaves at a specific scale.
2.4. Microscopic Investigations
For transmission electron microscopic imaging (TEM), ten seedlings of each treatment were randomly chosen. Leaves (not diseased and diseased by oak powdery mildew) from each treatment (two per seedling) were used for further investigations. The adsorption and assimilation of NPs to leaves, shoots and roots of tested oak plants was observed by TEM 8 weeks after the last treatment. Three samples (from the epidermis and mesophyll) from one leaf were used for TEM. Tissue samples were collected from the middle part of leaves, shoots and roots of oak seedlings. The obtained plant material was fixed in 2% (v/v) glutaraladehyde and 2% paraformaldehyde (v/w) in 0.05 M cacodylate buffer (pH 7.2) according to Karnovsky [43] for 4 h at room temperature. Then, the material was rinsed four times with cacodylate buffer. The samples were contrasted in 1% OsO4 for 2 h at 4 °C and rinsed four times with the same buffer. Next, the material was dehydrated in a series of aqueous solutions of ethanol and subsequently in propylene oxide. Finally, the material was gradually saturated with resin (Epon, Fluka) and polymerised for 24 h at 60 °C. Epon blocks with plant material were cut for TEM, with an ultramicrotome (Leica) into ultrathin sections (~90 nm thick), which were collected on Formvar coated slot-grids and contrasted with 1% lead citrate and 2% uranyl acetate. Thereafter, the material was examined using a Morgagni 268D (FEI) electron microscope. Photographic documentation was prepared with a Morada (SIS) digital camera and the iTEM (SIS) computer programme.
2.5. Ectomycorrhizae Assessment
The roots of harvested seedlings were analysed under a stereoscopic microscope (Delta IPOS-808, Delta Optical, Warsaw, Poland) coupled to a digital camera at 10–40× magnification. The degree of mycorrhization of seedlings was determined by classifying and counting the vital mycorrhizal (VM: turgid ECM tips, with ECM mantle), non-mycorrhizal (NM: well-developed, turgid fine root tips, mantle lacking) and non-vital (NV: non-turgid, with a scurfy surface and an easily detachable cortex, with or without the remnants of an ECM mantle) root tips. The proportion of vital mycorrhizal (VM) root tips was calculated as: (mycorrhizal vital root tips)/(mycorrhizal root tips + non-mycorrhizal root tips (NM) and non-vital (NV)) × 100 and presented as degree of mycorrhization. The proportions of NM and NV were expressed in percentages in the same way. The relative abundance of individual ectomycorrhizal fungal taxa was calculated as the number of ectomycorrhizal root tips of each morphotypes/taxon divided by the total number of ectomycorrhizal roots tips. The frequency of ectomycorrhizal taxa was expressed as the percentage of colonized plants. Ectomycorrhizal tips were identified under a dissecting microscope by the absence of root hairs, shape (hypertrophy) and colour of fine roots, the presence of mycelial mantles and emanating fungal structures (hyphae and rhizomorphs). The initial identification of morphotypes was based on available literature [44,45]. Representative mycorrhizal root tips of each morphotype were photographed and deposited together with the fungal description and molecular information in an internal database. To determinate the mycorrhizal species/taxa, we collected tip material from three to five mycorrhizal root tips per morphotype and transferred them into Eppendorf tubes filled with 70% EtOH for molecular analysis.
Identification of mycorrhizal fungi included PCR amplification of selected regions of internal transcribed spacer (ITS) rDNA using the primer pair ITS1F/ITS4 [46,47] and sequencing of the resulting PCR product. We used direct PCR amplification of fungal DNA from ECM samples, bypassing conventional DNA extraction procedures [48]. PCR products were assessed by loading 2 μL onto a 1% agarose gel (0.5 × TAE buffer) and visualized under UV light, using the GeneRulerTM 1 kb Plus DNA Ladder (Fermentas, Thermo Fisher Scientific, Waltham, MA, USA). Bi-directional Dye-Terminator Cycle Sequencing was performed using BigDye Terminator v3.1 Chemistry (Applied Biosystems, Thermo Fisher Scientific, Waltham, MA, USA) and one of the PCR primers each. The resulting fragments were analysed on a 3730 DNA Analyser (Applied Biosystems, Thermo Fisher Scientific, Waltham, MA, USA) at the Department of Botany and Biodiversity Research in University of Vienna. Identification of sequenced fungi was based on the results of BLAST [49] searches against the National Centre for Biotechnology Information (NCBI) public database and subsequent phylogenetic placement and on queries against the UNITE database [50].
2.6. Data Analysis
Normality of data distribution within blocks x treatment areas was tested using the Shapiro-Wilk test; homogeneity of variance was assessed with Levene’s test. We tested for multivariate relationships among biometrical parameters and infestation by powdery mildew depending on blocks and treatments (different concentrations of AgNPs and CuNPs) using a permutation multivariate analysis of variance (perMANOVA) with 9999 permutations. This analysis was conducted based on a Bray-Curtis dissimilarity matrix calculated on all observed biometrical parameters and on the powdery mildew infestation index. The statistical analysis was performed using R version 2.15.0 (The R Foundation for Statistical Computing, Vienna, Austria) [51] with the vegan package [52] for multivariate analysis (perMANOVA). Tukey contrasts were calculated with the multcomp package [53]. The accepted level of significance was p < 0.05. Potential effects of the nanoparticle treatments (AgNPs and CuNPs at different concentrations) on the abundance of VM, NM and NV root tips were analysed using Generalized Linear Models (GLM) for a randomized complete block design (with binomial error distribution and logit link function). The GLM model for the proportion of the defined root types (VM, NM and NV) is given by:
where pik is the numerical proportion of specific root tips (VM, NM and NV) for the j-th concentration of nanoparticles nested in the i-th type of metal nanoparticles in the k-th block, θ is the grand mean, ai is the effect of the i-th type of metal nanoparticles, bj(i) is effect of the j-th concentration nested in the i-th type of metal nanoparticles, gk is the random effect of the k-th block and eijk is the random effect of experimental error. Tukey´s contrast was used to test pairwise comparisons (post hoc test) between different concentrations of each type of metal nanoparticle separately.
pijk = θ + ai + bj(i) + gk + eijk
3. Results
3.1. Growth Parameters and Powdery Mildew
Within the range of tested concentrations of NPs, the foliar application of nano-silver and nano-copper did not affect the growth parameters of oak seedlings after the first growing season (perMANOVA, df = 2, p = 0.9191). However, the lowest number of root tips was found on roots of control plants (Cu0—856.90), while the Cu25 ppm treatment resulted in the highest number of tips (Cu25—1084). The growth parameters in relation to NP treatments are presented in Table 2. The infestation index of powdery mildew ranged from 02.59—Ag50 ppm to 2.80—Cu50 ppm. The effects of NP treatments on the degree of powdery mildew infection were not statistically significant (Table 2).
Table 2.
Biometrical parameters (mean ± SD) and scale infestation of powdery mildew disease of treated seedlings of Quercus robur at different concentrations of AgNPs and CuNPs. Ag0, Cu0—0 ppm; Ag5, Cu5—5 ppm; Ag25, Cu25—25 ppm; Ag50, Cu50—50 ppm.
3.2. Microscopic Investigations of Plant Material
The ultrastructure of shoots and roots in the nanoparticle-treated plants was similar to the control plants. In contrast, plants treated with Cu and Ag nanoparticles exhibited a disturbed ultrastructure in leaves, especially in the photosynthetic apparatus. In the control plants (no AgNP and CuNP), the chloroplasts filled the cells and contained small plastoglobules, whereas the CuNP-treated plants exhibit large plastoglobules; by contrast, the AgNP-treated plants contained large starch granules. Other organelles (e.g., mitochondria, cell walls, vacuoles) remained unchanged. Obtained results indicated that Cu- and AgNPs in high concentration (50 ppm) had influenced the ultrastructure of chloroplasts in oak leaves (Figure 2).
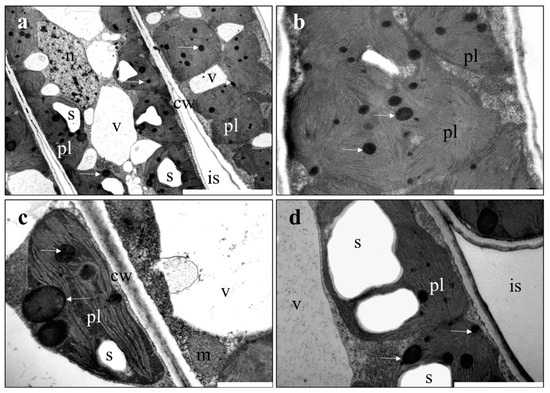
Figure 2.
The ultrastructure of oak leaves was studied using TEM. (a,b) show cross sections of palisade mesophyll cells in control oak leaves; (c) plants treated with 50 ppm CuNPs—chloroplast with disturbed ultrastructure, huge plastoglobules are visible). The chloroplasts from the 50 ppm AgNP—treated plants (d) are normal in shape but contain increased amounts of starch. Abbreviations: cw—cell wall; is—intercellular space; m—mitochondrion; pl—plastid; s—starch; v—vacuole; white arrow—plastoglobule. Scale bars represent: (a) = 5 μm; (b,d) = 2 μm; (c) = 1 μm.
3.3. Mycorrhizal Colonization
The analysed seedlings of Q. robur treated with different concentrations (0, 5, 25, 50 ppm) of two types of nanoparticles (AgNPs, CuNPs) differed significantly (GLM, df = 6, p < 0.0001) in ectomycorrhizal colonization (Figure 3A,B). Mycorrhizal colonisation ranged from 20.8% (Cu50 ppm) to 37.5% (Ag25 ppm). Seedlings treated with 25 ppm CuNPs and AgNPs had the highest degree of mycorrhization (VM): 37.1% and 37.5%, respectively. On the other hand, these seedlings had the highest proportion of non-vital root tips: 6.6%—Cu25 ppm and 4.8%—Ag25 ppm.
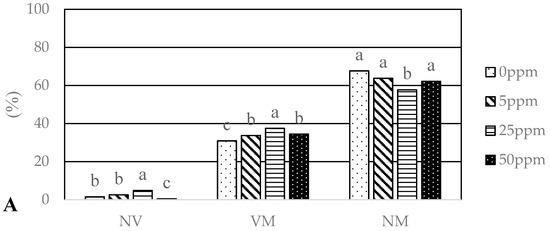
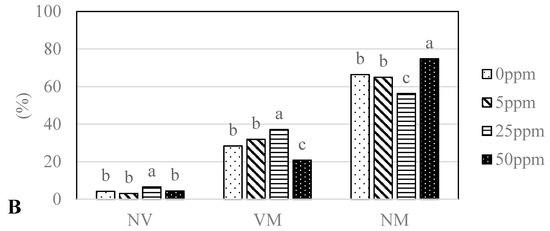
Figure 3.
Abundance of vital mycorrhizal (VM), non-vital (NV) and non-mycorrhizal (NM) root tips (%) of oak seedlings treated with different concentrations of AgNPs (A) and CuNPs (B); Ag0, Cu0—0 ppm; Ag5, Cu5—5 ppm; Ag25, Cu25—25 ppm; Ag50, Cu50—50 ppm. Different letters indicate significant differences among groups assessed with Tukey´s test.
3.4. Mycorrhizal Fungal Communities
Morphotyping and subsequent molecular analysis of the mycorrhizal root tips collected from Q. robur seedlings revealed the presence of 4 mycorrhizal fungal taxa (Table 3, Figure 4). Sphaerosporella brunnea, Thelephora terrestris and Paxillus involutus were recorded in samples from all treatments, while Laccaria proxima was exclusively detected on seedlings treated with Ag5 ppm (Table 3). The most abundantly and frequently observed ECM fungus was S. brunnea.
Table 3.
Frequency and relative abundance of fungal taxa (percent of colonized plants/percent of mycorrhizal roots colonized) on root tips of Quercus robur seedlings treated with different NPs. Ag0, Cu0—0 ppm; Ag5, Cu5—5 ppm; Ag25, Cu25—25 ppm; Ag50, Cu50—50 ppm.

Figure 4.
Mycorrhizal morphotypes observed on English oak seedlings grown in containers in a forest tree nursery. (a) Sphaerosporella brunnea; (b) Thelephora terrestris; (c) Laccaria proxima; (d) Paxillus involutus. Bars, 1 mm.
4. Discussion
4.1. The Effect of Foliar Applications of AgNPs and CuNPs on Biometric Parameters of Oak Seedlings and on Powdery Mildew Disease
In the current study, foliar application of AgNPs and CuNPs at four concentrations: 0, 5, 25 and 50 ppm had insignificant effects on biometrical parameters (Table 2), in agreement with the findings of some other studies (Table 1). Our results were similar to Seeger et al. [15], who showed insignificant acute toxicity of titanium dioxide nanoparticles to willow trees. Effects on transpiration rate, growth, and water use efficiency of willow cuttings exposed to aqueous TiO2 at different concentrations were not statistically significant. Other studies report that the effect of NPs on tree seedling growth can be positive or negative (e.g., [54]; Table 1). For example, Lin et al. [8] experimentally demonstrated that soaking roots of Larix olgensis seedlings in a solution of nanostructured silicon dioxide (TMS) greatly promoted seedling growth and improved seedling quality. In contrast, Sweet and Singleton [13] observed a reduction of root and shoot growth of Pinus muricata exposed to AgNPs.
Nanoparticles may be regarded as pollutants, but they are also used in plant protection [55]. In our study, the experimental application of AgNPs and CuNPs against spontaneously occurring oak powdery mildew was not successful (Table 2). Repeated foliar application of NPs at four concentrations: 0, 5, 25 and 50 ppm failed to reduce the degree of powdery mildew infection. This result is in agreement with Ashrafi et al. [24] but in contrast with Lamsal et al. [25]. Lamsal et al. [25] demonstrated antifungal activities of silver nanoparticles against powdery mildew of cucurbits (Cucurbitaceae). The application of 100 ppm AgNPs showed maximum inhibition of fungal hyphae in the field experiment [25].
4.2. Ultrastructure of Organs of Plants Treated with NPs
Uptake effectiveness and impact of NPs on plant cell structure and metabolism are species specific. In Allium cepa, different cytological effects including chromosomal aberrations, disturbed metaphase and cell disintegration of root tip cells were observed after application of AgNPs [56]. Observation of nanoparticles and their impact inside plant tissues using TEM is a very challenging task. Because of the restricted resolution of the TEM used, we cannot indicate the compartments where NPs accumulate. However, the ultrastructural changes are clearly visible. The most sensitive organelles are plastids, which exhibit evident disturbances in shape, plastoglobules and starch content. Similar effects are known to be caused by heavy metals in the non-nano state. Exposure to elevated levels of heavy metals was observed to result in changes of chloroplasts, thylakoid membranes, plastoglobuli and starch grains [26,28,29], in reduced chlorophyll (Chl) content, decreased Chl a/b ratio and enhanced chlorophyllase activity. Cu is an essential metal which is known to cause damage to various components of photosynthesis, if present in excess. It can (1) disturb the architecture of chloroplasts, particularly that of the thylakoid membranes; (2) inhibit photosynthetic electron transport of both Photosystem I (PSI) and Photosystem II (PSII) and (3) inhibit RUBISCO (Ribulose-1,5-bisphosphat-carboxylase, the key enzyme of the Calvin cycle and photosynthetic CO2 fixation) activity. Excess Cu commonly decreases photosynthetic activity (mainly PSII). Thereby causing damage to the structure and composition of the thylakoid membranes [57,58]. Cu can cause changes in lipid and pigment composition [6]. In Cu-treated plants, degraded intergranal thylakoid membranes, fine starch grains and numerous plastoglobuli are seen in place of normal intergranal thylakoids [6]. Alteration in the structure and composition of the thylakoid membranes caused by Cu(II) influences the conformation and function of photosystems [59]. Moreover, Navarro et al. [60] demonstrated that AgNPs inhibited the photosynthetic yield of algae (Chlamydomonas reinhardtii). It was shown that all differently coated AgNPs (chitosan, lactate, polyvinylpyrrolidone, polyethelene glycol, gelatin, sodium dodecylbenzenesulfonate, citrate, dexpanthenol, carbonate and AgNO3) and dissolved Ag proved to be toxic, decreasing the photosynthetic yield of algae as a function of increasing total Ag concentrations.
Similar effects, mainly an increased number of plastoglobules, were observed in the duckweed Landoltia punctata plants treated with copper oxide NPs [61]. Plastoglobules are subcompartments of thylakoids, containing enzymes that participate in lipid metabolic pathways. It is well documented that under biotic and abiotic stress conditions, the size, as well as the number of plastoglobules increase [62,63]. In watermelon plants, the transport of NPs from the leaves through the stem and into the roots could be traced [64]. In oak seedlings, we have not observed any changes in the ultrastructure of stems and roots in plants treated with NPs. There are two possible explanations for this situation: the NPs are delivered into the roots without causing damaging effects or the NPs accumulate in leaves only. Hong et al. [59] demonstrated translocation and physiological impacts of foliar-applied CeO2 nanoparticles (both powder and spray treatments) on cucumber (Cucumis sativus) plants. After foliage application of CeO2-NP, Ce was detected in all tissues, suggesting uptake through the leaves and translocation to the other plant parts, through stems and roots. Moreover, Ce concentrations in stems and flowers increased as the external concentration of CeO2NPs increased. This finding is in contrast with a study conducted by Birbaum et al. [65]. It was demonstrated that exposure of maize plants to either aerosol or suspension of cerium dioxide nanoparticles resulted in cerium adsorbed to or incorporated into maize leaves. However, no translocation into newly growing leaves was found when cultivating the maize plants after airborne particle exposure. The similarity of ultrastructural changes induced by metal nanoparticles and by metal ions as reported in the literature deserves further study. If both growth-promoting and phytotoxic effects are analogous in metal nanoparticles and metal ions, modelling of the effects of treatment with NPs might be simplified.
NPs from different manufacturers vary in many properties, such as diameter, surface area, surface charge, active groups, and composition, etc. Therefore, the results obtained in our study with AgNPs and CuNPs cannot be transferred to other types of nanomaterials.
4.3. Mycorrhizal Colonization
English oak is strongly dependent on ectomycorrhizal symbiosis [66]. It was shown that healthy oak (Quercus ilex) trees have significantly greater proportions of vital ectomycorrhizal root tips than declining trees [67]. On the other hand, crown defoliation may result in the reduction of photosynthetic activity, which negatively affects the below-ground carbon allocation on which ECM symbionts depend [31,68].
In our study, the ectomycorrhizal colonisation ranged from 20.8% (CuNPs 50 ppm) to 37.5% (AgNPs 25 ppm). Such levels of ectomycorrhizal colonization are common in forest nurseries, albeit low if compared to the high levels typically observed in vital forest stands. Menkis et al. [69] observed that cultivation systems in forest nurseries had a marked effect on the degree of mycorrhization. In pine seedlings, the highest level of colonisation (48%) was observed in bare-root systems, followed by plants from plastic container trays (41.9%) and by bare-root seedlings cultivated in the greenhouse (19.4%).
Different effects of NPs on AM colonization were shown [16,39,70], but only very limited evidence is available concerning the effects of NPs on ECM colonisation and community structure. To the best of our knowledge, only one scientific study relates to this issue. It was demonstrated that experimental soil contamination with AgNPs reduces Bishop pine growth, as well as the level of mycorrhizal colonisation and ectomycorrhizal diversity on pine roots. Five different genera of ECM fungi were found on roots of control plants, but only the genus Laccaria was found on roots of plants grown in soil containing 350 mg Ag/kg. At higher levels of AgNP (790 mg Ag/kg) contamination, no ECM fungi were observed.
In the current study, the lowest degree of mycorrhization of oak was observed on seedlings treated with the highest CuNPs concentrations (Figure 2). In oak seedlings, foliar application of AgNPs and CuNPs at concentrations of 25 ppm resulted in a stimulation of mycorrhizal colonisation. We hypothesise that foliar treatment with 25 ppm CuNPs may have released the seedlings from latent Cu deficiency. Cu is an essential micronutrient involved in a multitude of metabolic processes, particularly redox reactions and a potentially toxic source of free radicals. Cu metabolism is precisely regulated by plants and ectomycorrhizal fungi to maintain adequate levels of this trace metal [71]. In particular, the copper protein plastocyanin in higher plants is essential for photosynthetic electron transport [72]. Furthermore, Cu acts as a prosthetic group of the copper/zinc superoxide dismutase (SOD Cu), which protects the photosynthetic apparatus from oxidative stress. Higher efficiency of photosynthesis due to improved Cu micronutrient status might explain the observed increase in ectomycorrhizal colonisation, since the level of mycorrhizal colonisation dependents on allocation of organic C to mycorrhizal roots. However, the Cu-deficiency release hypothesis is challenged by two facts: 1. Osmocote Exact Standard fertilizer used in our experiment includes copper as a trace element. 2. The absence of a plant growth response to CuNPs.
Direct or indirect inhibitory effects of higher Cu concentrations on root growth and ECM formation are possible. Root elongation and branching are negatively affected by excess Cu, potentially limiting nutrient and water uptake [73]. The application of 1 μM Cu supplied as CuSO4 was reported to reduce taproot elongation in Pinus pinea and P. pinaster [74]. In the case of fungicides, foliar application affects the formation and development of mycorrhizae to a lesser extent than soil treatment [75]. The effects of foliar applications of nanoparticles may be limited in an analogous way.
In the presented study, ECM fungi found on oak roots, such as S. brunnea, T. terrestris, L. proxima and P. involutus were identified as typical mycobionts of young oaks in early succession [76]. The pyrophilous ascomycete S. brunnea is a frequent companion in forest nurseries and considered a contaminant in the production of tree seedlings mycorrhized with edible truffle species [77]. Baum et al. [78] observed a higher percentage of Populus trichocarpa fine roots colonized with Laccaria bicolor and P. involutus in soil rich in organic carbon and in nitrogen compared to a nitrogen-poor soil. Furthermore, fruiting bodies of the ECM fungi recorded in this study have been frequently recorded in heavily polluted soils. It has been shown that inoculation of willow trees with P. involutus can increase the phytoextraction of heavy metals [79]. Toxic heavy metals are believed to affect fungal communities by reducing abundance and species diversity and by selecting for resistant/tolerant taxa [79]. However, the phylogenetic diversity of ECM communities colonizing sites with potentially toxic soil contents of heavy metals was found high by some studies [71,80], suggesting that the capacity of acquiring avoidance and resistance mechanisms is widespread among fungi.
Nanoparticles might be compared to systemic pesticides, if they are taken up and internally redistributed by the target plants. For instance, the persistent systemic fungicide triadimefon, currently prohibited in the EU and other countries, was shown to be translocated to pine roots, strongly inhibiting the formation of ectomycorrhizal symbioses [81]. Are the tested NPs being translocated from leaves into the roots, do they accumulate in the roots or are they released into the rhizosphere? Concerning the translocation of NPs within plants, the route from soil via roots and xylem to shoots and leaves (and ultimately to consumer food webs, including humans) is better studied than the reverse direction [82,83]. Foliar uptake and potential phloem-based transport of metal particles including nanoparticles by vascular plants depend on a variety of factors, such as exposure conditions, metal element, particle size, chemical speciation, physiochemical properties (crystallinity, surface charge, hydrobicity/hydrophility, etc.) and plant species and condition [82,83,84]. Cerium dioxide NPs applied as suspension or aerosol were found to be either adsorbed to or taken up into maize leaves. Uptake of NPs into plant tissues was not dependent on stomata. aperture, and no translocation of NPs into newly grown leaves was observed [65]. Basipetal translocation (from leaves to roots) was demonstrated in maize for CuONPs [64] and for CeO2NPs [59], showing that NPs transport via phloem could potentially affect root-associated symbionts. However, we are not aware of any research assessing the potential impact of foliar application of NPs on ectomycorrhizal symbiosis apart from our study. Our results indicate that foliar application of NPs might not necessarily harm the biodiversity and function of ectomycorrhizal fungi. However, the ectomycorrhizal fungal species typically present under greenhouse conditions, including opportunistic extremophiles which are well adapted to stressful environments, such as S. brunnea [77], are just a minute subset of ectomycorrhizal biodiversity. It was demonstrated experimentally that S. brunnea propagates rapidly through asexual propagation by conidiospores and homothallism, which makes this fungus a pioneer and opportunist ectomycorrhizal species, and the most common fungal competitor in nurseries producing plants [77]. Therefore, the results of this greenhouse trial cannot be extrapolated to field conditions.
5. Conclusions
None of the tested AgNPs and CuNPs were manifested as being phytotoxic to the examined Q. robur seedlings under container nursery conditions, and no significant effect on growth parameters of oak seedlings was observed. Ultrastructural changes after NP treatments were detected in leaf chloroplasts but not in shoot and root tissues. The degree of mycorrhization increased significantly in plants treated with AgNPs and CuNPs at concentrations 25 ppm, while the treatment with CuNPs at 50 ppm resulted in the lowest degree of mycorrhization, indicating non-linear dose-dependent effects of NP treatments. The application of four NPs treatments was not effective in controlling oak powdery mildew infection.
Acknowledgments
The first author gratefully acknowledges the Austrian Agency for International Mobility and Cooperation in Education, Science and Research (OeAD) for supporting his stay at the University of Vienna in 2015 and the WULS-SGGW International Research Scholarship Fund for supporting his stay at the University of Vienna in 2016. The study was financed by the Rector of the Warsaw University of Live Sciences—SGGW in the research project: 505-10-030400-L00374-99.
Author Contributions
Jacek Olchowik generated the data. All authors analysed and discussed the data. The manuscript was written by Roman Mariusz Bzdyk, Jacek Olchowik, Alexander Urban and Magdalena Bederska-Błaszczyk. Statistical analysis using R version 2.15.0 (The R Foundation for Statistical Computing, Vienna, Austria) with the vegan package for multivariate analysis: Marcin Studnicki. The general conception: Marta Aleksadrowicz–Trzcińska.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ball, P. Natural strategies for the molecular engineer. Nanotechnology 2002, 13, 15–28. [Google Scholar] [CrossRef]
- Roco, M.C. Broader societal issue of nanotechnology. J. Nanopart. Res. 2003, 5, 181–189. [Google Scholar] [CrossRef]
- Nel, A.; Xia, T.; Madler, L.; Li, N. Toxic potential of materials at the nanolevel. Science 2006, 311, 622–627. [Google Scholar] [CrossRef] [PubMed]
- Ciancio, A.; Mukerji, K.G. General Concepts in Integrated Pest and Disease Management; Springer: Dordrecht, The Netherlands, 2007; pp. 27–43. [Google Scholar]
- Smita, S.; Gupta, S.K.; Bartonova, A.; Dusinska, M.; Gutleb, A.C.; Rahman, Q. Nanoparticles in the environment: Assessment using the causal diagram approach. Environ. Health 2012, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Han, J.; Xiao, J.Q.; Jin, Y. Uptake, translocation and accumulation of manufactured iron oxide nanoparticles by pumpkin plants. J. Environ. Monit. 2008, 10, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Husen, A.; Siddiqi, K.S. Phytosynthesis of nanoparticles: Concept, controversy and application. Nanoscale Res. Lett. 2014. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.S.; Diao, S.Q.; Li, C.H.; Fang, L.J.; Qiao, S.C.; Yu, M. Effect of TMS (nanostructured silicon dioxide) on growth of Changbai larch seedlings. J. For. Res. 2004, 15, 138–140. [Google Scholar]
- Lin, D.; Xing, B. Phytotoxicity of nanoparticles: Inhibition of seed germination and root growth. Environ. Pollut. 2007, 150, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Geiser-Lee, J.; Deng, Y.; Kolmakov, A. Interactions between engineered nanoparticles (ENPs) and plants: Phytotoxicity, uptake and accumulation. Sci. Total Environ. 2010, 408, 3053–3061. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Lian, K.; Wu, Q.; Li, Y.; Danzy, M.; Menard, R.; Chin, K.L.; Collins, D.; Oliveria, F.; Klepzig, K. Nanotechnology application in forest protection. In Proceedings of the TAPPI International Conference on Nanotechnology for Renewable Materials, Washington, DC, USA, 6–8 June 2011; pp. 271–301. [Google Scholar]
- Lee, W.; An, Y.; Yoon, H.; Kweon, H. Toxicity and bioavailability of copper nanoparticles to the terrestrial plants mung bean (Phaseolus radiatus) and wheat (Triticum aestivum): Plant uptake for water insoluble nanoparticles. Environ. Toxicol. Chem. 2008, 27, 1915–1921. [Google Scholar] [CrossRef] [PubMed]
- Sweet, M.J.; Singleton, I. Soil contamination with silver nanoparticles reduces Bishop pine growth and ectomycorrhizal diversity on pine roots. J. Nanopart. Res. 2015. [Google Scholar] [CrossRef] [PubMed]
- Rostami, A.A.; Shahsavar, A. Nano-silver particles eliminate the in vitro contaminations of olive ‘mission’ explants. Asian J. Plant Sci. 2009, 8, 505–509. [Google Scholar] [CrossRef]
- Seeger, E.M.; Baun, A.; Kästner, M.; Trapp, S. Insignificant acute toxicity of TiO2 nanoparticles to willow trees. J. Soil. Sediment 2009, 9, 46–53. [Google Scholar] [CrossRef]
- Feng, Y.; Cui, X.; He, S.; Dong, G.; Chen, M.; Wang, J. The role of metal nanoparticles in influencing arbuscular mycorrhizal fungi effects on plant growth. Environ. Sci. Technol. 2013, 47, 9496–9504. [Google Scholar] [CrossRef] [PubMed]
- El-Temsah, Y.S.; Joner, E.J. Impact of Fe and Ag nanoparticles on seed germination and differences in bioavailability during exposure in aqueous suspension and soil. Environ. Toxicol. 2012, 27, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Savithramma, N.; Ankanna, S.; Bhumi, G. Effect of nanoparticles on seed germination and seedling growth of Boswellia ovalifoliolata an endemic and endangered medicinal tree taxon. Nano Vis. 2012, 2, 61–68. [Google Scholar]
- Stampoulis, D.; Sinha, S.K.; White, J.C. Assay-dependent phytotoxicity of nanoparticles to plants. Environ. Sci. Technol. 2009, 43, 9473–9479. [Google Scholar] [CrossRef] [PubMed]
- Dimkpa, C.O.; McLean, J.E.; Latta, D.E.; Manangón, E.; Britt, D.W.; Johnson, W.P.; Boyanov, M.I.; Anderson, A.J. CuO and ZnO nanoparticles: Phytotoxicity, metal speciation, and induction of oxidative stress in sand-grown wheat. J. Nano Res. 2012, 14, 1–15. [Google Scholar] [CrossRef]
- Lilja, A.; Poteri, M. Infectious Forest Diseases; Gonthier, P., Nicolotti, G., Eds.; CBA International: Wallingford, UK, 2013; pp. 567–592. [Google Scholar]
- Ouda, S.M. Antifungal Activity of Silver and Copper Nanoparticles on Two Plant Pathogens, Alternaria alternata and Botrytis cinerea. Res. J. Microbiol. 2014, 9, 34–42. [Google Scholar] [CrossRef]
- Gorczyca, A.; Pociecha, E.; Kasprowicz, M.; Niemiec, M. Effect of nanosilver in wheat seedlings and Fusarium culmorum culture systems. Eur. J. Plant Pathol. 2015, 142, 251–261. [Google Scholar] [CrossRef]
- Ashrafi, S.J.; Rastegar, M.F.; Nekouei, M.K. Use of silver Nano particle for controlling the powdery mildew diseases at Celastrales plants. Am. Eurasian J. Agric. Environ. Sci. 2010, 7, 70–74. [Google Scholar]
- Lamsal, K.; Kim, S.W.; Jung, J.H.; Kim, Y.S.; Kim, K.S.; Lee, Y.S. Inhibition Effects of Silver Nanoparticles against Powdery Mildews on Cucumberand Pumpkin. Mycobiology 2011, 39, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: Cambridge, UK, 2008. [Google Scholar]
- Marin, M. Effects of pesticides on the growth of ectomycorrhizal fungi and ectomycorrhiza formation. In Diversity and Biotechnology of Ectomycorrhizae; Rai, M., Varma, A., Eds.; Springer: Berlin, Germany, 2010; pp. 323–346. [Google Scholar]
- Scholl, L.; Hoffland, E.; van Breemen, N. Organic anion exudation by ectomycorrhizal fungi and Pinus sylvestris in response to nutrient deficiencies. New Phytol. 2006, 170, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Scholl, L.; Kuyper, T.W.; Smits, M.M.; Landeweert, R.; Hoffland, E.; van Breemen, N. Rock-eating mycorrhizas: Their role in plant nutrition and biogeochemical cycles. Plant Soil 2008, 303, 35–47. [Google Scholar] [CrossRef]
- Finlay, R.D. Ecological aspects of mycorrhizal symbiosis: With special emphasis on the functional diversity of interactions involving the extraradical mycelium. J. Exp. Bot. 2008, 59, 1115–1126. [Google Scholar] [CrossRef] [PubMed]
- Finlay, R.D.; Söderström, B. Mycorrhiza and carbon flow to soil. In Mycorrhizal Functioning; Allen, M.F., Ed.; Chapman and Hall: London, UK, 1992; pp. 134–160. [Google Scholar]
- Colpaert, J.V. Heavy metal pollution and genetic adaptations in ectomycorrhizal fungi. In Stress in Yeasts and Filamentous Fungi; Avery, S., Stratford, M., van West, P., Eds.; Elsevier: Amsterdam, The Netherlands, 2008; pp. 157–173. [Google Scholar]
- Finlay, R.D.; Lindahl, B.D.; Taylor, A.F.S. Responses of mycorrhizal fungi to stress. In Stress in Yeasts and Filamentous Fungi; Avery, S., Stratford, M., van West, P., Eds.; Elsevier: Amsterdam, The Netherlands, 2008; pp. 201–220. [Google Scholar]
- Izumi, H.; Anderson, I.C.; Alexander, I.J.; Killam, K.; Moore, E.R.B. Diversity and expression of nitrogenase genes (nifH) from ectomycorrhizas of Corsican pine (Pinus nigra). Environ. Microbiol. 2006, 8, 2224–2230. [Google Scholar] [CrossRef] [PubMed]
- Power, S.A.; Ashmore, M.R. Nutrient relations and root mycorrhizal status of healthy and declining beech (Fagus sylvatica L) in southern Britain. Water Air Soil Pollut. 1996, 86, 317–333. [Google Scholar] [CrossRef]
- Kovacs, G.; Pausch, M.; Urban, A. Diversity of ectomycorrhizal morphotypes and oak decline. Phyton Ann. Rei Bot. A 2000, 40, 109–116. [Google Scholar]
- Scattolin, L.; Dal Maso, E.; Mutto Accord, S.; Sella, L.; Montecchio, L. Detecting asymptomatic ink-diseased chestnut trees by the composition of the ectomycorrhizal community. For. Pathol. 2012, 42, 501–509. [Google Scholar] [CrossRef]
- Manceau, A.; Nagy, K.L.; Marcus, M.A.; Lanson, M.; Geoffroy, N.; Jacquet, T.; Kirpichtchikova, T. Formation of metallic copper nanoparticles at the soil-root interface. Environ. Sci. Technol. 2008, 42, 1766–1772. [Google Scholar] [CrossRef] [PubMed]
- Dubchak, S.; Ogar, A.; Mietelski, J.W.; Turnau, K. Influence of silver and titanium nanoparticles on arbuscular mycorrhiza colonization and accumulation of radiocaesium in Helianthus annuus. Span. J. Agric. Res. 2010, 8, 103–108. [Google Scholar] [CrossRef]
- Judy, J.D.; Kirby, J.K.; Creamer, C.; McLaughlin, M.J.; Fiebiger, C.; Wright, C.; Cavagnaro, T.R.; Bertsch, P.M. Effects of silver sulfide nanomaterials on mycorrhizal colonization of tomato plants and soil microbial communities in biosolid-amended soil. Environ. Pollut. 2015, 206, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, S. Zasady postępowania technologicznego przy zastosowaniu polskiego biopreparatu z grzybem Hebeloma crustuliniforme w zabiegu sterowanej mikoryzacji sadzonek drzew leśnych. In Ektomikoryzy. Nowe Biotechnologie w Polskim Szkółkarstwie Leśnym; Kowalski, S., Ed.; CILP: Warszawa, Poland, 2007; pp. 101–103. [Google Scholar]
- Nanogrp.com. Available online: http://www.nanogrp.com/pl/ (accessed on 15 April 2017).
- Karnovsky, M.J. A formaldehyde-glutaraldehyde fixative of high osmolality for use in electron microscopy. J. Cell. Biol. 1965, 27, 1A–149A. [Google Scholar]
- Agerer, R. Colour Atlas of Ectomycorrhizae, 1st–15th ed.; Einhorn: Munich, Germany, 1987–2008. [Google Scholar]
- Agerer, R. Exploration types of ectomycorrhizae. A proposal to classify ectomycorrhizal mycelial systems according to their patterns of differentiation and putative ecological importance. Mycorrhiza 2001, 11, 107–114. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Iotti, M.; Zambonelli, A. A quick and precise technique for identifying ectomycorrhizas by PCR. Mycol. Res. 2006, 110, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of Fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B. Vegan: Community Ecology Package. R Package Version 2.4-2. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 24 August 2017).
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed]
- Khot, L.R.; Sankaran, S.; Maja, J.M.; Ehsani, R.; Schuster, E.W. Applications of nanomaterials in agricultural production and crop protection. Crop. Prot. 2012, 35, 64–70. [Google Scholar] [CrossRef]
- Zhang, Z.; Kong, F.; Vardhanabhuti, B.; Mustapha, A.; Lin, M. Detection of engineered silver nanoparticle contamination in pears. J. Agric. Food Chem. 2012, 60, 10762–10767. [Google Scholar] [CrossRef] [PubMed]
- Kumari, M.; Mukherjee, A.; Chandrasekaran, N. Genotoxicity of silver nanoparticles in Allium cepa L. Sci. Total Environ. 2010, 407, 5243–5246. [Google Scholar] [CrossRef] [PubMed]
- Battke, F.; Leopold, K.; Maier, M.; Schmidhalter, U.; Schuster, M. Palladium exposure of barley: Uptake and effects. Plant. Biol. 2009, 10, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Harris, A.T.; Bali, R. On the formation and extent of uptake of silver nanoparticles by live plants. J. Nanopart. Res. 2008, 10, 691–695. [Google Scholar] [CrossRef]
- Hong, J.; Peralta-Videa, J.R.; Rico, C.; Sahi, S.; Viveros, M.N.; Bartonjo, J.; Zhao, L.; Gardea-Torresdey, J.L. Evidence of translocation and physiological impacts of foliar applied CeO2 nanoparticles on cucumber (Cucumis sativus) plants. Environ. Sci. Technol. 2014, 48, 4376–4385. [Google Scholar] [CrossRef] [PubMed]
- Navarro, E.; Wagner, B.; Odzak, N.; Sigg, L.; Behra, R. Effects of Differently Coated Silver Nanoparticles on the Photosynthesis of Chlamydomonas reinhardtii. Environ. Sci. Technol. 2015, 49, 8041–8047. [Google Scholar] [CrossRef] [PubMed]
- Lalau, C.M.; de Almeida Mohedano, R.; Schmidt, E.C.; Bouzon, Z.; Ouroques, L.C.; dos Santos, R.; da Costa, C.H.; Vicentini, D.S.; Matia, W.G. Toxicological effects of copper oxide nanoparticles on the growth rate, photosynthetic pigment content, and cell morphology of the duckweed Landoltia punctata. Protoplasma 2014, 252, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Selga, T.; Selga, M. Response of Pinus silvestris L. needles to electromagnetic fields. Cytological and ultrastructural aspects. Sci. Total Environ. 1996, 180, 65–73. [Google Scholar] [CrossRef]
- Piller, L.E.; Abraham, M.; Dormann, P.; Kessler, F.; Besagni, C. Plastid lipid droplets at the crossroads of prenylquinone metabolism. J. Exp. Bot. 2012, 63, 1609–1618. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Tarafdar, J.C.; Biswa, S.P. Nanoparticle synthesis and delivery by an aerosol route for water melon plant foliar uptake. J. Nanopart. Res. 2013. [Google Scholar] [CrossRef]
- Birbaum, K.; Brogioli, R.; Schellenberg, M.; Martinoia, E.; Stark, W.J.; Gunter, D.; Limbach, L.K. No evidence for cerium dioxide NP translocation in maize plants. Environ. Sci. Technol. 2010, 44, 8718–8723. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Qiu, Y.L. Phylogenetic distribution and evolution of mycorrhizas in land plants. Myzorrhiza 2006, 16, 299–363. [Google Scholar] [CrossRef] [PubMed]
- Corcobado, T.; Vivas, M.; Moreno, G.; Solla, A. Ectomycorrhizal symbiosis in declining and non-declining Quercus ilex trees infected with or free of Phytophthora Cinnamomi. Forest Ecol. Manag. 2014, 324, 72–80. [Google Scholar] [CrossRef]
- Kuikka, K.; Härmä, E.; Markkola, A.; Rautio, P.; Roitto, M.; Saikkonen, K.; Ahonen-Jonnarth, U.; Finlay, R.; Tuomi, J. Severe defoliation of Scots pine reduces reproductive investment by ectomycorrhizal symbionts. Ecology 2003, 84, 2051–2061. [Google Scholar] [CrossRef]
- Menkis, A.; Vasiliauskas, R.; Taylor, A.F.; Stenlid, J.; Finlay, R. Fungal communities in mycorrhizal roots of conifer seedlings in forest nurseries under different cultivation systems, assessed by morphotyping, direct sequencing and mycelial isolation. Mycorrhiza 2005, 16, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Abbasian, M.; Kashani, A.; Ardakani, M.R.; Rejali, F.; Timajchi, M.; Seifi, S.M.; Mafakheri, S. The effects of chemical, biological and nano fungicides on mycorrhizal colonization and quality of sunflower. Ann. Biol. Res. 2012, 3, 4239–4245. [Google Scholar]
- Urban, A. Metal elements and the diversity and function of ectomycorrhizal communities. In Diversity and Biotechnology of Ectomycorrhiza; Rai, M., Varma, A., Eds.; Springer: Berlin, Germany, 2011; Volume 25, pp. 231–254. [Google Scholar]
- Yamasaki, H.; Pilon, M.; Shikanai, T. How do plants respond to copper deficiency? Plant Signal Behav. 2008, 3, 231–232. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.H.; Bradshaw, A.D. A comparison of the toxicity of heavy metals, using root elongation of rye grass, Lolium perenne. New Phytol. 1982, 91, 255–261. [Google Scholar] [CrossRef]
- Arduini, I.; Godbold, D.L.; Onnis, A. Influence of copper on root growth and morphology of Pinus pinea L. and Pinus pinaster Ait. seedlings. Tree Physiol. 1995, 15, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Trappe, J.M.; Molina, R.; Castellano, M. Reactions of mycorrhizal fungi and mycorrhiza formation to pesticides. Annu. Rev. Phytopathol. 1984, 22, 331–359. [Google Scholar] [CrossRef]
- Gherghel, F.; Krause, K. Role of mycorrhiza in re-forestation at heavy metal-contaminated sites. In Bio-Geo Interactions in Metal-Contaminated Soils; Kothe, E., Varma, A., Eds.; Springer: Berlin, Germany, 2012; Volume 31, pp. 183–199. [Google Scholar]
- Sánchez, S.; Gómez, E.; Martín, M.; De Miguel, A.M.; Urban, A.; Barriuso, J. Experiments on the life cycle and factors affecting reproduction of Sphaerosporella brunnea provide evidence for rapid asexual propagation by conidiospores and for homothallism in an ectomycorrhizal competitor of cultivated truffle species. Fungal Ecol. 2014, 8, 59–65. [Google Scholar] [CrossRef]
- Baum, C.; Schmid, K.; Makeschin, F. Interactive effects of substrates and ectomycorrhizal colonization on growth of a poplar clone. J. Plant Nutr. Soil Sci. 2000, 163, 221–226. [Google Scholar] [CrossRef]
- Baum, C.; Hrynkiewicz, K.; Leinweber, P.; Meißner, R. Heavy-metal mobilization and uptake by mycorrhizal and nonmycorrhizal willows (Salix × dasyclados). J. Plant Nutr. Soil Sci. 2006, 169, 516–522. [Google Scholar] [CrossRef]
- Urban, A.; Puschenreiter, M.; Strauss, J.; Gorfer, M. Diversity and structure of ectomycorrhizal and co-associated fungal communities in a serpentine soil. Mycorrhiza 2008, 18, 339–354. [Google Scholar] [CrossRef] [PubMed]
- Marx, D.H.; Cordell, C.E.; France, R.C. Effects of triadimefon on growth and ectomycorrhizal development of loblolly and slash pines in nurseries. Phytopathology 1986, 76, 824–831. [Google Scholar] [CrossRef]
- Deng, Y.; White, J.C.; Xing, B. Interactions between Engineered Nanomaterials and Agricultural Crops: Implications for Food Safety. J. Zhejiang Univ. Sci. A 2014, 15, 552–572. [Google Scholar] [CrossRef]
- Pacheco, I.; Bueza, C. Nanoparticle Interaction with Plants. In Nanoscience and Plant–Soil Systems; Ghorbanpour, M., Manika, K., Varma, A., Eds.; Springer International Publishing: Cham, Switzerland, 2017; Volume 48, pp. 323–355. [Google Scholar]
- Schreck, E.; Foucault, Y.; Sarret, G.; Sobanska, S.; C´ecillon, L.; Castrec-Rouelle, M.; Uzu, G.; Dumat, C. Metal and metalloid foliar uptake by various plant species exposed to atmospheric industrial fallout: Mechanisms involved for lead. Sci. Total Environ. 2012, 427, 253–262. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).