
The Effects of Climate Change on the Development of Tree Plantations for Biodiesel Production in China



Abstract
:1. Introduction
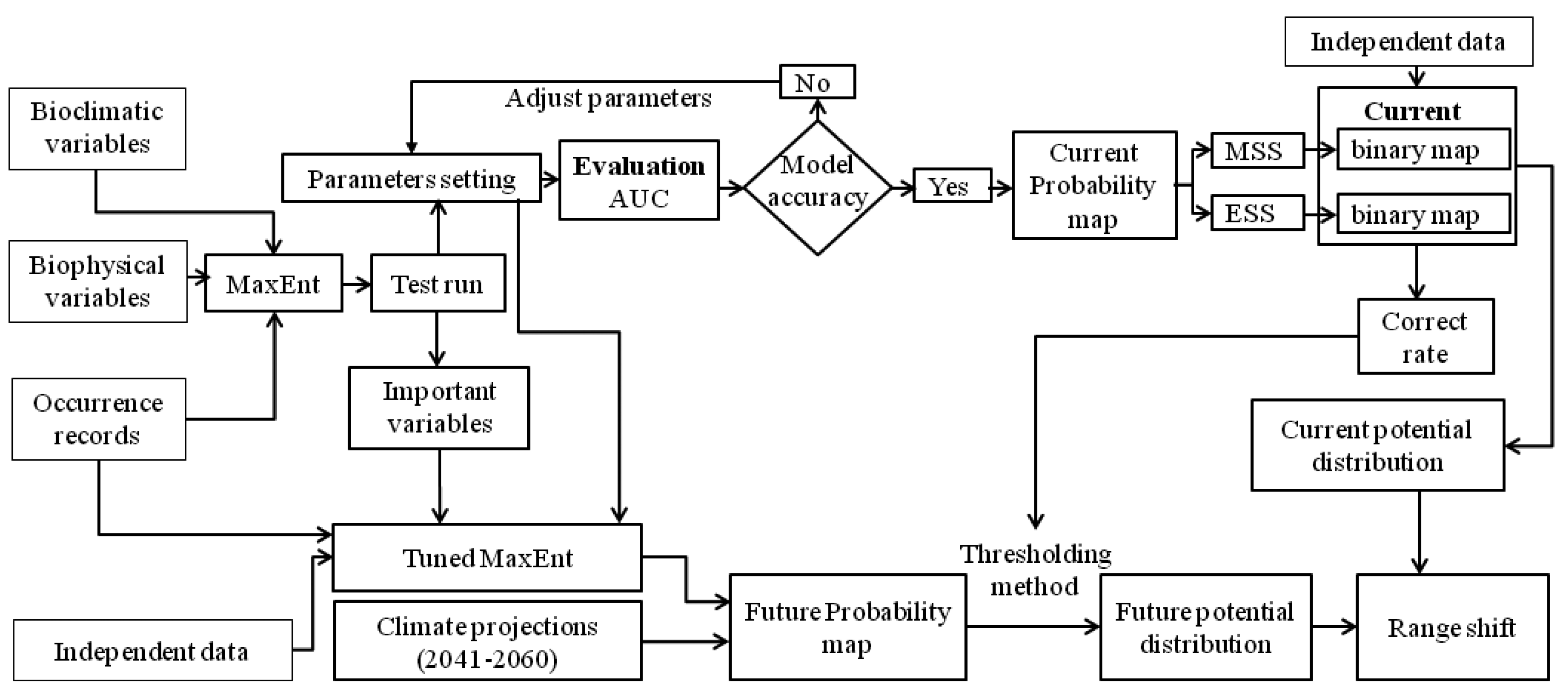
2. Methods
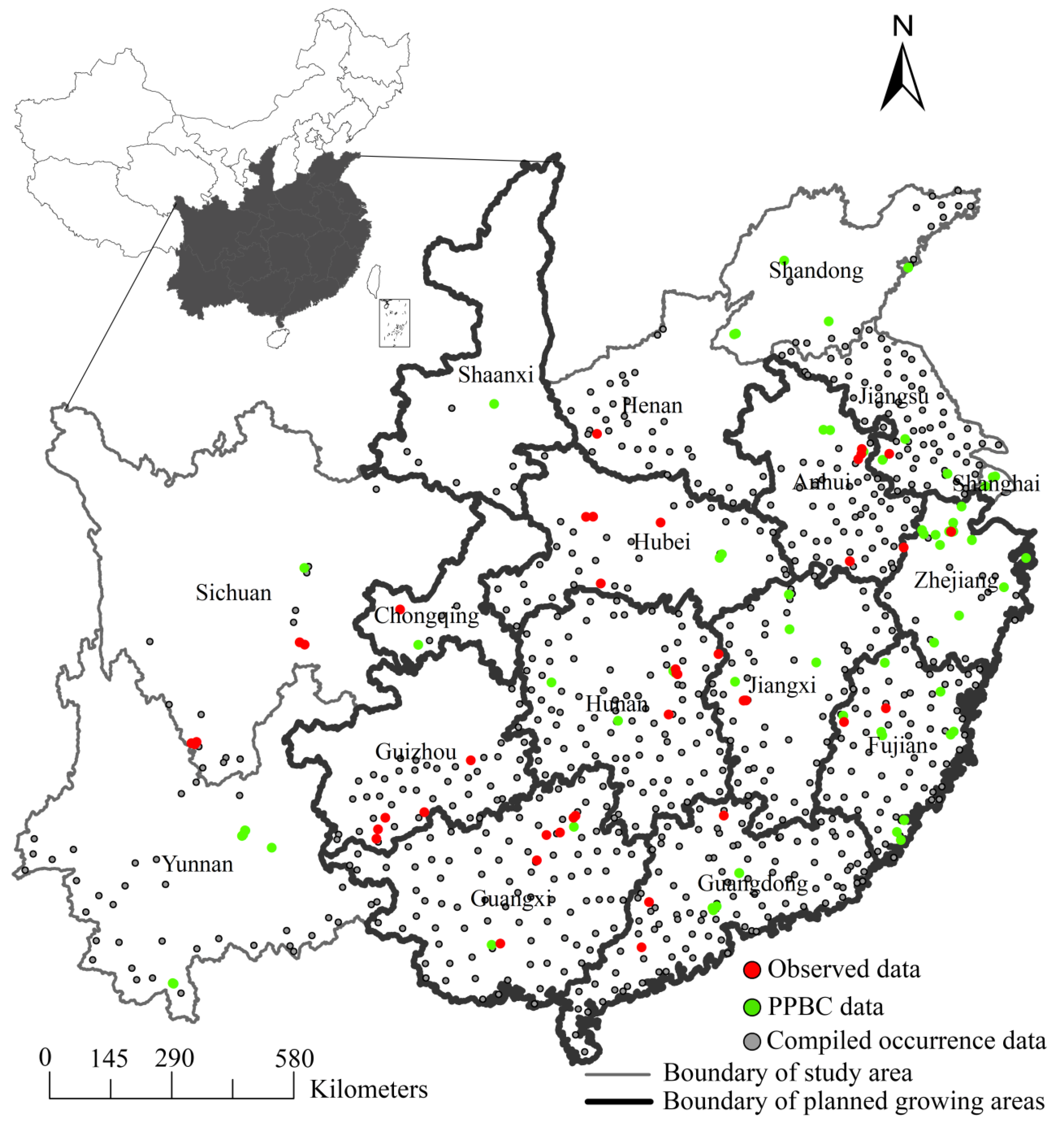
2.1. Compilation of Occurrence Data
2.2. Climatic and Environmental Inputs
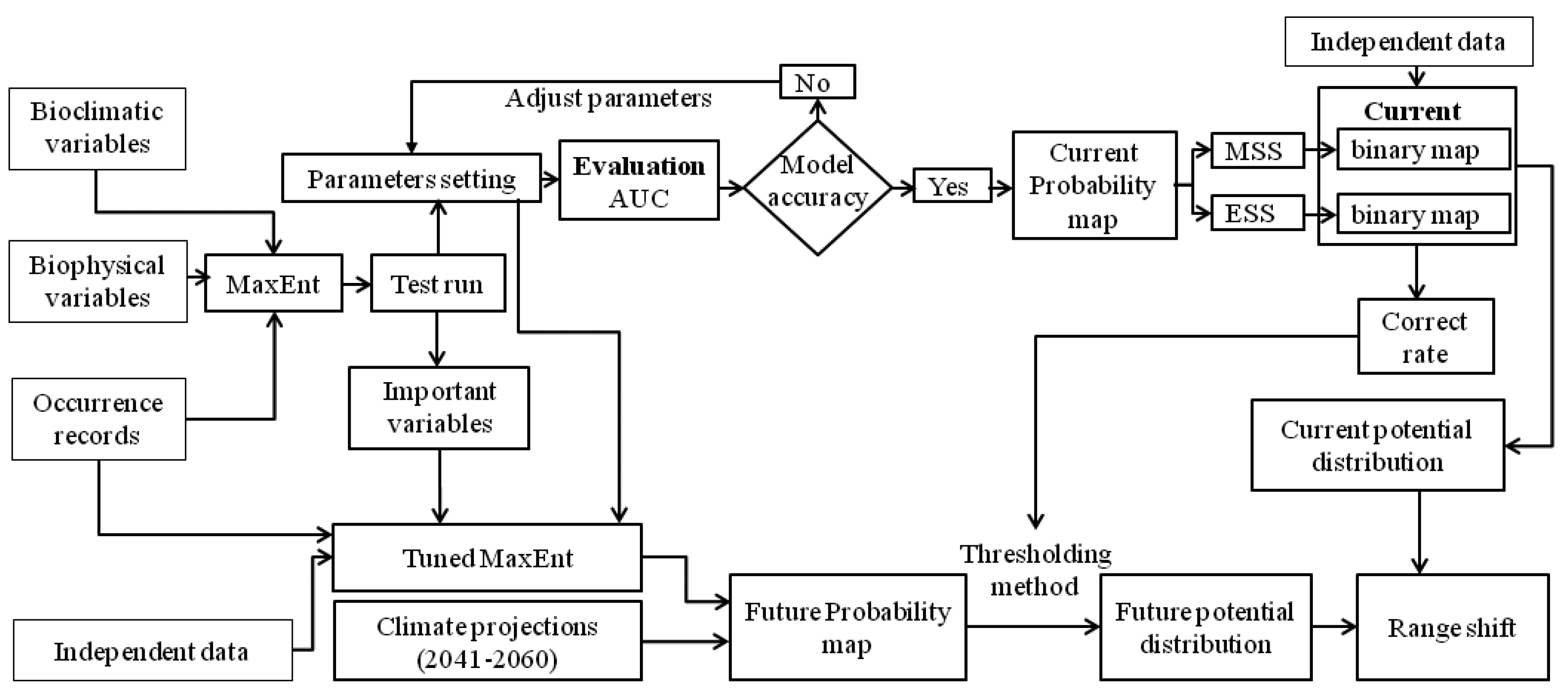
2.3. Model Simulation
2.4. Analysis of Model Outputs
3. Results
3.1. Selection of Model Variables and the Modeling Accuracy
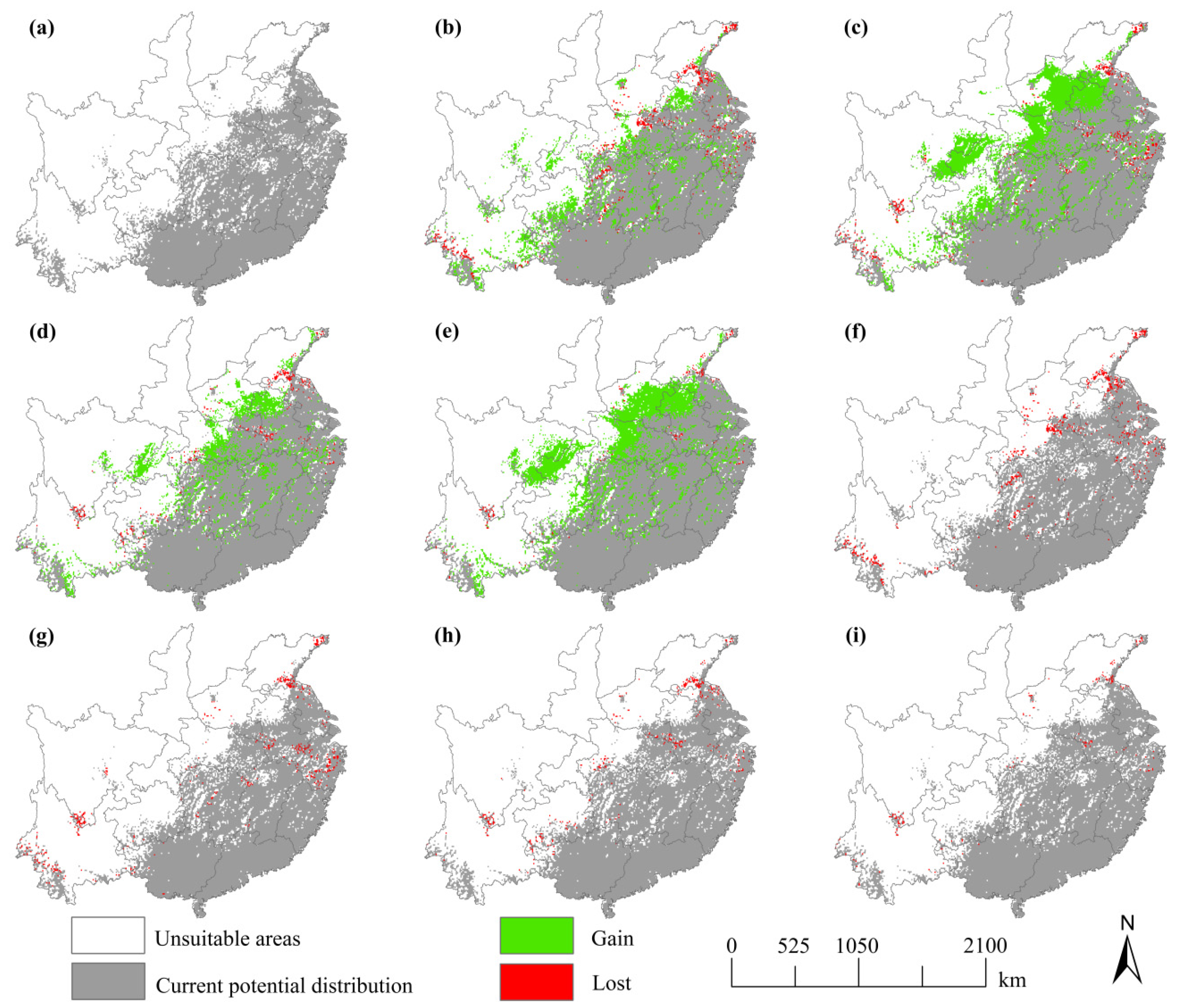
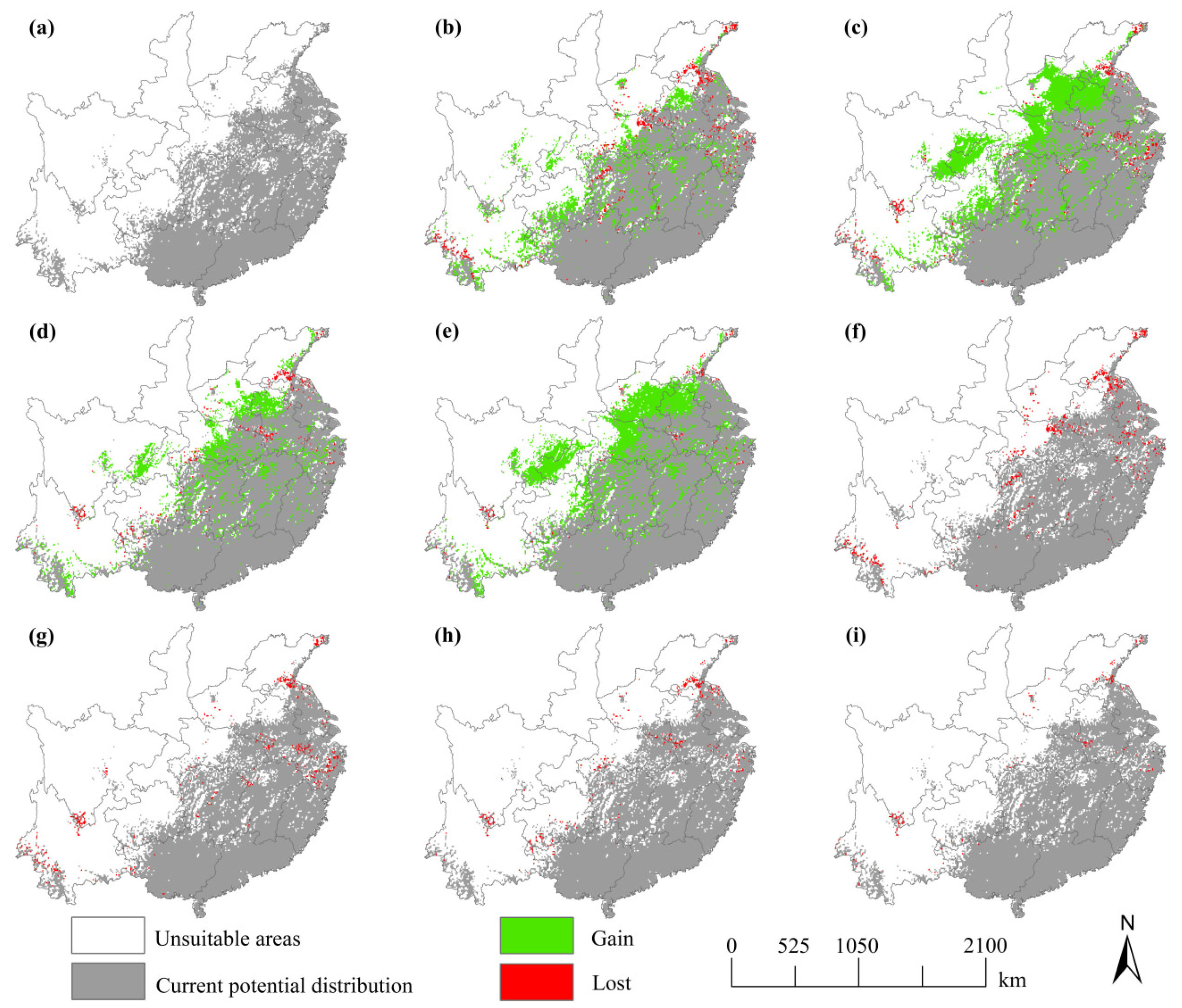
3.2. Current Potential Distributions of S. mukorossi
3.3. Future Distribution of S. mukorossi
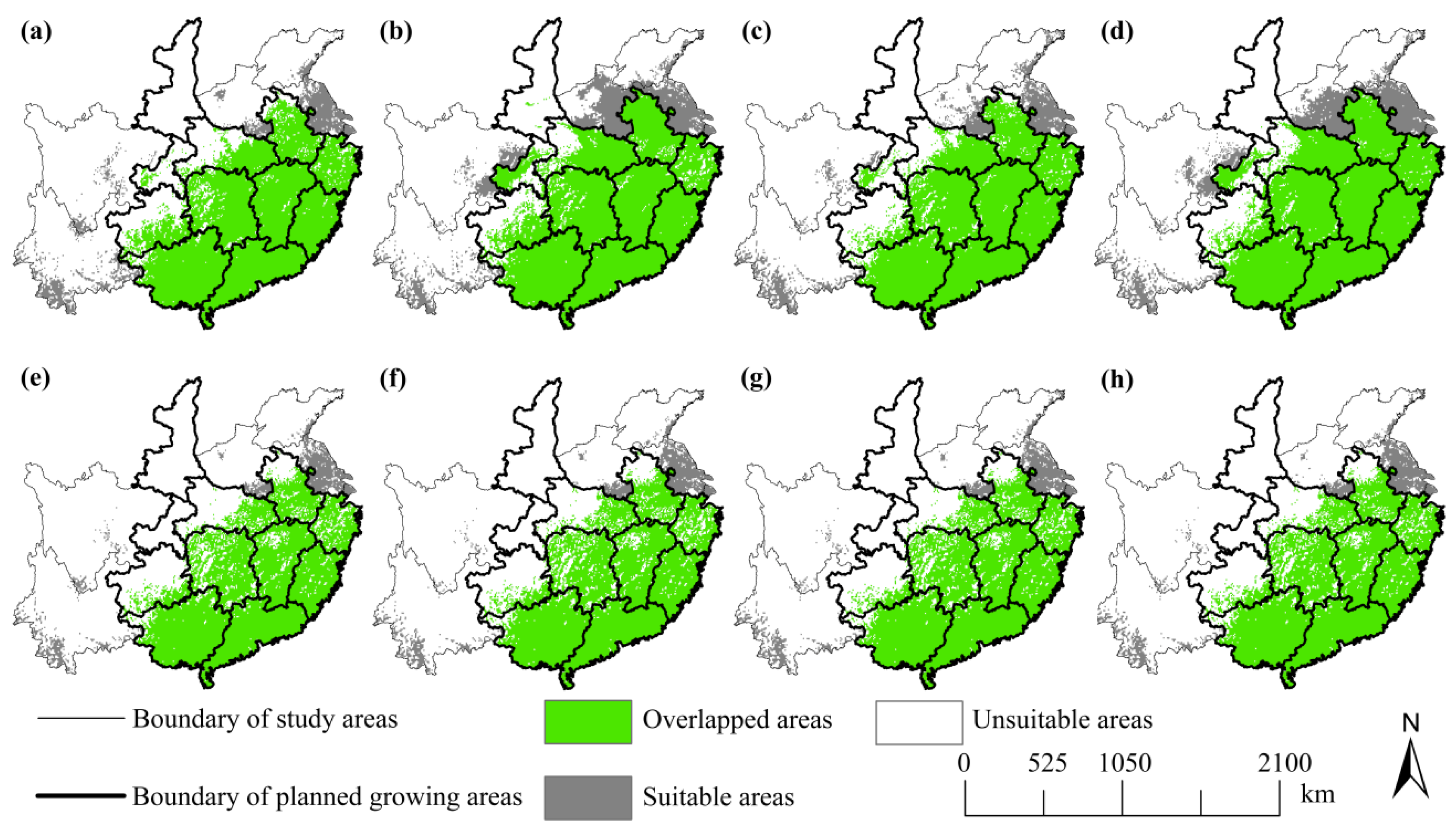
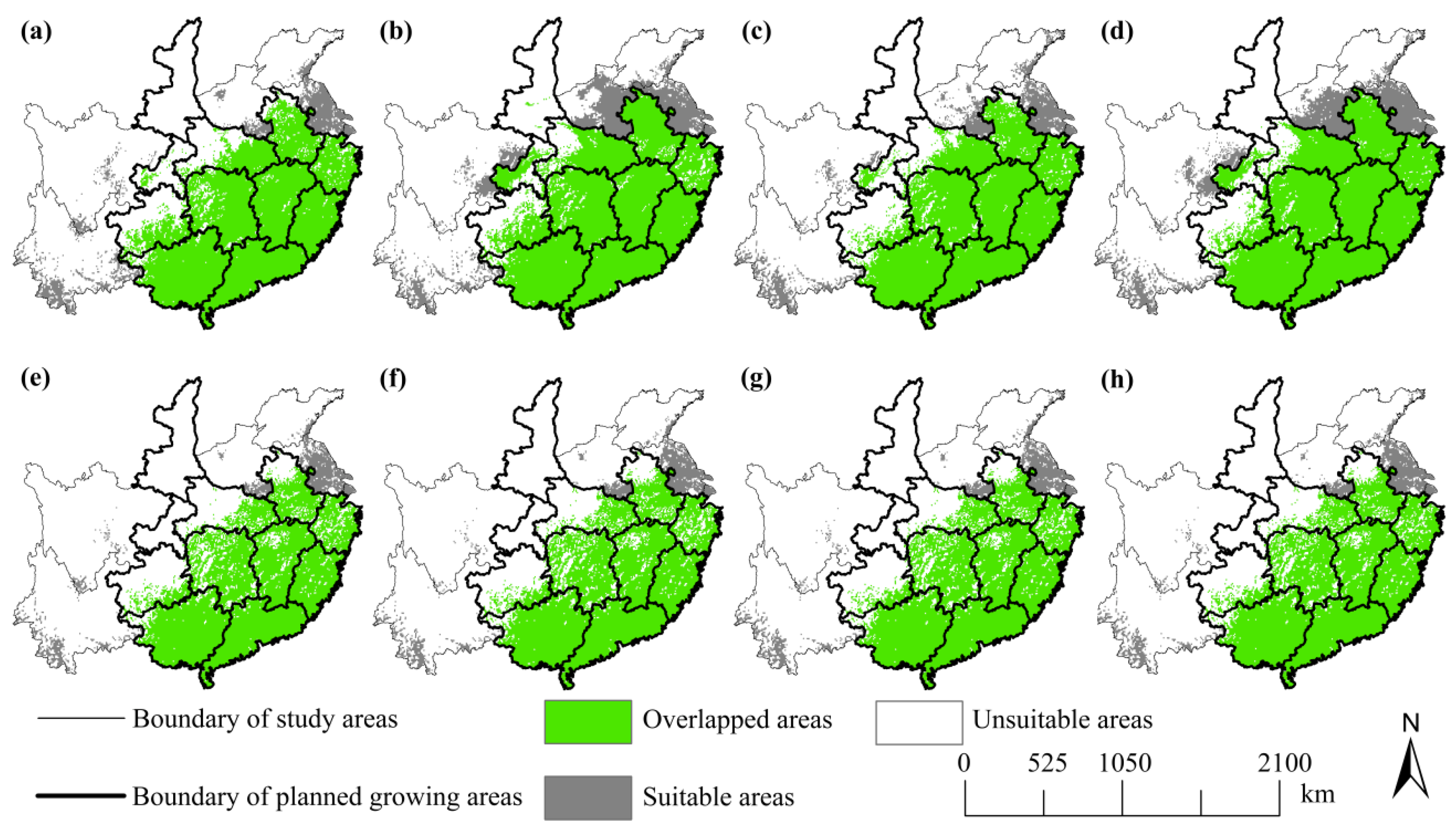
3.4. Comparison between the SFA’s Plan and Future Distributions
4. Discussion
4.1. Changes of the Distribution Range of S. mukorossi
4.2. Uncertainties in Projections
4.3. Implications for Forest-Based Bioenergy Planning
4.4. Limitations of the Current Study
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Tambo, E.; Wang, D.Q.; Zhou, X.N. Tackling air pollution and extreme climate changes in china: Implementing the paris climate change agreement. Environ. Int. 2016, 95, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Wagner, L.; Ross, I.; Foster, J.; Hankamer, B. Trading off global fuel supply, co2 emissions and sustainable development. PLoS ONE 2016, 11, e0149406. [Google Scholar] [CrossRef] [PubMed]
- Repo, A.; Tuovinen, J.-P.; Liski, J. Can we produce carbon and climate neutral forest bioenergy? GCB Bioenergy 2015, 7, 253–262. [Google Scholar] [CrossRef]
- Tilman, D.; Socolow, R.; Foley, J.A.; Hill, J.; Larson, E.; Lynd, L.; Pacala, S.; Reilly, J.; Searchinger, T.; Somerville, C.; et al. Beneficial biofuels—the food, energy, and environment trilemma. Science 2009, 325, 270–271. [Google Scholar] [CrossRef] [PubMed]
- Rowe, R.L.; Hanley, M.E.; Goulson, D.; Clarke, D.J.; Doncaster, C.P.; Taylor, G. Potential benefits of commercial willow short rotation coppice (src) for farm-scale plant and invertebrate communities in the agri-environment. Biomass Bioenergy 2011, 35, 325–336. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, G.; Xu, K.; Li, Z. Developing forest bioenergy and pushing up degraded lands amelioration in china. Energy Educ. Sci. Technol. Part A 2012, 30, 1089–1094. [Google Scholar]
- Kumar, V.; Kant, P. Biodiesel: Beneficial for environment and human health. Pet. Coal 2013, 55, 207–215. [Google Scholar]
- Sundstrom, S.; Nielsen-Pincus, M.; Moseley, C.; McCaffery, S. Woody biomass use trends, barriers, and strategies: Perspectives of us forest service managers. J. For. 2012, 110, 16–24. [Google Scholar] [CrossRef]
- Moiseyev, A.; Solberg, B.; Kallio, A.M.I.; Lindner, M. An economic analysis of the potential contribution of forest biomass to the eu res target and its implications for the eu forest industries. J. For. Econ. 2011, 17, 197–213. [Google Scholar] [CrossRef]
- Yang, J.; Dai, G.; Ma, L.; Jia, L.; Wu, J.; Wang, X. Forest-based bioenergy in china: Status, opportunities, and challenges. Renew. Sustain. Energy Rev. 2013, 18, 478–485. [Google Scholar] [CrossRef]
- Yin, F.S.; Teah, Y.K. Status of bio-diesel industry in china. Deterg. Cosmet. 2008, 31, 1–3. [Google Scholar]
- Yang, C.Y.; Fang, Z.; Li, B.; Long, Y.F. Review and prospects of jatropha biodiesel industry in china. Renew. Sustain. Energy Rev. 2012, 16, 2178–2190. [Google Scholar] [CrossRef]
- Stocker, T.F.; Qin, D.; Plattner, G.-K.; Alexander, L.V.; Allen, S.K.; Bindoff, N.L.; Bréon, F.-M.; Church, J.A.; Cubasch, U.; Emori, S.; et al. Technical summary. In Climate Change 2013: The Physical Science Basis: Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; pp. 33–115. [Google Scholar]
- Yao, Z.Y.; Qi, J.H.; Yin, L.M. Biodiesel production from xanthoceras sorbifolia in china: Opportunities and challenges. Renew. Sustain. Energy Rev. 2013, 24, 57–65. [Google Scholar] [CrossRef]
- Francis, G.; Edinger, R.; Becker, K. A concept for simultaneous wasteland reclamation, fuel production, and socio-economic development in degraded areas in india: Need, potential and perspectives of jatropha plantations. Nat. Resour. Forum 2005, 29, 12–24. [Google Scholar] [CrossRef]
- Kriticos, D.J.; Murphy, H.T.; Jovanovic, T.; Taylor, J.; Herr, A.; Raison, J.; O’Connell, D. Balancing bioenergy and biosecurity policies: Estimating current and future climate suitability patterns for a bioenergy crop. GCB Bioenergy 2014, 6, 587–598. [Google Scholar] [CrossRef]
- Wang, W.G.; Tang, X.Y.; Zhu, Q.L.; Pan, K.; Hu, Q.C.; He, M.X.; Li, J.T. Predicting the impacts of climate change on the potential distribution of major native non-food bioenergy plants in china. PLoS ONE 2014, 9, e111587. [Google Scholar] [CrossRef] [PubMed]
- Trabucco, A.; Achten, W.M.J.; Bowe, C.; Aerts, R.; Van Orshoven, J.; Norgrove, L.; Muys, B. Global mapping of jatropha curcas yield based on response of fitness to present and future climate. GCB Bioenergy 2010, 2, 139–151. [Google Scholar] [CrossRef]
- Thuiller, W.; Lavergne, S.; Roquet, C.; Boulangeat, I.; Lafourcade, B.; Araujo, M.B. Consequences of climate change on the tree of life in Europe. Nature 2011, 470, 531–534. [Google Scholar] [CrossRef] [PubMed]
- Araújo, M.B.; New, M. Ensemble forecasting of species distributions. Trends Ecol. Evol. 2007, 22, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Diniz-Filho, J.A.; Bini, L.M.; Rangel, T.F.; Loyola, R.D.; Hof, C.; Nogués-Bravo, D.; Araújo, M.B. Partitioning and mapping uncertainties in ensembles of forecasts of species turnover under climate change. Ecography 2009, 32, 897–906. [Google Scholar] [CrossRef]
- Buisson, L.; Thuiller, W.; Casajus, N.; Lek, S.; Grenouillet, G. Uncertainty in ensemble forecasting of species distribution. Glob. Chang. Biol. 2010, 16, 1145–1157. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H. Do they? How do they? Why do they differ? On finding reasons for differing performances of species distribution models. Ecography 2009, 32, 66–77. [Google Scholar] [CrossRef]
- Real, R.; Márquez, A.L.; Olivero, J.; Estrada, A. Species distribution models in climate change scenarios are still not useful for informing policy planning: An uncertainty assessment using fuzzy logic. Ecography 2010, 33, 304–314. [Google Scholar] [CrossRef]
- Wang, T.; Campbell, E.M.; O’Neill, G.A.; Aitken, S.N. Projecting future distributions of ecosystem climate niches: Uncertainties and management applications. For. Ecol. Manag. 2012, 279, 128–140. [Google Scholar] [CrossRef]
- Wright, A.N.; Hijmans, R.J.; Schwartz, M.W.; Shaffer, H.B. Multiple sources of uncertainty affect metrics for ranking conservation risk under climate change. Divers. Distrib. 2015, 21, 111–122. [Google Scholar] [CrossRef]
- Lindner, M.; Fitzgerald, J.B.; Zimmermann, N.E.; Reyer, C.; Delzon, S.; van der Maaten, E.; Schelhaas, M.J.; Lasch, P.; Eggers, J.; van der Maaten-Theunissen, M.; et al. Climate change and european forests: What do we know, what are the uncertainties, and what are the implications for forest management? J. Environ. Manag. 2014, 146, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Fu, D.J.; Huang, H.W. Brief introduction of expoitation and utilization of fuel plants resources. J. Wuhan Bot. Res. 2006, 24, 183–190. [Google Scholar]
- Zhuang, D.F.; Jiang, D.; Liu, L.; Huang, Y.H. Assessment of bioenergy potential on marginal land in China. Renew. Sustain. Energy Rev. 2011, 15, 1050–1056. [Google Scholar] [CrossRef]
- State Forestry Administration (SFA). The National Forestry Biomass Energy Development Plan. Available online: http://www.forestry.gov.cn//portal/main/s/218/content-633246.html (accessed on 4 September 2014).
- Phillips, S.J.; Dudik, M.; Elith, J.; Graham, C.H.; Lehmann, A.; Leathwick, J.; Ferrier, S. Sample selection bias and presence-only distribution models: Implications for background and pseudo-absence data. Ecol. Appl. 2009, 19, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.J.; Ma, K.P.; Kreft, H. Geographical sampling bias in a large distributional database and its effects on species richness-environment models. J. Biogeogr. 2013, 40, 1415–1426. [Google Scholar] [CrossRef]
- Chinese Academy of Science (CAS). Plant Photo Bank of China (PPBC). Available online: http://www.plantphoto.cn (accessed on 15 October 2014).
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (FAO)/International Institute for Applied Systems Analysis (IIASA)/International Soil Reference and Information Centre (ISRIC)/Institute of Soil Science–Chinese Academy of Sciences (ISSCAS)/Joint Research Centre of the European Commission (JRC). Harmonized World Soil Database (Version 1.2). Available online: http://webarchive.iiasa.ac.at/Research/LUC/External-World-soil-database/HTML/ (accessed on 2 September 2013).
- Iverson, L.R.; Prasad, A.M. Predicting abundance of 80 tree species following climate change in the eastern united states. Ecol. Monogr. 1998, 68, 465–485. [Google Scholar] [CrossRef]
- Cruz-Cárdenas, G.; Villaseñor, J.L.; López-Mata, L.; Ortiz, E. Potential distribution of humid mountain forest in mexico. Bot. Sci. 2012, 90, 331–340. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A. A practical guide to maxent for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of maxent for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Garcia, K.; Lasco, R.; Ines, A.; Lyon, B.; Pulhin, F. Predicting geographic distribution and habitat suitability due to climate change of selected threatened forest tree species in the philippines. Appl. Geogr. 2013, 44, 12–22. [Google Scholar] [CrossRef]
- Hosmer, D.W.; Lemeshow, S. Applied Logistic Regression; Wiley: New York, NY, USA, 1989. [Google Scholar]
- Kramer-Schadt, S.; Niedballa, J.; Pilgrim, J.D.; Schröder, B.; Lindenborn, J.; Reinfelder, V.; Stillfried, M.; Heckmann, I.; Scharf, A.K.; Augeri, D.M.; et al. The importance of correcting for sampling bias in maxent species distribution models. Divers. Distrib. 2013, 19, 1366–1379. [Google Scholar] [CrossRef]
- Jiménez-Valverde, A.; Lobo, J.M. Threshold criteria for conversion of probability of species presence to either-or presence-absence. Acta Oecol 2007, 31, 361–369. [Google Scholar] [CrossRef]
- Fordham, D.A.; Wigley, T.M.L.; Brook, B.W. Multi-model climate projections for biodiversity risk assessments. Ecol Appl. 2011, 21, 3317–3331. [Google Scholar] [CrossRef]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; Ferreira de Siqueira, M.; Grainger, A.; Hannah, L.; et al. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Van Vliet, J.; Bregt, A.K.; Hagen-Zanker, A. Revisiting kappa to account for change in the accuracy assessment of land-use change models. Ecol Model 2011, 222, 1367–1375. [Google Scholar] [CrossRef]
- Walther, G.R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.D. Climate, climate change and range boundaries. Divers. Distrib. 2010, 16, 488–495. [Google Scholar] [CrossRef]
- Pattison, R.R.; Mack, R.N. Potential distribution of the invasive tree triadica sebifera (euphorbiaceae) in the united states: Evaluating climex predictions with field trials. Glob Chang. Biol 2008, 14, 813–826. [Google Scholar] [CrossRef]
- Tanasijevic, L.; Todorovic, M.; Pereira, L.S.; Pizzigalli, C.; Lionello, P. Impacts of climate change on olive crop evapotranspiration and irrigation requirements in the mediterranean region. Agric. Water Manag. 2014, 144, 54–68. [Google Scholar] [CrossRef]
- Moriondo, M.; Trombi, G.; Ferrise, R.; Brandani, G.; Dibari, C.; Ammann, C.M.; Lippi, M.M.; Bindi, M. Olive trees as bio-indicators of climate evolution in the mediterranean basin. Glob. Ecol. Biogeogr. 2013, 22, 818–833. [Google Scholar] [CrossRef]
- Zomer, R.J.; Trabucco, A.; Wang, M.; Lang, R.; Chen, H.; Metzger, M.J.; Smajgl, A.; Beckschäfer, P.; Xu, J. Environmental stratification to model climate change impacts on biodiversity and rubber production in xishuangbanna, yunnan, china. Biol. Conserv. 2014, 170, 264–273. [Google Scholar] [CrossRef]
- Ray, D.; Behera, M.D.; Jacob, J. Indian brahmaputra valley offers significant potential for cultivation of rubber trees under changed climte. CSci 2014, 107, 461–469. [Google Scholar]
- Diao, S.F.; Shao, W.H.; Dong, R.X.; Sun, H.G.; Jiang, X.M. Diurnal variation of photosynthesis and relationship with the eco-physiological factors of sapindus mukorossi. Acta Bot. Boreali-Occident Sin. 2014, 34, 0828–0834. [Google Scholar]
- Jiang, C.C.; Lu, X.K.; Ye, X.F.; Guo, Y.Z.; Jiang, Y.X. Characteristics and cultivation techniques of sapindus mukorossi. Southeast Horticulture 2014, 6, 118–121. [Google Scholar]
- Shao, W.H.; Yue, H.F.; Xu, Y.Q.; Dong, R.X.; Jiang, J.M.; Shen, F.Q.; Zhang, J.Z. Effects of site type on the growth and main economic traits of sapindus mukorossi gaertn. J. Henan Agric. Univ. 2015, 49, 783–786. [Google Scholar]
- Gu, S.H.; Tu, J.L.; Liu, L.Y.; Zhong, G.J.; Bao, T.F. Waterlogging tolerance of native tree species in jiaxing, zhejiang. Technology of Soil and Water Conservation 2015, 5, 7–9. [Google Scholar]
- Wiens, J.A.; Stralberg, D.; Jongsomjit, D.; Howell, C.A.; Snyder, M.A. Niches, models, and climate change: Assessing the assumptions and uncertainties. Proc. Natl. Acad. Sci. USA 2009, 106, 19729–19736. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Chen, F.; Wang, S.H. Distribution area prediction of biofuel crop jatropha cucars L. J. Sichuan Univ. (Nat. Sci. Ed.) 2012, 49, 239–245. [Google Scholar]
- VanDerWal, J.; Shoo, L.P.; Graham, C.; Williams, S.E. Selecting pseudo-absence data for presence-only distribution modeling: How far should you stray from what you know? Ecol. Model. 2009, 220, 589–594. [Google Scholar] [CrossRef]
- Boria, R.A.; Olson, L.E.; Goodman, S.M.; Anderson, R.P. Spatial filtering to reduce sampling bias can improve the performance of ecological niche models. Ecol. Model. 2014, 275, 73–77. [Google Scholar] [CrossRef]
- Nenzén, H.K.; Araújo, M.B. Choice of threshold alters projections of species range shifts under climate change. Ecol. Model. 2011, 222, 3346–3354. [Google Scholar] [CrossRef]
- Dennis, R.L.H.; Thomas, C.D. Bias in butterfly distribution maps: The influence of hot spots and recorder’s home range. J. Insect Conserv. 2000, 4, 73–77. [Google Scholar] [CrossRef]
- Goberville, E.; Beaugrand, G.; Hautekeete, N.C.; Piquot, Y.; Luczak, C. Uncertainties in the projection of species distributions related to general circulation models. Ecol. Evol. 2015, 5, 1100–1116. [Google Scholar] [CrossRef] [PubMed]
- Visconti, P.; Bakkenes, M.; Baisero, D.; Brooks, T.; Butchart, S.H.M.; Joppa, L.; Alkemade, R.; Di Marco, M.; Santini, L.; Hoffmann, M.; et al. Projecting global biodiversity indicators under future development scenarios. Conserv. Lett. 2016, 9, 5–13. [Google Scholar] [CrossRef]
- Kosinkova, J.; Doshi, A.; Maire, J.; Ristovski, Z.; Brown, R.; Rainey, T.J. Measuring the regional avaiability of biomass for biofuels and potential for microalgae. Renew. Sustain. Energy Rev. 2015, 49, 1271–1285. [Google Scholar] [CrossRef]
- Puettmann, K.J. Silvicultural challenges and options in the context of global change: "Simple" fixes and opportunities for new management approaches. J. For. 2011, 109, 321–331. [Google Scholar]
- FAO. Climate change to forest management; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013. [Google Scholar]
- Pawson, S.M.; Brin, A.; Brockerhoff, E.G.; Lamb, D.; Payn, T.W.; Paquette, A.; Parrotta, J.A. Plantation forests, climate change and biodiversity. Biodivers Conserv 2013, 22, 1203–1227. [Google Scholar] [CrossRef]
- Garcia-Gonzalo, J.; Borges, J.G.; Palma, J.H.N.; Zubizarreta-Gerendiain, A. A decision support system for management planning of eucalyptus plantations facing climate change. Ann. For. Sci. 2014, 71, 187–199. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Set-Up of the Model | Current | Future | ||
|---|---|---|---|---|
| Test AUC | Training AUC | Test AUC | Training AUC | |
| Full dispersal + average GCMs + RCP2.6 | 0.74 | 0.79 | 0.72 | 0.78 |
| Full dispersal + average GCMs + RCP8.5 | 0.72 | 0.79 | 0.72 | 0.78 |
| Full dispersal + consensus GCMs + RCP2.6 | 0.72 | 0.78 | 0.72 | 0.78 |
| Full dispersal + consensus GCMs + RCP8.5 | 0.72 | 0.79 | 0.72 | 0.78 |
| No dispersal + average GCMs + RCP2.6 | 0.74 | 0.79 | 0.72 | 0.78 |
| No dispersal + average GCMs + RCP8.5 | 0.72 | 0.79 | 0.72 | 0.78 |
| No dispersal + consensus GCMs + RCP2.6 | 0.72 | 0.78 | 0.72 | 0.78 |
| No dispersal + consensus GCMs + RCP8.5 | 0.72 | 0.79 | 0.72 | 0.78 |
| Dispersal Ability | GCMs | Emission Scenario | Lost | Remain | Gain | Net | |||
|---|---|---|---|---|---|---|---|---|---|
| Area (km2) | % | Area (km2) | % | Area (km2) | % | % | |||
| Full dispersal | Consensus GCMs | RCP2.6 | 24,982 | −1.99 | 1,233,473 | 98.01 | 241,925 | 19.22 | 17.23 |
| Consensus GCMs | RCP8.5 | 11,829 | −0.94 | 1,246,626 | 99.06 | 434,513 | 34.53 | 33.59 | |
| Average GCMs | RCP2.6 | 47,724 | −3.79 | 1,210,731 | 96.21 | 230,123 | 18.29 | 14.50 | |
| Average GCMs | RCP8.5 | 36,301 | −2.88 | 1,222,154 | 97.12 | 453,559 | 36.04 | 33.16 | |
| No dispersal | Consensus GCMs | RCP2.6 | 24,982 | −1.99 | 1,233,473 | 98.01 | 0 | 0.00 | −1.99 |
| Consensus GCMs | RCP8.5 | 11,829 | −0.94 | 1,246,626 | 99.06 | 0 | 0.00 | −0.94 | |
| Average GCMs | RCP2.6 | 47,724 | −3.79 | 1,210,731 | 96.21 | 0 | 0.00 | −3.79 | |
| Average GCMs | RCP8.5 | 36,301 | −2.88 | 1,222,154 | 97.12 | 0 | 0.00 | −2.88 | |
| Set-Up of the Model | Full Dispersal + Average GCMs + RCP2.6 | Full Dispersal + Average GCMs + RCP8.5 | Full Dispersal + Consensus GCMs + RCP2.6 | Full Dispersal + Consensus GCMs + RCP8.5 | No Dispersal + Average GCMs + RCP2.6 | No Dispersal + Average GCMs + RCP8.5 | No Dispersal + Consensus GCMs + RCP2.6 | No Dispersal + Consensus GCMs + RCP8.5 |
|---|---|---|---|---|---|---|---|---|
| Full dispersal + average GCMs + RCP2.6 | 1.00 | 0.76 | 0.85 | 0.76 | 0.85 | 0.81 | 0.82 | 0.82 |
| Full dispersal + average GCMs + RCP8.5 | - | 1.00 | 0.81 | 0.86 | 0.68 | 0.71 | 0.69 | 0.69 |
| Full dispersal + consensus GCMs + RCP2.6 | - | - | 1.00 | 0.85 | 0.81 | 0.82 | 0.84 | 0.83 |
| Full dispersal + consensus GCMs + RCP8.5 | - | - | - | 1.00 | 0.69 | 0.71 | 0.71 | 0.73 |
| No dispersal + average GCMs + RCP2.6 | - | - | - | - | 1.00 | 0.96 | 0.97 | 0.97 |
| No dispersal + average GCMs + RCP8.5 | - | - | - | - | - | 1.00 | 0.98 | 0.98 |
| No dispersal + consensus GCMs + RCP2.6 | - | - | - | - | - | - | 1.00 | 0.99 |
| No dispersal + consensus GCMs + RCP8.5 | - | - | - | - | - | - | - | 1.00 |
| Set-Up of the Model | Zhejiang | Anhui | Fujian | Jiangxi | Hubei | Hunan | Guangdong | Guangxi | Chongqing | Guizhou | Shaanxi | Total |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Full dispersal + average GCMs + RCP2.6 | 87.19 | 76.57 | 98.84 | 97.37 | 46.73 | 87.01 | 99.71 | 94.03 | 10.87 | 36.64 | 0.00 | 68.38 |
| Full dispersal + average GCMs + RCP8.5 | 85.74 | 94.23 | 99.22 | 97.05 | 68.13 | 91.67 | 99.86 | 94.77 | 48.84 | 41.51 | 0.75 | 74.75 |
| Full dispersal + consensus GCMs + RCP2.6 | 92.41 | 85.64 | 98.84 | 97.51 | 54.92 | 88.26 | 99.43 | 91.41 | 21.03 | 22.08 | 0.00 | 68.96 |
| Full dispersal + consensus GCMs + RCP8.5 | 96.48 | 96.94 | 99.52 | 99.16 | 68.95 | 93.24 | 99.81 | 93.55 | 46.66 | 32.88 | 0.04 | 74.82 |
| No dispersal + average GCM + RCP2.6 | 77.52 | 59.05 | 94.15 | 85.89 | 31.38 | 71.48 | 98.17 | 89.00 | 0.62 | 19.29 | 0.00 | 58.70 |
| No dispersal + average GCMs + RCP8.5 | 75.64 | 58.51 | 94.28 | 85.27 | 34.25 | 72.90 | 98.22 | 89.16 | 0.71 | 19.26 | 0.00 | 59.00 |
| No dispersal + consensus GCMs + RCP2.6 | 81.02 | 59.07 | 94.35 | 86.15 | 33.46 | 73.24 | 98.22 | 89.20 | 0.71 | 17.79 | 0.00 | 59.23 |
| No dispersal + consensus GCMs + RCP8.5 | 81.96 | 60.24 | 94.35 | 86.19 | 34.34 | 73.66 | 98.22 | 89.36 | 0.71 | 19.33 | 0.00 | 59.69 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, G.; Yang, J.; Huang, C.; Sun, C.; Jia, L.; Ma, L. The Effects of Climate Change on the Development of Tree Plantations for Biodiesel Production in China. Forests 2017, 8, 207. https://doi.org/10.3390/f8060207
Dai G, Yang J, Huang C, Sun C, Jia L, Ma L. The Effects of Climate Change on the Development of Tree Plantations for Biodiesel Production in China. Forests. 2017; 8(6):207. https://doi.org/10.3390/f8060207
Chicago/Turabian StyleDai, Guanghui, Jun Yang, Conghong Huang, Caowen Sun, Liming Jia, and Luyi Ma. 2017. "The Effects of Climate Change on the Development of Tree Plantations for Biodiesel Production in China" Forests 8, no. 6: 207. https://doi.org/10.3390/f8060207
APA StyleDai, G., Yang, J., Huang, C., Sun, C., Jia, L., & Ma, L. (2017). The Effects of Climate Change on the Development of Tree Plantations for Biodiesel Production in China. Forests, 8(6), 207. https://doi.org/10.3390/f8060207