Changes in Seed Germination Ability, Lipid Peroxidation and Antioxidant Enzyme Activities of Ginkgo biloba Seed during Desiccation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Designs
2.2. Measurements
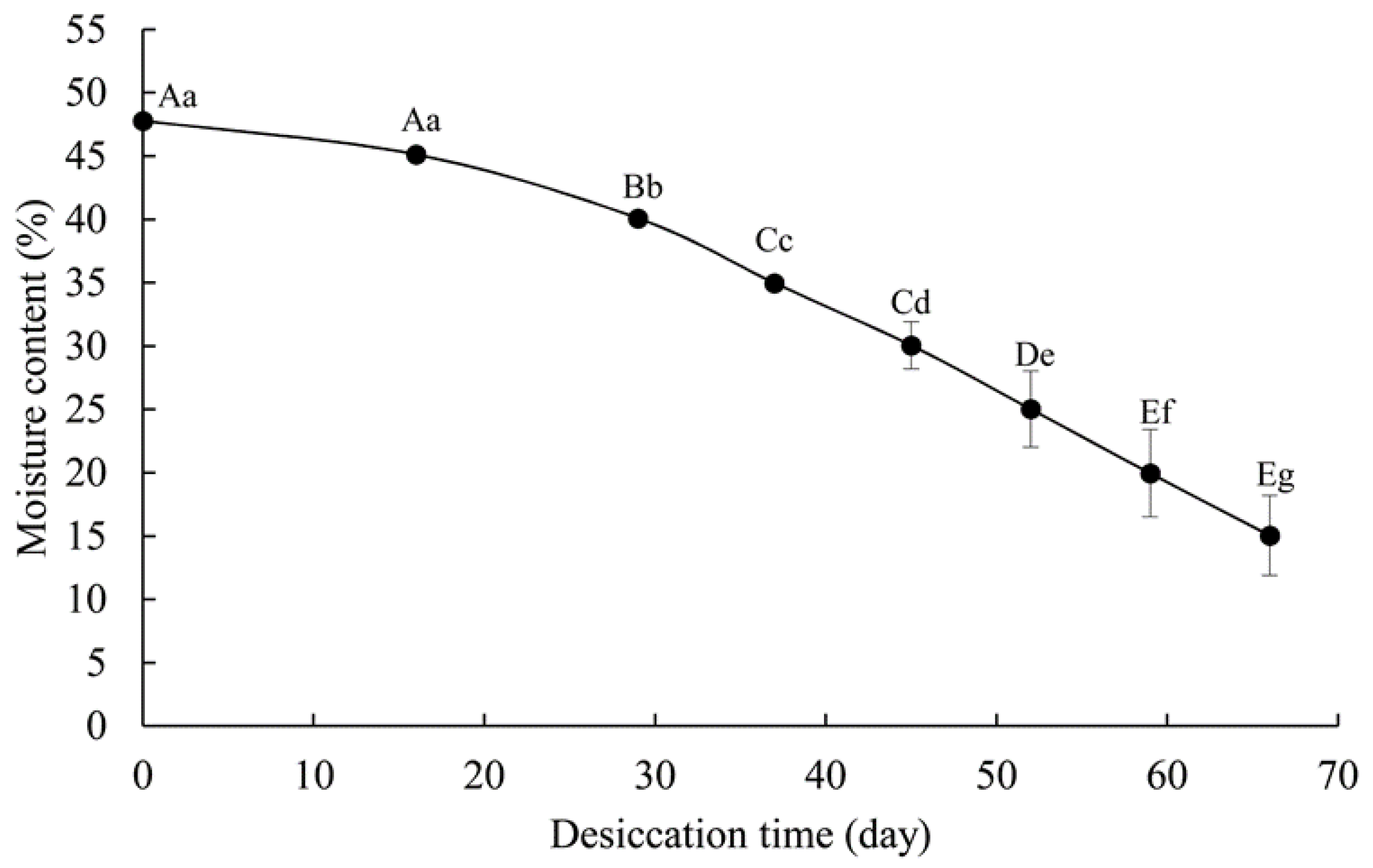
2.2.1. Determination of Moisture Content (MC)
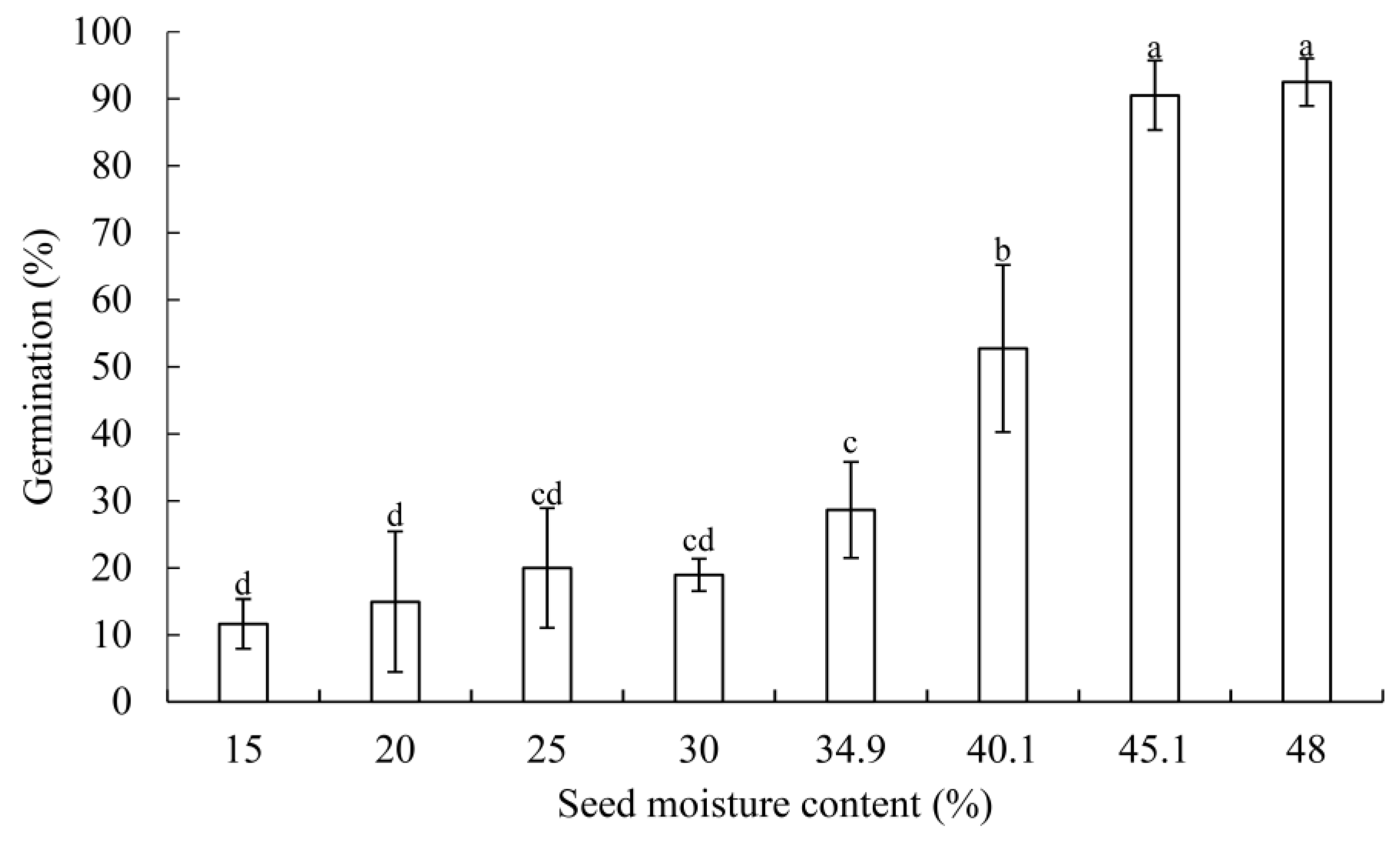
2.2.2. Germination Test
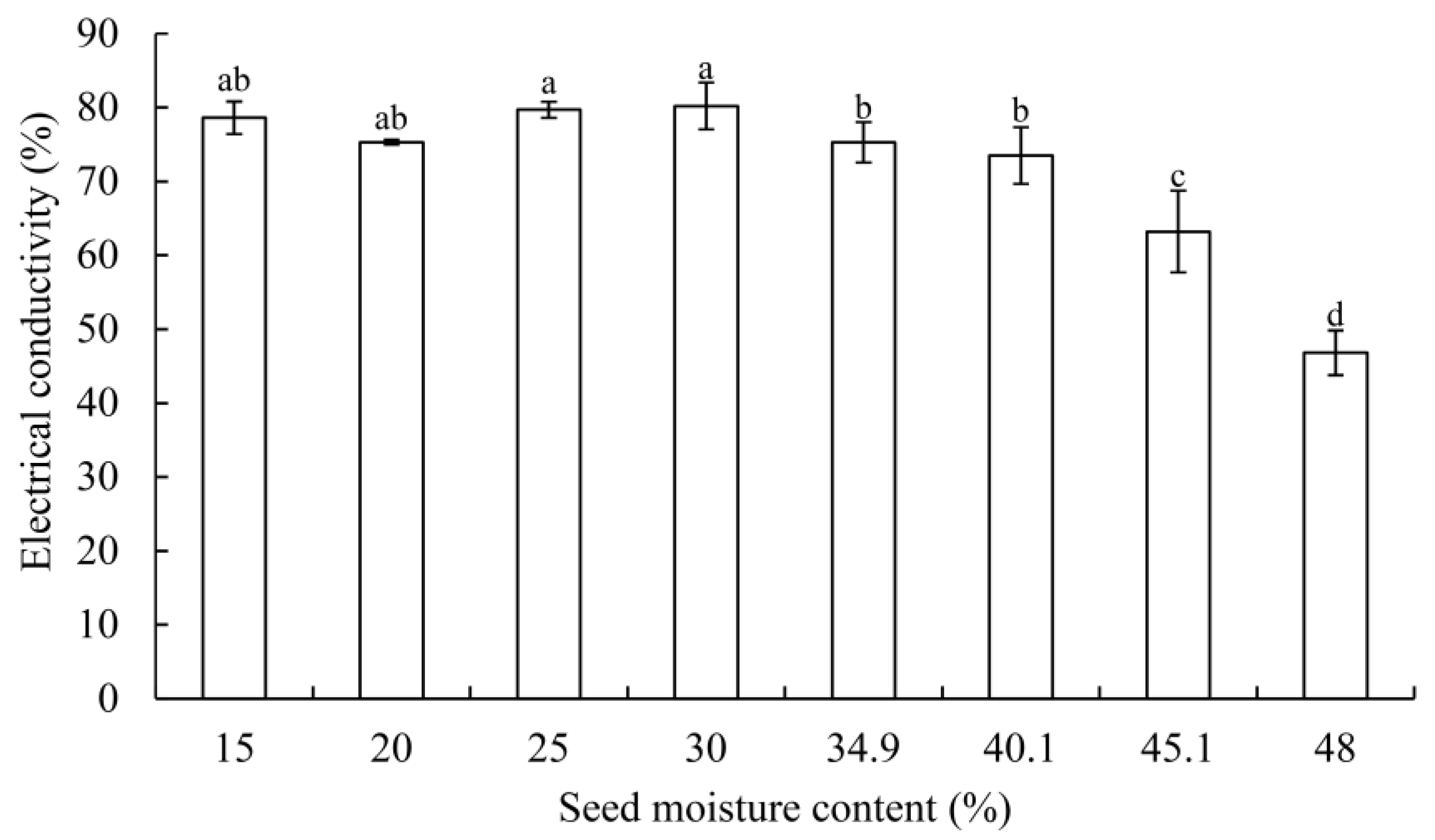
2.2.3. Electrical Conductivity Assay
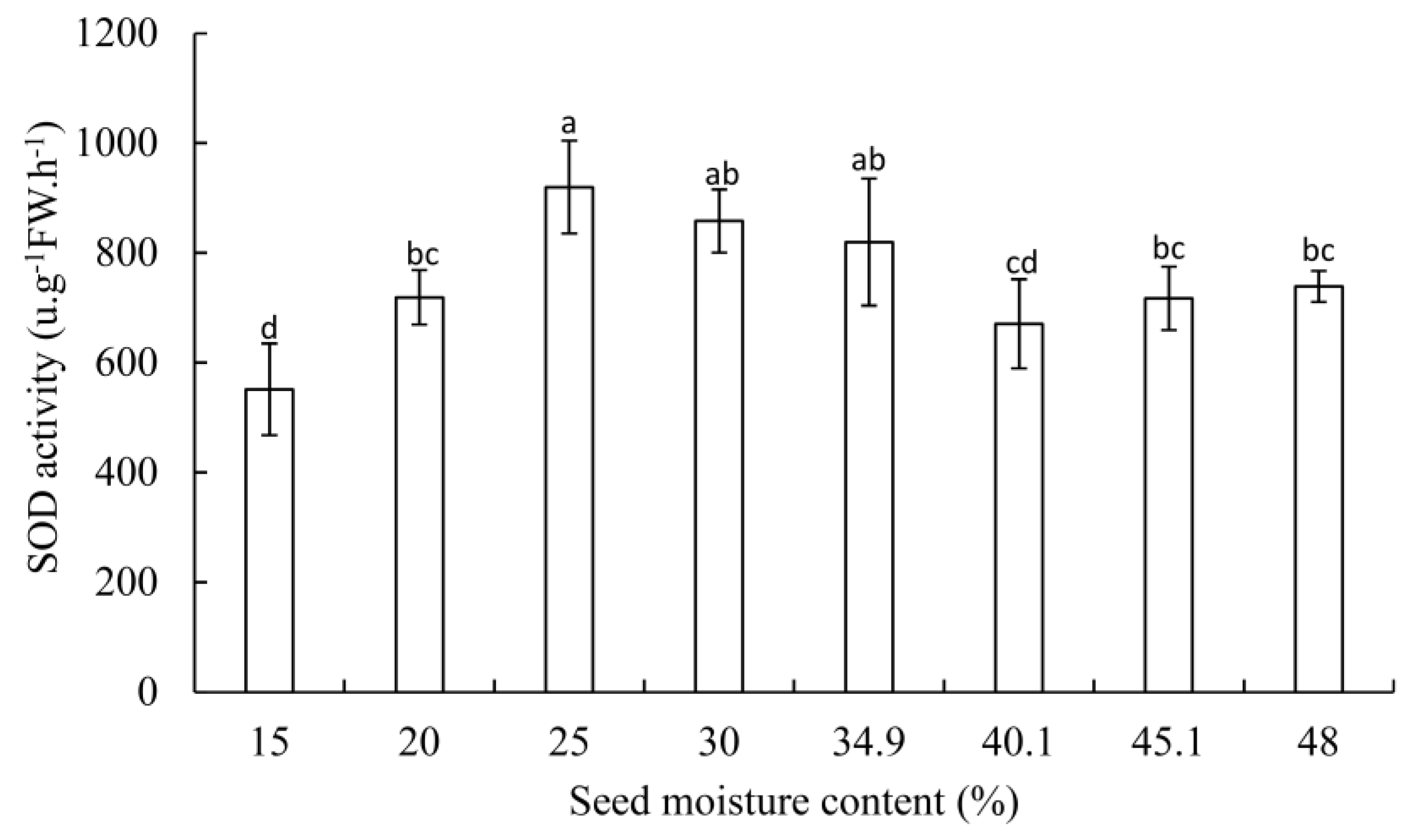
2.2.4. Determination of Anti-Oxidation System (SOD, POD Activity, and Malondialdehyde (MDA) Content)
2.3. Statistical Analysis
3. Results
3.1. Germination Changes and Electrical Conductivity
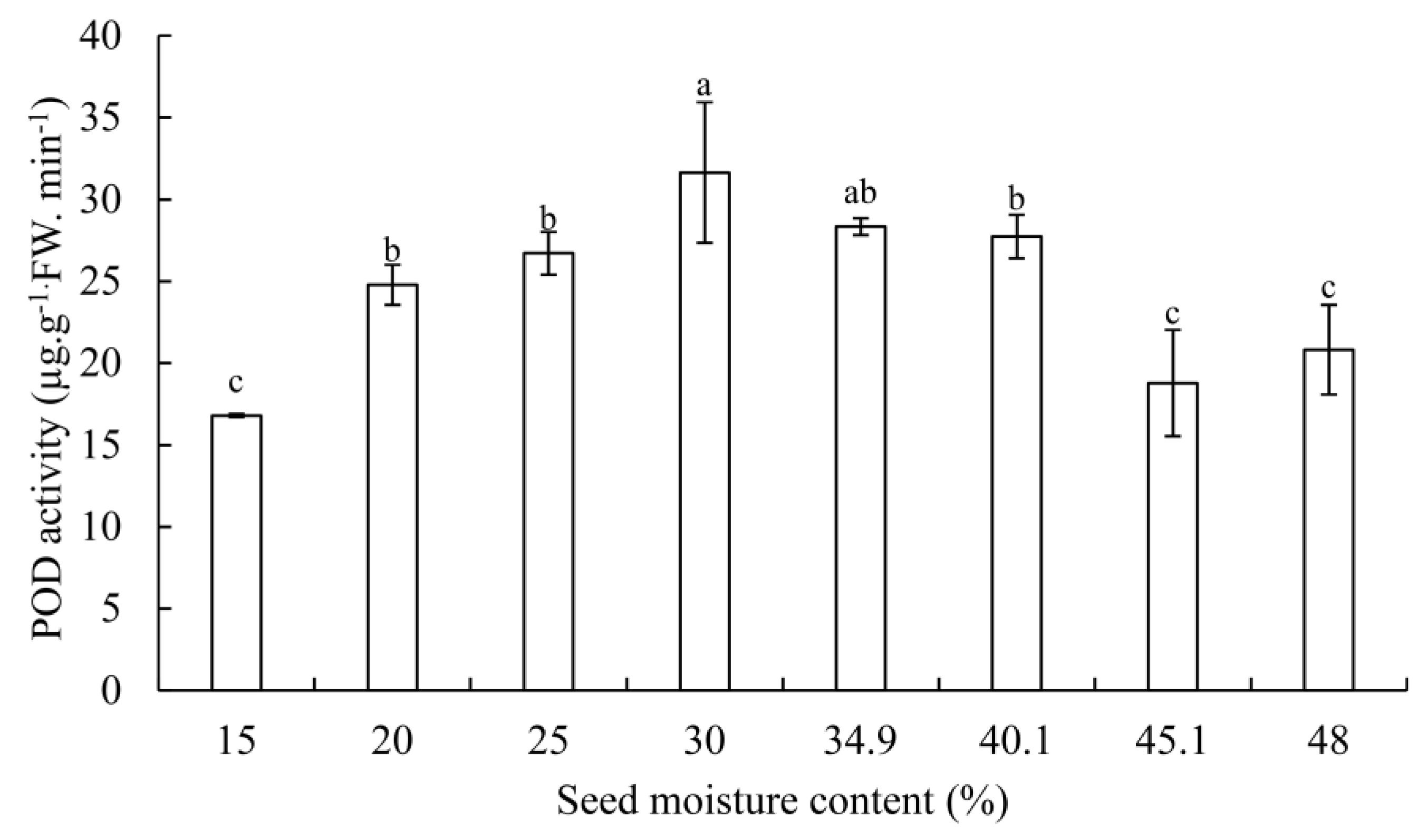
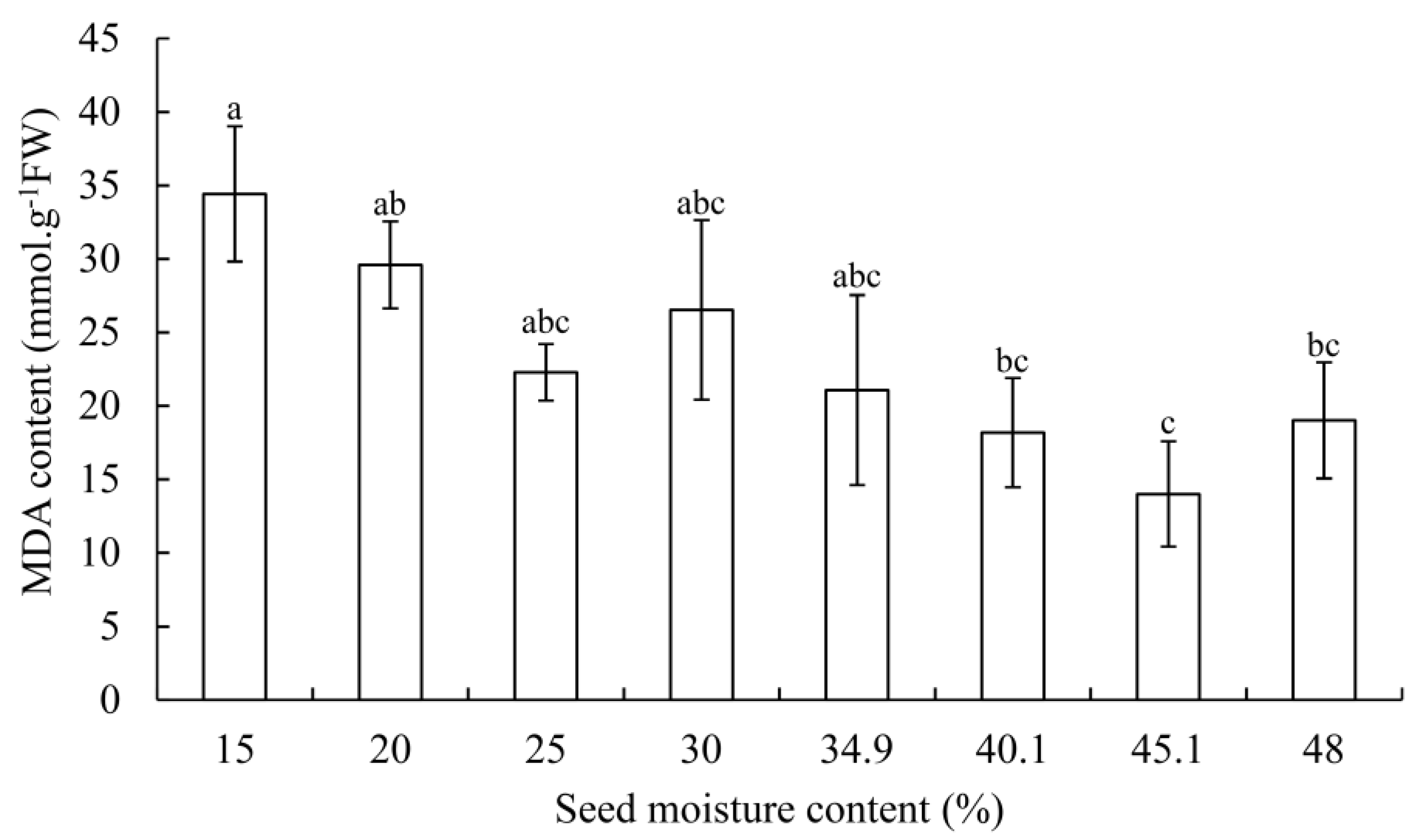
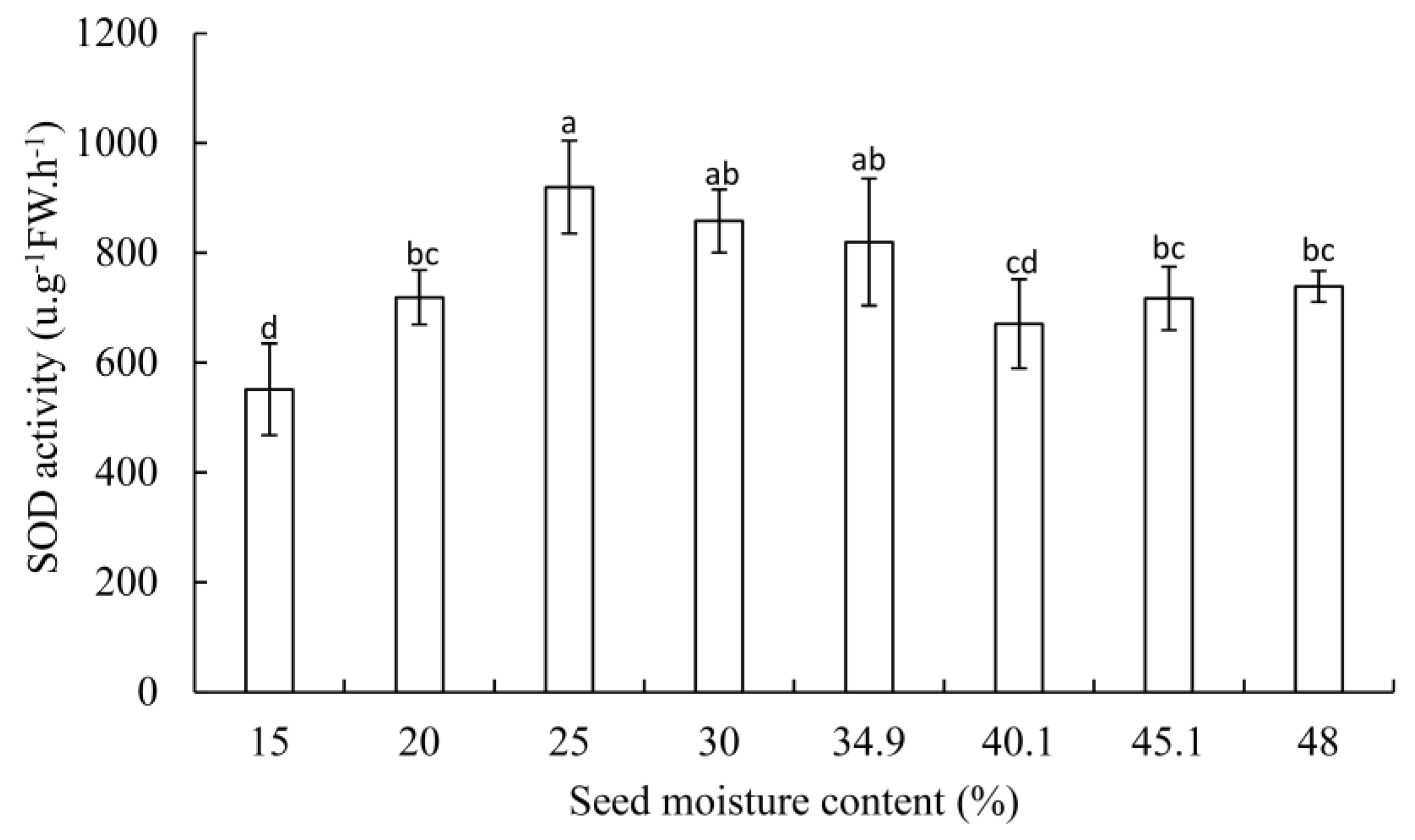
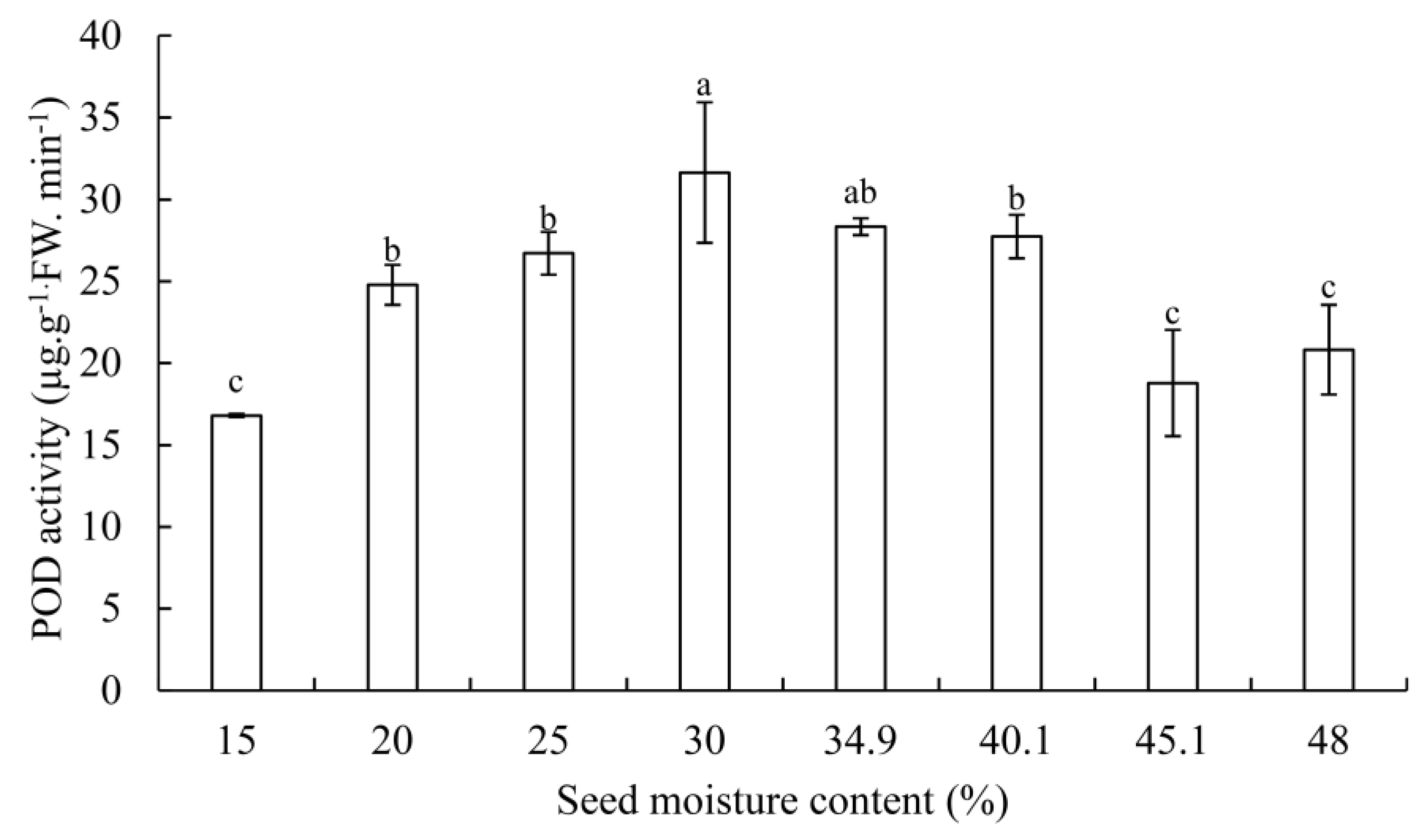
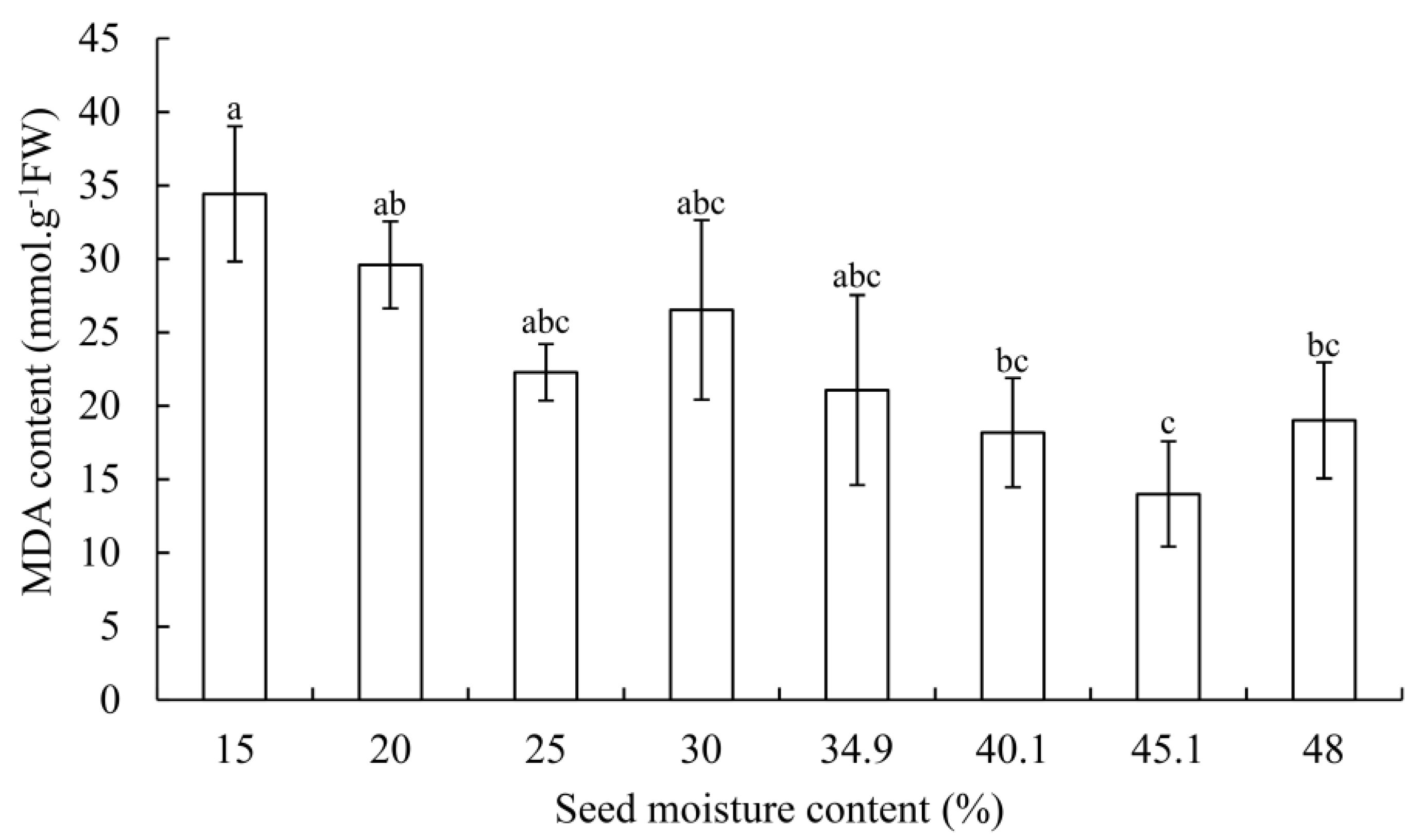
3.2. SOD, POD Activity, and MDA Contents
3.3. Correlation Analysis
4. Discussion
4.1. Changes of Germination in G. biloba Seeds during Desiccation
4.2. Changes of Electrical Conductivity in G. biloba Seeds during Desiccation
4.3. Changes of SOD and POD Activity in G. biloba Seeds during Desiccation
4.4. Changes of MDA Content in G. biloba Seeds during Desiccation
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhou, Z. Mesozoic ginkgoaleans: Phylogeny, classification and evolutionary trends. Acta Bot. Yunnanica 2003, 25, 377–396. [Google Scholar]
- Del Tredici, P. The evolution, ecology, and cultivation of Ginkgo biloba. In Ginkgo biloba; Beek, T.A.V., Ed.; Harwood Academic Publishers: Amsterdam, The Netherlands, 2000; pp. 7–23. [Google Scholar]
- Mahadevan, S.; Park, Y. Multifaceted therapeutic benefits of Ginkgo biloba L.: Chemistry, efficacy, safety, and uses. J. Food Sci. 2008, 73, R14–R19. [Google Scholar] [CrossRef] [PubMed]
- Nakao, Y.; Kawase, K.; Shiozaki, S.; Ogata, T.; Horiuchi, S. The growth of pollen and female reproductive organs of ginkgo between pollination and fertilization. J. Jpn. Soc. Hortic. Sci. 2001, 70, 21–27. [Google Scholar] [CrossRef]
- Jin, B.; Zhang, L.; Lu, Y.; Wang, D.; Jiang, X.X.; Zhang, M.; Wang, L. The mechanism of pollination drop withdrawal in Ginkgo biloba L. BMC Plant Biol. 2012, 12, 59. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Clayton, S.C.; Cui, K.; Lee, C. Developmental synchronization of male and female gametophytes in Ginkgo biloba and its neck mother cell division prior to fertilization. Physiol. Plant. 2013, 147, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Daws, M.I.; Garwood, N.C.; Pritchard, H.W. Prediction of desiccation sensitivity in seeds of woody species: A probabilistic model based on two seed traits and 104 species. Ann. Bot. 2006, 97, 667–674. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Song, S. Desiccation sensitivity of Calophyllum polyanthum seeds and factors affecting their germination. Acta Bota. Yunnanica 2003, 25, 687–692. [Google Scholar]
- Singh, B.; Kaur, P.; Singh, R.D.; Ahuja, P.S. Biology and chemistry of Ginkgo biloba. Fitoterapia 2008, 79, 401–418. [Google Scholar] [CrossRef] [PubMed]
- Tommasi, F.; Paciolla, C.; Concetta de Pinto, M.; De Gara, L. Effects of storage temperature on viability, germination and antioxidant metabolism in Ginkgo biloba L. Seeds. Plant Physiol. Biochem. 2006, 44, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Hong, T.D.; Ellis, R.H. A Protocol to Determine Seed Storage Behaviour; International Plant Genetic Resources Institute: Rome, Italy, 1996. [Google Scholar]
- Del Tredici, P. The phenology of sexual reproduction in Ginkgo biloba: Ecological and evolutionary implications. Bot. Rev. 2007, 73, 267–278. [Google Scholar] [CrossRef]
- Jing, F.; Shen, Y. Effects of female gametophyte inclusions on embryonic development during development of Ginkgo biloba seed (manuscript in preparation).
- Roberts, E.H. Predicting the storage life of seeds. Seed Sci. Technol. 1973, 1, 499–514. [Google Scholar]
- Berjak, P.; Pammenter, N. Orthodox and recalcitrant seeds. In Tropical Tree Seed Manual; Vozzo, J.V., Ed.; Agricultural Handbook 721; USDA Forest Service: Washington, DC, USA, 2002; pp. 137–147. [Google Scholar]
- Wen, B. On the compound quantitative characteristic trait of seed recalcitrance. Acta Bot. Yunnanica 2008, 30, 78–85. [Google Scholar]
- Bailly, C. Active oxygen species and antioxidants in seed biology. Seed Sci. Res. 2004, 14, 93–107. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Hoekstra, F.A.; Golovina, E.A.; Buitink, J. Mechanisms of plant desiccation tolerance. Trends Plant Sci. 2001, 6, 431–438. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef]
- Smith, M.; Berjak, P. Deteriorative changes associated with the loss of viability of stored desiccation-tolerant and desiccation-sensitive seeds. In Seed Development and Germination; Marcel Dekker: New York, NY, USA, 1995; pp. 701–746. [Google Scholar]
- Kigel, J.; Galili, G. Seed Development and Germination; Marcel Dekker: New York, NY, USA, 1995. [Google Scholar]
- Kranner, I.; Birtić, S. A modulating role for antioxidants in desiccation tolerance. Integr. Comp. Biol. 2005, 45, 734–740. [Google Scholar] [CrossRef] [PubMed]
- Bailly, C.; Audigier, C.; Ladonne, F.; Wagner, M.H.; Coste, F.; Corbineau, F.; Come, D. Changes in oligosaccharide content and antioxidant enzyme activities in developing bean seeds as related to acquisition of drying tolerance and seed quality. J. Exp. Bot. 2001, 52, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Shen, Y. Changes on physiological characteristics of Osmanthus fragrans ‘zibing ziyingui’ seeds during dehydration. Acta Hortic. Sin. 2009, 36, 279–284. [Google Scholar]
- Farrant, J.M.; Pammenter, N.W.; Berjak, P.; Walters, C. Subcellular organization and metabolic activity during the development of seeds that attain different levels of desiccation tolerance. Seed Sci. Res. 1997, 7, 135–144. [Google Scholar] [CrossRef]
- Pukacka, S.; Malec, M.; Ratajczak, E. Ros production and antioxidative system activity in embryonic axes of Quercus robur seeds under different desiccation rate conditions. Acta Physiol. Plant. 2011, 33, 2219–2227. [Google Scholar] [CrossRef]
- ISTA. International Rules for Seed Testing; The International Seed Testing Association (ISTA): Bassersdorf, Switzerland, 2005. [Google Scholar]
- Xu, L.; Pan, Y.; Yu, F. Effects of water-stress on growth and physiological changes in Pterocarya stenoptera seedlings. Sci. Hortic. 2015, 190, 11–23. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: II. Purification and quantitative relationship with water-soluble protein in seedlings. Plant Physiol. 1977, 59, 315–318. [Google Scholar] [CrossRef] [PubMed]
- Kochba, J.; Lavee, S.; Spiegelroy, P. Differences in peroxidase activity and isoenzymes in embryogenic and non-embryogenic ‘shamouti’ orange ovular callus lines. Plant Cell Physiol. 1977, 18, 463–467. [Google Scholar] [CrossRef]
- Cakmak, I.; Horst, W.J. Effect of aluminium on lipid peroxidation, superoxide dismutase, catalase, and peroxidase activities in root tips of soybean (Glycine max). Physiol. Plant. 1991, 83, 463–468. [Google Scholar] [CrossRef]
- King, M.W.; Roberts, E.H. The Storage of Recalcitrant Seeds: Achievements and Possible Approaches; International Board for Plant Genetic Resources: Rome, Italy, 1979. [Google Scholar]
- Dousseau, S.; Alvarenga, A.A.D.; Guimarães, R.M.; Lara, T.S.; Custódio, T.N.; Chaves, I.D.S. Ecofisiologia da germinação de sementes de Campomanesia pubescens. Ciência Rural 2011, 41, 1362–1368. [Google Scholar] [CrossRef]
- Yu, F.-Y.; Du, Y.; Shen, Y.-B. Physiological characteristics changes of Aesculus chinensis seeds during natural dehydration. J. For. Res. 2006, 17, 103–106. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S. Plant drought stress: Effects, mechanisms and management. In Sustainable Agriculture; Lichtfouse, E., Navarrete, M., Debaeke, P., Souchère, V., Alberola, C., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 153–188. [Google Scholar]
- Chandra, J.; Tandon, M.; Keshavkant, S. Increased rate of drying reduces metabolic inequity and critical water content in radicles of Cicer arietinum L. Physiol. Mol. Biol. Plants 2015, 21, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Dresch, D.M.; Masetto, T.E.; Scalon, S.P. Campomanesia adamantium (cambess.) O. Berg seed desiccation: Influence on vigor and nucleic acids. An. Acad. Brasil. Cie. 2015, 87, 2217–2228. [Google Scholar] [CrossRef] [PubMed]
- Berjak, P.; Pammenter, N.W. Implications of the lack of desiccation tolerance in recalcitrant seeds. Front. Plant Sci. 2013, 4, 478. [Google Scholar] [CrossRef] [PubMed]
- Veljovic-Jovanovic, S.; Kukavica, B.; Stevanovic, B.; Navari-Izzo, F. Senescence- and drought-related changes in peroxidase and superoxide dismutase isoforms in leaves of Ramonda serbica. J. Exp. Bot. 2006, 57, 1759–1768. [Google Scholar] [CrossRef] [PubMed]
- Veljovic-Jovanovic, S.; Kukavica, B.; Navari-Izzo, F. Characterization of polyphenol oxidase changes induced by desiccation of Ramonda serbica leaves. Phys. Plant 2008, 132, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.P.; Le, N.T.; Brandt, W.F.; Driouich, A.; Farrant, J.M. Towards a systems-based understanding of plant desiccation tolerance. Trends Plant Sci. 2009, 14, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Jamalomidi, M.; Gholami, M. Effect of desiccation on antioxidant enzymes activity of recalcitrant tea (Camellia sinensis L.) seeds. Int. Res. J. Appl. Basic Sci. 2013, 4, 4318–4322. [Google Scholar]
- Wang, Y.; Li, S.; He, J.; Fu, J. Changes in activity of reactive-oxygen-scavenging enzymes in recalcitrant wampee (Clausena lansium) seeds duing desiccation. Acta Phytophysiol. Sin. 2000, 27, 81–86. [Google Scholar]
- Greggains, V.; Finch-Savage, W.E.; Atherton, N.M.; Berjak, P. Viability loss and free radical processes during desiccation of recalcitrant Avicennia marina seeds. Seed Sci. Res. 2001, 11, 235–242. [Google Scholar]
- Obroucheva, N.; Sinkevich, I.; Lityagina, S. Physiological aspects of seed recalcitrance: A case study on the tree Aesculus hippocastanum. Tree Physiol. 2016, 36, 1127–1150. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Yang, L.; Ahmad, P.; Wan, X.; Hu, X. Proteomic profiling and redox status alteration of recalcitrant tea (Camellia sinensis) seed in response to desiccation. Planta 2011, 233, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Ntuli, T.M.; Finch-Savage, W.E.; Berjak, P.; Pammenter, N.W. Increased drying rate lowers the critical water content for survival in embryonic axes of english oak (Quercus robur L.) seeds. J. Integr. Plant Biol. 2011, 53, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Song, S.-Q.; Berjak, P.; Pammenter, N. Desiccation sensitivity of Trichilia dregeana axes and antioxidant role of ascorbic acid. Acta Bot. Sin. 2004, 46, 803–810. [Google Scholar]
- Varghese, B.; Naithani, S. Desiccation-induced changes in lipid peroxidation, superoxide level and antioxidant enzymes activity in neem (Azadirachta indica A. Juss) seeds. Acta Physiol. Plant. 2002, 24, 79–87. [Google Scholar] [CrossRef]
- Li, Y.; Ma, Y. Effects of drying at different rates on desiccation sensitivity and membrane lipid peroxidation of Pachira macrocarpa seeds. Chin. J. Trop. Crops 2008, 29, 738–743. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Sum of Squares | df | Mean Square | F | p | |
|---|---|---|---|---|---|---|
| Germination percentage | Between Groups | 2.359 | 7 | 0.337 | 116.831 | <0.001 |
| Within Groups | 0.046 | 16 | 0.003 | |||
| Total | 2.405 | 23 | ||||
| Electrical conductivity | Between Groups | 0.269 | 7 | 0.038 | 39.305 | <0.001 |
| Within Groups | 0.016 | 16 | 0.001 | |||
| Total | 0.284 | 23 | ||||
| SOD activity | Between Groups | 279,608.451 | 7 | 39,944.064 | 7.252 | 0.001 |
| Within Groups | 88,130.424 | 16 | 5508.152 | |||
| Total | 367,738.876 | 23 | ||||
| POD activity | Between Groups | 553.311 | 7 | 79.044 | 15.173 | <0.001 |
| Within Groups | 83.350 | 16 | 5.209 | |||
| Total | 636.661 | 23 | ||||
| MDA content | Between Groups | 996.525 | 7 | 142.361 | 3.085 | 0.029 |
| Within Groups | 738.356 | 16 | 46.147 | |||
| Total | 1734.880 | 23 | ||||
| Physiological Indicators | MC | SOD Activity | MDA Content | POD Activity | Electrical Conductivity | Germination Percentage |
|---|---|---|---|---|---|---|
| MC | 1 | |||||
| SOD activity | 0.118 | 1 | ||||
| MDA content | −0.619 * | −0.373 | 1 | |||
| POD activity | −0.002 | 0.595 * | −0.065 | 1 | ||
| Electrical conductivity | −0.745 * | 0.092 | 0.421 * | 0.439 * | 1 | |
| Germination percentage | 0.910 * | −0.107 | −0.545 * | −0.364 | −0.864 * | 1 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, J.; Shen, Y.; Shi, F.; Li, C. Changes in Seed Germination Ability, Lipid Peroxidation and Antioxidant Enzyme Activities of Ginkgo biloba Seed during Desiccation. Forests 2017, 8, 286. https://doi.org/10.3390/f8080286
Feng J, Shen Y, Shi F, Li C. Changes in Seed Germination Ability, Lipid Peroxidation and Antioxidant Enzyme Activities of Ginkgo biloba Seed during Desiccation. Forests. 2017; 8(8):286. https://doi.org/10.3390/f8080286
Chicago/Turabian StyleFeng, Jing, Yongbao Shen, Fenghou Shi, and Chengzhong Li. 2017. "Changes in Seed Germination Ability, Lipid Peroxidation and Antioxidant Enzyme Activities of Ginkgo biloba Seed during Desiccation" Forests 8, no. 8: 286. https://doi.org/10.3390/f8080286
APA StyleFeng, J., Shen, Y., Shi, F., & Li, C. (2017). Changes in Seed Germination Ability, Lipid Peroxidation and Antioxidant Enzyme Activities of Ginkgo biloba Seed during Desiccation. Forests, 8(8), 286. https://doi.org/10.3390/f8080286