
Genetic Diversity among Age Classes of a Pinus sylvestris (L.) Population from the Białowieża Primeval Forest, Poland


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Sampling
2.2. DNA Extraction and Amplification
2.3. Data Analysis
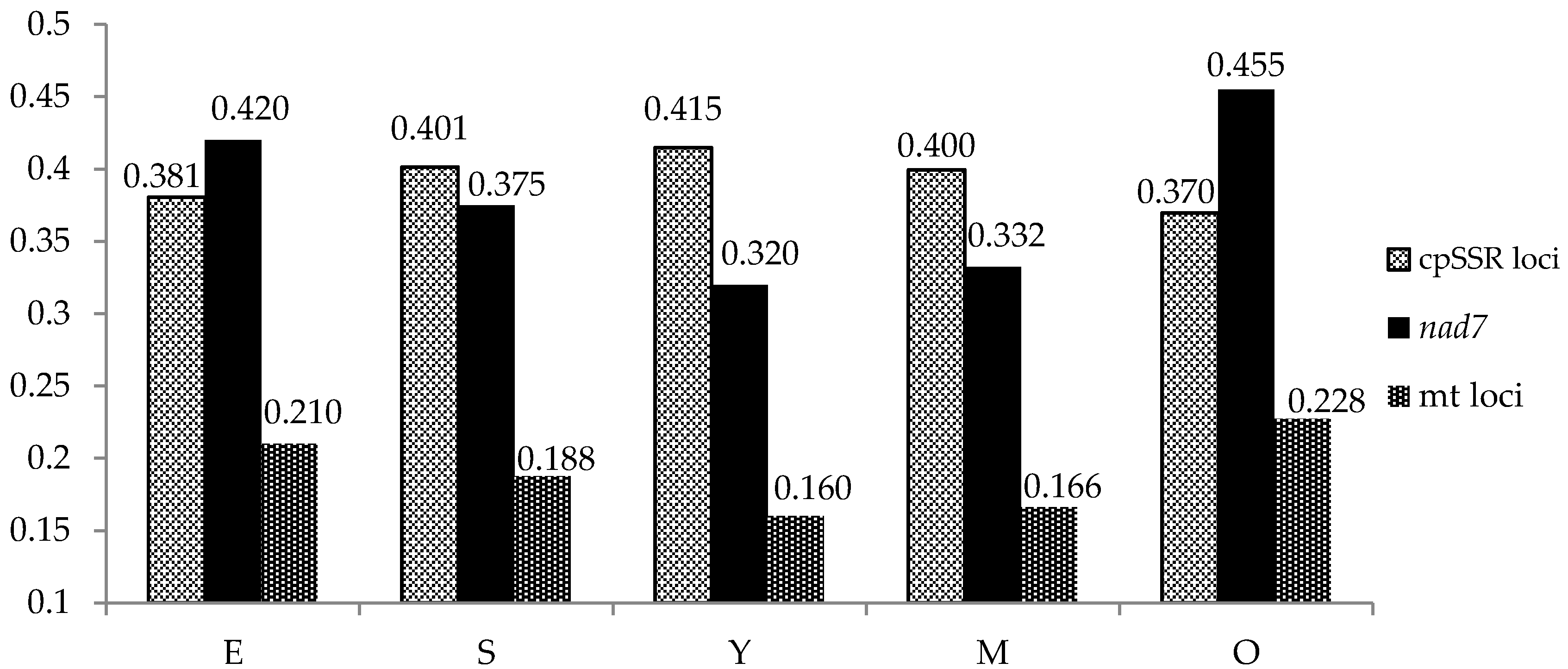
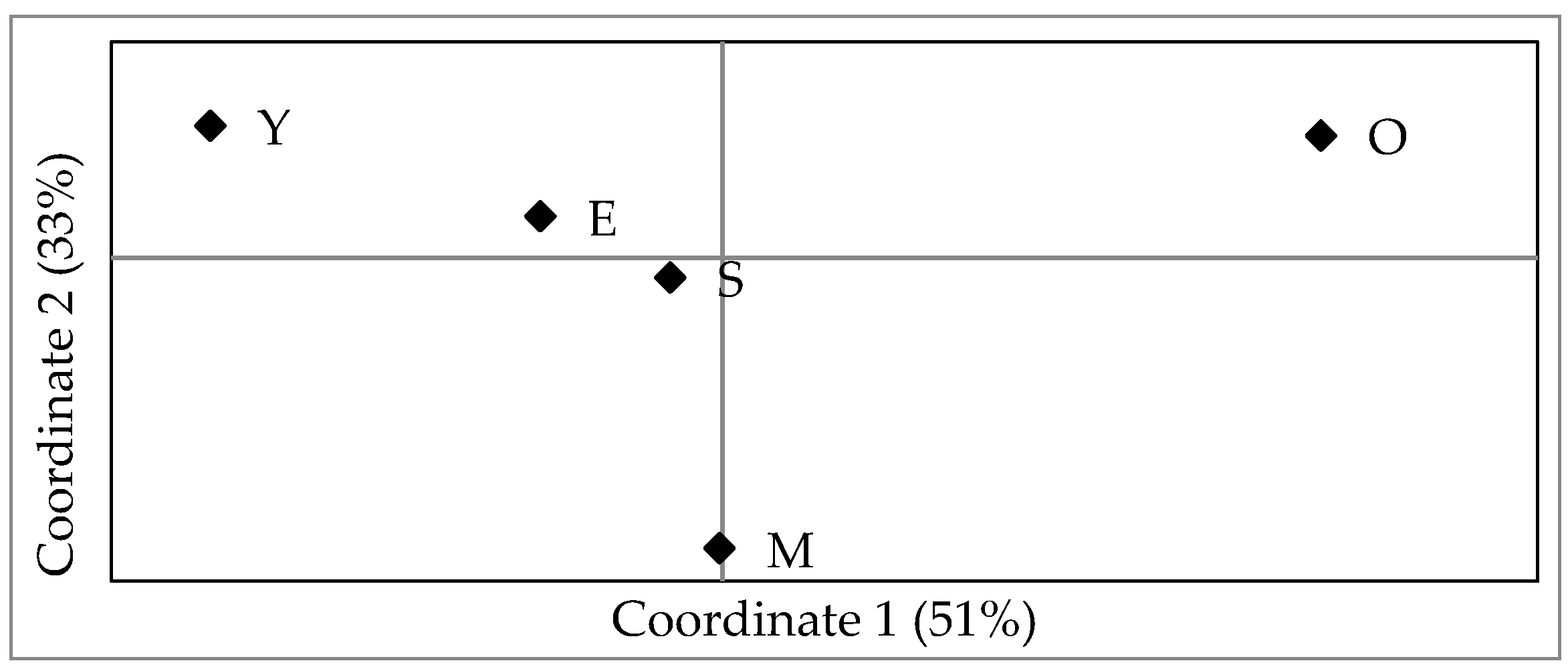
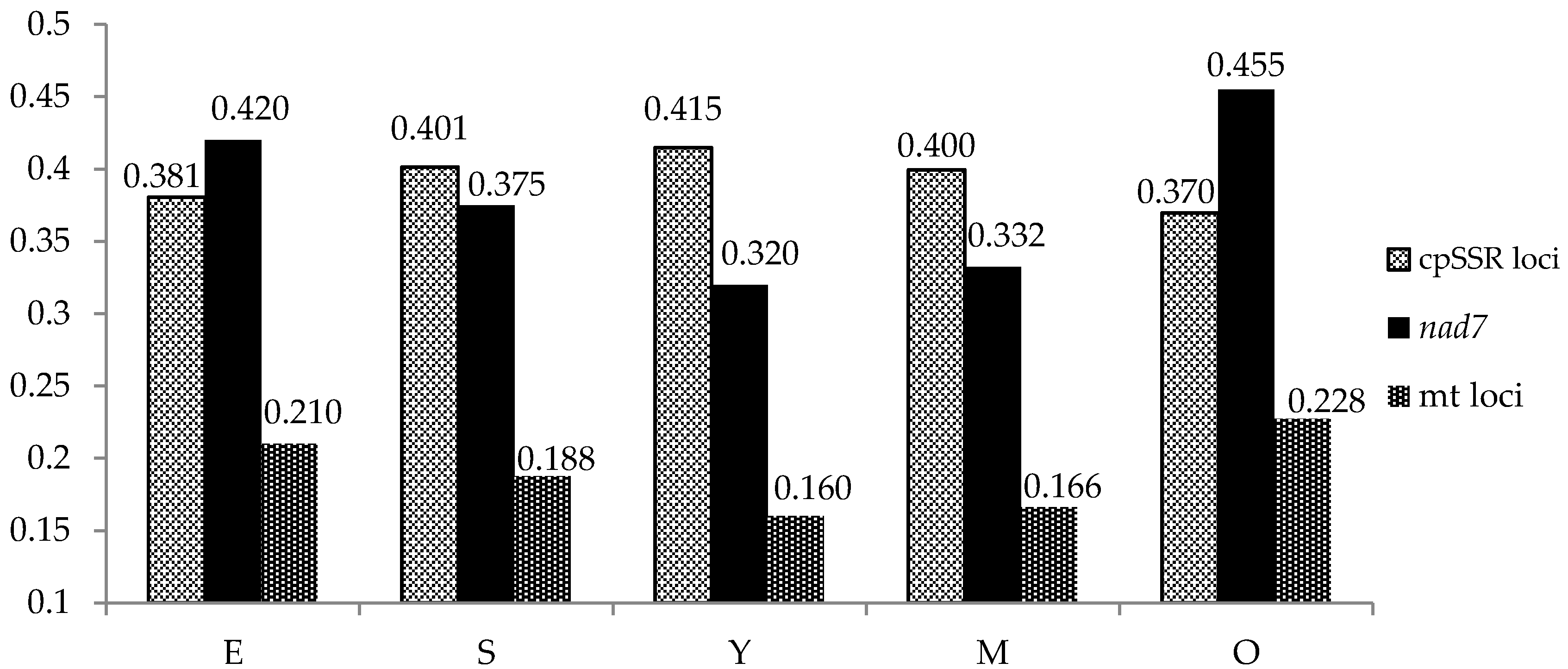
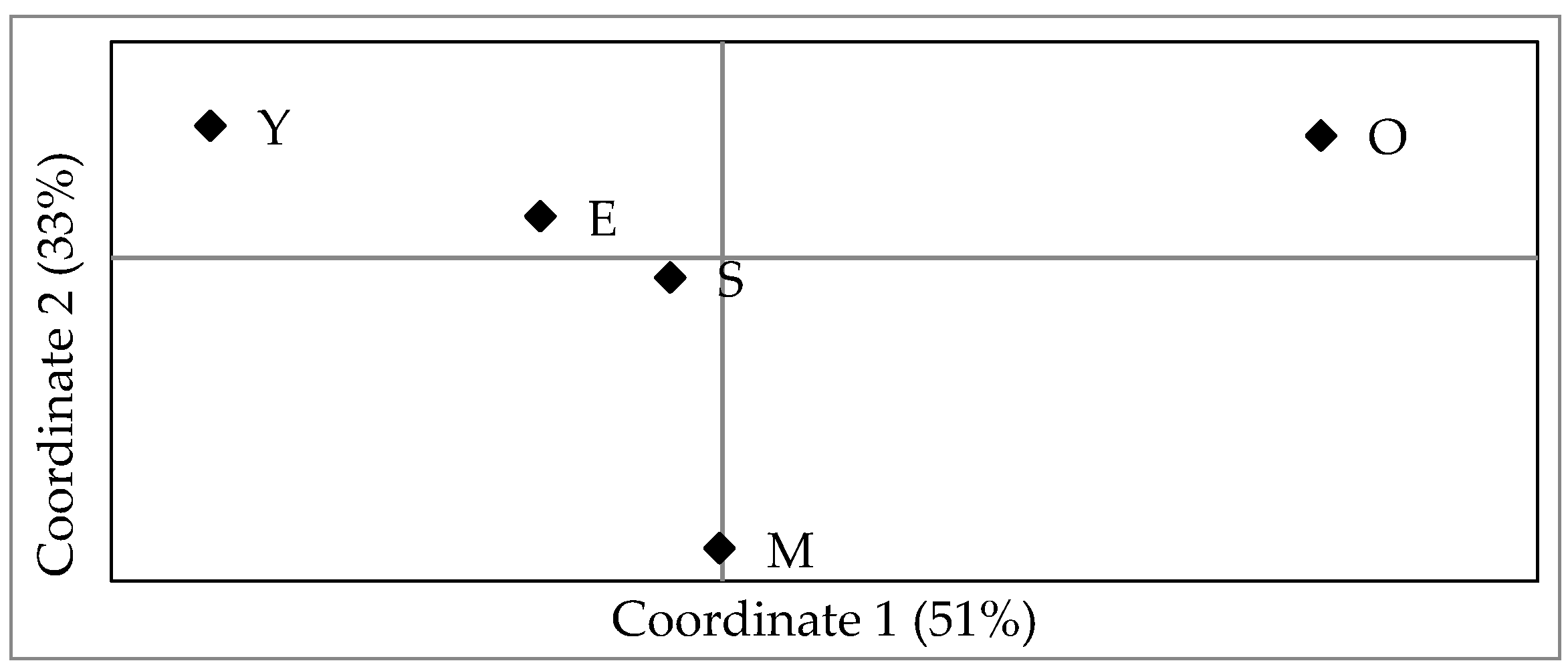
3. Results
3.1. Chloroplast simple sequence repeats (cpSSRs)
3.2. Mitochondrial Markers
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Baker, A.G.; Zimny, M.; Keczyński, A.; Bhagwat, S.A.; Willis, K.J.; Latałowa, M. Pollen productivity estimates from old-growth forest strongly differ from those obtained in cultural landscapes: Evidence from the Białowieża National Park, Poland. Holocene 2016, 26, 80–92. [Google Scholar] [CrossRef]
- Faliński, J.B. Vegetation Dynamics in Temperate Lowland Primeval Forests: Ecological Studies in Białowieża Forest; Junk Publishers: Dordrecht, The Netherlands, 1986. [Google Scholar]
- Bobiec, A. Białowieża Primeval Forest.The largest area of natural deciduous lowland forest in Europe. Int. J. Wilderness 2002, 8, 33–37. [Google Scholar]
- Lorenz, M.; Fischer, R.; Mues, V. Forest Resources in Europe and Their Condition; Arbora Publishers: Zvolen, Slovakia, 2005. [Google Scholar]
- Koop, H. Forest Dynamics. SILVI-STAR: A Comprehensive Monitoring System; Springer: Berlin, Germany, 1989. [Google Scholar]
- Ratnam, W.; Rajora, O.P.; Finkeldey, R.; Aravanopoulos, F.; Bouvet, J.M.; Vaillancourtf, R.E.; Kanashiro, M.; Fady, B.; Tomita, M.; Vinson, C. Genetic effects of forest management practices: Global synthesis and perspectives. For. Ecol. Manag. 2014, 333, 52–65. [Google Scholar] [CrossRef]
- Ackzell, L. Comparison of planting, sowing and natural regeneration for Pinus sylvestris (L.) in Boreal Sweden. For. Ecol. Manag. 1993, 61, 229–245. [Google Scholar] [CrossRef]
- Chomicz, E. Factor shaping genetic diversity of forest trees. Kosmos 2013, 62, 597–605, (In Polish with English summary). [Google Scholar]
- Poorter, L.; Bongers, F.; Sterck, F.J.; Woll, H. Beyond the regeneration phase: Differentiation of height-light trajectories among tropical tree species. J. Ecol. 2005, 93, 256–267. [Google Scholar] [CrossRef]
- Grey, B.M. Comment on transfer functions. In Climate from Tree Ring; Huges, M.K., Kelly, P.M., Pilcher, J.M., La Marche, V.C., Eds.; Cambridge University Press: Cambridge, UK, 1982; pp. 56–58. [Google Scholar]
- Szeicz, J.M.; MacDonald, G.M. Age-dependent tree ring growth on subarctic white spruce to climate. Can. J. For. Res. 1994, 24, 120–132. [Google Scholar] [CrossRef]
- Linderholm, H.W.; Linderholm, K. Age-dependent climate sensitivity of Pinus sylvestris L. in the central Scandinavian Mountains. Boreal Environ. Res. 2004, 9, 307–317. [Google Scholar]
- Chen, H.Y.H.; Luo, Y.; Reich, P.B.; Searle, E.B.; Biswas, S.R. Climate change-associated trends in net biomass change are age dependent in western boreal forests of Canada. Ecol. Lett. 2016, 19, 1150–1158. [Google Scholar] [CrossRef] [PubMed]
- Korczyk, A. Protection and conservation of gene resources of forest trees in the Białowieża Forest. In Protection of Forest Ecosystems Biodiversity of Białowieża Primeval Forest; Paschalis, P., Rykowski, K., Zajączkowski, S., Eds.; Pol Forest Biodiversity Protection Project; Warsaw, Poland, 1995; pp. 95–102. [Google Scholar]
- Wojnicka-Półtorak, A.; Celiński, K.; Chudzińska, E. Genetic differentiation between generations of Pinus sylvestris natural population: A case study from the last European primeval forest. Austrian J. For. Sci. 2017, in press. [Google Scholar]
- Sokołowski, A. Lasy Puszczy Białowieskiej; Centrum Informacyjne Lasów Państwowych: Warszawa, Poland, 2004. (In Polish) [Google Scholar]
- Korczyk, A.F. Identification of Genetic Diversity. Part I. Demographic Structure of Natural Populations of Pinus sylvestris L. and Picea abies Karst in the Białowieża Forest; Report GEF 05/21655 Pol part 3.04.; Global Environment Facility: Warsaw, Poland, 1994; (In Polish with English abstract). [Google Scholar]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 1, 13–15. [Google Scholar]
- Provan, J.; Soranzo, N.; Wilson, N.J.; McNicol, J.W.; Forrest, G.I.; Cottrell, J.; Powell, W. Gene-pool variation in Caledonian and European Scots pine (Pinus sylvestris L.) revealed by chloroplast simple-sequence repeats. Proc. R. Soc. B Biol. Sci. 1998, 265, 1697–1705. [Google Scholar] [CrossRef] [PubMed]
- Vendramin, G.G.; Lelli, L.; Rossi, P.; Morgante, M. A set of primers for the amplification of 20 chloroplast microsatellites in Pinaceae. Mol. Ecol. 1996, 5, 595–598. [Google Scholar] [CrossRef] [PubMed]
- Celiński, K.; Pawlaczyk, E.M.; Wojnicka-Półtorak, A.; Chudzińska, E.; Prus-Głowacki, W. Cross-species amplification and characterization of microsatellite loci in Pinus mugo Turra. Biologia 2013, 68, 621–626. [Google Scholar] [CrossRef]
- Soranzo, N.; Alia, R.; Provan, J.; Powell, W. Patterns of variation at a mitochondrial sequence-tagged-site locus provides new insights into the postglacial history of European Pinus sylvestris populations. Mol. Ecol. 2000, 9, 1205–1211. [Google Scholar] [CrossRef] [PubMed]
- Naydenov, K.; Senneville, S.; Beaulieu, J.; Tremblay, F.; Bosquet, J. Glacial vicariance in Eurasia: Mitochondrial DNA evidence from Scots pine for a complex heritage involving genetically distinct refugia at mid-northern latitudes and in Asia Minor. BMC Evol. Biol. 2007, 7, 233. [Google Scholar] [CrossRef] [PubMed]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar] [PubMed]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-un update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Raymond, M.; Rousset, F. An exact test for population differentiation. Evolution 1995, 49, 1280–1283. [Google Scholar] [CrossRef] [PubMed]
- Raymond, M.; Rousset, F. GenePop (version 1.2): A population genetics software for exact tests and ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Rousset, F. GENEPOP’007: A complete re-implementation of the GENEPOP software for Windows and Linux. Mol. Ecol. Res. 2008, 8, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Meirmans, P.G. Using the AMOVA framework to estimate a standardized genetic differentiation measure. Evolution 2006, 60, 2399–2402. [Google Scholar] [CrossRef] [PubMed]
- Nei, M. Genetic distance between populations. Am. Nat. 1972, 106, 283–292. [Google Scholar] [CrossRef]
- Pazouki, L.; Shanjani, P.S.; Fields, P.D.; Martins, K.; Suhhorutsenko, M.; Viinalass, H.; Niinemets, U. Large within-population genetic diversity of the widespread conifer Pinus sylvestris at its soil fertility limit characterized by nuclear and chloroplast microsatellite markers. Eur. J. For. Res. 2016, 135, 161–177. [Google Scholar] [CrossRef]
- Semerikov, V.L.; Semerikova, S.A.; Dymshakova, O.S.; Zatsepina, K.G.; Tarakanov, V.V.; Tikhonova, I.V.; Ekart, A.K.; Vidyakin, A.I.; Jamiyansuren, S.; Rogovtsev, R.V.; et al. Microsatellite loci polymorphism of chloroplast DNA of the pine tree (Pinus sylvestris L.) in Asia and Eastern Europe. Genetika 2014, 50, 660–669. [Google Scholar] [PubMed]
- Ribeiro, M.M.; Plomion, P.; Petit, R.; Vendramin, G.G.; Szmidt, A.E. Variation in chloroplast single-sequence repeats in Portuguese maritime pine (Pinus pinaster Ait.). Theor. Appl. Genet. 2001, 102, 97–103. [Google Scholar] [CrossRef]
- Parducci, R.; Szmidt, A.E.; Madaghiele, M.; Anzidei, M.; Vendramin, G.G. Genetic variation at chloroplast microsatellites (cpSSRs) in Abies nebrodensis (Lojac.) Mattei and three neighboring Abies species. Theor. Appl. Genet. 2001, 102, 733–740. [Google Scholar] [CrossRef]
- Echt, C.S.; DeVerno, L.L.; Anzidei, M.; Vendramin, G.G. Chloroplast microsatellites reveal population genetic diversity in red pine, Pinus resinosa Ait. Mol. Ecol. 1998, 7, 307–316. [Google Scholar] [CrossRef]
- Schmidt, A.; Semerikova, S.A.; Semerikov, V.L. The diversity of chloroplast microsatellite loci in Siberian fir (Abies sibirica Ledeb.) and two Far East fir species A. nephrolepis (Trautv.) Maxim and A. sachalinensis Fr. Schmidt. Russ. J. Genet. 2007, 43, 1373–1381. [Google Scholar]
- Robledo-Arnuncio, J.J.; Alia, R.; Gil, L. High levels of genetic diversity in a long-term European glacial refugium of Pinus sylvestris L. For. Genet. 2004, 11, 239–248. [Google Scholar]
- Naydenov, K.D.; Tremblay, F.M.; Alexandrov, A.; Fenton, N.J. Structure of Pinus sylvestris L. populations in Bulgaria revealed by chloroplast microsatellites and terpenes analysis: Provenance tests. Biochem. Syst. Ecol. 2005, 33, 1226–1245. [Google Scholar] [CrossRef]
- Semerikov, V.L.; Semerikova, S.A.; Dymshakova, O.S.; Zatsepina, K.G.; Tarakanov, V.V.; Tikhonova, I.V.; Ekart, A.K.; Vidyakin, A.I.; Jamiyansuren, S.; Rogovtsev, R.V.; et al. Microsatellite loci polymorphism of chloroplast DNA of Scots pine (Pinus sylvestris L.) in Asia and Eastern Europe. Russ. J. Genet. 2014, 50, 577–585. [Google Scholar] [CrossRef]
- Wang, X.-R.; Szmidt, A.E.; Lindgren, D.A.G. Allozyme differentiation among populations of Pinus sylvestris (L.) from Sweden and China. Hereditas 1991, 114, 219–226. [Google Scholar] [CrossRef]
- Goncharenko, G.G.; Silin, A.E.; Padutov, V.E. Allozyme variation in natural populations of Eurasian pines.3. Population structure, diversity, differentiation and gene flow in central and isolated populations of Pinus sylvestris L. in Eastern-Europe and Siberia. Silvae Genet. 1994, 43, 119–132. [Google Scholar]
- Shigapov, Z.K.H.; Bakhtiyarova, R.M.; Yanbaev, Y.A. Genetic variation and differentiation in natural populations of the Scots Pine, Pinus silvestris L. Russ. J. Genet. 1995, 31, 1180–1186. [Google Scholar]
- Sugita, S. Pollen representation of vegetation in Quaternary sediments: Theory and method in patchy vegetation. J. Ecol. 1994, 82, 881–897. [Google Scholar] [CrossRef]
- Bacilieri, R.; Labbe, T.; Kremer, R. Intraspecific genetic structure in a mixed population of Quercus petraea (Matt.) Liebl and Q. robur L. Heredity 1994, 73, 130–141. [Google Scholar] [CrossRef]
- Joung, Y.H.I.; Roh, M.S. Mapping characterization of Pinus sylvestris var. silvestriformis based on chloroplast DNA microsatellite markers. For. Genet. 2005, 12, 89–97. [Google Scholar]
- Acheré, V.; Favre, J.M.; Besnard, S.; Jeandroz, S. Genomic organization of molecular differentiation in Norway spruce (Picea abies). Mol. Ecol. 2005, 14, 3191–3201. [Google Scholar] [CrossRef] [PubMed]
- Lind-Riehl, J.F.; Sullivan, A.R.; Gailing, O. Evidence for selection on a CONSTANS-like gene between two red oak species. Ann. Bot. 2014, 113, 967–975. [Google Scholar] [CrossRef] [PubMed]
- Bock, D.G.; Andrew, R.L.; Seberg, L.H. On the adaptive value of cytoplasmic genomes in plants. Mol. Ecol. 2014, 23, 4899–4911. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, M.; Rau, D.; Angioi, S.A.; Bellucci, E.; Bitocchi, E.; Nanni, L.; Knüpffer, H.; Negri, V.; Papa, R.; Attene, G. European Phaseolus coccineus L. landraces: Population structure and adaptation, as revealed by cpSSRs and phenotypic analyses. PLoS ONE 2013, 8, e57337. [Google Scholar] [CrossRef] [PubMed]
- Latta, R.G.; Linhart, Y.B.; Mitton, J.B. Cytonuclear disequilibrium and genetic drift in a natural population of Ponderosa pine. Genetics 2001, 158, 843–850. [Google Scholar] [PubMed]
- Laroche, J.; Li, P.; Maggia, L.; Bousquet, J. Molecular evolution of angiosperm mitochondrial introns and exons. Proc. Natl. Acad. Sci. USA 1997, 94, 5722–5727. [Google Scholar] [CrossRef] [PubMed]
- Jaramillo-Correa, J.P.; Bousquet, J.; Beaulieu, J.; Isabel, N.; Perron, M.; Bouille, M. Cross-species amplification of mitochondrial DNA sequence-tagged-site markers in conifers: The nature of polymorphism and variation within and among species in Picea. Theor. Appl. Genet. 2003, 106, 1353–1367. [Google Scholar] [CrossRef] [PubMed]
- Burban, C.; Petit, R.J. Phylogeography of maritime pine inferred with organelle markers having contrasted inheritance. Mol. Ecol. 2003, 12, 1487–1495. [Google Scholar] [CrossRef] [PubMed]
- Hamrick, J.L.; Godt, M.J.W.; Sherman-Broyles, S.L. Factors influencing levels of genetic diversity in woody plant species. New For. 1992, 6, 95–124. [Google Scholar] [CrossRef]
- Burczyk, J.; DiFazio, S.P.; Adams, W.T. Gene flow in forest trees: How far do genes really travel? For. Genet. 2004, 1, 179–192. [Google Scholar]
- Vidyakin, A.I.; Semerikov, V.L.; Polezhaeva, M.A.; Dymshakova, O.S. Spread of mitochondrial DNA haplotypes in population of Scots pine (Pinus sylvestris L.) in northern European Russia. Russ. J. Genet. 2012, 48, 1267–1271. [Google Scholar] [CrossRef]
- Latta, R.G.; Linhart, Y.B.; Fleck, D.; Elliot, M. Direct and indirect estimates of seed versus pollen movement within a population of Ponderosa pine. Evolution 1998, 52, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Wojnicka-Półtorak, A.; Celiński, K.; Chudzińska, E. Temporal dynamics in the genetic structure of a natural population of Picea abies. Biologia 2016, 71, 875–884. [Google Scholar] [CrossRef]
- Chung, M.Y.; Epperson, B.K.; Chung, M.G. Genetic structure of age classes in Camellia japonica (Theaceae). Evolution 2003, 57, 62–73. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Locus | Range | Number of Alleles | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Total | E | S | Y | M | O | Mean | Total H | ||
| Number of individuals | 156 | 31 | 32 | 32 | 29 | 32 | |||
| cpSSR | |||||||||
| Total | 48 | 31 | 36 | 36 | 36 | 34 | 9.6 | 0.393 | |
| Pt 15169 | 123–130 | 8 | 4 | 7 (1) | 6 | 6 (1) | 5 | 5.6 | 0.734 |
| Pt 26081 | 105–108 | 4 | 3 (1) | 3 | 3 | 3 | 3 | 3.0 | 0.405 |
| Pt 30204 | 139–146 | 8 | 5 | 6 | 7 (1) | 6 (1) | 6 | 6.0 | 0.719 |
| Pt 36480 | 141–151 | 3 | 1 | 2 | 2 | 3 (1) | 2 | 2.0 | 0.130 |
| Pt 45002 | 165–170 | 4 | 3 | 4 (1) | 3 | 3 | 3 | 3.2 | 0.472 |
| Pt 71936 | 143–147 | 5 | 4 | 4 | 5 | 5 | 5 | 4.6 | 0.666 |
| PCP 1289 | 102–106 | 5 | 3 | 3 | 4 (1) | 4 (1) | 3 | 3.4 | 0.415 |
| PCP 41131 | 134–139 | 4 | 3 | 3 | 2 | 1 | 3 | 2.4 | 0.096 |
| PCP 87314 | 111–114 | 4 | 4 (1) | 3 | 3 | 3 | 2 | 3.0 | 0.271 |
| PCP 102652 | 113–115 | 3 | 1 | 1 | 1 | 2 (1) | 2 (1) | 1.4 | 0.025 |
| mt markers | |||||||||
| Total | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 0.188 | |
| nad 1 | 217 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0.000 |
| nad 7 | 1170 and 1175 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 0.375 |
| cp and mt | - | 51 | 34 | 39 | 39 | 39 | 37 | 10.2 | 0.358 |
| Locus | E Embryos | S 1–3 | Y 10–20 | M 40–80 | O >100 | Mean |
|---|---|---|---|---|---|---|
| cpSSR | ||||||
| Number of alleles per locus Na | 3.1 | 3.6 | 3.6 | 3.6 | 3.4 | 3.5 |
| Effective number of alleles per locus Ne | 2.0 | 2.0 | 2.2 | 2.0 | 2.0 | 2.0 |
| Number of haplotypes | 29 (22) | 30 (19) | 31 (19) | 23 (13) | 32 (19) | 29 |
| Percentage of polymorphic loci | 80% | 90% | 90% | 90% | 100% | 90% |
| mt markers | ||||||
| Number of alleles per locus Na | 1.5 [2] | 1.5 [2] | 1.5 [2] | 1.5 [2] | 1.5 [2] | 1.5 [2] |
| Effective number of alleles per locus Ne | 1.4 [1.7] | 1.4 [1.6] | 1.2 [1.5] | 1.2 [1.5] | 1.5 [1.8] | 1.3 [1.6] |
| Number of mitotypes | 2 | 2 | 2 | 2 | 2 | 2 |
| Frequency of mitotypes | ||||||
| a | 0.70 | 0.75 | 0.80 | 0.79 | 0.65 | 0.738 |
| b | 0.30 | 0.25 | 0.20 | 0.21 | 0.36 | 0.264 |
| Percentage of polymorphic loci | 50% | 50% | 50% | 50% | 50% | 50% |
| Source | d.f. | Sum of Squares | Variance Components | Percentage of Variation |
|---|---|---|---|---|
| Among age classes | 4 | 35.893 | 0.106 | 2% (p = 0.027) |
| Within age classes | 196 | 877.589 | 5.519 | 98% |
| Total | 200 | 913.482 | 5.625 | 100% |
| Locus | PhiPT | p |
|---|---|---|
| cpSSR | ||
| Total | 0.018 | 0.037 |
| Pt 15169 | −0.020 | 0.865 |
| Pt 26081 | −0.001 | 0.434 |
| Pt 30204 | 0.068 | 0.009 |
| Pt 36480 | −0.004 | 0.630 |
| Pt 45002 | 0.044 | 0.035 |
| Pt 71936 | 0.014 | 0.216 |
| PCP 1289 | −0.024 | 0.944 |
| PCP 41131 | 0.014 | 0.176 |
| PCP 87314 | −0.004 | 0.452 |
| PCP 102652 | 0.004 | 0.186 |
| mt markers | ||
| Total | −0.005 | 0.560 |
| nad1 | - | - |
| nad7 | −0.005 | 0.537 |
| E | S | Y | M | O | |
|---|---|---|---|---|---|
| 0.013 | 0.011 | 0.016 | 0.021 | E | |
| 0.022 | 0.018 | 0.017 | 0.019 | S | |
| 0.000 | 0.008 | 0.023 | 0.029 | Y | |
| 0.000 | 0.022 | 0.000 | 0.027 | M | |
| 0.084 *** | 0.002 | 0.058 ** | 0.051 ** | O |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wojnicka-Półtorak, A.; Celiński, K.; Chudzińska, E. Genetic Diversity among Age Classes of a Pinus sylvestris (L.) Population from the Białowieża Primeval Forest, Poland. Forests 2017, 8, 227. https://doi.org/10.3390/f8070227
Wojnicka-Półtorak A, Celiński K, Chudzińska E. Genetic Diversity among Age Classes of a Pinus sylvestris (L.) Population from the Białowieża Primeval Forest, Poland. Forests. 2017; 8(7):227. https://doi.org/10.3390/f8070227
Chicago/Turabian StyleWojnicka-Półtorak, Aleksandra, Konrad Celiński, and Ewa Chudzińska. 2017. "Genetic Diversity among Age Classes of a Pinus sylvestris (L.) Population from the Białowieża Primeval Forest, Poland" Forests 8, no. 7: 227. https://doi.org/10.3390/f8070227
APA StyleWojnicka-Półtorak, A., Celiński, K., & Chudzińska, E. (2017). Genetic Diversity among Age Classes of a Pinus sylvestris (L.) Population from the Białowieża Primeval Forest, Poland. Forests, 8(7), 227. https://doi.org/10.3390/f8070227