Abstract
Drought is a major constraint of forest productivity and tree growth across diverse habitat types. In this study, we investigated the drought responses of four conifer species growing within two locations of differing elevation and climatic conditions in northern Mexico. Two species were selected at a mesic site (Cupressus lusitanica Mill., Abies durangensis Martínez) and the other two species were sampled at a xeric site (Pinus engelmannii Carr., Pinus cembroides Zucc.). Using a dendrochronological approach, we correlated the radial-growth series of each species and the climatic variables. All study species positively responded to wet-cool conditions during winter and spring. Despite the close proximity of species at a mesic site, A. durangensis had high responsiveness to hydroclimatic variability, but C. lusitanica was not responsive. At the xeric site, P. engelmannii and P. cembroides were very responsive to drought severity, differentiated only by the longer time scale of the response to accumulated drought of P. engelmannii. The responsiveness to hydroclimate and drought of these tree species seems to be modulated by site conditions, or by the functional features of each species that are still little explored. These findings indicate that differentiating between mesic and xeric habitats is a too coarse approach in diverse forests with a high topographic heterogeneity.
1. Introduction
The hydroclimatic variability experienced by forests has shown differential responsiveness in recent decades as a consequence of global climate change [1]. Among the effects of climate warming, there has been an increase in water deficit, both in intensity and duration [2]. Water deficit has been reported as one of the major triggering factors of forest growth decline, dieback, and tree mortality [3,4,5]. Droughts reduce radial growth and alter the hydraulic conductivity of trees [6,7,8], with a negative impact on carbon sequestration [9,10] and forest productivity [11,12]. Moreover, given that aridity is expected to intensify in the future in many forested areas, this may have negative and long-term implications on forest productivity [7,13].
These effects of drought stress on growth and productivity may differ between forest type and tree species [7]. Some studies have argued that the radial growth of a tree species growing under xeric habitats is more affected by drought than those from mesic habitats [14]. Conversely, other studies have argued that species from xeric habitats are better adapted to withstand drought stress than species from mesic habitats [15]. This last idea implies that species in areas subjected to marked or prolonged seasonal water deficit (xeric habitats) may be less vulnerable to drought stress in terms of radial growth than species of wetter habitats (mesic habitats). Therefore, it is important to assess how different tree species from mesic and xeric habitats respond to drought severity. It is well known that site features including climate or elevation often influence forest productivity, composition, and structure [16]. Therefore, comparing tree growth responses to drought in xeric and mesic habitats could allow for a better understanding of the influence of hydroclimate variability on forest productivity in the face of expected drier conditions over the coming years which could severely affect entire regions.
To investigate the factors responsible for the temporal and spatial response patterns of forests to drought, elevation or precipitation gradients have been used as a surrogate of climate gradients [16,17]. Along with these gradients, radial growth (usually measured as changes in tree-ring width) varies in response to changes in water deficiency [5,18], site conditions [19], and tree characteristics (e.g., tree size, age, dominance, or vigor) [20,21]. Stand density [22], neighborhood composition [23], and past management history including thinning regimes [24] may also affect trees’ responses to drought.
Since northern Mexico has been reported to be susceptible to forest dieback and growth decline due to the increase in temperature and the occurrence of more severe droughts [25], this area is of great interest in understanding how tree species respond to drought [26]. This region has one of the highest conifer species richness in the world [27], where they grow in climatically and topographically contrasting environments. The main conifers include a diverse range of species: from Pinus cembroides Zucc., a species representative of low elevation and dry locations, to Cupressus lusitanica Mill., a species dominant in humid sites [27]. Here we compare a xeric habitat where trees grow under marginal environmental conditions (e.g., low precipitation and high evapotranspiration rates) with a mesic habitat characterized by higher productivity and more favorable climate conditions such as higher rainfall and lower evapotranspiration rates [27,28].
Often, tree species are replaced along elevation gradients and this shift in forest composition could be reflected in changes of growth and responses to drought. Although previous studies have been conducted in these Mexican mountains to associate climate variability with the radial growth of certain tree species [28,29,30,31], to date, there has been no analysis containing a multi-species perspective in locations of differing water availability. Several studies agree on the need to improve knowledge of the long-term drought impact at contrasting sites and forest types [11,32]. This improvement may provide insights into the mechanisms involved in growth response to hydroclimate and in understanding the growth patterns that these species could present under warmer and drier conditions. In this paper, we aim to study the drought growth responses of four conifer species growing in mesic and xeric habitats of differing elevation located in northern Mexico. We hypothesize that species from the mesic habitat will show the lowest responsiveness to drought.
2. Materials and Methods
2.1. Study Area and Sampled Tree Species
This study was conducted in the Sierra Madre Occidental, located to the north of Durango, Mexico (Figure 1a). The Sierra Madre Occidental is a mountain range formed by a volcanic plateau with an average elevation of 2000 m, but some areas rise to 3000 m. This range extends northwest from the south of the Tropic of Cancer to the west of Durango, ending in southern Arizona. The landscape is characterized by rolling highlands and deep canyons that rise at the edge of the plateaus and extend to the mountain peaks [33]. This area has different types of soil, such as Eutric Regosol and Dystric Cambisol soils. The former is characterized as poorly developed soil, with low organic matter content, and is often shallow; while the latter are young, acidic, and nitrogen-rich soils. Coniferous forests of the region are relatively rich floristically [27].
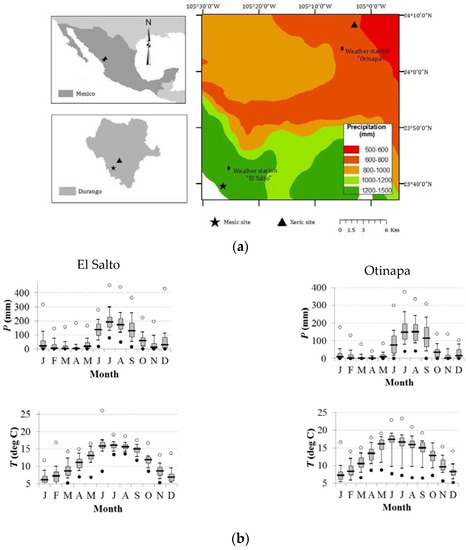
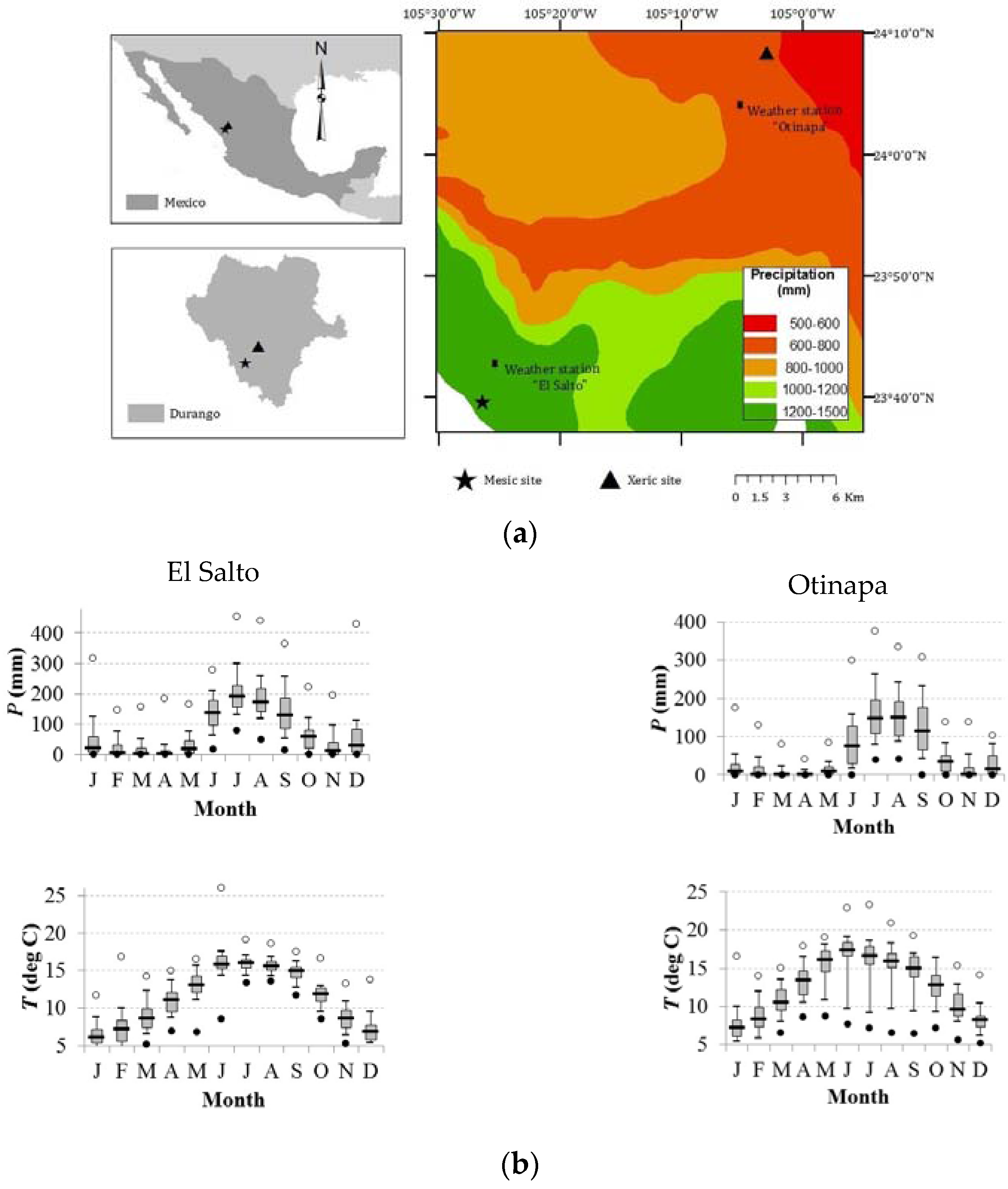
Figure 1.
(a) Site locations in northern México and the Durango state; and (b) climate diagrams of the El Salto and Otinapa stations corresponding to the mesic and xeric habitats, respectively. The map shows the study area, where trees of A. durangensis, C. lusitanica, from the mesic site (star), and P. engelmannii and P. cembroides from the xeric site (triangle) were sampled. The diagrams show the distribution (box plots) of monthly precipitation (P) and temperature (T) data. Months are abbreviated by J = January, F = February, …, D = December.
This region is one of the most important timber reserves in Mexico, supplying more than 25% of the national timber, and it is therefore relevant for sustainable forestry in the country [31]. In addition, these forests are major water sources used to supply agriculture in Sinaloa, Nayarit, and Coahuila [34]. The Sierra Madre Occidental is also an important and valuable source of ecosystem services for local populations [17], and it is a strategically important region for studies in the conservation and biodiversity of conifer forest ecosystems.
In this study, we selected four species of conifers representative of the study area inhabiting wet and dry sites [27]. Their representativeness in the study area and their importance for conservation and management purposes made them suitable for the objectives of this study. These species were: Abies durangensis Martínez, Cupressus lusitanica Mill., Pinus engelmannii Carr., and Pinus cembroides Zucc. The first two species are found in a humid and protected area (hereafter referred to as mesic habitat) located at 2700 m, while the latter two correspond to an irregular forest situated in a dry region (hereinafter xeric habitat) at 2200 m and managed with the selection system [28]. The mesic habitat has been considered a protected area for 30 years in order to produce environmental benefits and preserve biodiversity, whilst stumps appear in the xeric habitat, thereby confirming anthropogenic impact.
The climatic stations of El Salto (period 1946–2015, xeric habitat) and Otinapa (period 1962–2015, mesic habitat) illustrate the climate differences in the study area (Figure 1b). In El Salto, the average annual temperature is 11.5 °C and annual precipitation is 1100 mm, whereas in Otinapa the average annual temperature is 12.6 °C and annual precipitation is 600 mm (Figure 1b).
2.2. Collection of Samples and Development of Growth Chronologies
We selected and sampled 15 dominant individuals of each species, in which at least two radial wood cores were sampled at a height of 1.3 m by using a Pressler increment borer. In addition, tree diameter was measured at 1.3 m (Table 1).

Table 1.
Dendrochronological statistics of the sampled sites and tree species considering the common interval 1955–2015 (except for the best replicated period).
Wood samples were air dried, glued in wooden mounts and polished using sandpaper. Next, we detected and synchronized the series of characteristic rings (narrow and wide rings) among cores of the same tree and among tree series of each species. The tree-ring width was measured at 0.01 mm resolution using the LINTAB system (Rinntech, Heidelberg, Germany). Cross-dating was checked using the COFECHA program, which compares the series of each tree with a master chronology by species by calculating correlations [36].
To generate the chronologies of each species, long-term growth trends not associated with climatic variables were removed using an exponential negative function and obtaining standardized ring-width indexes. Autoregressive models were adjusted to remove most of the first-order temporal autocorrelation and were averaged using biweight robust means to obtain the mean residual series for each species [37]. The chronologies were developed using the library dplR [38] of the R Statistical package [39].
2.3. Relationships between Climate, Drought, and Growth
The influence of the local climate and drought severity on radial growth was evaluated by calculating the Pearson correlation index (r) among the residual growth series of each species, and the following climatic variables: evaporation (Ev), maximum temperature (Tmax) and minimum temperature (Tmin), and precipitation (Pr). These instrumental climatic data were obtained monthly from the two nearest climatic seasons: El Salto (period 1946–2015) and Otinapa (period 1962–2015) [40]. The correlation analysis window was from June of the previous year to September of the growing year, based on previous dendroclimatic analyses in the study area [17,25]. To estimate drought severity and duration we used the standardized precipitation evapotranspiration index (SPEI) [41], as this index considers the effects of temperature on evapotranspiration rates on water availability and it quantifies the intensity of drought at different time scales. The SPEI was calculated from local instrumental climatological data using the SPEI library of R [42]. Given that each tree species can show different temporal responses to accumulated water deficit [31], the SPEI was calculated at time scales from 1–28 months and correlations were calculated between these values and the growth series (residual indices) of each tree species.
Finally, to characterize the relationship between growth and climate and drought on a regional scale, we calculated spatial Pearson correlations across Mexico and the southern USA by relating the residual indices to mean maximum temperature and SPEI data. These variables were selected based on previous analyses that aimed to characterize the growth responses to climate or drought across large scales which reflect atmospheric patterns driving regional and continental droughts [43]. In both cases, we used the climate database Climate Research Unit (CRU) version 3.22 which considered 0.5° gridded data of temperature and SPEI [44]. These correlations were calculated using the Climate Explorer website [45].
3. Results
3.1. Growth Patterns
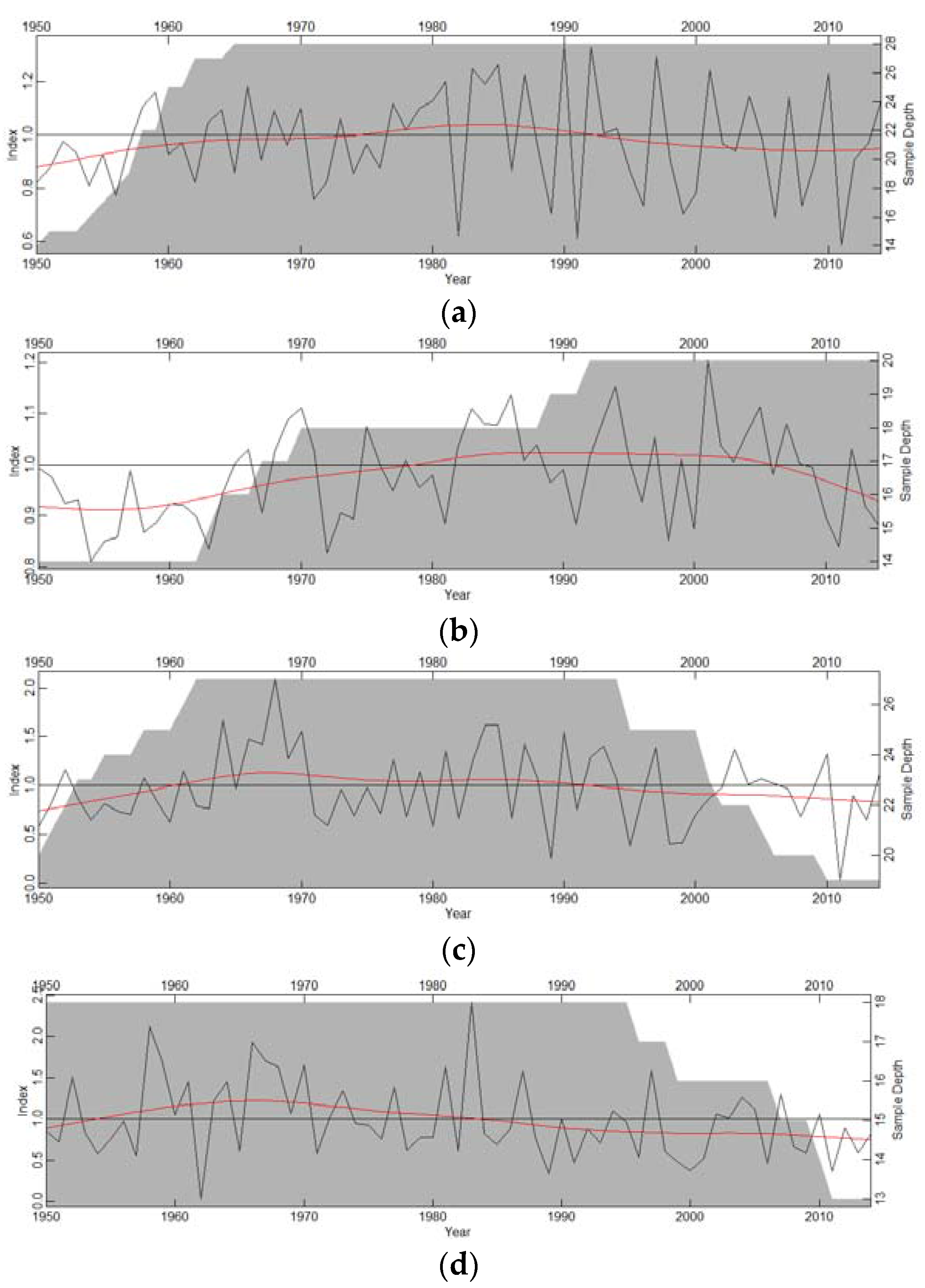
Tree-ring widths ranged from 1.04 mm (Pinus cembroides) to 2.17 mm (Pinus engelmannii). For the first-order autocorrelation (AC), the highest values were obtained by species from the mesic habitat (C. lusitanica, 0.70; A. durangensis, 0.72), while species from the xeric habitat had lower values (P. engelmannii, 0.60; P. cembroides, 0.61). The highest correlation between trees was observed in the xeric habitat (P. engelmannii), while the smaller values corresponded to C. lusitanica from the mesic habitat (Table 1). Growth patterns were synchronized for most species, except for C. lusitanica (Figure 2).
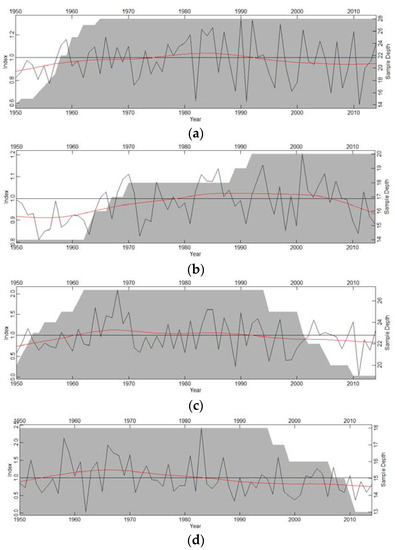
Figure 2.
Residual chronologies of ring-width indices (black lines) of (a) A. durangensis; (b) C. lusitanica; (c) P. engelmannii; and (d) P. cembroides. The red lines show moving averages and the grey areas indicate the number of measured radii (sample depth) for each year (right y-axis).
3.2. Climate-Growth Correlations
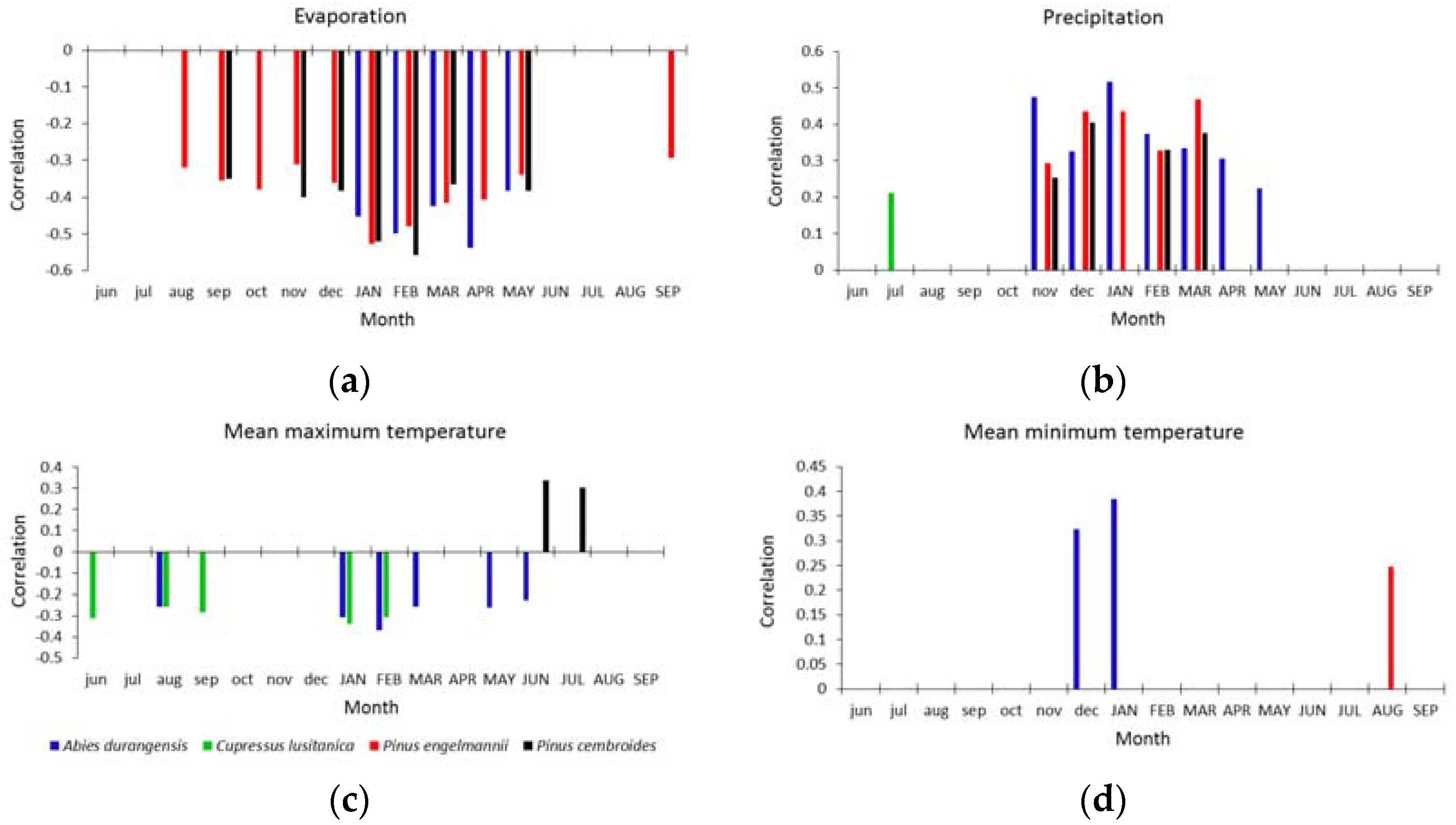
Abies durangensis was the only species that had significant correlations with all climatic variables (Figure 3). The growth of this species was negatively correlated (r = −0.42–0.52) with evaporation (Ev) from January–May of the growth year, and with Tmax (r = −0.25–0.35) from August of the previous year, and from January–March and May–June of the current year. The precipitation (Pr) during the previous winter (November–February) and spring (March–May) was positively associated with growth (r = 0.22–0.52), as was the Tmin during the previous winter (December–January, r = 0.34–0.40).
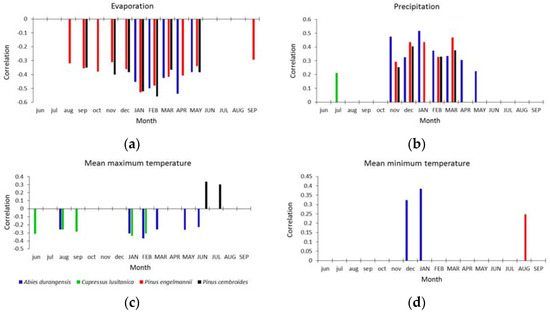
Figure 3.
Climate–growth associations (Pearson correlations) for the four species studied with regard to the following climatic variables: (a) evaporation; (b) precipitation; (c) maximum temperature; and (d) minimum temperature. Only the significant correlations (p < 0.05) are shown. The previous- and current-year months are symbolized by lowercase and uppercase letters, respectively.
In the case of Cupressus lusitanica, this species only had a significant and negative correlation with Tmax (r = −0.25–−0.23) in June, August, and September of the previous year and from January–February of the current year. Positive correlations were observed with Pr (r = 0.20) in July of the previous year (Figure 3).
In the xeric habitat, Pinus engelmannii showed negative correlations with the Ev from August of the previous year to September of the current year (r = −0.31–−0.50), with particularly strong relationships from January–March. We obtained positive and significant correlations (r = 0.30–0.47) for Pr in this same period, and also with Tmin of August of the current year (r = 0.25).
For Pinus cembroides, the variables that positively influenced their growth were Pr (r = 0.25–0.40) from November–March and Tmax (r = 0.30–0.35) during June and July of the growth year. A negative correlation was observed in this species with Ev (r = −0.35–−0.55) for the months of September from the previous year, and November–February and May of the current year (Figure 3).
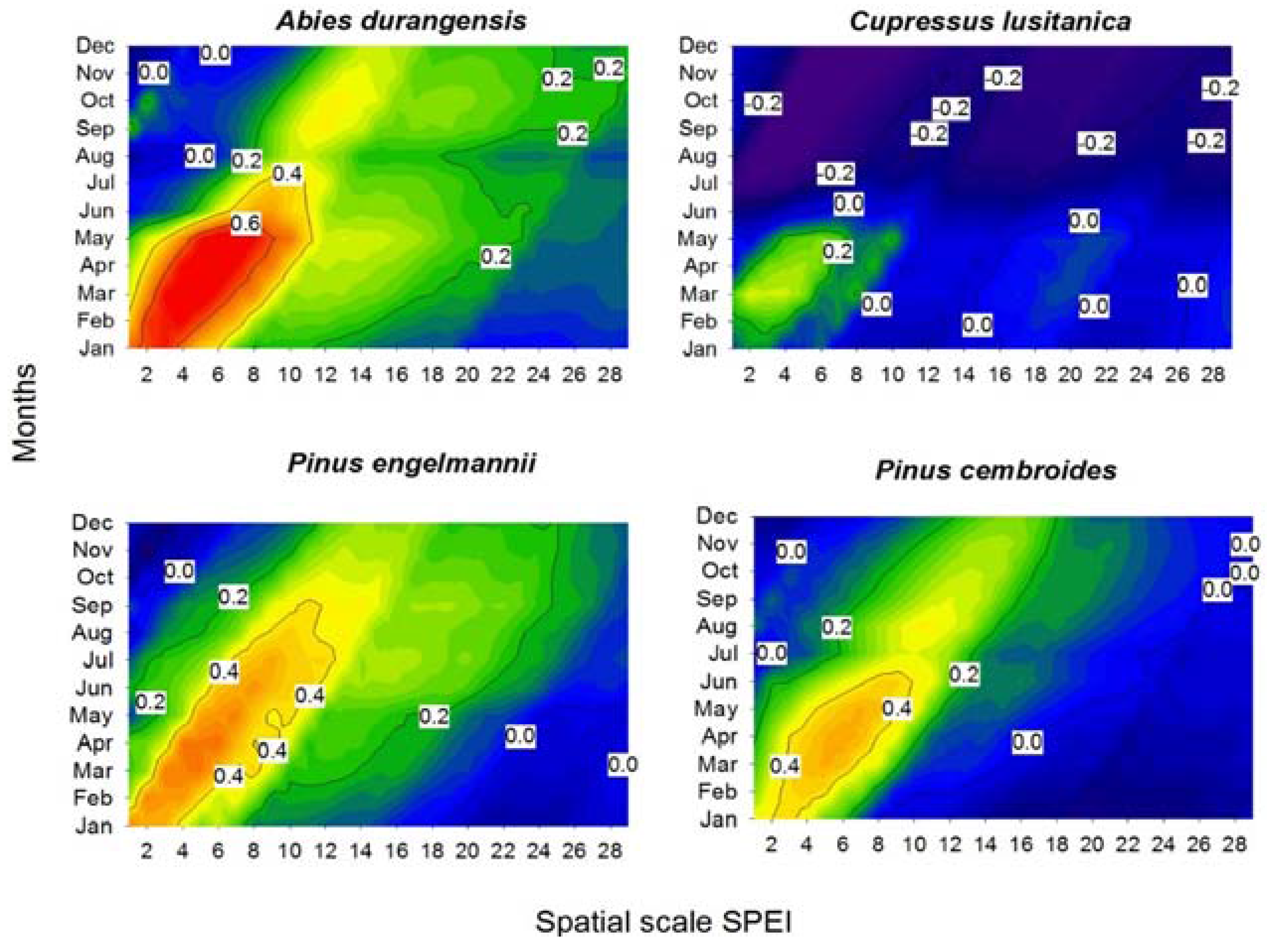
The growth of the four species showed a positive response to the SPEI calculated at scales of 1–8 months (Figure 4). Specifically, this correlation was the maximum for A. durangensis from January–July when considering 2–8 month long scales. For the xeric habitat, the drought-growth relationship intensified from two to 10 months from January–July in both pine species. Thus, A. durangensis was the species with the greatest response to drought in the mesic habitat (r = 0.81 in May for an estimated SPEI at a 7-month scale), while species from the xeric site such as P. engelmannii and P. cembroides showed moderate associations (r = 0.50–0.57 in April for SPEI calculated at a 6-month long scale) and C. lusitanica barely responded to drought (r = 0.35 in March for SPEI calculated at a 2-month long scale).
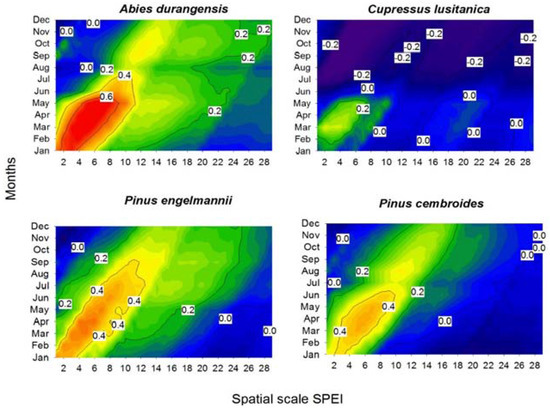
Figure 4.
Correlations between the standardized precipitation evapotranspiration index (SPEI) drought index and residual ring-width chronologies for the four study conifer species. The SPEI was calculated on a scale of 1–28 months (x-axis) from January–December (y-axis) of the growth year. The graphs show the Pearson correlation coefficients (r) using color scales. Values of r above 0.2 are significant (p < 0.05).
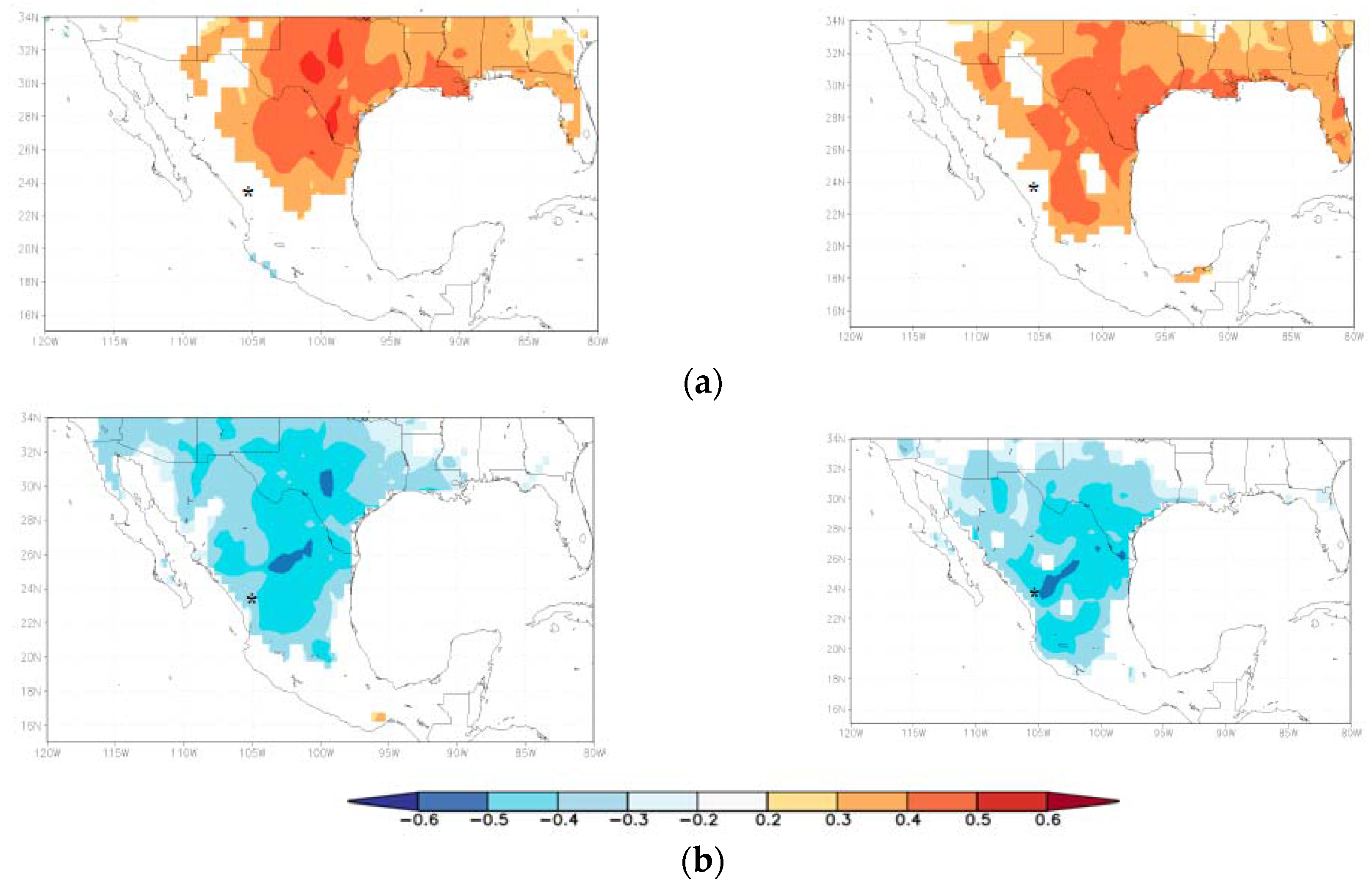
The growth of the four species was negatively associated with mean annual maximum temperature values in Northeastern Mexico (Figure 5a), and species of the xeric habitat responded to the temperatures of a wider territorial extent. This correlation seems to indicate higher rates of evapotranspiration and higher water deficit as temperature increased, as indicated by the positive correlations detected between the residual growth series and the SPEI drought index (Figure 5b).
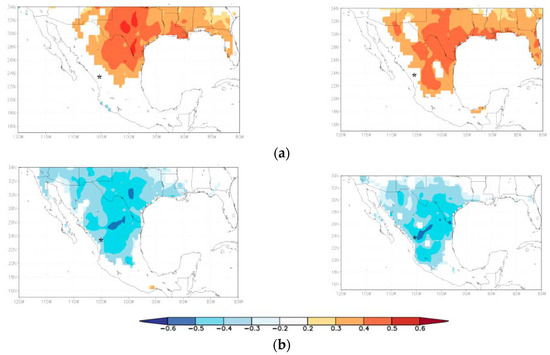
Figure 5.
Spatial correlations calculated by relating residual ring-width series of tree species from the mesic (A. durangensis, C. lusitanica; left plots), and xeric habitats (P. cembroides, P. engelmannii right plots) with (a) mean maximum temperatures; and (b) the 6-month long SPEI drought index. The asterisk indicates the approximate situation of the study area. Field correlations have p < 0.10 in all cases.
4. Discussion
We found that a fir (Abies) species dominant in mesic habitats (A. durangensis) showed the highest responsiveness to drought and water availability, thus refuting our main hypothesis and highlighting the relevance of dry spells as drivers of forest dynamics in sites where water availability is usually high.
The suitability of the species for dendrochronology was adequate, except for C. lusitanica (Table 1), which proved to be a complacent species with little responsiveness to hydroclimatic variability. This may be attributed to the humid sites (e.g., creeks) in which it often inhabits [46]. The dominance of C. lusitanica near streams and in the deepest parts of the canyons [46] assured sufficient water for carbohydrate metabolism and growth and explains why this species is not affected by warm (evapotranspirative demand) and dry conditions. As, to our knowledge, these are the first chronologies for C. lusitanica and A. durangensis, it was not possible to compare their dendrochronological parameters with the same species from other sites of Northern Mexico. The dendrochronological statistics obtained for P. engelmannii were similar to those presented by Bickford et al. [26], but higher than those reported by González-Cásares et al. [31] for the variables AC and Rbar. In the case of P. cembroides, the obtained results were different from those reported by Constante-García et al. [47] at a neighboring site, with higher and better values for the mean correlation between trees, which indicated a high common variance of tree growth patterns (e.g., probably also tree growth responses to climatic variability).
A common feature across all study species was their sensitivity to the winter-spring climate conditions. Studies at neighboring sites on similar species have demonstrated the positive influence of a wet and cool climate during this period on the growth of trees [31,34]. The conditions experienced by the trees during the previous winter are precursors of soil mixture during the early season of growth in spring, and determine the available water or the amount of reserves (e.g., carbohydrates, nutrients) accumulated at the beginning of the growth period [17,48]. This influence prior to the beginning of tree-ring formation could become more relevant due to the presence of warmer and drier winters, putting some forests in higher decline risks [25].
The response of A. durangensis to climate was characterized by its growth being enhanced by cold and rainy winters, which is in accordance with reports for similar Mexican conifer species [30]. High winter evaporation rates, associated with high maximum temperatures, promote moisture deficit by decreasing soil moisture reserves at the beginning of the growing season [49]. However, these results were in contrast with those reported for Silvefr fir (Abies alba) in the Spanish Pyrenees [32], which is more sensitive to late-summer drought stress. This difference could be attributed to the conditions of the thin, stony soils and high slopes in which A. durangensis grows, which result in a low water retention capacity.
In the case of P. engelmannii, this species benefited from warm and humid conditions in the months preceding the growing season, while drought conditions (higher Ev and Tmax values) decreased wood production. These conditions were expected, as they were previously observed in Mexican tree species of similar characteristics [31]. In addition, this species grew better under higher minimum temperatures in winter, and in response to cool and wet winter-spring conditions (see Figure 3). Elevated winter moisture can contribute to the formation of carbohydrates for the generation of earlywood during the following growing season and recharge the soil with water reserves before the ring starts to form [32], while drought conditions constrain growth [26]. As these last authors pointed out, water deficit may accentuate in sites of high slope, with stony and shallow soils (low water retention capacity), or areas exposed to the effects of dry winds that contribute to increased evaporation. P. engelmannii had similar climate-growth relationships to our results at a nearby site [31]. The growth response to SPEI was an indicator that P. engelmannii was sensitive to water deficit but it was less responsive to drought than A. durangensis (Figure 4). This lends itself to hypothesizing that P. engelmannii strategies to tolerate drought depend on the availability of water reserves in the soil, which should be checked in subsequent studies (study of root systems, inferences of water-use efficiency using carbon isotopes) [50]. Comparatively, P. cembroides showed greater adaptability to drought (it was the least responsive species at the xeric habitat) which conferred a differentiated phenological response. This tree species could take advantage of its short height and extended root system [51], despite living in poorer soil sites [48]. Its marked negative growth response to high evaporation rates (Figure 3) placed this species at a high degree of vulnerability to extreme weather events such as heat waves and dry spells.
The results of this study contribute data to better understand the responses of forests in mesic habitats to hydroclimatic variability, in the sense that these usually more humid places are also subject to drought stress [32]. For instance, A. durangensis (Figure 3 and Figure 4) showed high sensitivity to water deficit derived from high temperatures and high evapotranspiration rates, which had previously been observed in other fir species in drought-prone areas from Europe [11]. Our results indicate that dominant species in mesic habitats, such as fir species, respond to water deficit, particularly to atmospheric drought, which is supported by studies based on photosynthesis and water-use efficiency in other fir forests of Mexico [52] and the Mediterranean Basin [53]. Consequently, we expect that water-use efficiency inferred from carbon isotope data would be lower for A. durangensis than in the pine species studied at the xeric habitat. If this drought-sensitive species faces climate conditions which become increasingly hot and dry, the relative increase of water-use efficiency could be greater in this fir species (typical of mesic habitats) than in other tree species from xeric habitats such as P. engelmannii [50].
Moreover, despite the close proximity between species of the mesic habitat, there were microsite conditions that defined differentiated growth responses, as shown by the low sensitivity to drought of C. lusitanica, an abundant species in moist areas or near watercourses with nutrient-poor soils [54]. According to Pasho et al. [32], site-specific conditions, characterized by particular topographical features or soil types, contribute to decreasing soil water reserves and consequently alter the sensitivity of tree species and populations to drought.
The species of the xeric habitat showed similar vulnerability to drought, differentiated only by the temporality of the response to accumulated drought (Figure 4). P. engelmannii was slightly affected by longer droughts than P. cembroides, whereas P. cembroides seemed to gain a slight advantage over P. engelmannii, which can be attributed to different root systems [51]. The lower response of P. cembroides to drought suggested that it was more water efficient than P. engelmannii and better adapted to withstand prolonged water deficit. Additional factors such as stand density [22], neighborhood tree composition [23], and management history including thinning [23] could also affect our results. Management could partially explain some of the findings observed at the xeric habitat, which is managed with the selection system [28], whilst thinning seems to not be relevant in the protected mesic habitat.
It was also notable that the responses to temperature and drought at large spatial scales of the two most sensitive species at each site were similar (Figure 5). Geographically, our results suggested similar responses of pine growth to climate over northern Mexico [31]. This indicated that the differentiation in drought response between species occurred locally or between sites along the elevational and climatic gradient. Therefore, species susceptible to drought of mesic habitats such as A. durangensis are useful for reconstructing spatiotemporal hydroclimatic patterns. In this way, the results of the four species showed the absence of a strictly elevational pattern or differentiation in mesic and xeric habitats. The modulation of drought, according to contrasting soil moisture characteristics, seemed to differ from that reported in other studies [32] that showed greater response to drought at xeric habitats than at mesic habitats. Drought appeared to modulate growth depending on the conditions of the microsite, or may be dependent on the functional features of each species—which have yet to be sufficiently explored (growth rate, anatomy, and density of wood, etc.)—which is evident in the comparative responses of the two tree species coexisting at the mesic habitat. That is, that precipitation pattern does not regulate the expected growth in the mesic habitats versus the xeric habitats, but that drought does (Figure 4). This is crucial to the more intense and long-lasting droughts in the north of Mexico according to the predicted climatic scenarios, which may be unprecedented when compared to the last six centuries of data [42].
Further research is required to explore whether other drivers are able to explain drought responses. For instance, the similar response to drought of the two species from the xeric site can be strongly influenced by the phylogenetic relatedness of the conifer species. In addition, extreme climate events as severe drought should be considered a relevant disturbance to understand forest dynamics even in mesic habitats (e.g., [55]), as our findings with A. durangensis reveal.
This study represents a preliminary attempt at determining the vulnerability threshold of four Mexican conifers to drought. This effort is very relevant given the warmer and more arid climatic scenarios predicted for the study region [43]. However, determining such a threshold requires obtaining multiple and often difficult measures and variables such as: xylem cavitation [56], crown die-back [57], or growth data of healthy and declining trees [58]. Future studies should deal with those measures so as to integrate them in vulnerability thresholds of tree species in response to drought.
5. Conclusions
The growth responses to drought changed according to the conifer species and the local conditions of the site. In the mesic habitat, we found species with a high responsiveness to drought such as A. durangensis, whilst C. lusitanica did not respond to hydroclimate variability. In the xeric habitat, P. engelmannii was slightly more sensitive than P. cembroides. Multi-specific studies encompassing climatic and environmental gradients represent a first step towards defining vulnerability thresholds of tree species in the face of more frequent severe droughts.
Acknowledgments
Funding was provided by CONACYT (Consejo Nacional de Ciencia y Tecnología) through the CB-2013/222522 project. Many thanks to the community-based site known as “Ejido el Brillante”, and we also thank the forester responsible for the area (Javier Bretado) for supporting the data gathering. We thank the Dirección General de Vida Silvestre, SEMARNAT (Secretaría de Medio Ambiente y Recursos Naturales), Mexico, for providing technical facilities. The authors are grateful to the editors and anonymous reviewers for their useful comments and suggestions.
Author Contributions
M.P.-G. conceived and designed the experiments; M.P.-G., M.G.-C., A.C.A.-H. and M.R.-C. performed the experiments; J.J.C. analyzed the data. All authors wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Intergovernmental Panel on Climate Change (IPCC). Summary for Policymakers. In Climate Change 2013: The Physical Science Basis; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Dai, A. Increasing drought under global warming in observations and models. Nat. Clim. Change 2013, 3, 52–58. [Google Scholar] [CrossRef]
- Bigler, C.; Gavin, D.G.; Gunning, C.; Veblen, T.T. Drought induces lagged tree mortality in a subalpine forest in the Rocky Mountains. Oikos 2007, 116, 1983–1994. [Google Scholar] [CrossRef]
- Sánchez-Salguero, R.; Navarro, R.M.; Camarero, J.J.; Fernández-Cancio, Á. Drought-induced growth decline of Aleppo and maritime pine forests in south-eastern Spain. For. Syst. 2010, 19, 458–470. [Google Scholar] [CrossRef]
- Anderegg, W.R.; Hicke, J.A.; Fisher, R.A.; Allen, C.D.; Aukema, J.; Bentz, B.; Hood, S.; Lichstein, J.W.; Macalady, A.K.; McDowell, N.; et al. Tree mortality from drought, insects, and their interactions in a changing climate. New Phytol. 2015, 208, 674–683. [Google Scholar] [CrossRef] [PubMed]
- Linares, J.C.; Camarero, J.J.; Bowker, M.A.; Ochoa, V.; Carreira, J.A. Stand-structural effects on Heterobasidion abietinum-related mortality following drought events in Abies pinsapo. Oecologia 2010, 164, 1107–1119. [Google Scholar] [CrossRef] [PubMed]
- Camarero, J.J.; Gazol, A.; Sangüesa-Barreda, G.; Oliva, J.; Vicente-Serrano, S.M. To die or not to die: Early warnings of tree dieback in response to a severe drought. J. Ecol. 2015, 103, 44–57. [Google Scholar] [CrossRef]
- Chhin, S. Influence of climate on the growth of hybrid poplar in Michigan. Forests 2010, 1, 209–229. [Google Scholar] [CrossRef]
- Brzostek, E.R.; Dragoni, D.; Schmid, H.P.; Rahman, A.F.; Sims, D.; Wayson, C.A.; Johnson, D.J.; Phillips, R.P. Chronic water stress reduces tree growth and the carbon sink of deciduous hardwood forests. Glob. Chang. Biol. 2014, 20, 2531–2539. [Google Scholar] [CrossRef] [PubMed]
- Correa-Díaz, A.; Gómez-Guerrero, A.; Villanueva-Díaz, J.; Castruita-Esparza, L.U.; Martínez-Trinidad, T.; Cervantes-Martínez, R. Análisis dendroclimático de Ahuehuete (Taxodium mucronatum Ten.) en el centro de México. Agrociencia 2014, 48, 537–551. [Google Scholar]
- Vicente-Serrano, S.M.; Camarero, J.J.; Zabalza, J.; Sangüesa-Barreda, G.; López-Moreno, J.I.; Tague, C.L. Evapotranspiration deficit controls net primary production and growth of silver fir: Implications for circum-Mediterranean forests under forecasted warmer and drier conditions. Agric. For. Meteorol. 2015, 206, 45–54. [Google Scholar] [CrossRef]
- Rodríguez-Catón, M.; Villalba, R.; Morales, M.; Srur, A. Influence of droughts on Nothofagus pumilio forest decline across northern Patagonia, Argentina. Ecosphere 2016, 7, 1–17. [Google Scholar] [CrossRef]
- Sánchez-Salguero, R.; Camarero, J.J.; Gutiérrez, E.; González-Rouco, F.; Gazol, A.; Sangüesa-Barreda, G.; Andreu-Hayles, L.; Linares, J.C.; Seftigen, K. Assessing forest vulnerability to climate warming using a process-based model of tree growth: Bad prospects for rear-edges. Glob. Chang. Biol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Adams, H.D.; Kolb, T.E. Tree growth response to drought and temperature along an elevation gradient on a mountain landscape. J. Biogeogr. 2005, 32, 1629–1640. [Google Scholar] [CrossRef]
- Linares, J.C.; Tíscar, P.A. Climate change impacts and vulnerability of the southern populations of Pinus nigra subsp. salzmannii. Tree Physiol. 2010, 30, 795–806. [Google Scholar] [CrossRef] [PubMed]
- Suarez, L.; Kitzberger, T. Differential effects of climate variability on forest dynamics along a precipitation gradient in northern Patagonia. J. Ecol. 2010, 98, 1023–1034. [Google Scholar] [CrossRef]
- Pompa-García, M.; Cerano-Paredes, J.; Fulé, P.Z. Variation in radial growth of Pinus cooperi response to climatic signals across an elevational gradient. Dendrochronologia 2013, 31, 198–204. [Google Scholar] [CrossRef]
- Sanders, T.G.M.; Heinrich, I.; Günther, B.; Beck, W. Increasing water use efficiency comes at a cost for Norway spruce. Forests 2016, 7, 296. [Google Scholar] [CrossRef]
- Bunn, A.G.; Hughes, M.K.; Salzer, M.W. Topographically modified tree-ring chronologies as a potential means to improve paleoclimate inference. Clim. Chang. 2011, 105, 627–634. [Google Scholar] [CrossRef]
- Rodríguez-Catón, M.; Villalba, R.; Srur, A.M.; Luckman, B. Long-term trends in radial growth associated with Nothofagus pumilio forest decline in Patagonia: Integrating local- into regional-scale patterns. For. Ecol. Manag. 2015, 339, 44–56. [Google Scholar] [CrossRef]
- Sangüesa-Barreda, G.; Camarero, J.J.; Linares, J.C.; Hernández, R.; Oliva, J.; Gazol, A.; González, E.; Montes, F.; García-Martín, A.; De la Riva, J. Papel de los factores bióticos y las sequías en el decaimiento del bosque: Aportaciones desde la dendroecología. Ecosistemas 2015, 24, 15–23. [Google Scholar] [CrossRef]
- Jiang, Z.H.; Wang, X.Q.; Fei, B.H.; Ren, H.Q.; Liu, X.E. Effect of stand and tree attributes on growth and wood quality characteristics from a spacing trial with Populus xiaohei. Ann. For. Sci. 2007, 64, 807–814. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schütze, G.; Uhl, E. Resistance of European tree species to drought stress in mixed versus pure forests: Evidence of stress release by inter-specific facilitation. Plant Biol. 2013, 15, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Sohn, J.A.; Saha, S.; Bauhus, J. Potential of forest thinning to mitigate drought stress: A meta-analysis. For. Ecol. Manag. 2016, 380, 261–273. [Google Scholar] [CrossRef]
- Pompa-García, M.; Camarero, J.J. Reconstructing evaporation from pine tree rings in northern Mexico. Tree Ring Res. 2015, 71, 95–105. [Google Scholar] [CrossRef]
- Bickford, I.N.; Fulé, P.Z.; Kolb, T.E. Growth sensitivity to drought of co-occurring Pinus spp. along an elevation gradient in northern Mexico. West. N. Am. Nat. 2011, 71, 338–348. [Google Scholar] [CrossRef]
- González-Elizondo, M.S.; González-Elizondo, M.; Tena-Flores, J.A.; Ruacho-González, L.; López-Enríquez, I.L. Vegetación de la Sierra Madre Occidental, México: Una síntesis. Acta Bot. Mex. 2012, 100, 351–403. [Google Scholar] [CrossRef]
- Pompa-García, M.; Domínguez-Calleros, P.A. Respuesta de madera temprana y tardía a la sequía en una conífera mexicana bajo dos condiciones ecológicas. Rev. Ecosistemas 2015, 24, 37–42. [Google Scholar] [CrossRef]
- Cerano, J.; Rivera, M.; Estrada, J.; Trucios, R.; Rios, J.C. Análisis dendrocronológico de Pinus cooperi en Durango, México. Agrofaz 2012, 12, 81–88. [Google Scholar]
- Villanueva, J.; Cerano, J.; Fulé, P.Z.; Cortés, C.; Vázquez, L.; Yocom, L.L.; Ruiz-Corral, J.A. Cuatro siglos de variabilidad hidroclimática en el noroeste de Chihuahua, México, reconstruida con anillos de árboles. Investig. Geogr. 2015, 87, 141–153. [Google Scholar] [CrossRef]
- González-Cásares, M.; Pompa-García, M.; Camarero, J.J. Differences in climate–growth relationship indicate diverse drought tolerances among five pine species coexisting in Northwestern Mexico. Trees 2016, 31, 531–544. [Google Scholar] [CrossRef]
- Pasho, E.; Camarero, J.J.; De Luis, M.; Vicente-Serrano, S.M. Impacts of drought at different time scales on forest growth across a wide climatic gradient in north-eastern Spain. Agric. For. Meteorol. 2011, 151, 1800–1811. [Google Scholar] [CrossRef]
- Aguirre-Díaz, G.J.; Labarthe-Hernández, G. Fissure ignimbrites: Fissure-source origin for voluminous ignimbrites of the Sierra Madre Occidental and its relationship with Basin and Range faulting. Geology 2003, 31, 773–776. [Google Scholar] [CrossRef]
- González-Elizondo, M.; Jurado, E.; Návar, J.; González-Elizondo, M.S.; Villanueva, J.; Aguirre, O.; Jiménez, J. Tree-rings and climate relationships for Douglas-fir chronologies from the Sierra Madre Occidental, Mexico: A 1681–2001 rain reconstruction. For. Ecol. Manag. 2005, 213, 39–53. [Google Scholar] [CrossRef]
- Wigley, T.M.L.; Briffa, K.R.; Jones, P.D. On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J. Appl. Meteorol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Holmes, R.L. Computer-assisted quality control in treering dating and measurement. Tree Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Cook, E.R.; Kairiukstis, L.A. Methods of Dendrochronology: Applications in the Environmental Science; Kluwer: Alphen aan den Rijn, The Netherlands, 1990; p. 394. [Google Scholar]
- Bunn, A.G. A dendrochronology program library in R (dplR). Dendrochronologia 2008, 26, 115–124. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Comisión Nacional del Agua (CNA). Datos Climáticos de Estaciones Meteorológicas de Durango: El Salto, Otinapa. México, 2016. Available online: http://smn1.conagua.gob.mx/index.php?option =com_content& view=article&id=180:durango&catid=14:normales-por-estacion (accessed on 10 February 2017).
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A multiscalar drought index sensitive to global warming: The Standardized Precipitation Evapotranspiration Index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef]
- Beguería, S.; Vicente-Serrano, S.M. SPEI: Calculation of the Standardised Precipitation-Evapotranspiration Index, R Package Version 1.6. 2013. Available online: http://CRAN.R-project.org/package=SPEI (accessed on 10 February 2017).
- Stahle, D.W.; Cook, E.R.; Burnette, D.J.; Villanueva, J.; Cerano, J.; Burns, J.N.; Griffin, D.; Cook, B.I.; Acuña, R.; Torbenson, M.C.A.; et al. The Mexican Drought Atlas: Tree-ring reconstructions of the soil moisture balance during the late pre-Hispanic, colonial, and modern eras. Quat. Sci. Rev. 2016, 149, 34–60. [Google Scholar] [CrossRef]
- Harris, I.; Jones, P.D.; Osborn, T.J.; Lister, D.H. Updated high-resolution grids of monthly climatic observations—The CRU TS3.10 Dataset. Int. J. Climatol. 2014, 34, 623–642. [Google Scholar] [CrossRef]
- Van Oldenborgh, G.J.; Burgers, G. Searching for decadal variations in ENSO precipitation teleconnections. Geophys. Res. Lett. 2005, 32, L15701. [Google Scholar] [CrossRef]
- García-Arévalo, A. Vegetación y flora de un bosque relictual de Picea chihuahuana Martínez del norte de México. Polibotánica 2008, 25, 45–68. [Google Scholar]
- Constante-García, V.; Villanueva-Díaz, J.; Cerano-Paredes, J.; Cornejo-Oviedo, E.H.; Valencia-Manzo, S. Dendrocronología de Pinus cembroides Zucc. y reconstrucción de precipitación estacional para el Sureste de Coahuila. Cienc. For. Méx. 2009, 34, 17–39. [Google Scholar]
- Pompa-García, M.; Yerena-Yamalliel, J.I. Concentración de carbono en Pinus cembroides Zucc: Fuente potencial de mitigación del calentamiento global. Rev. Chapingo Ser. Cie. 2014, 20, 169–175. [Google Scholar]
- Pasho, E.; Camarero, J.J.; Vicente-Serrano, S.M. Climatic impacts and drought control of radial growth and seasonal wood formation in Pinus halepensis. Trees 2012, 26, 1875–1886. [Google Scholar] [CrossRef]
- Castruita-Esparza, L.U. Variabilidad Climática, Eficiencia de uso de Agua Intrínseca y Crecimiento del Área Basal en Bosques del Norte de México. Ph.D. Thesis, Colegio de Postgraduados, Texcoco, México, 2014. [Google Scholar]
- Gutiérrez-García, J.V.; Rodríguez-Trejo, D.A.; Villanueva-Morales, A.; García-Díaz, S.; Romo-Lozano, J.L. Calidad del agua en la producción de Pinus cembroides Zucc. en vivero. Agrociencia 2015, 49, 205–219. [Google Scholar]
- Gómez-Guerrero, A.; Silva, L.C.; Barrera-Reyes, M.; Kishchuk, B.; Velázquez-Martínez, A.; Martínez-Trinidad, T.; Plascencia-Escalante, F.O.; Horwath, W.R. Growth decline and divergent tree ring isotopic composition (δ13C and δ18O) contradict predictions of CO2 stimulation in high altitudinal forests. Glob. Change Biol. 2013, 19, 1748–1758. [Google Scholar] [CrossRef] [PubMed]
- Linares, J.C.; Camarero, J.J. From pattern to process: Linking intrinsic water-use efficiency to drought-induced forest decline. Glob. Chang. Biol. 2012, 18, 1000–1015. [Google Scholar] [CrossRef]
- Farjon, A. A Monograph of Cupressaceae and Sciadopitys; Royal Botanic Gardens: Kew, UK, 2005. [Google Scholar]
- Pederson, N.; Dyer, J.M.; McEwan, R.W.; Hessl, A.E.; Mock, C.J.; Orwig, D.A.; Rieder, H.E.; Cook, B.I. The legacy of episodic climatic events in shaping temperate, broadleaf forests. Ecol. Monogr. 2014, 84, 599–620. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Flint, A.; Huang, C.Y.; Flint, L.; Berry, J.A.; Davis, F.W.; Sperry, J.S.; Field, C.B. Tree mortality predicted from drought-induced vascular damage. Nat. Geosci. 2015, 8, 367–371. [Google Scholar] [CrossRef]
- Chakrabortya, T.; Sahab, S.; Matzarakisd, A.; Reifa, A. Influence of multiple biotic and abiotic factors on the crown die-back of European beech trees at their drought limit. Flora 2017, 229, 58–70. [Google Scholar] [CrossRef]
- Cavin, L.; Mountford, E.P.; Peterken, G.F.; Jump, A.S. Extreme drought alters competitive dominance within and between tree species in a mixed forest stand. Funct. Ecol. 2013, 27, 1424–1435. [Google Scholar] [CrossRef]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).