
Effects of Fire Frequency and Soil Temperature on Soil CO2 Efflux Rates in Old-Field Pine-Grassland Forests


1
School of Forest Resources and Conservation, University of Florida, Gainesville, FL 32611, USA
2
College of Natural Resources, Natural Resources and Society, University of Idaho, Moscow, ID 83844, USA
3
Tall Timbers Research Station and Land Conservancy, Leon County, FL 32308, USA
*
Author to whom correspondence should be addressed.
Forests 2017, 8(8), 274; https://doi.org/10.3390/f8080274
Submission received: 28 June 2017
/
Revised: 25 July 2017
/
Accepted: 28 July 2017
/
Published: 30 July 2017
Abstract
:Soil CO2 efflux (Rs) is a significant source of carbon dioxide from soils to the atmosphere and is a critical component of total ecosystem carbon budgets. Prescribed fire is one of the most prevalent forest management tools employed in the southeastern USA. This study investigated the influence of prescribed fire on Rs rates in old-field pine-grassland forests in north Florida, USA, that had been managed with prescribed fire annually and biennially for over 40 years, or left unburned for approximately the same period. Monthly measurements were taken of Rs, soil temperature (Ts), and soil moisture from August 2009 to May 2011. Results showed that sites managed with annual and biennial dormant season prescribed fire had significantly lower monthly mean Rs rates and estimated annual soil carbon fluxes than sites where fire had been excluded. While Ts explained a significant amount of the temporal variations in Rs, it did not explain the differences in Rs among prescribed fire treatments. Our results provide new insight into the effects of prescribed fire and fire exclusion on soil carbon fluxes, and suggest that future methods to model ecosystem carbon budgets should incorporate not only current vegetative conditions, but also prescribed fire management activities.
1. Introduction
The influence of long-term forest management practices on soil CO2 efflux (Rs) rates plays a key role in determining total ecosystem carbon budgets [1,2,3]. It has been estimated that soil CO2 efflux represents one of the largest global terrestrial fluxes of carbon to the atmosphere, with the total annual Rs flux (75 Pg C year−1) an order of magnitude greater than current annual anthropogenic carbon (C) emissions from fossil fuel combustion (6 Pg C year−1) [4]. Soil CO2 efflux is a function of various interrelated biogeochemical factors that govern the production of autotrophic soil CO2 efflux by plant roots and associated mycorrhizal fungi (Ra) and heterotrophic soil CO2 efflux by soil micro and macro biota(Rh) [3,4,5,6].
Fire can influence Rs rates by differentially impacting the Rh and Ra sources of CO2 [4,7]. For example, short-term autotrophic production of CO2 can be reduced by fire due to both aboveground and belowground plant mortality and injury. While the long-term impacts of fire on Ra are variable, Ra has been shown in some cases to increase with time since fire as vegetation recovers following disturbance [8]. In many cases, the burning of vegetation and surface fuels reallocates nutrient resources via incomplete combustion and subsequent deposition of ash, char, and other residues [8,9]. In the short-term period following the deposition of those residues, both plants and soil microbes may respond positively to the availability of such resources, subsequently increasing Rs rates. Long-term prescribed fire management regimes may also impact Rh sources of CO2, by influencing vegetation composition, structure and associated litter and duff quality, production, and accumulation rates [10] which in turn have been shown to influence soil microbial populations and metabolic activity [11]. Fire can also reduce Rh by killing soil microbes through heating of litter and duff layers and upper soil horizons [4]. Previous studies in multiple ecosystems have shown that both fire and forest management can influence Rs rates, soil carbon pools, and various coupled biogeochemical processes [12,13,14,15,16]. For example, in a study of a mixed conifer forest in California, USA, Ryu et al. [17] found that prescribed fire reduced Rs rates while simultaneously altering soil conditions that would otherwise be associated with increased Rs rates. However, two studies that investigated the influence of prescribed fire in a different mixed conifer forest in California and an upload oak (Quercus spp. L) forest in Missouri, USA, found that while prescribed burning significantly altered forest floor conditions, there was no clear effect on Rs rates [18]. The contrasting results reported by these studies demonstrate the complex nature of predicting the influence of prescribed fire on Rs. While many factors influencing Rs rates have been identified, much remains to be determined regarding the effects of specific forest and land management practices on overall Rs rates [2,3,4], especially where management has been long-term rather than experimental.
Frequent prescribed fire is one of the dominant tools for forest management in the pine-grassland forests of the southeastern USA [19,20]. Many of the current pine-grassland forests in the region are old-field forest assemblages that exist on former agricultural lands estimated to cover up to 21 million ha across the southeastern USA [21]. Across many of these forests, frequent (1–3 year return interval) prescribed fire is used to perpetuate native species assemblages, reduce the risk of destructive wildfires, and promote and restore wildlife habitat [22]. Although less frequently cited, the impacts of prescribed fire regimes on soil carbon sequestration are likely to increase in importance as important commodity trade partners adopt carbon credit exchanges (e.g., Canada). The long term consequences of varying fire return intervals for carbon cycling in pine grasslands can inform fire management decisions and projections of carbon sequestration capacity.
This study sought to investigate the impacts of fire regime, specifically annual burning, biennial burning, and prolonged fire exclusion, on Rs rates in old-field pine-grassland forests. In addition, this study sought to interpret the potential response of Rs to biotic and abiotic factors, including soil temperature, soil moisture, forest stand characteristics, and soil physical properties and chemistry. The intent of this research is to build upon our understanding of the effects of frequent fire on carbon dynamics and sequestration in pine-grasslands and similar woodland and savanna communities worldwide. Studies such as this also provide insight into the response of ecosystem carbon dynamics to forecasted changes in temperature and moisture regimes due to global climate change.
2. Materials and Methods
2.1. Study Site
The study was conducted on Tall Timbers Research Station (TTRS) in Leon County, FL, USA, approximately 30 km from the cities of Tallahassee, Florida (to the south) and Thomasville, Georgia (to the north) (30°39′ N, 84°12′ W; Figure 1). The study utilized research plots established on old field land for long-term study of fire regimes. The Stoddard Fire Plots, which are 0.2 ha plots established in 1960, have each been managed with a constant fire return interval until the present [10,23,24]. The study also utilized a 9.2 ha plot named NB66 that had been fire-excluded since 1966 [24]. Prior to establishment of the plots, the areas had been burned at 1–2 year intervals since agricultural abandonment which occurred from the late 1800s to the 1920s. Soils within the sites were heavily cultivated for growing corn and cotton from the 1820s until abandonment with subsequent understory and overstory vegetation assemblages highly influenced by past agricultural practices [23] but largely representing a subset of species occurring in native pine communities of the region [10,25]. Soils were classified as fine-loamy, kaolinitic, thermic Typic Kandiudults of the Orangeburg and Faceville series (Natural Resource Conservation Service (NRC) Soil Survey Geographic Database (SSURGO)).
For this study, sampling took place within three annually burned (1YR), three biennially burned (2YR), and two fire-excluded (UB) Stoddard Fire Plots, and an additional 0.2 ha fire-excluded study area (UB) was established within the Tall Timbers NB66 study site, making three replicates of each fire regime [10,23]. Replicate sets of plots each containing the three fire regimes were grouped within soil units, such that two sets were within Faceville (Fine, kaolinitic, thermic Typic Kandiudult) soil units and one was in an Orangeburg (Fine-loamy, kaolinitic, thermic Typic Kandiudult) soil unit. The study sites were located approximately 60 m a.s.l. Average annual precipitation was 137 cm with the majority falling during the summer months of June, July and August (National Climate Data Center 2009, Thomasville, GA, USA). Mean maximum and minimum temperatures for January and July for the area from long-term records (1971–2000) are 16.8 °C and 4.6 °C for January and 33 °C and 21.8 °C for July (National Climate Data Center 2009, Thomasville, GA, USA).
The overstory of the 1YR and 2YR burned plots consisted of a mixture of naturally regenerated shortleaf pine (Pinus echinata P. Mill), loblolly pine (P. taeda L.), and, to a lesser extent, longleaf pine (P. palustris P. Mill). The understory was composed of a mixture of grasses, forbs, and broadleaf woody plants which are typically topkilled by prescribed fire and then resprout [10,19,23,26]. The unburned plots, due to the prolonged fire exclusion, contained a closed midstory and overstory canopy of broadleaf deciduous trees of species including but not limited to: water oak (Q. nigra L.), laurel oak (Q. laurifolia Mich.), sweetgum (Liquidambar styraciflua L.), black cherry (Prunus serotina Ehrh.), and flowering dogwood (Cornus florida L.), in addition to mature pine trees.
2.2. Sampling
The Stoddard Fire Plots combined with NB66 representing each of three prescribed fire regimes (1YR, 2YR, UB) were grouped into blocks replicated three times for a total of nine plots. Sampling for this study took place within subplots randomly located within each of the Stoddard Fire Plots and NB66. To account for spatial variability within the subplots, measurements were taken at nine locations within in a square grid arrangement with 5 m separation, following Kobziar [16].
Rs (μmol CO2 m−2·s−1) was sampled at each subplot using a LI-COR Biosciences LI-8100 automated soil CO2 sampling instrument with a 20 cm diameter survey chamber (LI-COR Biosciences Inc., Lincoln, NE, USA). The survey chamber fit onto 20 cm diameter × 10 cm height PVC collars inserted 8 cm into the ground. Sampling of Rs for all plots occurred monthly. During each monthly visit, all measurements were taken within two days. Over the course of study, any vegetative growth within the sample collars was clipped and removed prior to Rs measurement. The PVC collars were installed in June and July 2009 and sampling began in August. The PVC collars and soil surfaces contained within them were protected from fire during burns in March 2010 (1YR plots) and March 2011 (1YR and 2YR plots) by placing slightly larger diameter sheet-metal cylinders around them. As such, the measurements were not expected to respond to direct impacts of prescribed fire, e.g., changes to soil chemistry or injury of microorganisms near the soil surface, but rather general environmental conditions corresponding to the respective fire regime. Sampling consisted of a 120 s measurement initiated by a 15 s “dead-band” during which gases were allowed time to mix within the chamber. During Rs measurements, adjacent to each PVC collar, soil temperature (Ts) (°C) was measured at 10 cm depth using an Omega 8831 type E T-Handle temperature probe, and soil moisture content (Ms) (m3/m3) was measured at 5 cm depth using a Decagon Systems EC-5 soil moisture probe mounted on the LI-8100 (Omega Inc., Stamford, CT, USA; Decagon Systems Inc., Pullman, WA, USA). From August of 2009 until February 2010, each monthly sampling period involved measurement of Rs, Ts, and Ms eight times per day in order to assess for diurnal variability. An analysis of the preliminary results found that fewer daily measurements would be sufficient to capture the variability, so daily measurements were scaled back to three times per day (morning, mid-day and late afternoon-early evening) from March 2010 until the end of the study in May 2011. Equipment problems resulted in no field measurements being taken during the month of October 2010, but otherwise there were only few interruptions due to equipment problems or severe thunderstorms. The resulting dataset for the entire twenty-one-month study totaled 7566 Rs measurements. Some strong outliers in Rs, Ts, and Ms were attributed to measurement or equipment error and were isolated and excluded from the analyses.
Within a 15 m radius circular plot (0.07 ha) centered on the middle PVC collar in each subplot, basal area (BA) (m2·ha−1), pine basal area (PBA) (m2·ha−1), hardwood (broadleaf) tree basal area (HW BA) (m2·ha−1), and stand density (TPH) (trees ha−1) were measured (only counting trees diameter at breast height > 10 cm). Forest stand characteristics were assessed once per subplot in the winter or spring of 2011. Soil total carbon and total nitrogen were measured in January 2013 (Table 1). Subplot level mean litter depth (Litter) (cm), duff depth (Duff) (cm) and total litter and duff depth (DL) (cm) were recorded as the average of three measurements taken from random locations with 30 cm of each PVC collar. Soil was sampled using a 2 cm diameter soil corer to 10 cm depth at 30 locations spread throughout each plot then combined, homogenized, and split to obtain subsamples for analysis. Samples were analyzed at Auburn University to determine percent total carbon and nitrogen with a TruSpec CN analyzer (Leco Corp., St. Joseph, MI, USA) using the dry combustion method, and mg−1·kg−1 of phosphorus, calcium, magnesium, and potassium were determined with an iCAP analyzer (Thermo Fisher Scientific Inc., Waltham, MA, USA) using Mehlich 3 extraction. Bulk density to 10 cm depth was sampled at five locations within each plot using a 4.5 cm diameter soil sampler (Eijkelkamp Corp., Giesbeek, The Netherlands). Bulk density measurements were used to convert soil chemistry measurements to mass per unit area (kg−1 or Mg−1 per·ha−1).
2.3. Analysis
Analyses sought to test for significant differences among the three fire regimes with regard to Rs, Ts, Ms and forest environmental conditions measured in the field. For each month, daily measurements per PVC collar were averaged, and the nine PVC collar means were averaged to produce a subplot-level mean value for each month. Repeated measures analysis of variance (ANOVA) was used to test for differences in these monthly means among the three fire regimes (n = 3 per fire regime) for Rs, Ts, and Ms over the twenty-one monthly sampling periods between August 2009 and May 2011. Significant treatment effects were identified at p-value < 0.05. To assess for differences in forest environmental conditions among the three fire regimes, one-way ANOVA tests were applied to each variable separately. Where significant differences were identified, differences were further investigated using Tukey’s HSD test. Additionally, linear and nonlinear regression models (Equations (1) and (2)) were developed to predict the monthly response of Rs to Ts as well as Rs to Ms in separate equations within each fire regime and within each season. Non-linear models of the relationships between Rs rates and Ts per fire regime were explored using an exponential equation (Equation (2)) frequently used to describe the response of Rs rates to soil temperature [15,18,27,28]. The β1 estimates developed using Equation (2) were used to estimate Q10 (Equation (3)), which describes the response of Rs to a 10° C change in soil temperature [4,15,27]. All statistical analyses were performed using JMP 9.0 (SAS Institute, Cary, NC, USA).
where MTemp was air temperature, β0, and β1 were coefficients estimated through regression analysis.
Total monthly and annual soil carbon emissions per fire regime were estimated using the nonlinear regression models of Rs responses (Equation (2)) to changes in ambient temperature (MTemp) following Samuelson et al. [28]. Twenty-four hour 2 m elevation ambient temperature measurements recorded hourly from 1 August 2009–31 July 2010 at the Quincy, FL Automated Weather Network site located approximately 30 km from the study sites were used as the MTemp input to estimate hourly Rs rates. The estimated Rs rates (μmol CO2 m−2·s−1) were then converted to hourly soil carbon fluxes (g C m−2·h−1) which were then summed to estimate monthly and annual soil carbon fluxes (Mg C ha−1·year−1).
3. Results
3.1. Differences in Rs, Ms, Ts, and Vegetation among Fire Regimes
Mean Rs rates varied significantly between fire regimes (p = 0.0007), with the highest Rs rates typically in the UB treatment and the lowest typically in the 1YR treatment (Figure 2; Table 2 and Table 3). The treatment effect of fire regime on Rs rates varied monthly (treatment × time p < 0.0001), with the greatest difference between fire regimes observed during the summer months and the least during the winter months (Table 3). Across all treatments, Rs ranged from 0–11.98 μmol CO2 m−2·s−1 during the study period, with the highest Rs rates during the warmer months and the lowest rates during the cooler months (Figure 2).
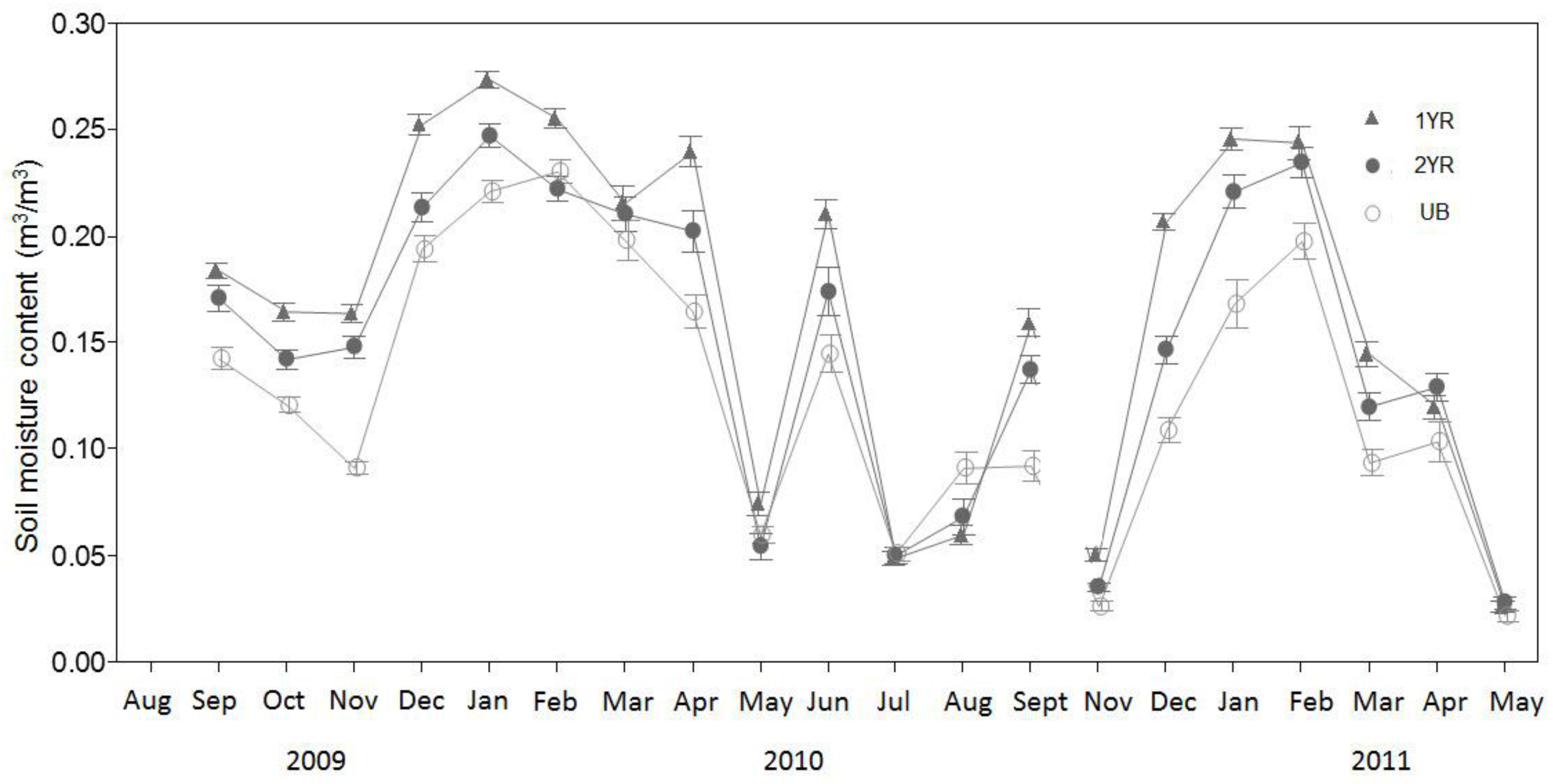
Although soil temperature did not differ significantly among the fire regimes (p = 0.1007), the lowest temperatures were in the UB sites and the highest in the 1YR sites (Figure 3). A distinct seasonal Ts trend among the fire regimes was observed, with the 1YR sites recording the highest mean Ts in the spring through fall seasons and the UB sites recording the highest mean Ts in the winter seasons. Soil moisture (Ms) did not differ significantly among the fire regimes (p = 0.11), although the lowest soil moisture contents were observed in the UB sites and the highest observed in the 1YR sites (Figure 4; Table 2).
Forest stand characteristics and litter and duff depths varied significantly among fire regimes (Table 4A). Patterns were related to higher tree stocking and duff and litter accumulation with lower occurrence of fire. Duff depth was over an order of magnitude higher in the UB sites than the 1YR sites (Table 4A). Soil bulk density was significantly higher in the 1YR and 2YR return interval treatments and percent total carbon was higher in the 2YR treatment than in the unburned treatment (Table 4B). Values for nitrogen, phosphorus, calcium, magnesium, potassium, and pH all showed a decreasing trend from the 1YR to unburned treatments, although results were significant only for magnesium (Table 4B).
3.2. Drivers of Rs within Treatment
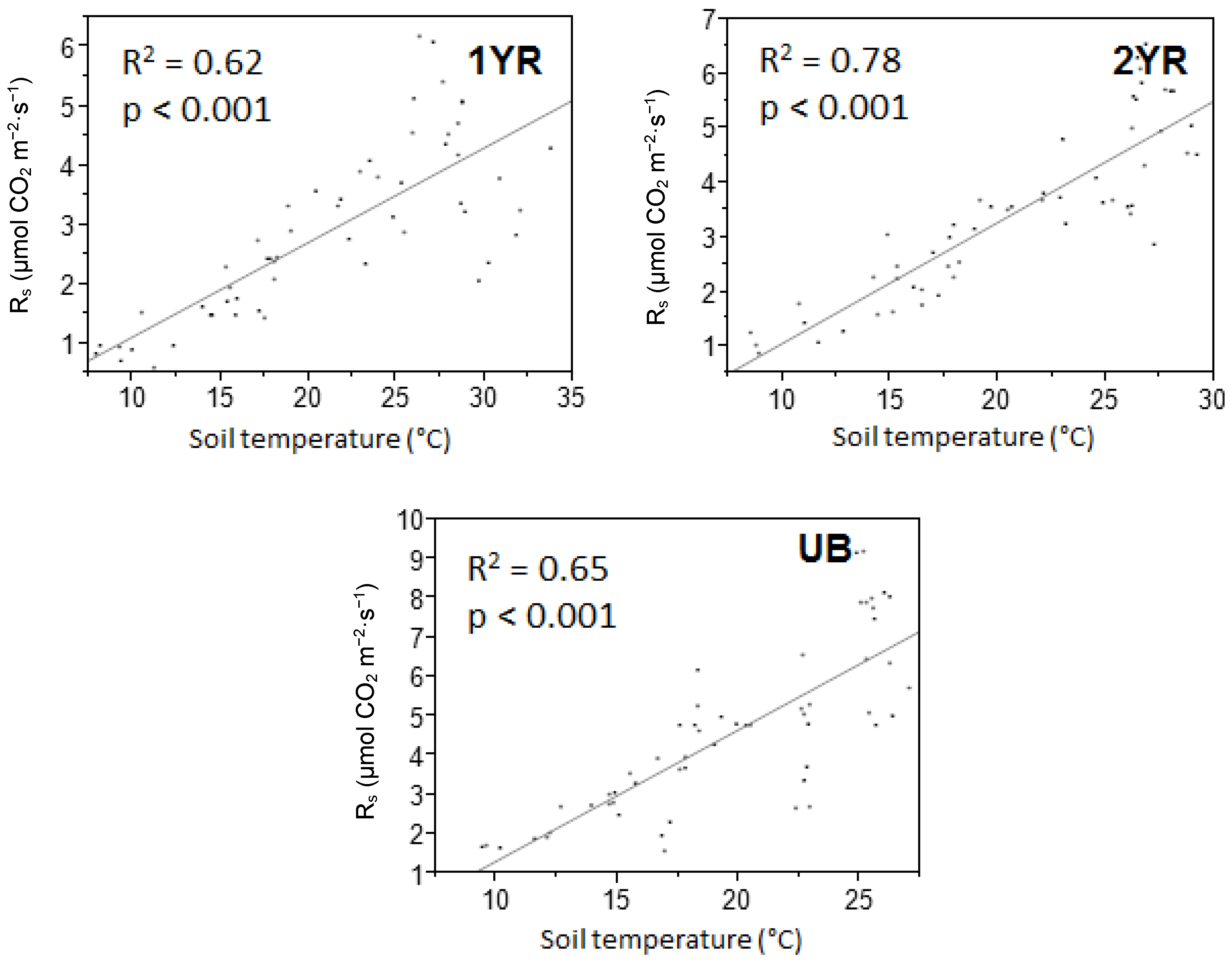
Linear regressions indicated monthly mean Ts significantly predicted monthly mean Rs within each fire regime (R2 > 0.62; Figure 5). In contrast, Ms did not predict Rs (R2 < 0.14) [29]. Nonlinear exponential models also used to predict Rs from Ts and Ms by fire regime showed similar results as the linear models, in which case Ms was also not significant and not reported (Table 5). Model coefficients β0 and β1 from the nonlinear models were similar to those reported by Samuelson et al. [28] and Kobziar and Stephens [15]. Q10 values ranged from 1.65 to 2.16 and differed among fire regimes (Table 5). The UB sites had the greatest Rs sensitivity to changes in soil temperature (Q10 = 2.16), while the 1YR sites were the least sensitive to soil temperature changes (Q10 = 1.65).
3.3. Seasonal Variation in Rs and Ts Relationships
In the simple linear models and non-linear models predicting mean monthly Rs from Ts within each fire regime showed that the relationship was strongest during the fall and winter (R2 = 0.69–0.90) and weakest during the spring and summer (R2 = 0.11–0.63) in all fire regimes (Table 6 and Table 7). Q10 also varied seasonally, with the lowest Q10 values observed during summer and the highest Q10 values observed during the winter in all fire regimes (Table 7).
3.4. Total Carbon Emissions
Estimated total monthly soil carbon emissions were consistently higher in the unburned UB sites than the frequently burned 1YR and 2YR sites. Similarly, estimated total annual soil carbon emissions per fire regime showed the highest soil carbon efflux in the UB sites (16.88 Mg ha−1·year−1) and the lowest in the 1YR (10.69 Mg ha−1·year−1) and 2YR (12.68 Mg ha−1·year−1) sites.
4. Discussion
Results of our study show that Rs was consistently higher in long fire-excluded pine-grasslands than in frequently burned pine-grasslands, while there was little difference between annually and biennially burned communities. Although determining specific mechanisms for the observed pattern was beyond the scope of this study, there is evidence that Rs responded to environmental conditions influenced by fire regime effects on forest structure.
In the UB sites, aboveground living tree biomass (≈150 Mg ha−1) was much greater than in the 1YR and 2YR sites (≈50 Mg ha−1 and ≈80 Mg ha−1, respectively), suggesting that the presumably corresponding higher tree root biomass in the unburned plots may have influenced Rs. Other studies investigating Rs rates across stand age and biomass gradients have found mean Rs rates to be higher in older stands with greater aboveground biomass [30,31]. In a trenching and exclusion experiment along a chronosequence of temperate forests in China, Luan et al. [32] found that Rs rates were significantly correlated with site basal area (R2 = 0.59, p < 0.05).
Higher soil respiration rates in the unburned sites may have also been influenced by tree species composition and associated litter quality, specifically dominance by deciduous broadleaf trees which were essentially absent in the burned plots. Soil CO2 efflux rates have been shown in other studies to be lower in coniferous forests than in broad-leafed forests of the same soil type [5]. In a study of Rs rates in a mixed conifer–deciduous forest in Belgium, Yuste et al. [33] found that mean Rs rates were lower under conifer tree canopies than under deciduous canopies, with total estimated annual carbon flux approximately 50% greater in the deciduous sites (8.8 ± 2.2 Mg C ha−1·year−1) than in the coniferous sites (4.8 ± 0.7 Mg C ha−1·year−1). In their review, Raich and Tufekciogul [5] suggested that the observed differences in Rs between coniferous and deciduous forests may have been driven by forest litter production and quality, carbon allocation, and autotrophic contributions to total Rs.
In our study, litter and duff depths were highest in the UB sites, reflecting higher tree stocking and lack of fire, in turn possibly contributing to higher Rs rates in the UB sites. Other studies have reported that frequent fire greatly reduces the accumulation of litter and duff [34]. The lower soil carbon levels and lowest Rs in the 1YR plots compared to the 2YR plots likely reflect limited inputs of organic matter because of annual burning. The 2YR plots in turn are assumed to have lower organic matter inputs to the soil than the UB plots because frequent fire, yet the total carbon was higher in the 2YR plots, suggesting lower rates of decomposition than the UB plots consistent with the lower Rs measured. It appears that some characteristic of the frequently burned environment suppressed soil respiration, whether through its effect on Ra or Rh. Increased bulk density associated with burning, found in this and other studies [35,36,37] has been attributed to infiltration of micropore space by ash and char, which might negatively influence microbial activity. Similar patterns of higher total carbon in the mineral soil of burned than unburned areas have been seen in many long-term fire experiments in the southeastern USA [36,38,39,40,41].
The Rs rates observed in this study resulted in lower estimated total annual soil carbon emissions in the 1YR (37%) and 2YR (25%) sites relative to the UB sites. The estimated monthly carbon emissions for all sites were similar to those reported by Samuelson et al. [28] for an unburned loblolly pine plantation in southwestern Georgia, USA, while the estimated total annual soil carbon emissions reported in our study were similar to those (14.10 Mg ha−1·year−1) reported by Maier and Kress [42] for an unburned loblolly pine plantation, but greater than those (7.78–9.66 Mg ha−1·year−1) reported by the Samuelson et al. [28] study.
Temporal variability in Rs rates was explained more by soil temperature than any other recorded parameter. These results were consistent with other studies in southeastern USA forest systems in which Rs rates were found to have strong correlations with soil temperature [28,42,43]. The Rs correlation with Ts reported in this study (R2 = 0.62–0.78 in linear models and R2 = 0.60–0.80 in nonlinear models) were higher than those reported by Samuelson et al. [28] for a Georgia, USA loblolly pine plantation (R2 = 0.38–0.56), much higher than those reported by Templeton et al. [43] across a range of loblolly pine plantations in 11 southeastern USA states (R2 = 0.45), and similar to those reported for a loblolly pine plantation in North Carolina, USA (R2 = 0.70) [42]. Caution must be used in comparing correlations among studies, as variations in the modeled Rs and Ts relationship may be influenced by statistical modeling techniques regardless of the biogeochemical couplings observed in the field. While Ts explained the majority of the temporal variability in Rs rates, a lack of significant differences in Ts among treatments suggests that temperature was not the key factor contributing to the differences in Rs rates among the treatments.
Over the course of this study’s sampling period, relationships between Rs and Ts varied seasonally, with stronger relationships during the winter seasons. This pattern was evident in the R2 values of the seasonal linear and nonlinear models as well as in the seasonal Q10 values. Distinct seasonal variations in Q10 values have been noted by others [44,45] and such variations have potential implications for modeling Rs values based on temperature measurements. Yuste et al. [45] suggested that while annual Q10 values may sufficiently model annual Rs rates, seasonal or shorter-term estimates of Rs should be based on season specific Q10 functions that capture the seasonal variation in Rs temperature response, as observed in our study.
The seasonal variation in the relationship between Rs and Ts may be the result of phenology-related shifts in the relative contributions of Ra and Rh to Rs. Previous research in partitioning the sources of Rs have shown that during periods of aboveground vegetative growth, Ra contributions to Rs can increase relative to Rh, as plants allocate recent C photosynthate belowground, driving higher root maintenance, root growth, and mycorrihizal fungal respiration rates [6,46]. In addition, other studies have shown that during periods of aboveground vegetative growth, the Ts and Rs relationship weakens as other variables such as soil moisture and available photosynthetically active radiation (PAR) become important in governing belowground C allocation by plants [47,48,49]. Similar to our findings, Fenn et al. [50] found that soil temperature explained less of the variation in Rs during the summer than during the spring in a multi-season study of Rs rates in a woodland in Oxfordshire, UK.
5. Conclusions
In this study comparing long-term fire disturbance intervals, soil temperature explained the majority of the temporal variation in Rs rates, but neither temperature nor moisture explained the differences in Rs observed among the fire frequencies. As such, we suggest that future efforts to model soil carbon fluxes in otherwise similar burned and unburned forests should account for the impacts of both seasonal variability and prescribed fire management regime on the relationships between Rs and Ts. Differences in CO2 efflux rates, as well as soil chemistry and bulk density, reflect the long-term impact of contrasting fire disturbance regimes, as well as the biophysical consequences of those regimes. Lower total annual soil C emissions, combined with lower rates of soil C efflux and higher stored soil C, suggest that even biennially burned sites are effective at sequestering soil carbon.
Acknowledgments
This research was supported by the Kobziar Fire Ecology Lab and the the School of Forest Resources and Conservation, University of Florida, Gainesville, FL, USA. Additional support was provided by the Joint Fire Science Program Graduate Research Innovation grant (11-3-1-21). The authors thank two anonymous reviewers who provided valuable feedback on earlier versions of this paper. Special thanks for prescribed burning, soil collection, and literature assistance from A. Reid, J. Patterson, and C. Armstrong of the Fire Ecology Program and the Parker-Williams Library at the Tall Timbers Research Station and Land Conservancy. Soil was collected by James Patterson. Logistics and fieldwork were supported by A. Reid and. A. Kattan.
Author Contributions
David R. Godwin, Leda N. Kobziar and Kevin M. Robertson conceived and designed the experiments; David R. Godwin performed the experiments and analyzed the data; Kevin M. Robertson provided soil chemical and physical measurements and supervised prescribed burns; David R. Godwin, Leda N. Kobziar and Kevin M. Robertson wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Raich, J.W.; Schlesinger, W.H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus B 1992, 44, 81–99. [Google Scholar] [CrossRef]
- Schlesinger, W.; Andrews, J. Soil respiration and the global carbon cycle. Biogeochemistry 2000, 48, 7–20. [Google Scholar] [CrossRef]
- Ryan, M.G.; Law, B.E. Interpreting, measuring, and modeling soil respiration. Biogeochemistry 2005, 73, 3–27. [Google Scholar] [CrossRef]
- Luo, Y.; Zhou, X. Soil Respiration and the Environment; Academic Press: Burlington, MA, USA, 2006. [Google Scholar]
- Raich, J.W.; Tufekciogul, A. Vegetation and soil respiration: Correlations and controls. Biogeochemistry 2000, 48, 71–90. [Google Scholar] [CrossRef]
- Subke, J.; Inglima, I.; Cotrufo, M.F. Trends and methodological impacts in soil CO2 efflux partitioning: A meta-analytical review. Glob. Chang. Biol. 2006, 12, 921–943. [Google Scholar] [CrossRef]
- Neary, D.; Klopatek, C.; DeBano, L.; Folliott, P. Fire effects on belowground sustainability: A review and synthesis. For. Ecol. Manag. 1999, 122, 51–71. [Google Scholar] [CrossRef]
- O’Neill, K.P.; Richter, D.; Kasischke, E. Succession-driven changes in soil respiration following fire in Black Spruce stands of interior. Alsk. Biogeochem. 2006, 80, 1–20. [Google Scholar] [CrossRef]
- Medvedeff, C. The Effect of an Extreme Restoration Approach on Microbial Carbon Cycling in a Restored Subtropical Wetland. Ph.D. Thesis, University of Florida, Gainesville, FL, USA, 2012. [Google Scholar]
- Glitzenstein, J.; Streng, D.; Masters, R.E.; Robertson, K.M.; Hermann, S.M. Fire-frequency effects on vegetation in north Florida pinelands: Another look at the long-term Stoddard Fire Research Plots at Tall Timbers Research Station. For. Ecol. Manag. 2012, 264, 197–209. [Google Scholar] [CrossRef]
- Sulzman, E.W.; Brant, J.B.; Bowden, R.D.; Lajtha, K.S.; Mar, S.R.; Brant, B. Contribution of aboveground litter, belowground litter, and rhizosphere respiration to total soil CO2 efflux in an old growth coniferous forest. Biogeochemistry 2012, 73, 231–256. [Google Scholar] [CrossRef]
- Johnson, D. Effects of forest management on soil C and N storage: Meta-analysis. For. Ecol. Manag. 2001, 140, 227–238. [Google Scholar] [CrossRef]
- Johnson, D.W.; Knoepp, J.D.; Swank, W.T.; Shan, J.; Morris, L.; Van Lear, D.H.; Kapeluck, P.R. Effects of forest management on soil carbon: Results of some long-term resampling studies. Eviron. Pollut. 2002, 116, S201–S208. [Google Scholar] [CrossRef]
- Certini, G. Effects of fire on properties of forest soils: A review. Acta Oecol. 2005, 143, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kobziar, L.N.; Stephens, S.L. The effects of fuels treatments on soil carbon respiration in a Sierra Nevada pine plantation. Agric. For. Meteorol. 2006, 141, 161–178. [Google Scholar] [CrossRef]
- Kobziar, L.N. The role of environmental factors and tree injuries in soil carbon respiration response to fire and fuels treatments in pine plantations. Biogeochemistry 2007, 84, 191–206. [Google Scholar] [CrossRef]
- Ryu, S.; Concilio, A.; Chen, J.; North, M.; Ma, S. Prescribed burning and mechanical thinning effects on belowground conditions and soil respiration in a mixed-conifer forest, California. For. Ecol. Manag. 2009, 257, 1324–1332. [Google Scholar] [CrossRef]
- Concilio, A.; Ma, S.; Li, Q.; LeMoine, J.; Chen, J.; North, M.; Moorhead, D.; Jensen, R. Soil respiration in response to prescribed burning and thinning in mixed-conifer and hardwood forests. Can. J. For. Res. 2005, 35, 1581–1591. [Google Scholar] [CrossRef]
- Engstrom, R.T.; Palmer, W.E. Two species in one ecosystem: Management of Northern bobwhite and Red-cockaded woodpecker in the Red Hills. In Bird Conservation Implementation and Integration in the Americas, Proceedings of the 3rd International Partners in Flight Conference, Asilomar, CL, USA, 20–24 March 2002; Ralph, C.J., Rich, T.D., Eds.; USDA Forest Service Pacific Southwest Research Station: Albany, CL, USA, 2005; pp. 1151–1157. [Google Scholar]
- Way, A.G. The Stoddard-Neel method: Forestry beyond one generation. For. Hist. Today 2006, 16–23. [Google Scholar]
- Frost, C. Four centuries of changing landscape patterns in the longleaf pine ecosystem. In Proceedings of the 18th Tall Timbers Fire Ecology Conference: The Longleaf Pine Ecosystem: Ecology, Restoration and Management, Tallahassee, FL, USA, 30 May–2 June 1991; Hermann, S.M., Ed.; Tall Timbers Research, Inc.: Tallahassee, FL, USA, 1993; pp. 17–43. [Google Scholar]
- Kobziar, L.N.; Watts, A.C.; Godwin, D.S.; Taylor, L. Perspectives on trends, effectiveness, and challenges to prescribed burning in the Southern US. Forests 2015, 6, 561–580. [Google Scholar] [CrossRef]
- Clewell, A.; Komarek, R. NB66: The Initiation of a Long-Term Experiment in Forest Succession. Unpublished work. 1975. [Google Scholar]
- Clewell, A. Forest development 44 years after fire exclusion in formerly annually burned old-field pine woodland, Florida. Castanea 2014, 79, 147–167. [Google Scholar] [CrossRef]
- Robertson, K.M.; Ostertag, T.E. Effects of land use on fuel characteristics and fire behavior in pinelands of southwest Georgia. In Proceedings of the 23rd Tall Timbers Fire Ecology Conference: Fire in Grassland and Shrubland Ecosystems, Tall Timbers Research Station, Tallahassee, FL, USA, 17–19 October 2005; Masters, R.E., Galley, K.E., Eds.; Tall Timbers Research Station, Inc.: Tallahassee, FL, USA, 2007; pp. 181–191. [Google Scholar]
- Myers, R.L.; Ewel, J.J. Ecosystems of Florida; University of Central Florida Press: Orlando, FL, USA, 1990. [Google Scholar]
- Lundegardh, H. Carbon dioxide evolution of soil and crop growth. Soil Sci. 1927, 23, 417–453. [Google Scholar] [CrossRef]
- Samuelson, L.J.; Johnson, K.; Stokes, T.; Lu, W. Intensive management modifies soil CO2 efflux in 6-year-old Pinus taeda L. stands. For. Ecol. Manag. 2004, 200, 335–345. [Google Scholar] [CrossRef]
- Godwin, D.R. The influence of prescribed fire and mechanical fuels mastication on soil CO2 efflux rates in two Southeastern U.S. pine ecosystems. Ph.D. Thesis, University of Florida, Gainesville, FL, USA, 2012. [Google Scholar]
- Ewel, K.; Cropper, W.; Gholz, H. Soil CO2 evolution in Florida slash pine plantations. I. Changes through time. Can. J. For. Res. 1987, 17, 325–329. [Google Scholar] [CrossRef]
- Amiro, B.D.; Barr, A.G.; Barr, J.G.; Black, T.A.; Bracho, R.; Brown, M.; Chen, J.; Clark, K.L.; Davis, K.J.; Desai, A.R.; et al. Ecosystem carbon dioxide fluxes after disturbance in forests of North America. J. Geophys. Res. 2010, 115, 2156–2202. [Google Scholar] [CrossRef]
- Luan, J.; Liu, S.; Wang, J.; Zhu, X.; Shi, Z. Rhizospheric and heterotrophic respiration of a warm-temperate oak chronosequence in China. Soil Biol. Biochem. 2011, 43, 503–512. [Google Scholar] [CrossRef]
- Yuste, J.C.; Nagy, M.; Janssens, I.A.; Carrara, A.; Cuelemans, R. Soil respiration in a mixed temperate forest and its contribution to total ecosystem respiration. Tree Physiol. 2005, 25, 609–619. [Google Scholar] [CrossRef]
- Bizzari, L.E.; Collins, C.D.; Brudvig, L.A.; Damschen, E. Historical agriculture and contemporary fire frequency alter soil properties in longleaf pine woodlands. For. Ecol. Manag. 2015, 349, 45–54. [Google Scholar] [CrossRef]
- Wahlenberg, W.G. Effect of fire and grazing on soil properties and the natural reproduction of longleaf pine. J. For. 1939, 33, 331–338. [Google Scholar]
- Boyer, W.D.; Miller, J.H. Effect of burning and brush treatments on nutrient and soil physical properties in young longleaf pine stands. For. Ecol. Manag. 1994, 70, 311–318. [Google Scholar] [CrossRef]
- Williams, R.J.; Hallgren, S.W.; Wilson, G.W. Frequency of prescribed burning in an upland oak forest determines soil and litter properties and alters the soil microbial community. For. Eco. Manag. 2012, 265, 241–247. [Google Scholar] [CrossRef]
- Heyward, F.; Barnette, R.M. Effect of Frequent Fires on Chemical Composition of Forest Soils in the Longleaf Pine Region; Technical Bulletin 265; University of Florida Agricultural Experiment Station: Gainesville, FL, USA, 1934. [Google Scholar]
- Greene, S.W. Effect of annual grass fires on organic matter and other constituents of virgin longleaf pine soils. J. Agric. Res. 1935, 50, 809–822. [Google Scholar]
- Wells, C.G. Effects of prescribed burning on soil chemical properties and nutrient availability. In Proceedings of the Prescribed Burning Symposium; USDA Forest Service Southeastern Forest Experiment Station: Asheville, NC, USA, 14–16 April 1970; pp. 86–99. [Google Scholar]
- McKee, W.H. Changes in Soil Fertility Following Prescribed Burning on Coastal Plain Pine Sites; Research Paper SE-234; USDA Forest Service Southeastern Experimental Forest Research Station: Charleston, SC, USA, 1982.
- Maier, C.A.; Kress, L.W. Soil CO2 evolution and root respiration in 11 year-old loblolly pine (Pinus taeda) plantations as affected by moisture and nutrient availability. Can. J. For. Res. 2000, 30, 347–359. [Google Scholar] [CrossRef]
- Templeton, B.S.; Seiler, J.R.; Peterson, J.A.; Tyree, M.C. Environmental and stand management influences on soil CO2 efflux across the range of loblolly pine. For. Ecol. Manag. 2015, 355, 15–23. [Google Scholar] [CrossRef]
- Janssens, I.A.; Pilegaard, K. Large seasonal changes in Q10 of soil respiration in a beech forest. Glob. Chang. Biol. 2003, 9, 911–918. [Google Scholar] [CrossRef]
- Yuste, J.C.; Janssens, I.A.; Carrara, A.; Cuelemans, R. Annual Q10 of soil respiration reflects plant phenological patterns as well as temperature sensitivity. Glob. Chang. Biol. 2004, 10, 161–169. [Google Scholar] [CrossRef]
- Kuzyakov, Y. Sources of CO2 efflux from soil and review of partitioning methods. Soil Biol. Biochem. 2006, 38, 425–448. [Google Scholar] [CrossRef]
- Ekblad, A.; Högberg, P. Natural abundance of 13C in CO2 respired from forest soils reveals speed of link between tree photosynthesis and root respiration. Acta Oecol. 2001, 127, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Davidson, E.; Richardson, A.; Savage, K.; Hollinger, D.A. distinct seasonal pattern of the ratio of soil respiration to total ecosystem respiration in a spruce-dominated forest. Glob. Chang. Biol. 2006, 12, 230–239. [Google Scholar] [CrossRef]
- Wertin, T.M.; Teskey, R.O. Close coupling of whole-plant respiration to net photosynthesis and carbohydrates. Tree Physiol. 2008, 28, 1831–1840. [Google Scholar] [CrossRef] [PubMed]
- Fenn, K.M.; Malhi, Y.; Morecroft, M.D. Soil CO2 efflux in a temperate deciduous forest: Environmental drivers and component contributions. Soil Biol. Biochem. 2010, 42, 1685–1693. [Google Scholar] [CrossRef]
Figure 1.
The research site, Tall Timbers Research Station, was located in Leon County, FL, USA. The site is approximately 30 km north of the city of Tallahassee, FL, USA.
Figure 1.
The research site, Tall Timbers Research Station, was located in Leon County, FL, USA. The site is approximately 30 km north of the city of Tallahassee, FL, USA.
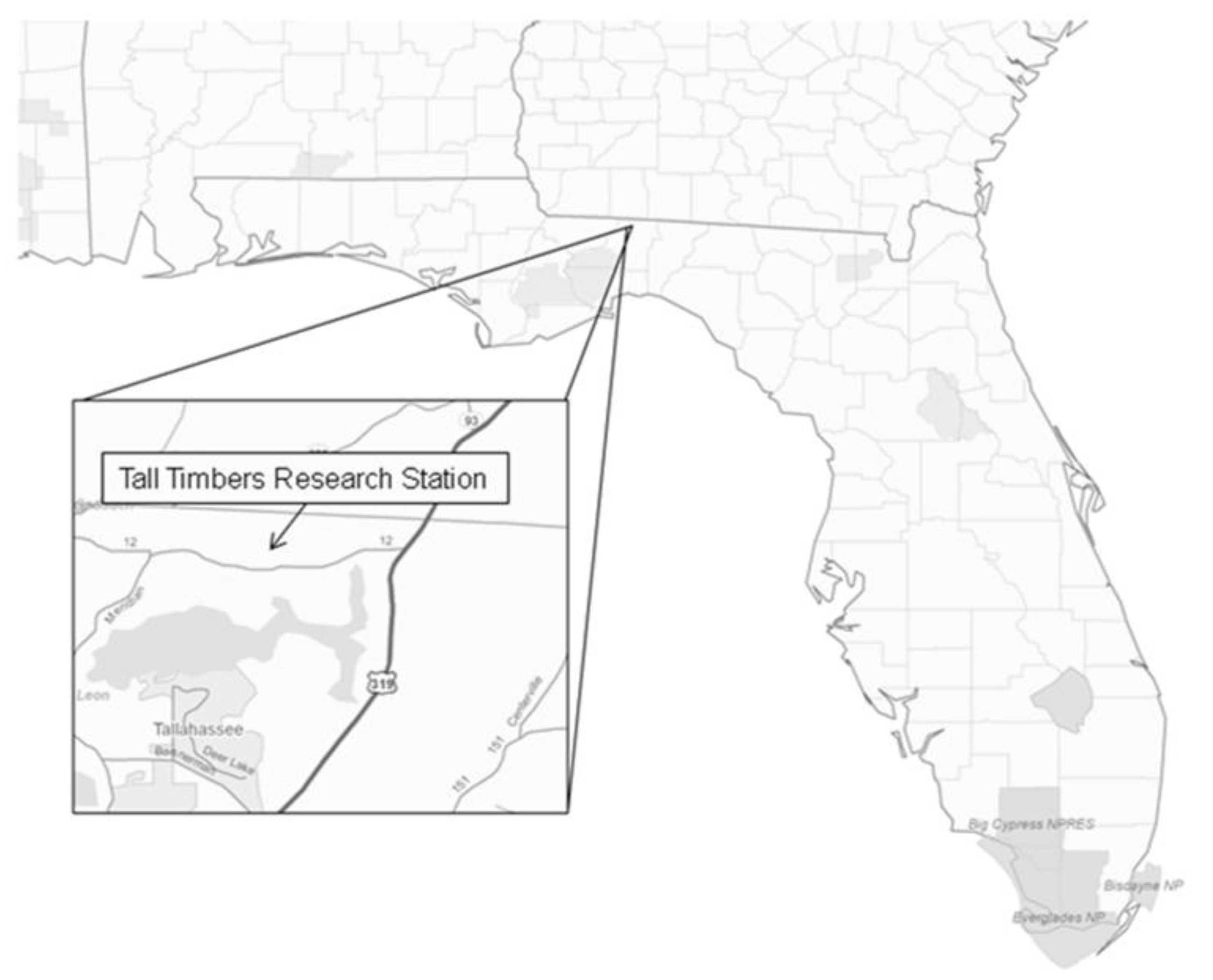
Figure 2.
Monthly mean soil CO2 efflux rates (Rs) (μmol CO2 m−2·s−1) for three prescribed fire regimes at the Tall Timbers Research Station near Tallahassee, FL, USA. Data were from 27 sample points per fire regime, measured three-times daily once per month. Measurements were not taken in October of 2010.
Figure 2.
Monthly mean soil CO2 efflux rates (Rs) (μmol CO2 m−2·s−1) for three prescribed fire regimes at the Tall Timbers Research Station near Tallahassee, FL, USA. Data were from 27 sample points per fire regime, measured three-times daily once per month. Measurements were not taken in October of 2010.
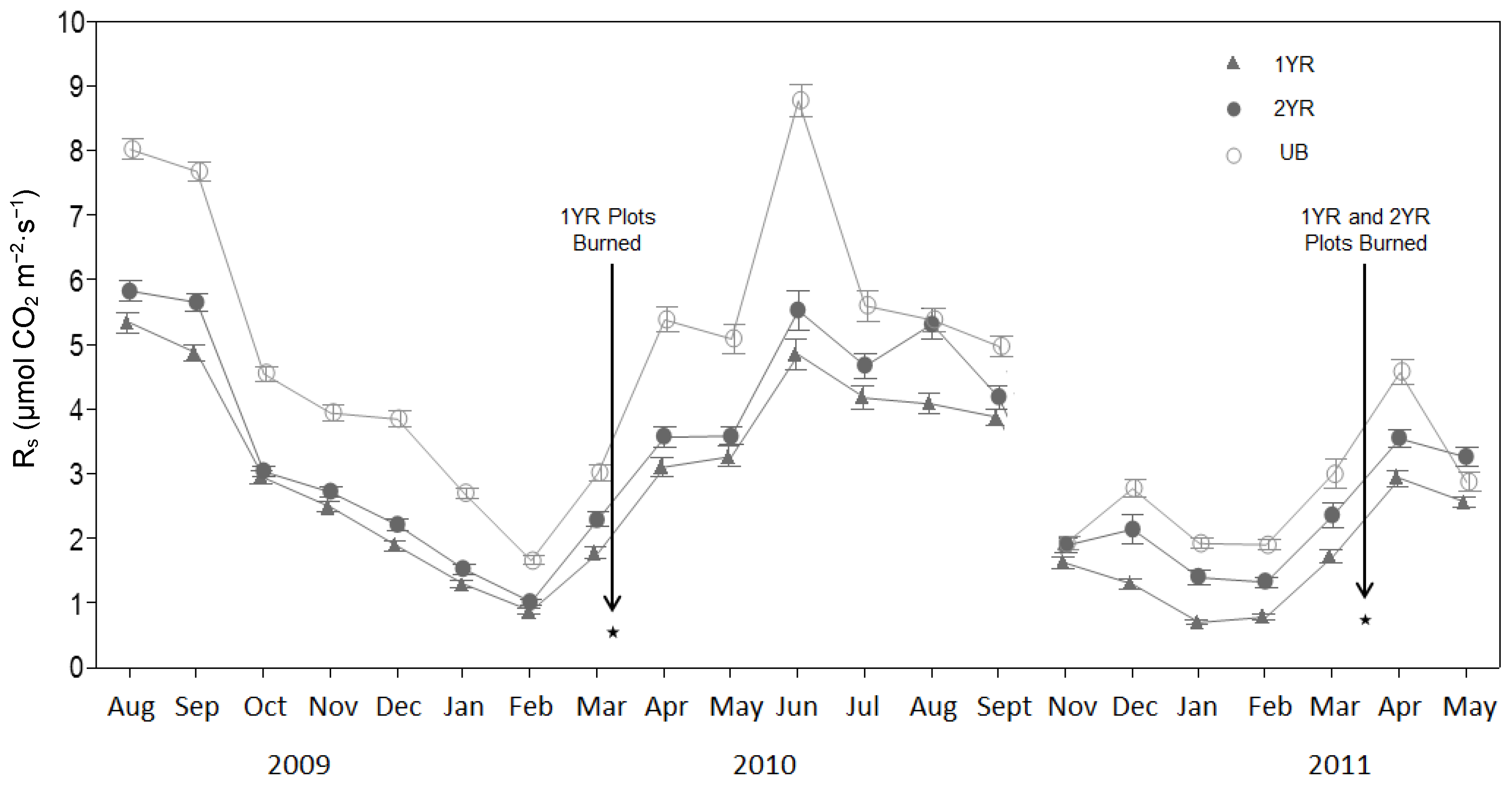
Figure 3.
Monthly mean soil temperature (Ts) (°C) for three prescribed fire regimes at the Tall Timbers Research Station near Tallahassee, FL, USA. Mean monthly 2 m air temperature also shown for comparison. Data were from 27 sample points per fire regime, measured three-times daily once per month. Equipment problems eliminated measurements taken in January 2009 and February 2011. Measurements were not taken in October of 2010.
Figure 3.
Monthly mean soil temperature (Ts) (°C) for three prescribed fire regimes at the Tall Timbers Research Station near Tallahassee, FL, USA. Mean monthly 2 m air temperature also shown for comparison. Data were from 27 sample points per fire regime, measured three-times daily once per month. Equipment problems eliminated measurements taken in January 2009 and February 2011. Measurements were not taken in October of 2010.
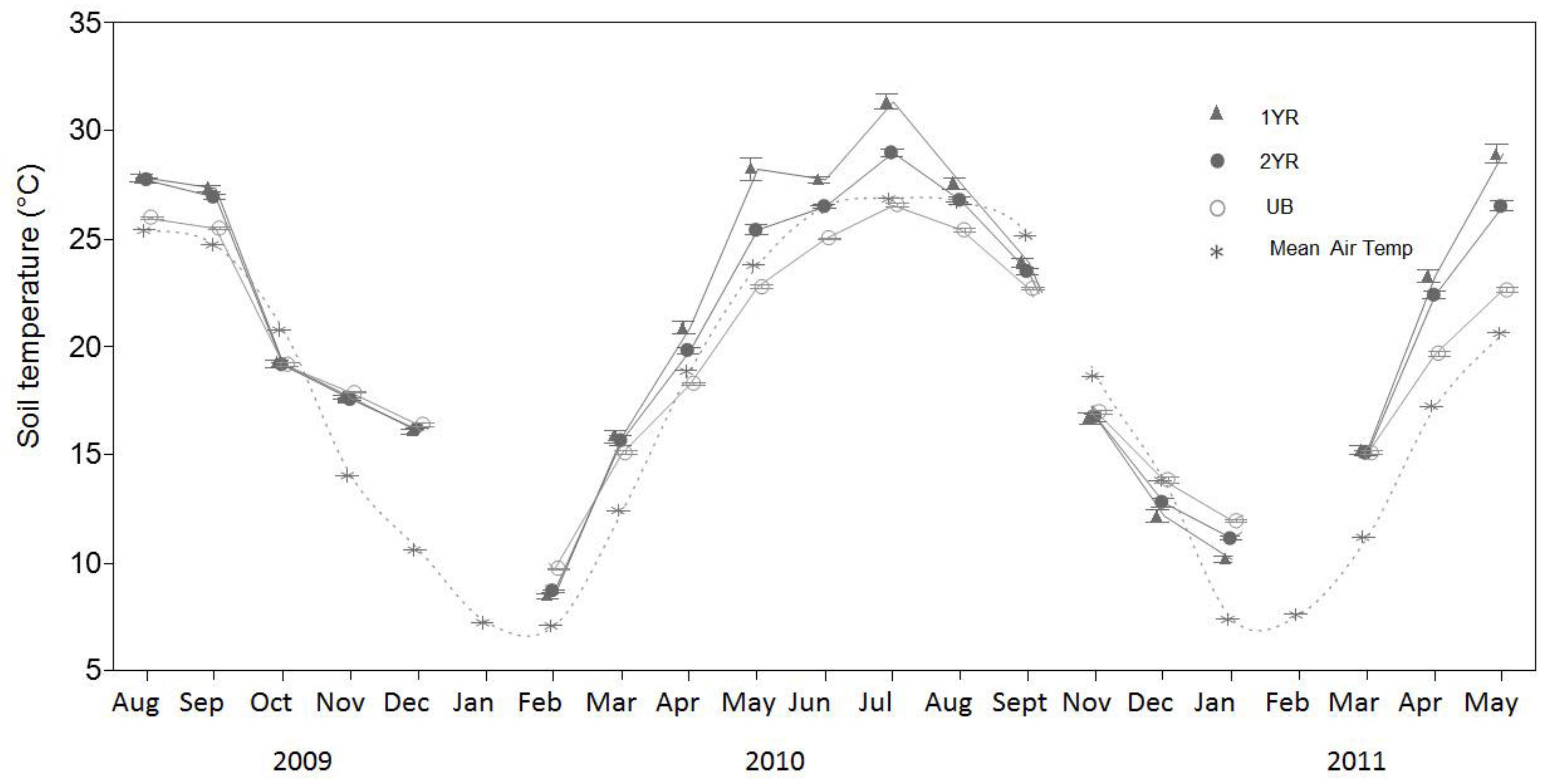
Figure 4.
Monthly mean soil moisture content (Ms) (m3/m3) for three prescribed fire regimes at the Tall Timbers Research Station near Tallahassee, FL, USA. Mechanical difficulty resulted in erroneous data collected in August 2009. Measurements were not taken in October of 2010.
Figure 4.
Monthly mean soil moisture content (Ms) (m3/m3) for three prescribed fire regimes at the Tall Timbers Research Station near Tallahassee, FL, USA. Mechanical difficulty resulted in erroneous data collected in August 2009. Measurements were not taken in October of 2010.
Figure 5.
Linear regression of the relationships between monthly mean soil CO2 efflux rates (Rs) (μmol CO2 m−2·s−1) and monthly mean soil temperature (Ts) (°C) for three prescribed fire intervals at the Tall Timbers Research Station near Tallahassee, FL, USA. Each point represents monthly mean values per sample plot. p-Values were considered significant at α = 0.05.
Figure 5.
Linear regression of the relationships between monthly mean soil CO2 efflux rates (Rs) (μmol CO2 m−2·s−1) and monthly mean soil temperature (Ts) (°C) for three prescribed fire intervals at the Tall Timbers Research Station near Tallahassee, FL, USA. Each point represents monthly mean values per sample plot. p-Values were considered significant at α = 0.05.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Environmental variables measured in each research plot, with abbreviation, unit of measurement, and measurement regime provided. Monthly measurements were made from August 2009 to April 2011.
Table 1.
Environmental variables measured in each research plot, with abbreviation, unit of measurement, and measurement regime provided. Monthly measurements were made from August 2009 to April 2011.
| Plot Variable | Abbreviation/Units | Measured |
|---|---|---|
| Soil temperature | Ts (°C) | Monthly |
| Soil moisture content | Ms (m3/m3) | Monthly |
| Basal area | BA (m2·ha−1) | Winter 2011 |
| Pine basal area | PBA (m2·ha−1) | Winter 2011 |
| Hardwood basal area | HWBA (m2·ha−1) | Winter 2011 |
| Stand density | TPH (trees ha−1) | Winter 2011 |
| Duff depth | Duff (cm) | Spring 2011 |
| Litter depth | Litter (cm) | Spring 2011 |
| Soil bulk density 0–10 cm | BD (g·cm−3) | Winter 2013 |
| Soil total carbon 0–10 cm | C (Mg·ha−1) | Winter 2013 |
| Soil total nitrogen 0–10 cm | N (Mg·ha−1) | Winter 2013 |
| Soil phosphorus 0–10 cm | P (kg·ha−1) | Winter 2013 |
| Soil calcium 0–10 cm | Ca (kg·ha−1) | Winter 2013 |
| Soil magnesium 0–10 cm | Mg (kg·ha−1) | Winter 2013 |
| Soil potassium 0–10 cm | K (kg·ha−1) | Winter 2013 |
| Soil pH 0–10 cm | pH | Winter 2013 |
Table 2.
Results of the repeated measures ANOVA showing effects of prescribed fire regime (FR); and time (monthly means of measurements) on soil CO2 efflux (Rs), soil temperature (Ts) and soil moisture content (Ms) (n = 9 three replicates within three treatments).
Table 2.
Results of the repeated measures ANOVA showing effects of prescribed fire regime (FR); and time (monthly means of measurements) on soil CO2 efflux (Rs), soil temperature (Ts) and soil moisture content (Ms) (n = 9 three replicates within three treatments).
| Analysis Period | Source | Rs | Ts | Ms | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| df | F | p-Value | df | F | p-Value | df | F | p-Value | ||
| Entire Study | Month | 20 | 105.19 | <0.0001 | 18 | 306.76 | <0.0001 | 19 | 97.51 | <0.0001 |
| FR × Month | 40 | 3.25 | <0.0001 | 36 | 4.04 | <0.0001 | 38 | 1.68 | 0.0190 | |
| FR | 2 | 20.72 | 0.0007 | 2 | 3.09 | 0.1007 | 2 | 2.95 | 0.1130 | |
| Fall | Month | 5 | 114.16 | <0.0001 | 5 | 241.78 | <0.0001 | 5 | 66.73 | <0.0001 |
| FR × Month | 10 | 4.12 | 0.0012 | 10 | 1.31 | 0.2684 | 10 | 2.06 | 0.0611 | |
| FR | 2 | 17.65 | 0.0031 | 2 | 2.70 | 0.1457 | 2 | 4.26 | 0.0705 | |
| Winter | Month | 5 | 25.16 | <0.0001 | 3 | 61.75 | <0.0001 | 5 | 25.45 | <0.0001 |
| FR × Month | 10 | 1.96 | 0.0761 | 6 | 0.40 | 0.8650 | 10 | 1.96 | 0.0753 | |
| FR | 2 | 13.40 | 0.0061 | 2 | 10.48 | 0.0036 | 2 | 6.30 | 0.0335 | |
| Spring | Month | 5 | 25.61 | <0.0001 | 5 | 243.07 | <0.0001 | 5 | 75.96 | <0.0001 |
| FR × Month | 10 | 2.73 | 0.0168 | 10 | 5.50 | 0.0001 | 10 | 1.03 | 0.4454 | |
| FR | 2 | 11.24 | 0.0054 | 2 | 4.11 | 0.0646 | 2 | 1.41 | 0.3003 | |
| Summer | Month | 2 | 16.70 | 0.0003 | 2 | 38.05 | <0.0001 | 1 | 139.44 | <0.0001 |
| FR × Month | 4 | 3.78 | 0.0327 | 4 | 4.70 | 0.0163 | 2 | 3.02 | 0.1235 | |
| FR | 2 | 16.03 | 0.0039 | 2 | 9.91 | 0.0126 | 2 | 1.32 | 0.3340 | |
p-Values were considered significant at α = 0.05, df = degrees of freedom.
Table 3.
Mean seasonal and total study period soil CO2 efflux rates (μmol CO2 m−2·s−1) (Rs) per prescribed fire regime.
Table 3.
Mean seasonal and total study period soil CO2 efflux rates (μmol CO2 m−2·s−1) (Rs) per prescribed fire regime.
| FR | Fall Rs | Winter Rs | Spring Rs | Summer Rs | Study Mean Rs |
|---|---|---|---|---|---|
| 1YR | 3.36 (1.71) b | 1.24 (0.87) b | 2.56 (1.21) b | 4.93 (1.95) b | 2.67 (1.87) b |
| 2YR | 3.75 (2.02) b | 1.60 (1.24) b | 3.09 (1.39) b | 5.49 (2.11) b | 3.09 (2.12) b |
| UB | 4.93 (2.36) a | 2.61 (1.49) a | 3.98 (1.94) a | 7.59 (2.25) a | 4.22 (2.52) a |
Values are seasonal means with standard deviations in parentheses. Letters show significant differences between fire regimes (Tukey’s HSD test). p-Values were considered significant at α = 0.05.
Table 4.
Mean forest characteristics per prescribed fire treatment (FR). See Table 1 for abbreviations and units. Data in parentheses are standard deviation. Letters per column show significant differences between fire return intervals (Tukey’s HSD test); p-Values were considered significant at α = 0.05.
Table 4.
Mean forest characteristics per prescribed fire treatment (FR). See Table 1 for abbreviations and units. Data in parentheses are standard deviation. Letters per column show significant differences between fire return intervals (Tukey’s HSD test); p-Values were considered significant at α = 0.05.
| (A) | ||||||||
|---|---|---|---|---|---|---|---|---|
| FR | TPH | HWBA | PBA | BA | Duff | Litter | ||
| 1YR | 282.9 (64.83) a | 3.87 (6.28) a | 7.92 (3.68) a | 11.79 (7.22) a | 0.08 (0.06) a | 1.77 (0.91) a | ||
| 2YR | 400.81 (344.87) a | 6.30 (4.28) ab | 9.16 (6.14) a | 15.45 (2.15) a | 0.46 (0.41) b | 2.17 (0.79) b | ||
| UB | 1716.41 (681.42) b | 15.73 (3.59) b | 21.99 (11.22) a | 37.72 (8.36) b | 1.58 (0.55) c | 2.81 (0.58) c | ||
| (B) | ||||||||
| FR | BD | C | N | P | Ca | Mg | K | pH |
| 1YR | 1.24 (0.04) a | 26.53 (2.49) ab | 1.23 (0.18) | 33.46 (25.18) | 853.1 (506.4) | 185.5 (69.8) a | 38.08 (18.80) | 5.48 (0.37) |
| 2YR | 1.26 (0.07) a | 30.33 (4.16) a | 1.18 (0.73) | 11.69 (7.58) | 737.8 (264.3) | 141.5 (45.6) ab | 27.54 (8.12) | 5.29 (0.18) |
| UB | 1.07 (0.04) b | 21.56 (2.38) b | 1.05 (0.16) | 11.16 (4.78) | 301.8 (185.4) | 66.3 (23.4) b | 25.21 (4.68) | 4.86 (0.31) |
Table 5.
Linear and nonlinear regression relationships between soil CO2 efflux rates (Rs) and soil temperature (Ts) by fire return interval (FR).
Table 5.
Linear and nonlinear regression relationships between soil CO2 efflux rates (Rs) and soil temperature (Ts) by fire return interval (FR).
| FR | Model Type | Model and Estimates | Q10 | R2 | p-Value |
|---|---|---|---|---|---|
| 1YR | Linear | 0.62 | <0.001 | ||
| 1YR | Nonlinear | 1.65 | 0.60 | <0.001 | |
| 2YR | Linear | 0.78 | <0.001 | ||
| 2YR | Nonlinear | 1.96 | 0.80 | <0.001 | |
| UB | Linear | 0.65 | <0.001 | ||
| UB | Nonlinear | 2.16 | 0.76 | <0.001 |
Model data use mean monthly measurements of Rs and Ts.
Table 6.
Linear regression relationship between soil CO2 efflux rates (Rs) and soil temperature (Ts) by fire return interval (FR) and season. p-Values were considered significant at α = 0.05.
Table 6.
Linear regression relationship between soil CO2 efflux rates (Rs) and soil temperature (Ts) by fire return interval (FR) and season. p-Values were considered significant at α = 0.05.
| FR | Season 1 | Model and Estimates | R2 | p-Value |
|---|---|---|---|---|
| 1YR | Spring | 0.29 | 0.021 | |
| Summer | 0.40 | 0.068 | ||
| Fall | 0.71 | <0.001 | ||
| Winter | 0.69 | 0.002 | ||
| 2YR | Spring | 0.48 | 0.002 | |
| Summer | 0.28 | 0.144 | ||
| Fall | 0.90 | <0.001 | ||
| Winter | 0.71 | 0.002 | ||
| UB | Spring | 0.11 | 0.170 | |
| Summer | 0.63 | 0.011 | ||
| Fall | 0.71 | <0.001 | ||
| Winter | 0.90 | <0.001 |
1 Spring (March–May), summer (June–August), fall (September–November), and winter (December–February).
Table 7.
Seasonal nonlinear models of soil CO2 efflux (Rs) rates using soil temperature (Ts) as a predictor. Models are of monthly mean Rs responses to Ts. p-values were considered significant at α= 0.05.
Table 7.
Seasonal nonlinear models of soil CO2 efflux (Rs) rates using soil temperature (Ts) as a predictor. Models are of monthly mean Rs responses to Ts. p-values were considered significant at α= 0.05.
| FR | Season | Equation | Q10 | R2 | p-Value |
|---|---|---|---|---|---|
| 1YR | Spring | 1.27 | 0.26 | 0.031 | |
| Summer | 0.49 | 0.45 | 0.049 | ||
| Fall | 1.82 | 0.66 | <0.001 | ||
| Winter | 3.32 | 0.77 | 0.000 | ||
| 2YR | Spring | 3.49 | 0.33 | 0.012 | |
| Summer | 0.50 | 0.27 | 0.149 | ||
| Fall | 2.28 | 0.89 | <0.001 | ||
| Winter | 3.05 | 0.78 | 0.001 | ||
| UB | Spring | 1.34 | 0.10 | 0.195 | |
| Summer | 0.25 | 0.62 | 0.012 | ||
| Fall | 2.42 | 0.69 | <0.001 | ||
| Winter | 3.99 | 0.96 | <0.001 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Godwin, D.R.; Kobziar, L.N.; Robertson, K.M. Effects of Fire Frequency and Soil Temperature on Soil CO2 Efflux Rates in Old-Field Pine-Grassland Forests. Forests 2017, 8, 274. https://doi.org/10.3390/f8080274
AMA Style
Godwin DR, Kobziar LN, Robertson KM. Effects of Fire Frequency and Soil Temperature on Soil CO2 Efflux Rates in Old-Field Pine-Grassland Forests. Forests. 2017; 8(8):274. https://doi.org/10.3390/f8080274
Chicago/Turabian StyleGodwin, David R., Leda N. Kobziar, and Kevin M. Robertson. 2017. "Effects of Fire Frequency and Soil Temperature on Soil CO2 Efflux Rates in Old-Field Pine-Grassland Forests" Forests 8, no. 8: 274. https://doi.org/10.3390/f8080274
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.