
Drivers of Plot-Scale Variability of CH4 Consumption in a Well-Aerated Pine Forest Soil



Abstract
:1. Introduction
2. Material and Methods
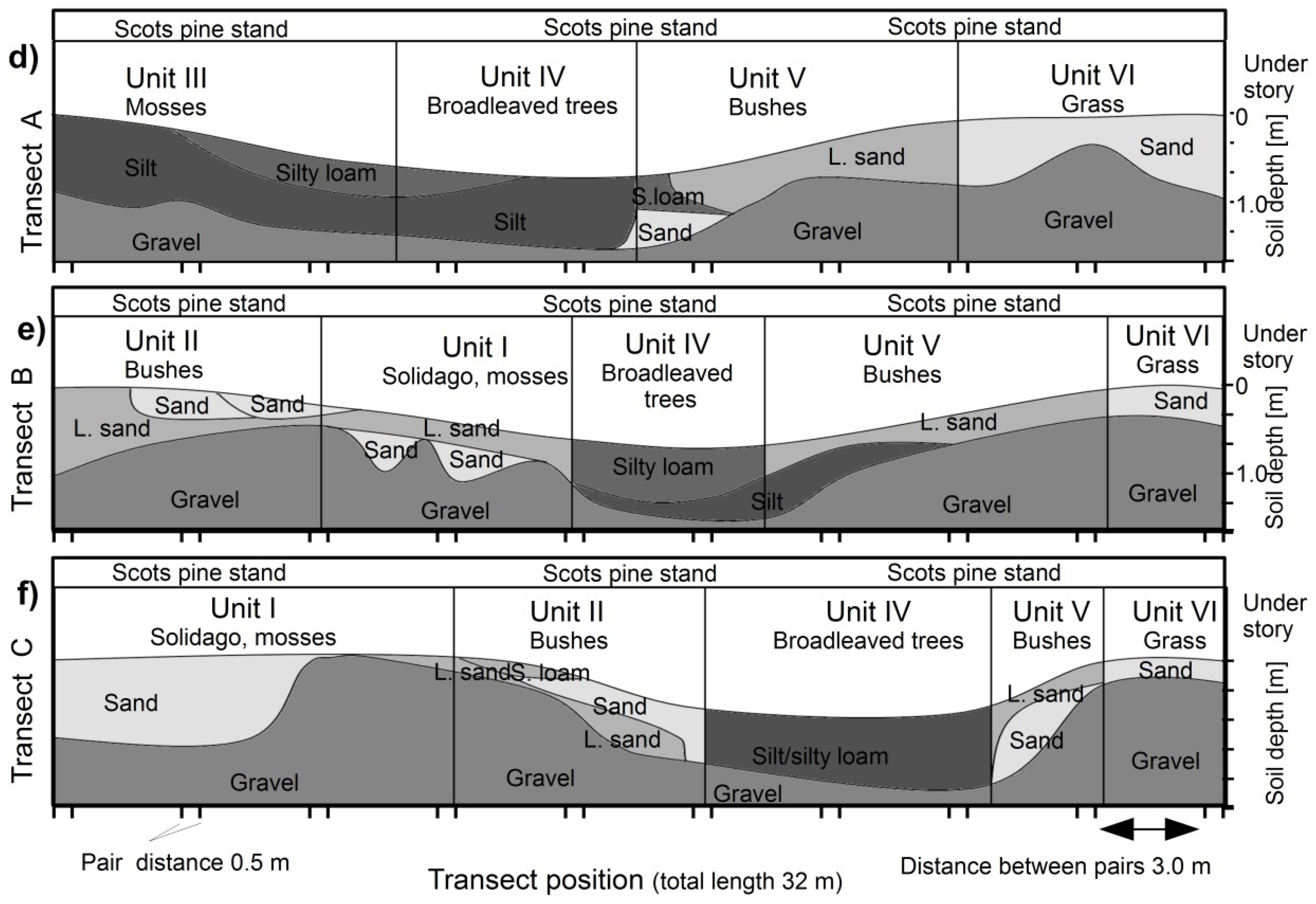
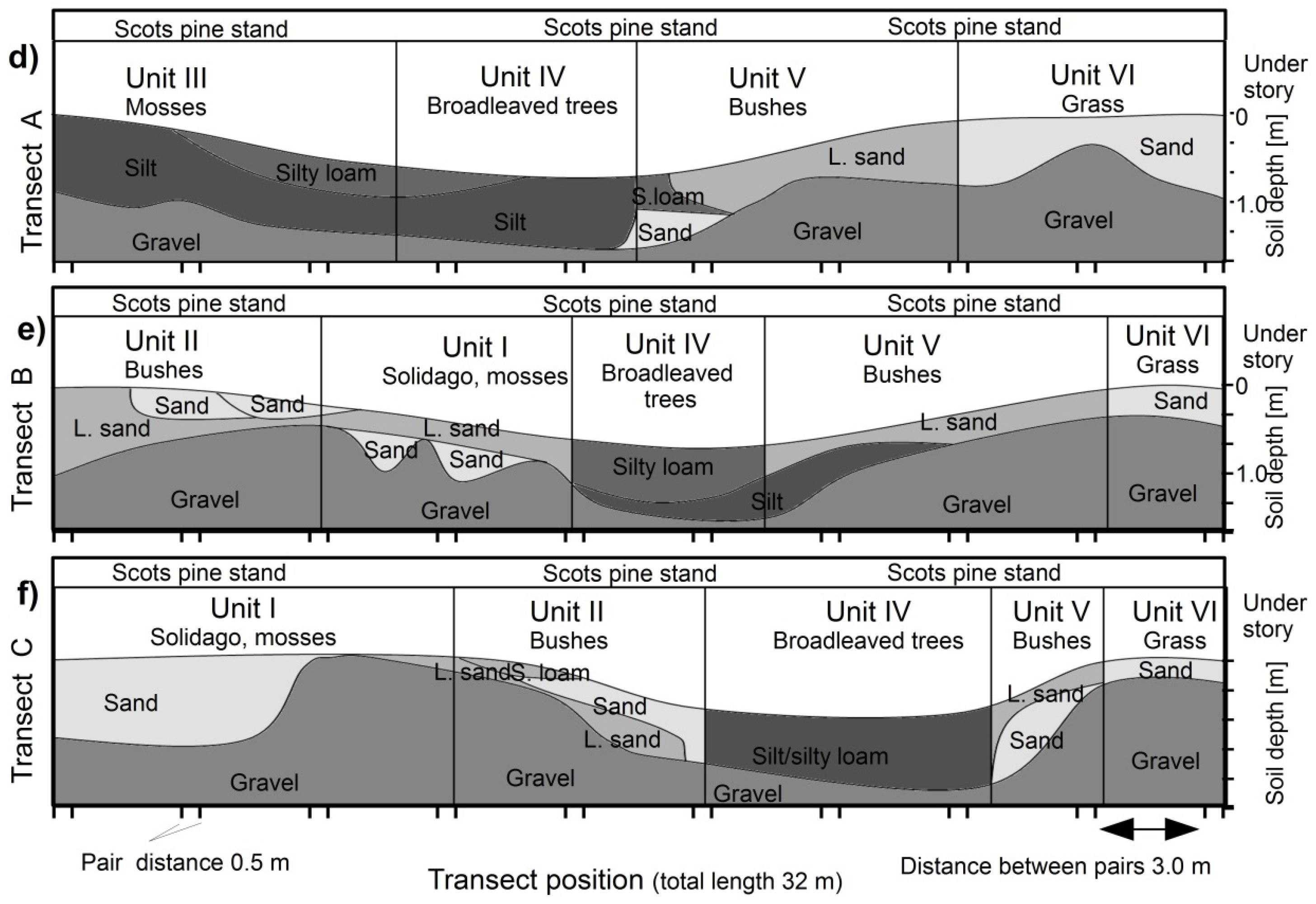
2.1. Experimental Site and Soil Characteristics
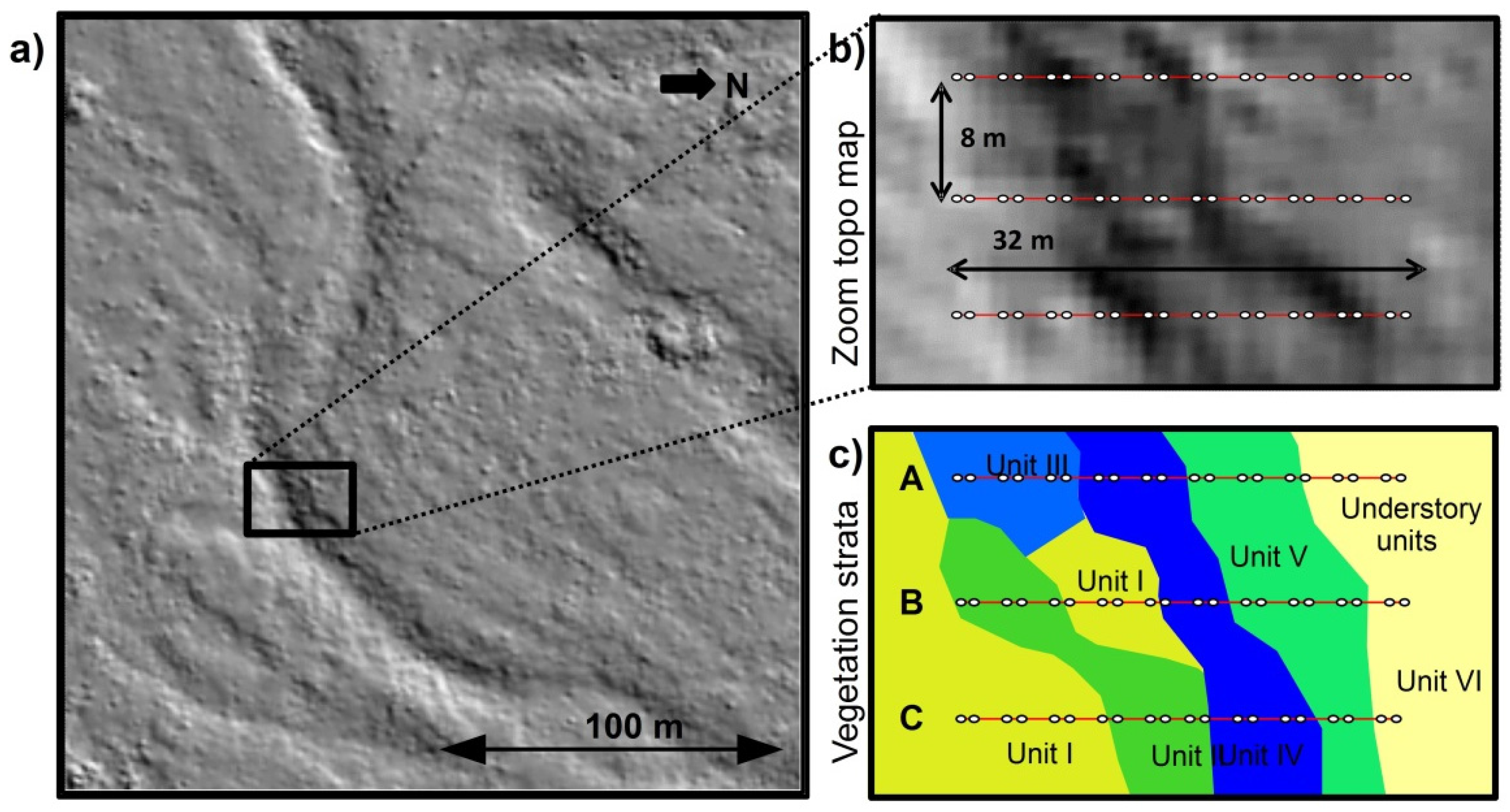
2.2. Experimental Design
2.3. Measurements of Soil Gas Fluxes
2.4. Measurements of Soil-Gas Diffusion Coefficients
2.5. Methanotrophic Activity
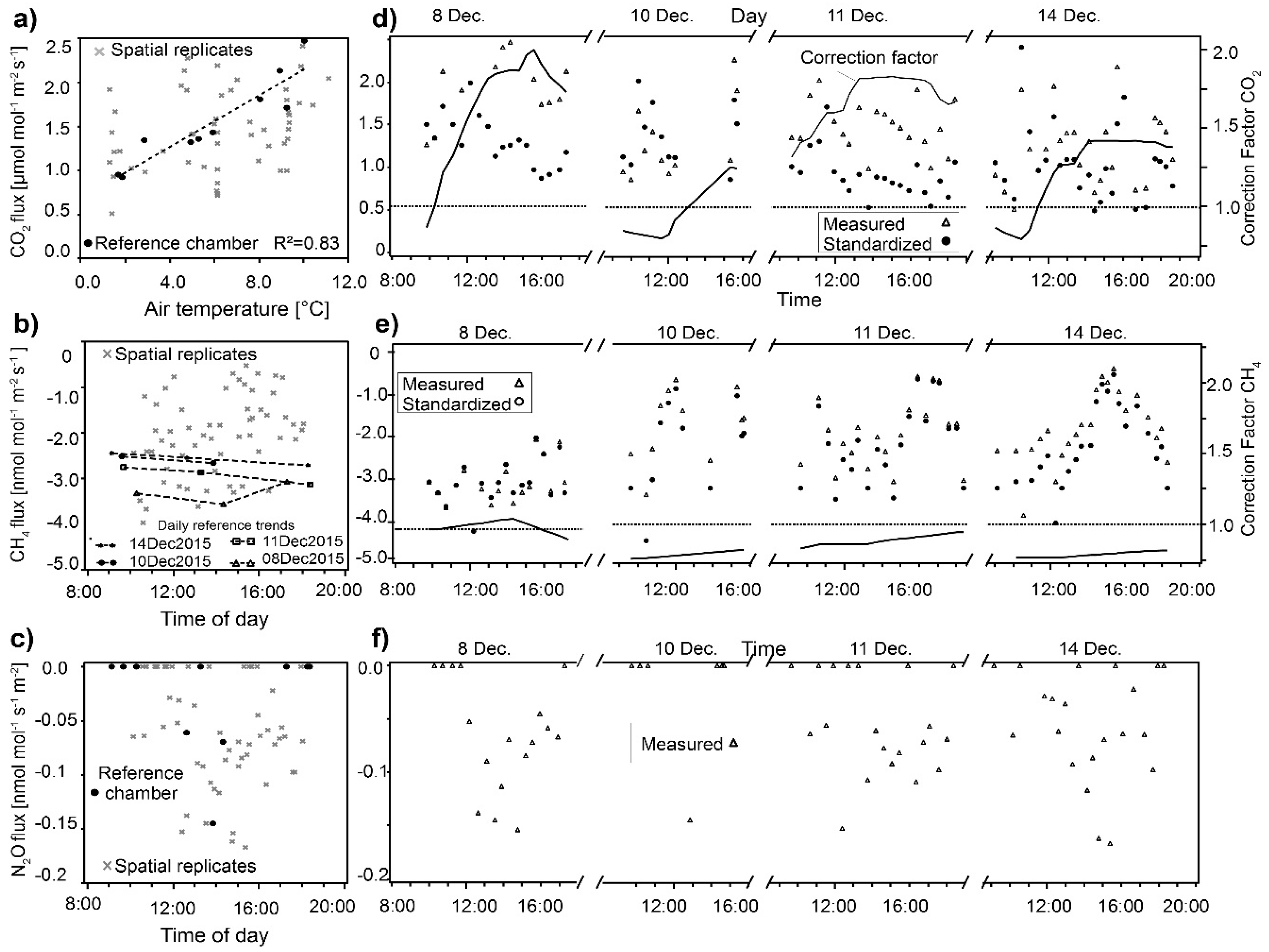
2.6. Standardization of GHG Fluxes
2.7. Statistical Analysis
3. Results and Discussion
3.1. Standardization of Temporal Variability of Greenhouse-Gas Fluxes
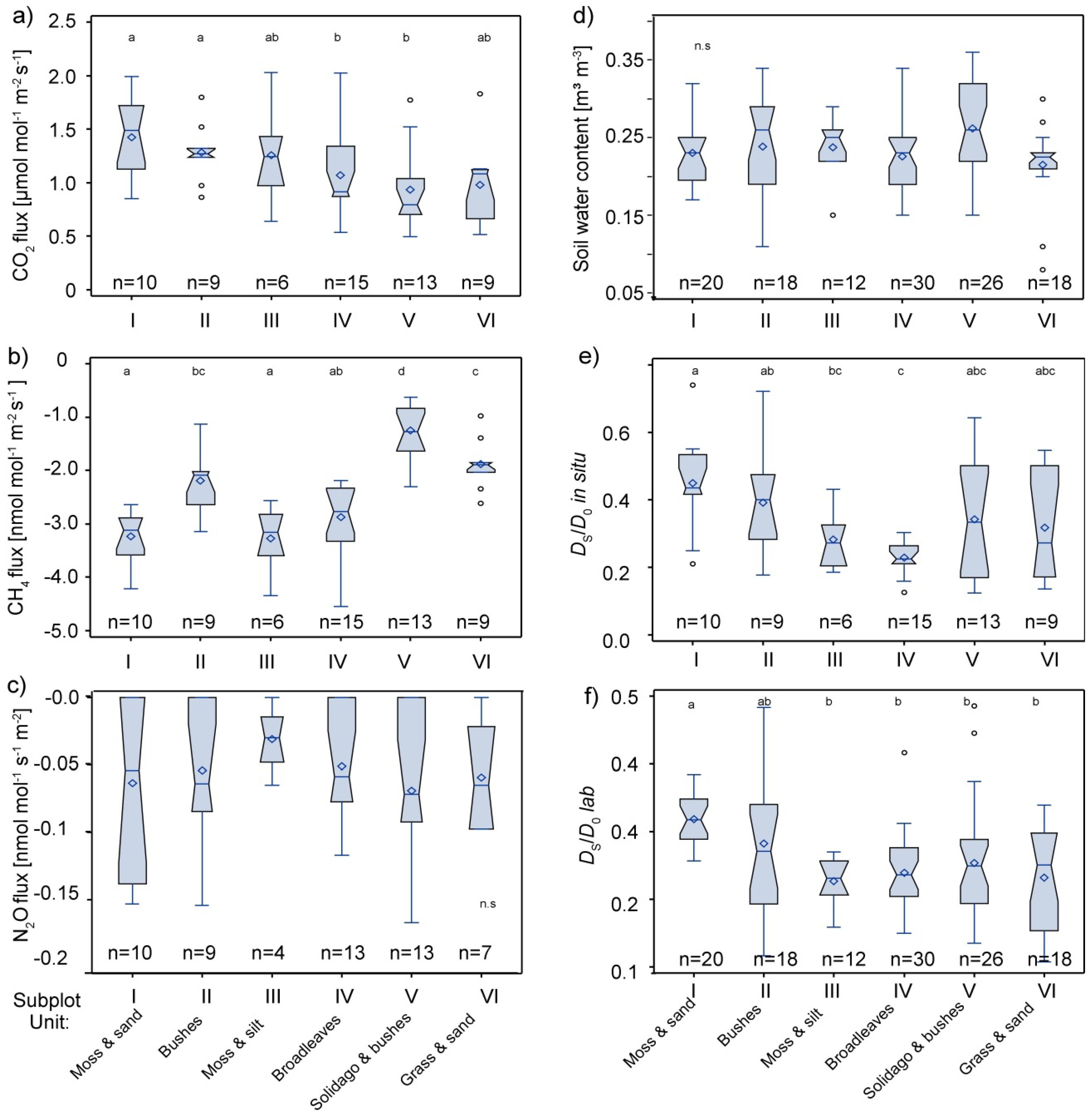
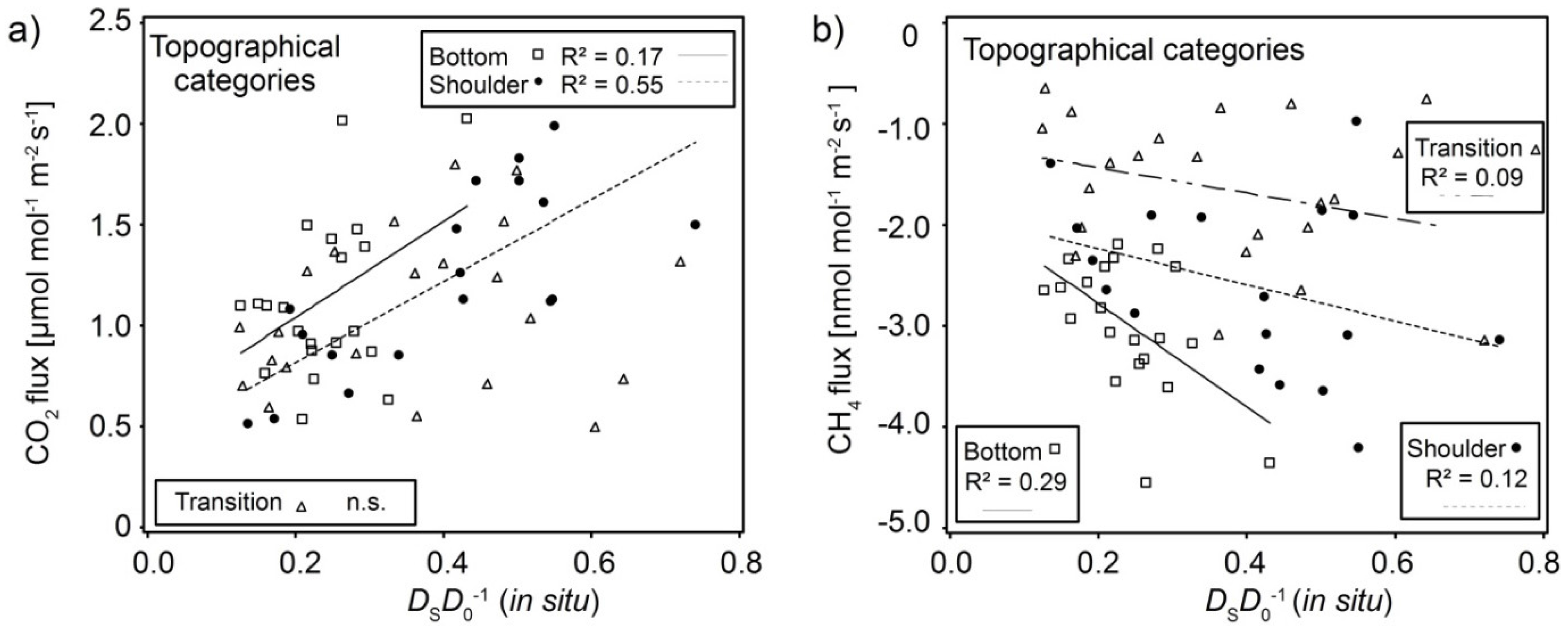
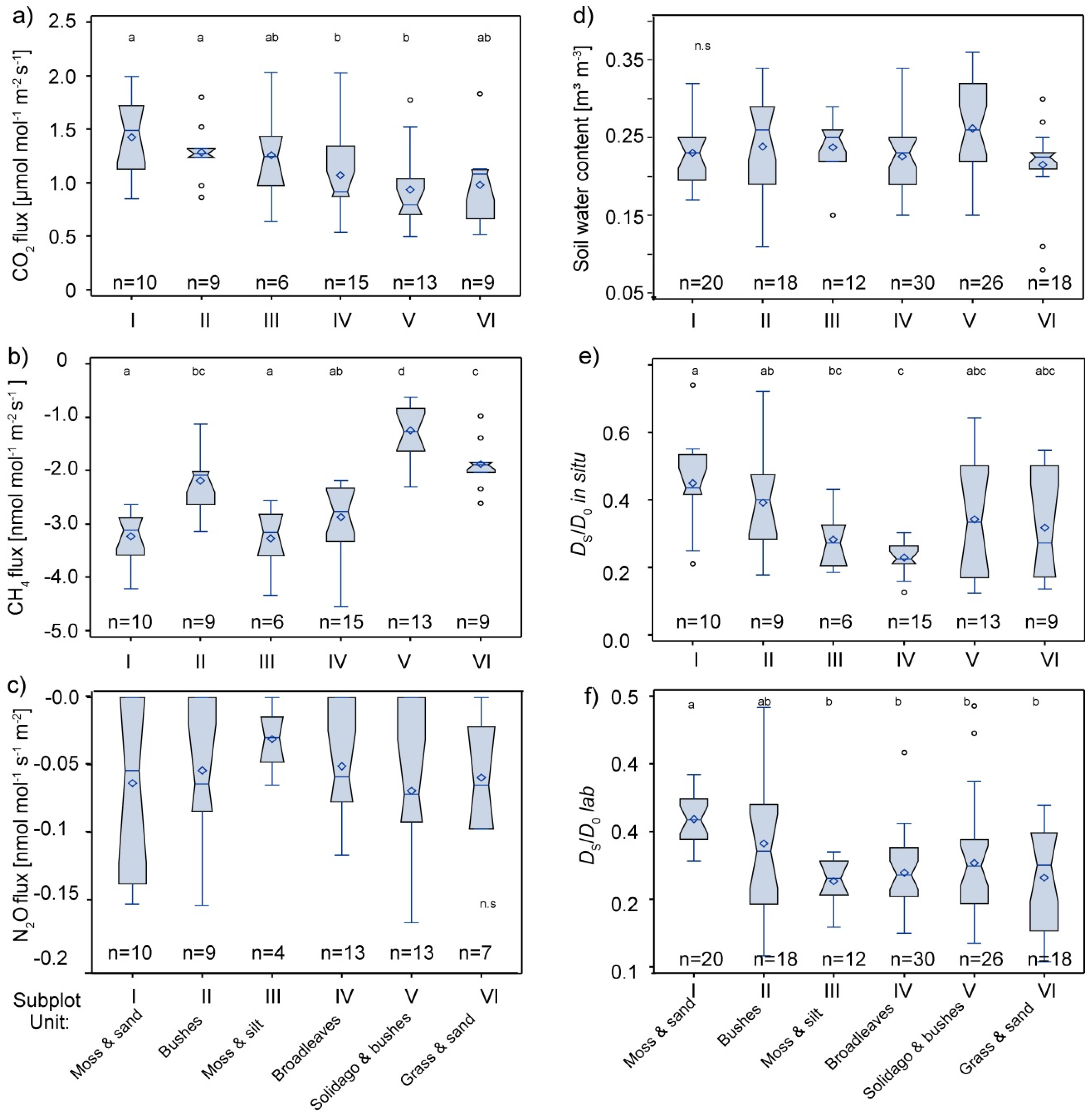
3.2. Spatial Variability of Soil Gas Fluxes
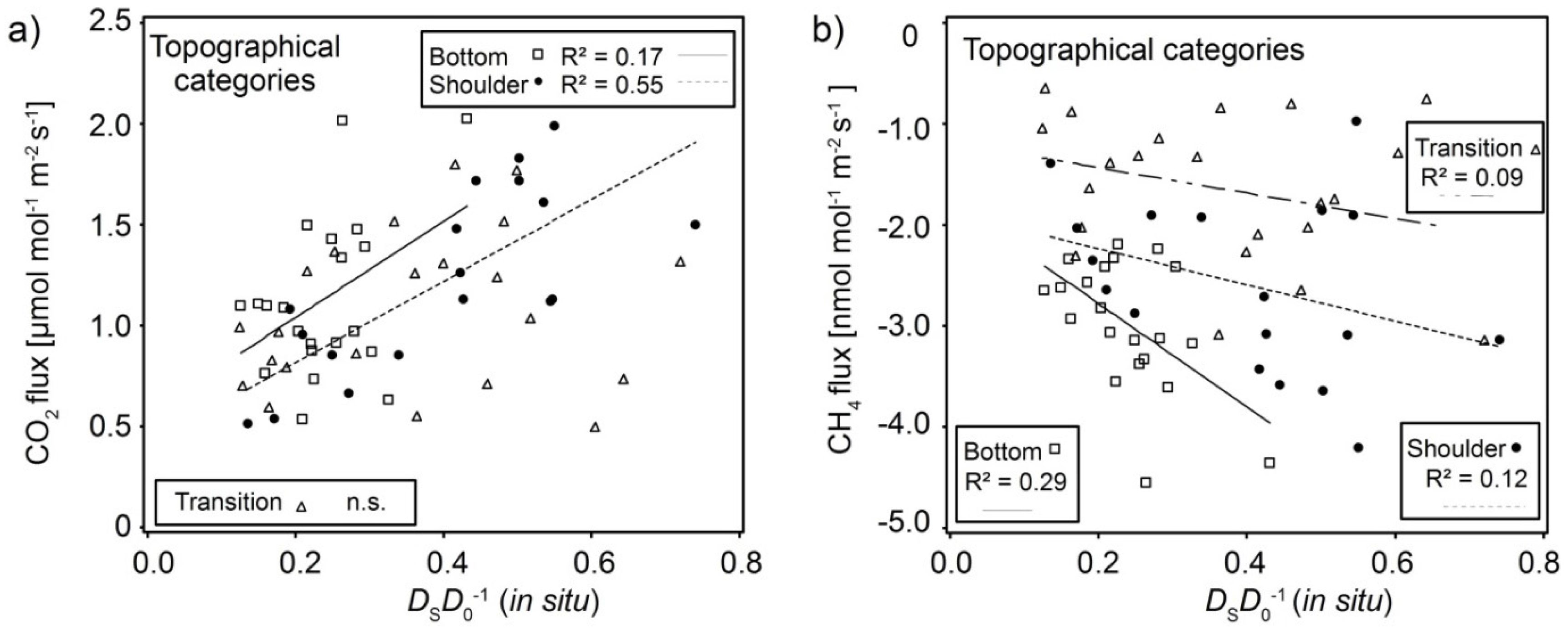
3.3. Interaction of Soil Physical Parameters, Soil Gas Fluxes, and Soil-Vegetation Units
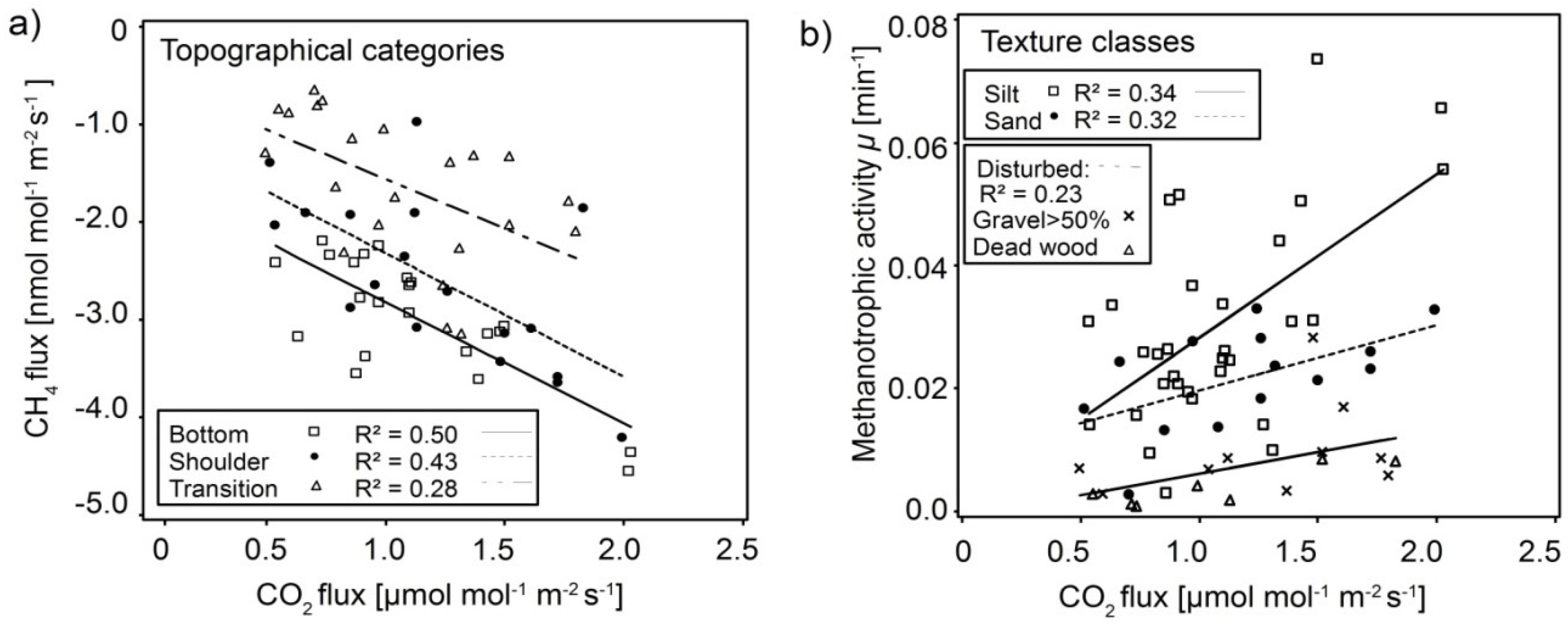
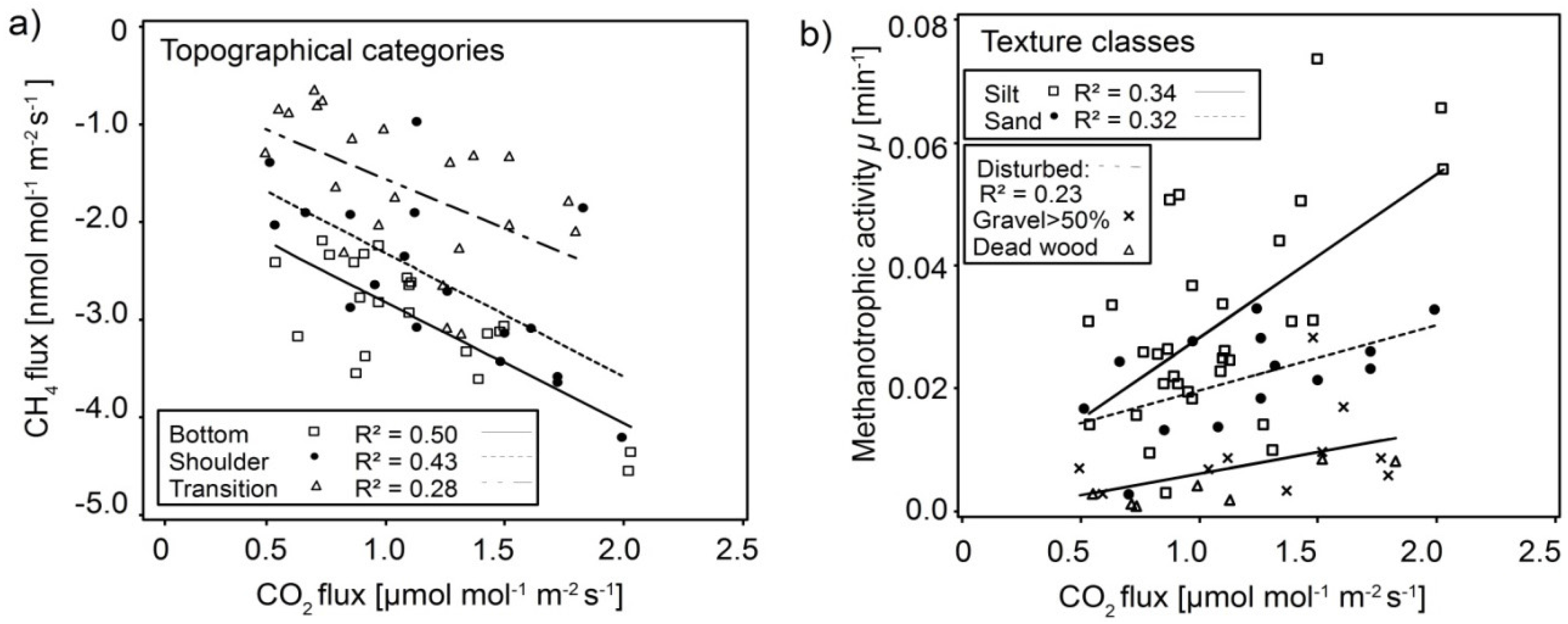
3.4. Interaction of CH4 Fluxes and Methanotrophic Activity with Soil Respiration
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hartmann, D.L.; Klein Tank, A.M.G.; Rusticucci, M.; Alexander, L.V.; Brönnimann, S.; Charabi, Y.; Dentener, F.J.; Dlugokencky, E.J.; Easterling, D.R.; Kaplan, A.; et al. Observations: Atmosphere and Surface. In Climate Change 2013. The Physical Science Basis; Working Group I contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Ed.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014. [Google Scholar]
- Ryan, M.G.; Law, B.E. Interpreting, measuring, and modeling soil respiration. Biogeochemistry 2005, 73, 3–27. [Google Scholar] [CrossRef]
- Forster, P.; Ramaswamy, V.; Artaxo, P.; Berntsen, T.; Betts, R.; Fahey, D.W.; Haywood, J.; Lean, J.; Lowe, D.C.; Myhre, G.; et al. Changes in Atmospheric Constituents and in Radiative Forcing. In Climate Change 2007: The Physical Science Basis; Solomon, S., Ed.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2007. [Google Scholar]
- Conrad, R. Soil Microorganisms as Controllers of Atmospheric Trace Gases (H2, CO, CH4, OCS, N2O, and NO). Microbiol. Rev. 1996, 60, 609–640. [Google Scholar] [PubMed]
- Smith, K.A.; Ball, T.; Conen, F.; Dobbie, K.E.; Massheder, J.; Rey, A. Exchange of greenhouse gases between soil and atmosphere: Interactions of soil physical factors and biological processes. Eur. J. Soil Sci. 2003, 54, 779–791. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Blagodatskaya, E. Microbial hotspots and hot moments in soil: Concept & review. Soil Biol. Biochem. 2015, 83, 184–199. [Google Scholar] [CrossRef]
- Le Mer, J.; Roger, P. Production, oxidation, emission and consumption of methane by soils: A review. Eur. J. Soil Biol. 2001, 37, 25–50. [Google Scholar] [CrossRef]
- Hanson, R.S.; Hanson, T.E. Methanotrophic bacteria. Microbiol. Rev. 1996, 60, 439–471. [Google Scholar] [PubMed]
- Dunfield, P.F. The soil methane sink. In Greenhouse Gas Sinks; Reay, D.S., Hewitt, C.N., Smith, K.A., Grace, J., Eds.; CABI: Wallingford, UK, 2007; pp. 152–170. [Google Scholar]
- Davidson, E.A.; Verchot, L.V. Testing a conceptual model of soil emissions of nitrous and nitric oxides. Glob. Biogeochem. Cycles 2000, 14, 1035–1043. [Google Scholar] [CrossRef]
- Chapuis-Lardy, L.; Wrage, N.; Metay, A.; Chotte, J.-L.; Bernoux, M. Soils, a sink for N2O? A review. Glob. Chang. Biol. 2007, 13, 1–17. [Google Scholar] [CrossRef]
- Davidson, E.A.; Belk, E.; Boone, R.D. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Glob. Chang. Biol. 1998, 4, 217–227. [Google Scholar] [CrossRef]
- Maier, M.; Schack-Kirchner, H.; Hildebrand, E.E.; Schindler, D. Soil CO2 efflux vs. soil respiration: Implications for flux models. Agric. For. Meteorol. 2011, 151, 1723–1730. [Google Scholar] [CrossRef]
- Darenova, E.; Pavelka, M.; Macalkova, L. Spatial heterogeneity of CO2 efflux and optimization of the number of measurement positions. Eur. J. Soil Biol. 2016, 75, 123–134. [Google Scholar] [CrossRef]
- Dalal, R.C.; Allen, D.E.; Livesley, S.J.; Richards, G. Magnitude and biophysical regulators of methane emission and consumption in the Australian agricultural, forest, and submerged landscapes: A review. Plant Soil 2008, 309, 43–76. [Google Scholar] [CrossRef]
- Davidson, E.A.; Ishida, F.Y.; Nepstad, D.C. Effects of an experimental drought on soil emissions of carbon dioxide, methane, nitrous oxide, and nitric oxide in a moist tropical forest. Glob. Chang. Biol. 2004, 10, 718–730. [Google Scholar] [CrossRef]
- Striegl, R.G.; McConnaughey, T.A.; Thorstenson, D.C.; Weeks, E.P.; Woodward, J.C. Consumption of atmospheric methane by desert soils. Nature 1992, 357, 145–147. [Google Scholar] [CrossRef]
- Smith, K.A.; Dobbie, K.E.; Ball, B.; Bakken, L.R.; Sitaula, B.K.; Hansen, S.; Brumme, R.; Borken, W.; Christensen, S.; Priemé, A.; et al. Oxidation of atmospheric methane in Northern European soils, comparison with other ecosystems, and uncertainties in the global terrestrial sink. Glob. Chang. Biol. 2000, 6, 791–803. [Google Scholar] [CrossRef]
- Fest, B.; Hinko-Najera, N.; von Fischer, J.C.; Livesley, S.J.; Arndt, S.K. Soil Methane Uptake Increases under Continuous Throughfall Reduction in a Temperate Evergreen, Broadleaved Eucalypt Forest. Ecosystems 2016, 20, 368–379. [Google Scholar] [CrossRef]
- Bender, M.; Conrad, R. Kinetics of CH4 oxidation in oxic soils exposed to ambient air or high CH4 mixing ratios. FEMS Microbiol. Lett. 1992, 101, 261–270. [Google Scholar] [CrossRef]
- Von Fischer, J.C.; Butters, G.; Duchateau, P.C.; Thelwell, R.J.; Siller, R. In situ measures of methanotroph activity in upland soils: A reaction-diffusion model and field observation of water stress. J. Geophys. Res. 2009, 114. [Google Scholar] [CrossRef]
- Wolf, B.; Chen, W.; Brüggemann, N.; Zheng, X.; Pumpanen, J.; Butterbach-Bahl, K. Applicability of the soil gradient method for estimating soil-atmosphere CO2, CH4, and N2O fluxes for steppe soils in Inner Mongolia. J. Plant Nutr. Soil Sci. 2011, 174, 359–372. [Google Scholar] [CrossRef]
- Flechard, C.; Neftel, A.; Jocher, M.; Amann, C.; Fuhrer, J. Bi-directional soil/atmosphere N2O exchange over two mown grassland systems with contrasting management practices. Glob. Chang. Biol. 2005, 11, 2114–2127. [Google Scholar] [CrossRef]
- Ball, B.; Smith, K.A.; Klemedtsson, L.; Brumme, R.; Sitaula, B.K.; Hansen, S.; Priemé, A.; MacDonald, J.; Horgan, G.W. The influence of soil gas transport properties on methane oxidation in a selection of northern European soils. J. Geophys. Res. 1997, 102, 23309. [Google Scholar] [CrossRef]
- Borken, W.; Beese, F. Methane and nitrous oxide fluxes of soils in pure and mixed stands of European beech and Norway spruce. Eur. J. Soil Sci. 2006, 57, 617–625. [Google Scholar] [CrossRef]
- Menyailo, O.V.; Abraham, W.-R.; Conrad, R. Tree species affect atmospheric CH4 oxidation without altering community composition of soil methanotrophs. Soil Biol. Biochem. 2010, 42, 101–107. [Google Scholar] [CrossRef]
- Nauer, P.A.; Dam, B.; Liesack, W.; Zeyer, J.; Schroth, M.H. Activity and diversity of methane-oxidizing bacteria in glacier forefields on siliceous and calcareous bedrock. Biogeosciences 2012, 9, 2259–2274. [Google Scholar] [CrossRef]
- Warner, D.L.; Villarreal, S.; McWilliams, K.; Inamdar, S.; Vargas, R. Carbon Dioxide and Methane Fluxes From Tree Stems, Coarse Woody Debris, and Soils in an Upland Temperate Forest. Ecosystems 2017. [Google Scholar] [CrossRef]
- WRB. World Reference Base for Soil Resources 2014. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; FAO: Rome, Italy, 2014. [Google Scholar]
- Ad-hoc-Arbeitsgruppe Boden der Staatlichen Geologischen Dienste und der Bundesanstalt für Geowissenschaften und Rohstoffe; Bundesanstalt für Geowissenschaften und Rohstoffe. Bodenkundliche Kartieranleitung; Auflage: 5; E. Schweizerbart’sche Verlagsbuchhandlung: Stuttgart, German, 2005. [Google Scholar]
- Schack-Kirchner, H.; Gaertig, T.; Wilpert, K.V.; Hildebrand, E.E. A modified McIntyre and Phillip approach to measure top-soil gas diffusivity in-situ. J. Plant Nutr. Soil Sci. 2001, 164, 253–258. [Google Scholar] [CrossRef]
- De Klein, C.; Harvey, M. Nitrous Oxide Chamber Methodology Guidelines. Available online: http://globalresearchalliance.org/wp-content/uploads/2015/11/Chamber_Methodology_Guidelines_Final-V1.1-2015.pdf (accessed on 17 November 2016).
- Maier, M.; Schack-Kirchner, H.; Aubinet, M.; Goffin, S.; Longdoz, B.; Parent, F. Turbulence effect on gas transport in three contrasting forest soils. Soil Sci. Soc. Am. J. 2012, 76, 1518–1528. [Google Scholar] [CrossRef]
- Raw, C.; Raw, T.T. Diffusion of gaseous fluoromethanes in air. Chem. Phys. Lett. 1976, 44, 255–256. [Google Scholar] [CrossRef]
- Redeker, K.R.; Baird, A.J.; Teh, Y.A. Quantifying wind and pressure effects on trace gas fluxes across the soil–atmosphere interface. Biogeosciences 2015, 12, 7423–7434. [Google Scholar] [CrossRef]
- Laemmel, T.; Mohr, M.; Schack-Kirchner, H.; Schindler, D.; Maier, M. Direct observation of wind-induced pressure-pumping on gas transport in soils. Soil Sci. Soc. Am. J. 2017. [Google Scholar] [CrossRef]
- Maier, M.; Schack-Kirchner, H.; Hildebrand, E.E.; Holst, J. Pore-space CO2 dynamics in a deep, well-aerated soil. Eur. J. Soil Sci. 2010, 61, 877–887. [Google Scholar] [CrossRef]
- Iversen, B.V.; Schjønning, P.; Poulsen, T.G.; Moldrup, P. In situ, on-site and laboratory measurements of soil air permeability. Soil Sci. 2001, 166, 97–106. [Google Scholar] [CrossRef]
- Gulledge, J.; Schimel, J.P. Moisture control over atmospheric CH4 consumption and CO2 production in diverse Alaskan soils. Soil Biol. Biochem. 1998, 30, 1127–1132. [Google Scholar] [CrossRef]
- Nauer, P.A.; Schroth, M.H. In Situ Quantification of Atmospheric Methane Oxidation in Near-Surface Soils. Vadose Zone J. 2010, 9, 1052. [Google Scholar] [CrossRef]
- Fuller, E.N.; Schettler, P.D.; Giddings, J.C. New method for prediction of binarz gas/phase diffusion coefficients. Ind. Eng. Chem. 1966, 58, 18–27. [Google Scholar] [CrossRef]
- Hastie, T.; Tibshirani, R.; Friedman, J.H. The Elements of Statistical Learning. Data Mining, Inference, and Prediction, 2nd ed.; Springer: New York, NY, USA, 2016. [Google Scholar]
- Cutler, D.R.; Edwards, T.C.; Beard, K.H.; Cutler, A.; Hess, K.T.; Gibson, J.; Lawler, J.J. Random Forests for classification in ecology. Ecology 2007, 88, 2783–2792. [Google Scholar] [CrossRef] [PubMed]
- Were, K.; Bui, D.T.; Dick, Ø.B.; Singh, B.R. A comparative assessment of support vector regression, artificial neural networks, and random forests for predicting and mapping soil organic carbon stocks across an Afromontane landscape. Ecol. Indic. 2015, 52, 394–403. [Google Scholar] [CrossRef]
- Liaw, A.; Wiener, M. Classification and Regression by randomForest. R News 2002, 2, 18–22. [Google Scholar]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Glatzel, S.; Stahr, K. Methane and nitrous oxide exchange in differently fertilised grassland in southern Germany. Plant Soil 2001, 231, 21–35. [Google Scholar] [CrossRef]
- Ryden, J.C. N2O exchange between a grassland soil and the atmosphere. Nature 1981, 292, 235–237. [Google Scholar] [CrossRef]
- Shvaleva, A.; Siljanen, H.M.P.; Correia, A.; Costa E Silva, F.; Lamprecht, R.E.; Lobo-do-Vale, R.; Bicho, C.; Fangueiro, D.; Anderson, M.; Pereira, J.S.; et al. Environmental and microbial factors influencing methane and nitrous oxide fluxes in Mediterranean cork oak woodlands: Trees make a difference. Front. Microbiol. 2015, 6, 1104. [Google Scholar] [CrossRef] [PubMed]
- Schrier-Uijl, A.P.; Kroon, P.S.; Leffelaar, P.A.; van Huissteden, J.C.; Berendse, F.; Veenendaal, E.M. Methane emissions in two drained peat agro-ecosystems with high and low agricultural intensity. Plant Soil 2010, 329, 509–520. [Google Scholar] [CrossRef]
- Konda, R.; Ohta, S.; Ishizuka, S.; Arai, S.; Ansori, S.; Tanaka, N.; Hardjon, O.A. Spatial structures of N2O, CO2, and CH4 fluxes from Acacia mangium plantation soils during a relatively dry season in Indonesia. Soil Biol. Biochem. 2008, 40, 3021–3030. [Google Scholar] [CrossRef]
- West, A.E.; Brooks, P.D.; Fisk, M.C.; Smith, L.K.; Holland, E.A.; Jaeger, I.C.; Babcock, S.; Lai, R.S.; Schmidt, S.K. Landscape patterns of CH4 fluxes in an alpine tundra ecosystem. Biogeochemistry 1999, 45, 243–264. [Google Scholar] [CrossRef]
- Reay, D.; NedwellL, D.; McNamara, N.; Ineson, P. Effect of tree species on methane and ammonium oxidation capacity in forest soils. Soil Biol. Biochem. 2005, 37, 719–730. [Google Scholar] [CrossRef]
- Hiltbrunner, D.; Zimmermann, S.; Karbin, S.; Hagedorn, F.; Niklaus, P.A. Increasing soil methane sink along a 120-year afforestation chronosequence is driven by soil moisture. Glob. Chang. Biol. 2012, 18, 3664–3671. [Google Scholar] [CrossRef]
- Niklaus, P.A.; Le Roux, X.; Poly, F.; Buchmann, N.; Scherer-Lorenzen, M.; Weigelt, A.; Barnard, R.L. Plant species diversity affects soil-atmosphere fluxes of methane and nitrous oxide. Oecologia 2016, 181, 919–930. [Google Scholar] [CrossRef] [PubMed]
- Raghoebarsing, A.A.; Smolders, A.J.P.; Schmid, M.C.; Rijpstra, W.I.C.; Wolters-Arts, M.; Derksen, J.; Jetten, M.S.M.; Schouten, S.; Sinninghe Damste, J.S.; Lamers, L.P.M.; et al. Methanotrophic symbionts provide carbon for photosynthesis in peat bogs. Nature 2005, 436, 1153–1156. [Google Scholar] [CrossRef] [PubMed]
- Putkinen, A.; Larmola, T.; Tuomivirta, T.; Siljanen, H.M.P.; Bodrossy, L.; Tuittila, E.-S.; Fritze, H. Peatland succession induces a shift in the community composition of Sphagnum-associated active methanotrophs. FEMS Microbiol. Ecol. 2014, 88, 596–611. [Google Scholar] [CrossRef] [PubMed]
- Brumme, R.; Borken, W. Site variation in methane oxidation as affected by atmospheric deposition and type of temperate forest ecosystem. Glob. Biogeochem. Cycles 1999, 13, 493–501. [Google Scholar] [CrossRef]
- Guckland, A.; Flessa, H.; Prenzel, J. Controls of temporal and spatial variability of methane uptake in soils of a temperate deciduous forest with different abundance of European beech (Fagus sylvatica L.). Soil Biol. Biochem. 2009, 41, 1659–1667. [Google Scholar] [CrossRef]
- Dubey, S.K.; Sinha, A.S.; Singh, J.S. Spatial variation in the capacity of soil for CH4 uptake and population size of methane oxidizing bacteria in dryland rice agriculture. Curr. Sci. 2000, 78, 617–620. [Google Scholar]
- Roslev, P.; Iversen, N.; Henriksen, K. Oxidation and assimilation of atmospheric methane by soil methane oxidizers. Appl. Environ. Microbiol. 1997, 63, 874–880. [Google Scholar] [PubMed]
- Ebrahimi, A.; Or, D. Hydration and diffusion processes shape microbial community organization and function in model soil aggregates. Water Resour. Res. 2015, 51, 9804–9827. [Google Scholar] [CrossRef]
- Butterfield, C.N.; Li, Z.; Andeer, P.F.; Spaulding, S.; Thomas, B.C.; Singh, A.; Hettich, R.L.; Suttle, K.B.; Probst, A.J.; Tringe, S.G.; et al. Proteogenomic analyses indicate bacterial methylotrophy and archaeal heterotrophy are prevalent below the grass root zone. PeerJ 2016, 4, e2687. [Google Scholar] [CrossRef] [PubMed]
- Stiehl-Braun, P.A.; Powlson, D.S.; Poulton, P.R.; Niklaus, P.A. Effects of N fertilizers and liming on the micro-scale distribution of soil methane assimilation in the long-term Park Grass experiment at Rothamsted. Soil Biol. Biochem. 2011, 43, 1034–1041. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Unit | Topography | Understory & Ground Vegetation | Soil Texture |
|---|---|---|---|
| Unit I | Southern shoulder & plain | Solidago gigantea (L.) and moss species | Sand & gravel to loamy sand |
| Unit II | Southern slope | Bushes, Ligustrum vulgare (L.); Crataegus monogyna (Jacq.) | Sandy gravel to sandy loam |
| Unit III | Bottom-slope | Mosses (e.g., Scleropodium purum, L.) | Silt and silty loam. |
| Unit IV | Bottom | 10–20 year old broadleaves Juglans regia (L.), Tilia platyphyllos (L.), Carpinus betulus (L.), less groundcover | Silt and silty loam. |
| Unit V | Norther slope & shoulder | Bushes, Ligustrum vulgare (L.); Crataegus monogyna (Jacq.) | Sandy gravel to sandy loam |
| Unit VI | Northern plain | (few) Solidago gigantea (L.), grass species | Sand and gravel |
| Flux | All Measurements | Weighted Sum of Strata | ||
|---|---|---|---|---|
| Mean | CV | Mean | CV | |
| CO2 (µmol s−1 m−2) | 1.13 | 36% | 1.13 | 33% |
| CH4 (nmol s−1 m−2) | −2.39 | 38% | −2.39 | 24% |
| N2O (nmol s−1 m−2) | −0.05 | 88% | −0.05 | 87% |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maier, M.; Paulus, S.; Nicolai, C.; Stutz, K.P.; Nauer, P.A. Drivers of Plot-Scale Variability of CH4 Consumption in a Well-Aerated Pine Forest Soil. Forests 2017, 8, 193. https://doi.org/10.3390/f8060193
Maier M, Paulus S, Nicolai C, Stutz KP, Nauer PA. Drivers of Plot-Scale Variability of CH4 Consumption in a Well-Aerated Pine Forest Soil. Forests. 2017; 8(6):193. https://doi.org/10.3390/f8060193
Chicago/Turabian StyleMaier, Martin, Sinikka Paulus, Clara Nicolai, Kenton P. Stutz, and Philipp A. Nauer. 2017. "Drivers of Plot-Scale Variability of CH4 Consumption in a Well-Aerated Pine Forest Soil" Forests 8, no. 6: 193. https://doi.org/10.3390/f8060193
APA StyleMaier, M., Paulus, S., Nicolai, C., Stutz, K. P., & Nauer, P. A. (2017). Drivers of Plot-Scale Variability of CH4 Consumption in a Well-Aerated Pine Forest Soil. Forests, 8(6), 193. https://doi.org/10.3390/f8060193