
Red Alder-Conifer Stands in Alaska: An Example of Mixed Species Management to Enhance Structural and Biological Complexity



{kind=link}
{kind=link}
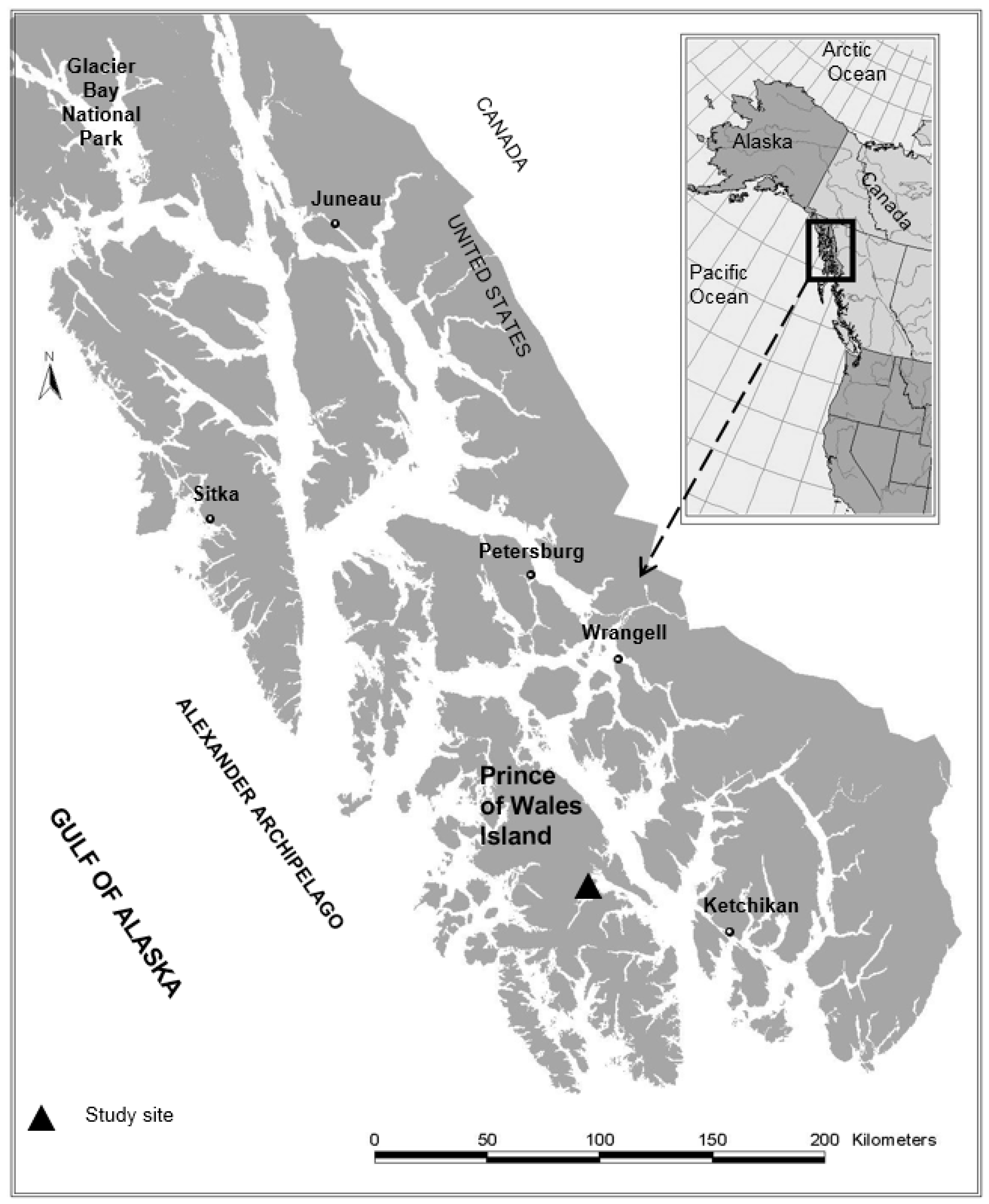{kind=link}
{kind=link}
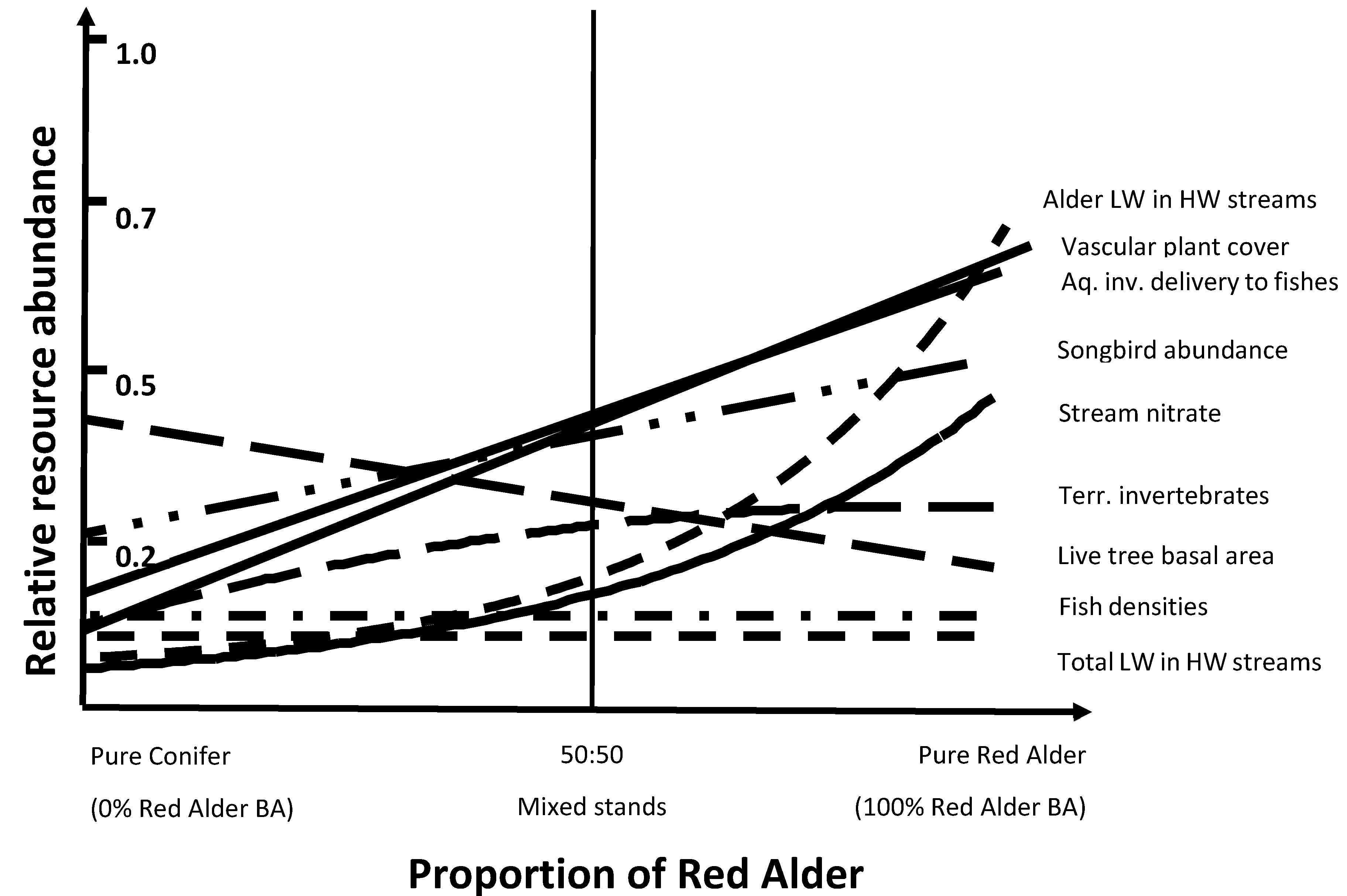{kind=link}
{kind=link}
Abstract
:1. Introduction
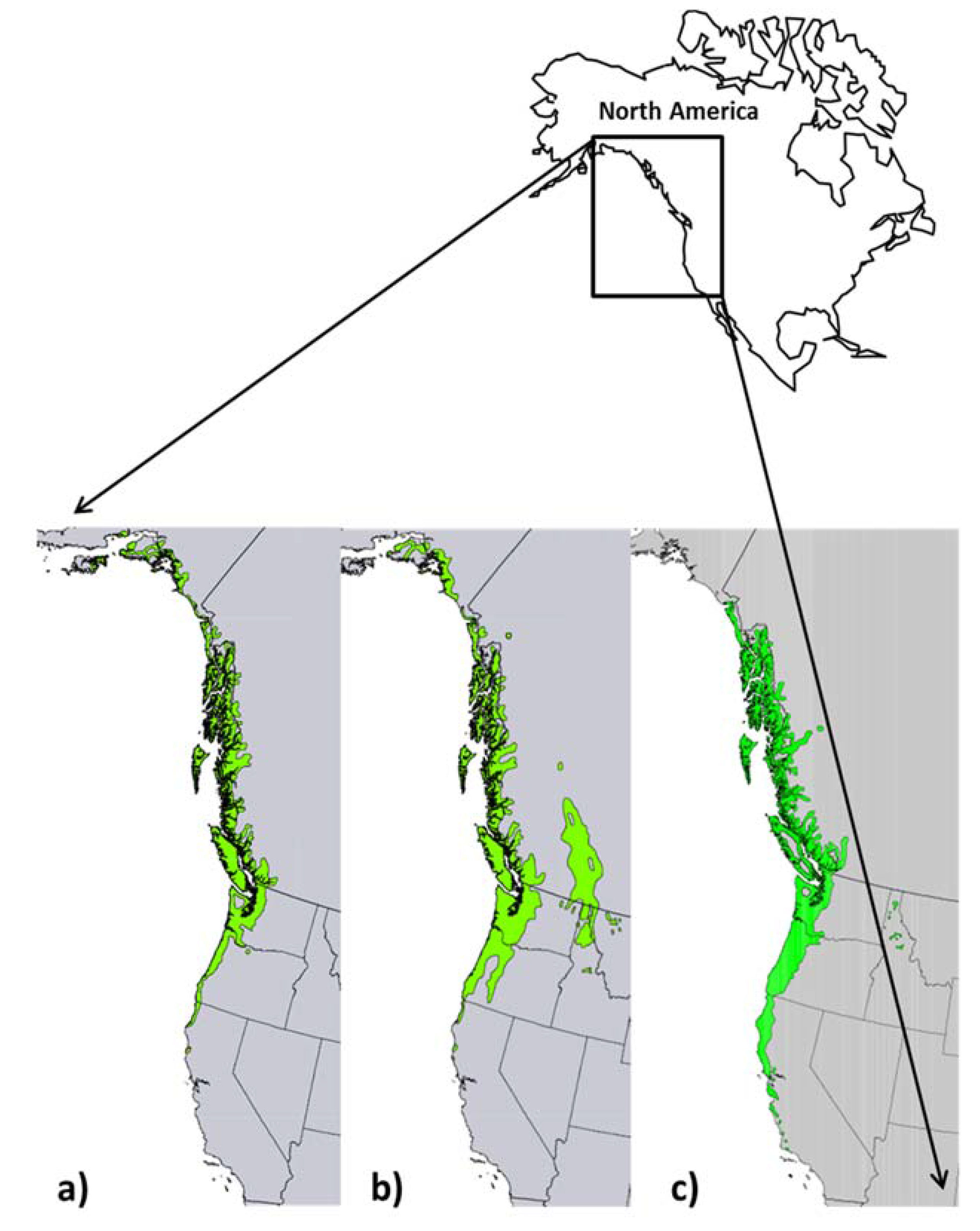
2. Distribution of Species
2.1. Range and Habitat of Sitka Spruce
2.2. Range and Habitat of Western Hemlock
2.3. Range and Habitat of Red Alder
3. Silvics and Species Growth of Natural Stands
3.1. Silvics of Spruce and Hemlock
3.2. Silvics of Red
3.3. Eight and Stand Growth of Spruce, Hemlock and Alder
4. Mixed Red Alder-Conifer Stands
4.1. Mixed Red Alder-Western Hemlock/Sitka Spruce Stands in Southeast Alaska
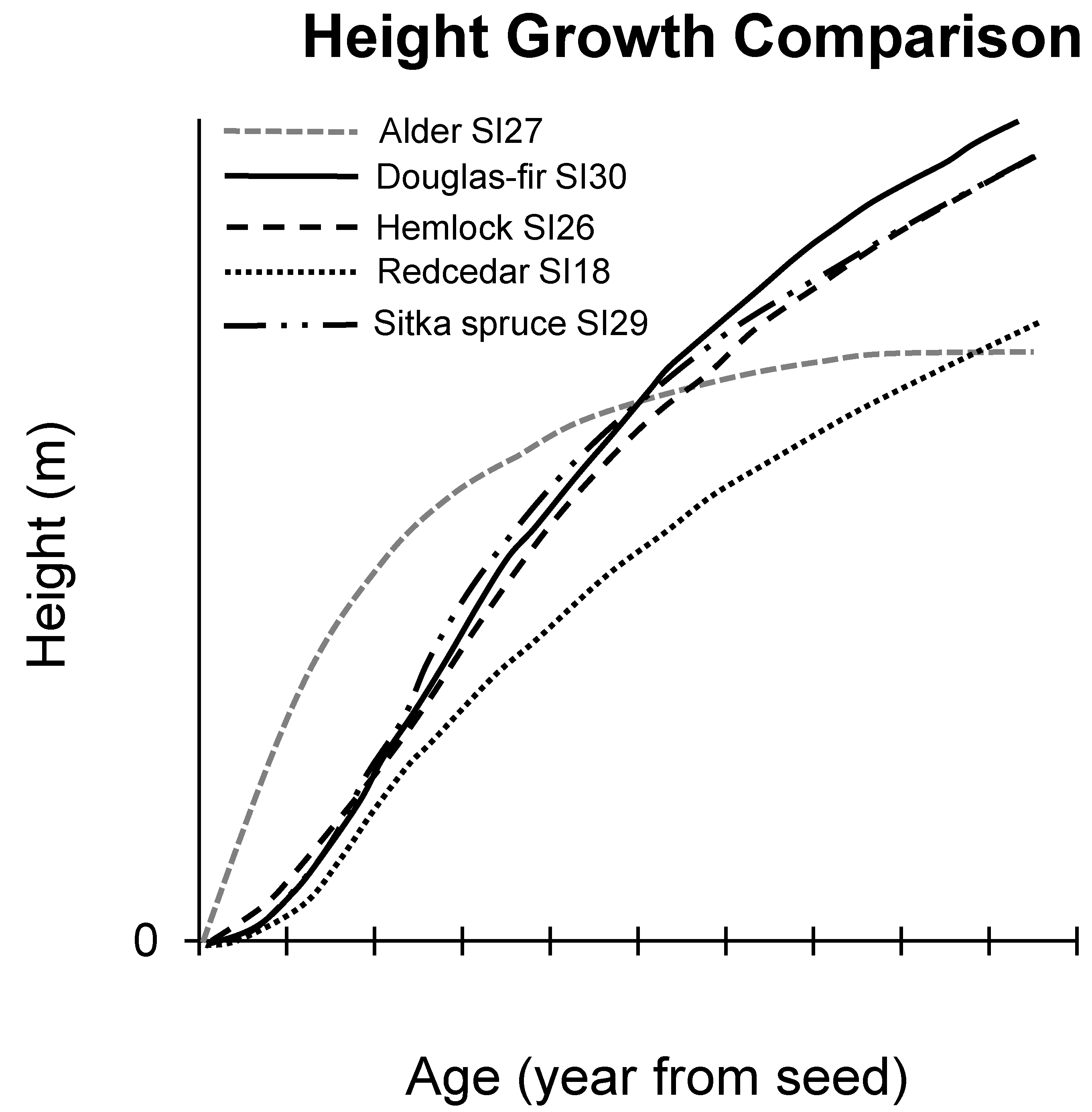
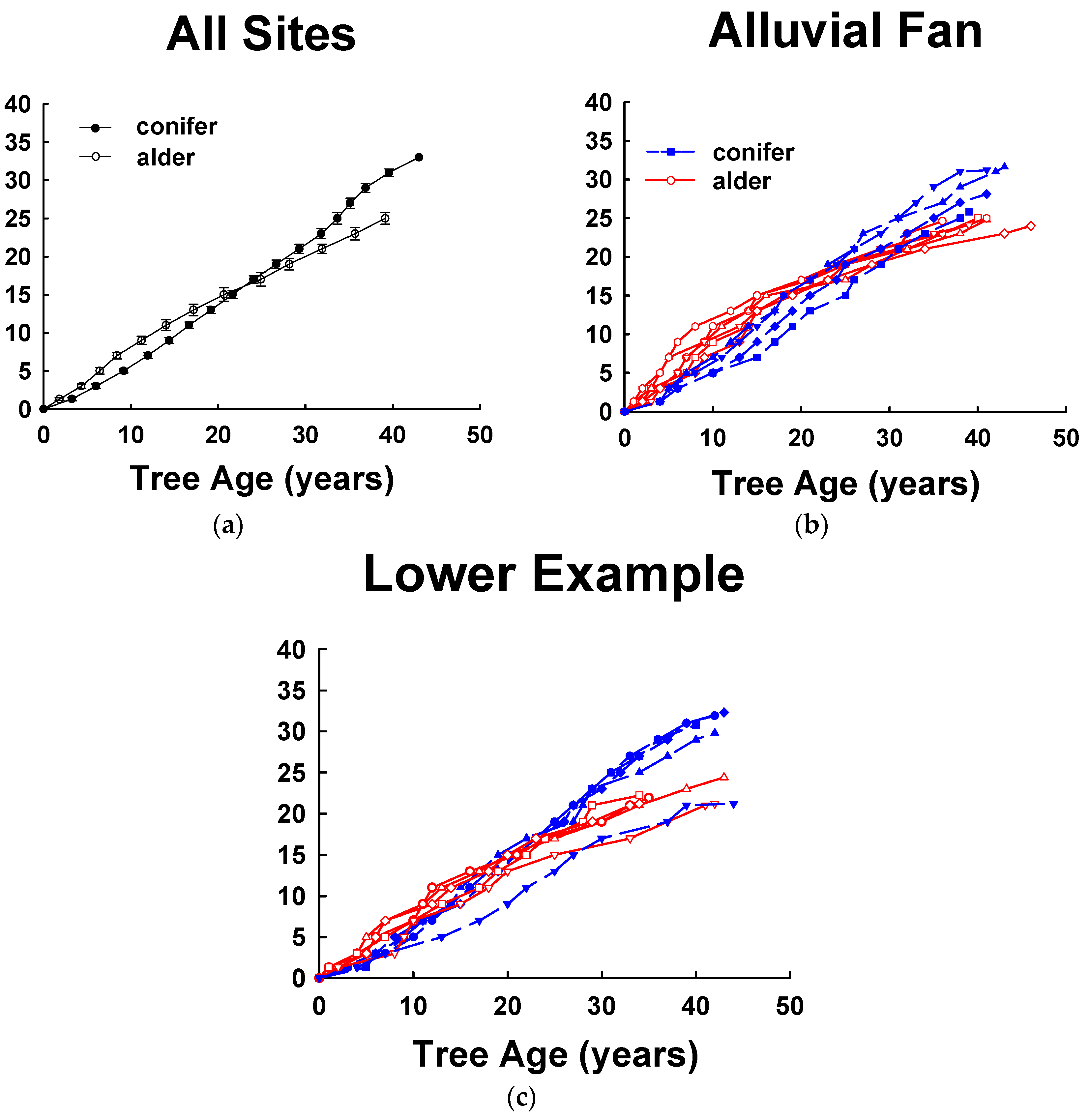
4.2. Tree Height Growth of Alder-Conifer Stands
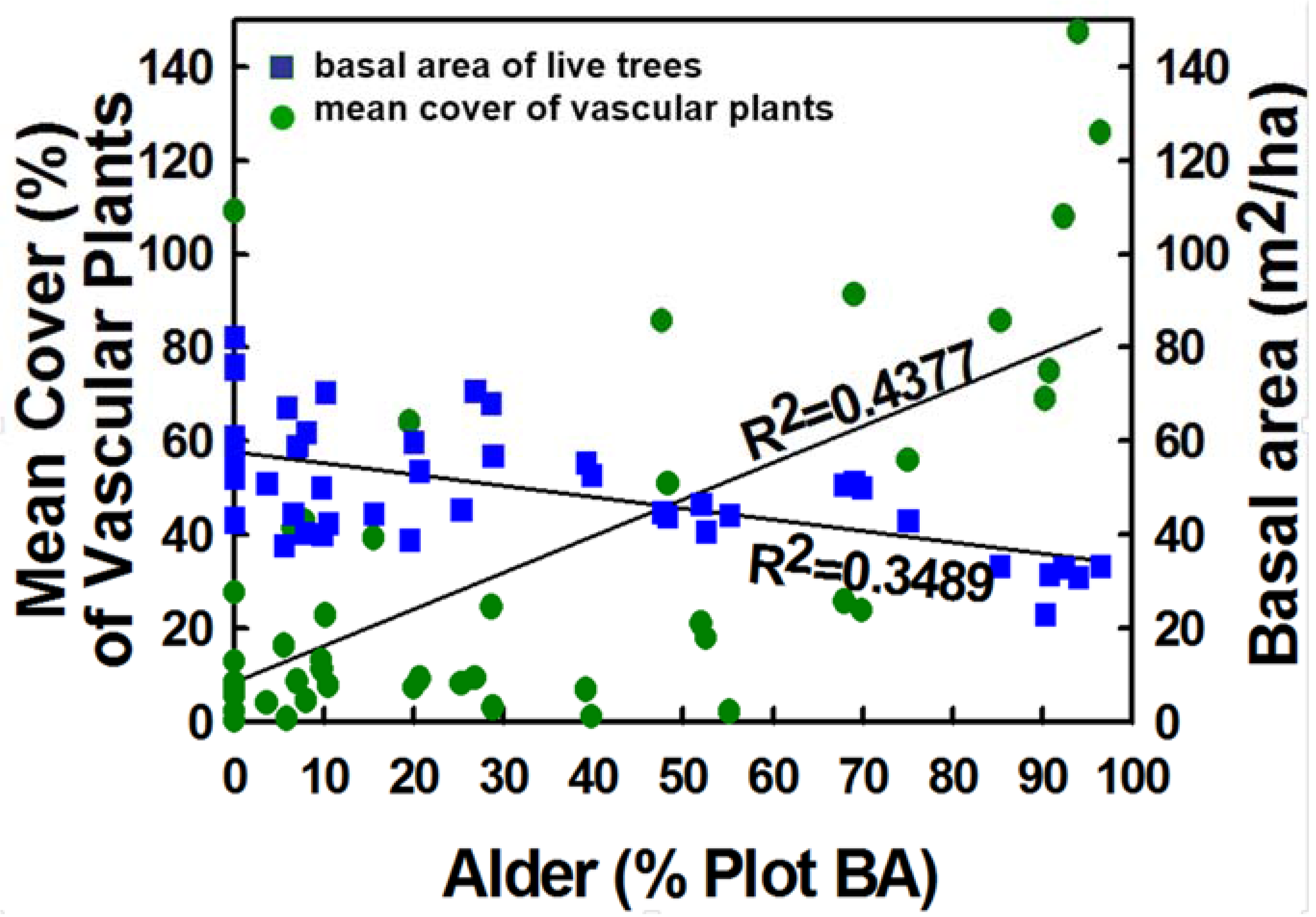
4.3. Stand Structure of Alder-Conifer Stands
4.4. Understory Plant Diversity and Wildlife Habitat of Alder-Conifer Stands
4.5. Aquatic and Riparian Habitat of Alder-Conifer Stands
5. Functional Role of Mixed Red Alder-Conifer Forests
5.1. Role of Red Alder in Young-Growth Forests
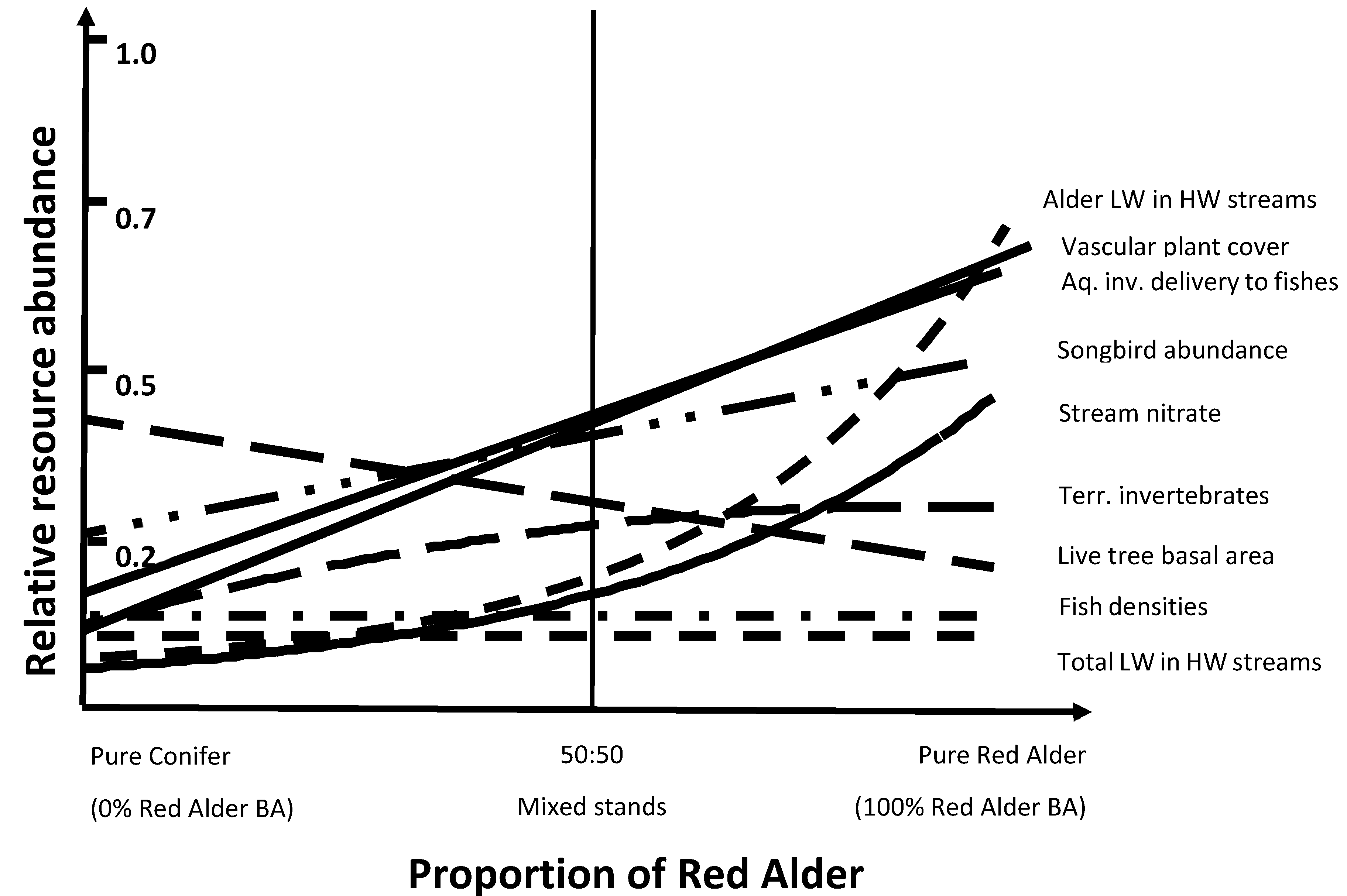
5.2. Improved Ecosystem Resilience of Mixed Alder-Conifer Stands
5.3. Role of Red Alder in Aquatic and Riparian Ecosystems
5.4. Compatibility and Potential Tradeoffs for the Inclusion of Alder in Conifer Forests
6. Summary
Acknowledgments
Conflicts of Interest
References
- Daily, G.C. Nature’s Services; Island Press: Washington, DC, USA, 1997; p. 392. [Google Scholar]
- Costanza, R.; d’Arge, R.; De Groot, R.; Farber, S.; Grasso, M.; Hannon, B.; Limburg, K.; Naeem, S.; O’Neill, R.V.; Paruelo, J.; et al. The value of the world’s ecosystem services and natural capital. Nature 1997, 387, 253–260. [Google Scholar] [CrossRef]
- MEA (Millenium Ecosystem Assessment). Ecosystems and Human Well-being: Synthesis; Island Press: Washington, DC, USA, 2005; p. 155. [Google Scholar]
- Franklin, J.F. Toward a new forestry. Am. For. 1989, 95, 37–44. [Google Scholar]
- Larsen, J.B. Ecological stability of forests and sustainable silviculture. For. Ecol. Manag. 1995, 73, 85–96. [Google Scholar] [CrossRef]
- Humphrey, J.W. Benefits to biodiversity from developing old-growth conditions in British upland spruce plantations: A review and recommendations. Forestry 2005, 78, 33–53. [Google Scholar] [CrossRef]
- Deal, R.L. Management strategies to increase stand structural diversity and enhance biodiversity in coastal rainforests of Alaska. Biol. Conserv. 2007, 137, 520–532. [Google Scholar] [CrossRef]
- Notaro, S.; Paletto, A.; Raffaelli, R. The economic valuation of non-productive forest functions as an instrument towards integrated forest management. The multifunctional role of forests—Policies, methods and case studies. EFI Proc. 2008, 55, 55–380. [Google Scholar]
- Deal, R.L.; Hennon, P.; O’Hanlon, R.; D’Amore, D. Lessons from native spruce forests in Alaska: Managing Sitka spruce plantations worldwide to benefit biodiversity and ecosystem services. Forestry 2014, 87, 193–208. [Google Scholar] [CrossRef]
- Grilli, G.; Jonkisz, J.; Ciolli, M.; Lesinski, J. Mixed forests and ecosystem services: Investigating stakeholders’ perceptions in a case study in the Polish Carpathians. For. Policy Econ. 2016, 66, 11–17. [Google Scholar] [CrossRef]
- Davis, L.S.; Johnson, K.N. Forest Management, 3rd ed.; McGraw-Hill Inc.: New York, NY, USA, 1987; p. 790. [Google Scholar]
- Fox, T.R.; Jokela, E.J.; Allen, H.L. The development of pine plantation silviculture in the southern United States. J. For. 2007, 105, 337–347. [Google Scholar]
- Evans, J.; Turnbull, J. Plantation Forestry in the Tropics, 3rd ed.; Oxford University Press: Oxford, UK, 2004; p. 480. [Google Scholar]
- Huffaker, C.B. (Ed.) Theory and Practice of Biological Control; Academic Press Inc.: New York, NY, USA, 2012; p. 810. [Google Scholar]
- Boyer, W.D. Pinus palustris Mill. Longleaf pine. In Silvics of North America; USDA Forest Service: Washington, DC, USA, 1990; pp. 405–412. [Google Scholar]
- McLaren, J.P.; Grace, J.C.; Kimberley, M.O.; Knowles, R.L.; West, G.G. Height growth of Pinus radiata as affected by stocking. N. Z. J. For. Sci. 1995, 25, 73–90. [Google Scholar]
- Kimberley, M.; West, G.; Dean, M.; Knowles, L. The 300 Index—A volume productivity index for radiate pine. N. Z. J. For. Sci. 2005, 30, 13–18. [Google Scholar]
- Salas, C.; Donoso, P.J.; Vargas, R.; Arriagada, C.A.; Pedraza, R.; Cesar, A.; Soto, D.P. The Forest Sector in Chile: An Overview and Current Challenges. J. For. 2016, 114. [Google Scholar] [CrossRef]
- Franklin, J.F.; Cromack, K.J.; Denison, W.; McKee, A.; Maser, C.; Sedell, J.; Swanson, F.; Juday, G. Ecological Characteristics of Old-Growth Douglas-Fir Forests; USDA Forest Service: Portland, OR, USA, 1981; p. 48.
- Hanley, T.A. Balancing economic development, biological conservation, and human culture: The Sitka black-tailed deer (Odocoileus hemionus sitkensis) as an ecological indicator. Biol. Conserv. 1993, 66, 61–67. [Google Scholar] [CrossRef]
- Gamfeldt, L.; Snall, T.; Bagchi, R.; Jonsson, M.; Gustafsson, L.; Kjellander, P.; Ruiz-Jaen, M.C.; Froberg, M.; Stendahl, J.; Philipson, C.D.; et al. Higher levels of multiple ecosystem services are found in forests with more tree species. Nat. Commun. 2013, 4, 1340. [Google Scholar] [CrossRef] [PubMed]
- Piotto, D. A meta-analysis comparing tree growth in monocultures and mixed plantations. For. Ecol. Manag. 2008, 255, 781–786. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; Mace, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity loss and its impact on humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Kelty, M.J. The role of species mixtures in plantation forestry. For. Ecol. Manag. 2006, 233, 195–204. [Google Scholar] [CrossRef]
- Humphrey, J.W.; Ferris, R.; Quine, C. Biodiversity in Britain’s Planted Forests; Forestry Commission: Edinburgh, UK, 2003; p. 126. [Google Scholar]
- Iremonger, S.; O’Halloran, J.; Kelly, D.L.; Wilson, M.W.; Smith, G.F.; Gittings, T.; Giller, P.S.; Mitchell, F.J.G.; Oxbrough, A.; Coote, L.; et al. Biodiversity in Irish Plantation Forests; Environmental Protection Agency: Wexford, Ireland, 2007; p. 69. [Google Scholar]
- Felton, A.; Lindbladh, M.; Brunet, J.; Fritz, Ö. Replacing coniferous monocultures with mixed-species production stands: An assessment of the potential benefits for forest biodiversity in northern Europe. For. Ecol. Manag. 2010, 260, 939–947. [Google Scholar] [CrossRef]
- Rothe, A.; Binkley, D. Nutritional interactions in mixed species forests: A synthesis. Can. J. For. Res. 2001, 31, 1855–1870. [Google Scholar] [CrossRef]
- Berger, T.W.; Inselsbacher, E.; Mutsch, F.; Pfeffer, M. Nutrient cycling and soil leaching in eighteen pure and mixed stands of beech (Fagus sylvatica) and spruce (Picea abies). For. Ecol. Manag. 2009, 258, 2578–2592. [Google Scholar] [CrossRef]
- Pretzsch, H.; Block, J.; Dieler, J.; Dong, P.H.; Kohnle, U.; Nagel, J.; Spellmann, H.; Zingg, A. Comparison between the productivity of pure and mixed stands of Norway spruce and European beech along an ecological gradient. Ann. For. Sci. 2010, 67, 712. [Google Scholar] [CrossRef]
- Griess, V.C.; Knoke, T. Growth performance, windthrow, and insects: Meta-analyses of parameters influencing performance of mixed-species stands in boreal and northern temperate biomes. Can. J. For. Res. 2011, 41, 1141–1159. [Google Scholar] [CrossRef]
- Harris, A.S. Picea sitchensis (Bong.) Carr. Sitka spruce. In Silvics of North America; Burns, R.M., Honkala, B.H., Eds.; USDA Forest Service: Washington, DC, USA, 1990; Volume 1, pp. 260–267. [Google Scholar]
- Tesch, S.D. The Pacific Northwest Region. In Regional Silviculture of the United States; Barrett, J.W., Ed.; John Wiley and Sons Inc.: New York, NY, USA, 1995; pp. 499–558. [Google Scholar]
- Cordes, L.D. An Ecological Study of the Sitka Spruce Forest on the West Coast of Vancouver Island. Ph.D. Thesis, University of British Columbia, Vancouver, BC, Canada, 1972. [Google Scholar]
- Godman, R.M. Seed Dispersal in Southeast Alaska; USDA Forest Service: Juneau, AK, USA, 1953; p. 2.
- Harris, A.S. Natural Reforestation on a Mile-Square Clearcut in Southeast Alaska; USDA Forest Service, Pacific Northwest Forest and Range Experiment Station: Juneau, AK, USA, 1967; p. 16.
- Harris, A.S.; Farr, W.A. The Forest Ecosystem of Southeast Alaska; USDA Forest Service, Pacific Northwest Forest and Range Experiment Station: Portland, OR, USA, 1974; p. 109.
- Deal, R.L.; Oliver, C.D.; Bormann, B.T. Reconstruction of mixed hemlock-spruce stands in coastal southeast Alaska. Can. J. For. Res. 1991, 21, 643–654. [Google Scholar] [CrossRef]
- Orlikowska, E.H.; Deal, R.L.; Hennon, P.E.; Wipfli, M.S. The role of red alder in riparian forest structure along headwater streams in southeastern Alaska. Northwest Sci. 2004, 78, 111–123. [Google Scholar]
- Packee, E.C. Tsuga heterophyla (Raf.) Sarg. Western hemlock. In Silvics of North America; Burns, R.M., Honkala, B.H., Eds.; USDA Forest Service: Washington, DC, USA, 1990; Volume 1, pp. 613–622. [Google Scholar]
- Deal, R.L.; Farr, W.A. Composition and development of conifer regeneration in thinned and unthinned natural stands of western hemlock and Sitka spruce in southeast Alaska. Can. J. For. Res. 1994, 24, 976–984. [Google Scholar] [CrossRef]
- Minore, D. Comparative Autoecological Characteristics of Northwestern Tree Species—A Literature Review; USDA Forest Service, Pacific Northwest Forest and Range Experiment Station: Portland, OR, USA, 1979.
- Minore, D. Germination and Early Growth of Coastal Tree Species on Organic Seed Beds; USDA Forest Service, Pacific Northwest Forest and Range Experiment Station: Portland, OR, USA, 1972; p. 40.
- Harmon, M.E.; Franklin, J.F. Tree seedlings on logs in Picea-Tsuga forests of Oregon and Washington. Ecology 1989, 70, 48–59. [Google Scholar] [CrossRef]
- Christy, E.J.; Mack, R.N. Variation in demography of juvenile Tsuga heterophylla across the substratum mosaic. J. Ecol. 1984, 72, 75–91. [Google Scholar] [CrossRef]
- Levy, L.S.Y.L.; Deal, R.L.; Tappeiner, J.C. The Density and Distribution of Sitka Spruce and Western Hemlock Seedling Banks in Partially Harvested Stands in Southeast Alaska; USDA Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 2010; p. 20.
- Harrington, C.A. Alnus rubra Bong.-Red alder. In Silvics of North America; Burns, R.M., Honkala, B.H., Eds.; USDA Forest Service: Washington, DC, USA, 1990; Volume 2, pp. 116–123. [Google Scholar]
- Hulten, E. Flora of Alaska and Neighboring Territories; Stanford University Press: Stanford, CA, USA, 1968; p. 1008. [Google Scholar]
- Viereck, L.A.; Little, E.L. Alaska Trees and Shrubs; U.S. Department of Agriculture: Washington, DC, USA, 1972; Volume 410, p. 265.
- Wipfli, M.S.; Deal, R.L.; Hennon, P.E.; Johnson, A.C.; DeSanto, T.L.; Hanley, T.A.; Schultz, M.E.; Bryant, M.D.; Edwards, R.T.; Orlikowska, E.H.; et al. Managing Young Upland Forest in Southeast Alaska for Wood Products, Wildlife, Aquatic Resources, and Fishes: Problem Analysis and Study Plan; USDA Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 2002; p. 71.
- Hanley, T.A.; Deal, R.L.; Orlikowska, E.H. Relations between red alder (Alnus rubra Bong.) and understory vegetation in a gradient of mixed hardwood-conifer, young-growth forest. Can. J. For. Res. 2006, 36, 738–748. [Google Scholar] [CrossRef]
- Harrington, C.A.; Zasada, J.C.; Allen, E.A. Biology of Red Alder (Alnus rubra Bong.). In The Biology and Management of Red Alder; Hibbs, D.E., DeBell, D.S., Tarrant, R.F., Eds.; Oregon State University Press: Corvallis, OR, USA, 1994; pp. 3–22. [Google Scholar]
- Meier, E. The Wood Database: The Truth behind Wood Identification. 2008. Available online: http://www.wood-database.com/wood-articles/the-truth-behind-wood-identification/ (accessed on 12 September 2016).
- Ruth, R.H.; Harris, A.S. Management of Western Hemlock-Sitka spruce Forests for Timber Production; USDA Forest Service: Washington, DC, USA, 1979; p. 197.
- Farr, W.A. Site Index and Height Growth Curves for Unmanaged Even-Aged Stands of Western Hemlock and Sitka Spruce in Southeast Alaska; USDA Forest Service, Pacific Northwest Forest and Range Experiment Station: Portland, OR, USA, 1984; p. 26.
- Van Pelt, R. Forest Giants of the Pacific Coast, 1st ed.; University of Washington Press: Seattle, WA, USA, 2001; p. 200. [Google Scholar]
- Taylor, R.F. Yield of Second-Growth Western Hemlock-Sitka Spruce Stands in South-Eastern Alaska; USDA: Washington, DC, USA, 1934; p. 29.
- Harris, A.S.; Ruth, R.H. Sitka Spruce: A Bibliography with Abstracts; USDA Forest Service, Pacific Northwest Forest and Range Experiment Station: Portland, OR, USA, 1970; p. 251.
- Ruth, R.H. Sitka spruce (Picea sitchensis (Bong.) Carr.). In Silvics of Forest Trees of the United States; Folwells, H.A., Ed.; USDA: Washington, DC, USA, 1965; pp. 311–317. [Google Scholar]
- Griggs, R.F. The edge of the forest in Alaska and the reason for its position. Ecology 1934, 15, 80–96. [Google Scholar] [CrossRef]
- Tappeiner, J.C.; Maguire, D.A.; Harrington, T.B. Silviculture and Ecology of Western U.S. Forests, 1st ed.; Oregon State University Press: Corvallis, OR, USA, 2007; p. 440. [Google Scholar]
- Farr, W.A.; Harris, A.S. Site index of Sitka spruce along the Pacific coast related to latitude and temperatures. For. Sci. 1979, 25, 145–153. [Google Scholar]
- Fujimori, T. Primary Productivity of a Young Tsuga Heterophylla Stand and Some Speculations about Biomass of Forest Communities on the Oregon Coast; USDA Forest Service, Pacific Northwest Forest and Range Experiment Station: Portland, OR, USA, 1971; p. 11.
- Handley, D.L. The yield potential of western hemlock. In Western Hemlock Management; Institute of Forest Products Contribution No. 34; Atkinson, W.A., Zasoski, R.J., Eds.; University of Washington: Seattle, WA, USA, 1976; pp. 221–227. [Google Scholar]
- Hibbs, D.E.; DeBell, D.S.; Tarrant, R.F. The Biology and Management of Red Alder, 1st ed.; Hibbs, D.E., DeBell, D.S., Tarrant, R.F., Eds.; Oregon State University Press: Corvallis, OR, USA, 1994; p. 256. [Google Scholar]
- Harrington, C.A. A Method of Site Quality Evaluation for Red Alder; USDA Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1986; p. 22.
- Heebner, C.F.; Bergener, M.J. Red Alder: A Bibliography with Abstracts; USDA Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1983; p. 186.
- Eyre, F.H. Forest Cover Types of the United States and Canada; Society of American Foresters: Washington, DC, USA, 1980; p. 148. [Google Scholar]
- Newton, M.; Cole, E.C. Stand development and successional implications: Pure and mixed stands. In The Biology and Management of Red Alder, 1st ed.; Hibbs, D.E., DeBell, D.S., Tarrant, R.F., Eds.; Oregon State University Press: Corvallis, OR, USA, 1994; pp. 106–115. [Google Scholar]
- DeMars, D. Stand Density Study of Spruce-Hemlock Stands in Southeastern Alaska; USDA Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 2000; p. 60.
- Harrington, C.A.; Curtis, R.O. Height Growth and Site Index Curves for Red Alder; USDA Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1986; p. 14.
- Worthington, N.P.; Johnson, F.A.; Staebler, G.R.; Lloyd, W.J. Normal Yield Tables for Red Alder; USDA Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1960; p. 32.
- Green, R.N.; Klinka, K. A Field Guide to Site Identification and Interpretation for the Vancouver Forest Region; British Columbia Ministry of Forests: Victoria, BC, Canada, 1994; p. 285.
- Omule, S. Comparative Height Growth to Age 28 for Seven Species in the CWHd Subzone; British Columbia Ministry of Forests: Victoria, BC, Canada, 1987; p. 9.
- Williamson, R.L. Productivity of Red Alder in Western Oregon and Washington. In Biology of Alder; Trappe, J.M., Franklin, J.E., Tarrant, R.E., Hansen, G.M., Eds.; USDA Forest Service, Pacific Northwest Forest and Range Experiment Station: Portland, OR, USA, 1968; pp. 287–292. [Google Scholar]
- Hermann, R.K.; Lavender, D.P. Pseudotsuga menziesii (Mirb.) Franco—Douglas-fir. In Silvics of North America; Burns, R.M., Honkala, B.H., Eds.; USDA Forest Service: Washington, DC, USA, 1990; Volume 1, pp. 527–540. [Google Scholar]
- Deal, R.L. Red alder stand development and dynamics. In Red Alder: A State of Knowledge; Deal, R.L., Harrington, C.A., Eds.; USDA Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 2006; pp. 45–54. [Google Scholar]
- DeBell, D.S.; Strand, R.F.; Reukema, D.L. Short-rotation production of red alder: Some options for future forest management. In Utilization and Management of Alder; Briggs, D.G., DeBell, D.S., Atkinson, W.A., Eds.; USDA Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1978; pp. 231–244. [Google Scholar]
- Chambers, C.J. Empirical Yield Tables for Predominantly Alder Stands in Western Washington; Washington State Department of Natural Resources: Olympia, WA, USA, 1983; Volume 31, p. 70.
- Skinner, E.C. Cubic Volume Tables for Red Alder and Sitka Spruce; USDA Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1959; p. 4.
- Bormann, B.T.; Gordon, J.C. Stand Density Effects in Young Red Alder Plantations: Productivity, Photosynthate Partitioning, and Nitrogen Fixation. Ecology 1984, 65, 394–402. [Google Scholar] [CrossRef]
- DeBell, D.S.; Giordano, P.A. Growth patterns of red alder. In The Biology and Management of Red Alder, 1st ed.; Hibbs, D.E., DeBell, D.S., Tarrant, R.F., Eds.; Oregon State University Press: Corvallis, OR, USA, 1994; pp. 116–130. [Google Scholar]
- Knowe, S.A.; Hibbs, D.E. Stand structure and dynamics of young red alder as affected by planting density. For. Ecol. Manag. 1996, 82, 69–85. [Google Scholar] [CrossRef]
- Puettmann, K.J. Growth and yield of red alder. In The Biology and Management of Red Alder, 1st ed.; Hibbs, D.E., DeBell, D.S., Tarrant, R.F., Eds.; Oregon State University Press: Corvallis, OR, USA, 1994; pp. 229–242. [Google Scholar]
- Knowe, S.A.; Ahrens, G.R.; DeBell, D.S. Comparison of diameter-distribution-prediction, stand-table-projection, and individual-tree-growth modeling approaches for young red alder plantations. For. Ecol. Manag. 1997, 98, 49–60. [Google Scholar] [CrossRef]
- Smith, J.H.G. Growth and yield of red alder in British Columbia. In Biology of Alder; Trappe, J.M., Franklin, J.F., Tarrant, R.F., Hansen, G.M., Eds.; USDA Forest Service, Pacific Northwest Range and Experiment Station: Portland, OR, USA, 1968; pp. 273–286. [Google Scholar]
- Gomi, T.; Johnson, A.C.; Deal, R.L.; Hennon, P.E.; Orlikowska, E.H.; Wipfli, M.S. Factors affecting distribution of wood, detritus, and sediment in headwater streams draining managed young-growth red alder-conifer forests in southeast Alaska. Can. J. For. Res. 2006, 36, 725–737. [Google Scholar] [CrossRef]
- Deal, R.L.; Hennon, P.E.; Orlikowska, E.H.; D’Amore, D.V. Stand dynamics of mixed red alder-conifer forests of southeast Alaska. Can. J. For. Res. 2004, 34, 969–980. [Google Scholar] [CrossRef]
- Kimbirauskas, R.K.; Merritt, R.W.; Wipfli, M.S.; Hennon, P.E. Headwater macroinvertebrate community response to riparian red alder (Alnus rubra Bong.) in southeast Alaska. Pan Pac. Entomol. 2008, 84, 220–237. [Google Scholar] [CrossRef]
- Wipfli, M.S.; Deal, R.L.; Hennon, P.E.; Johnson, A.C.; Edwards, R.T.; De Santo, T.L.; Gomi, T.; Orlikowska, E.H.; Bryant, M.D.; Schultz, M.E.; et al. Compatible management of red alder-conifer ecosystems in southeastern Alaska. In Compatible Forest Management, 1st ed.; Monserud, R.A., Haynes, R.W., Johnson, A.C., Eds.; Springer: Dordrecht, The Netherlands, 2003; pp. 55–81. [Google Scholar]
- Schultz, M.E.; De Santo, T.L. Comparison of terrestrial invertebrate biomass and richness in young mixed red alder-conifer, young conifer, and old conifer stands of southeast Alaska. Northwest Sci. 2006, 80, 120–132. [Google Scholar]
- Bryant, M.D.; Zymonas, N.D.; Wright, B.E. Salmonids on the Fringe: Abundance, Species Composition, and Habitat Use of Salmonids in High-Gradient Headwater Streams, Southeast Alaska. Trans. Am. Fish. Soc. 2004, 133, 1529–1538. [Google Scholar] [CrossRef]
- Hibbs, D.E.; Bower, A.L. Riparian forests in the Oregon Coast Range. For. Ecol. Manag. 2001, 154, 201–213. [Google Scholar] [CrossRef]
- Alaback, P.B. Plant Succession Following Logging in the Sitka Spruce-Western Hemlock Forests of Southeast Alaska: Implications for Management; USDA Forest Service, Pacific Northwest Forest and Range Experiment Station: Portland, OR, USA, 1984; p. 26.
- Miller, R.E.; Murray, M.D. The Effect of Red Alder on Growth of Douglas-Fir. In Utilization and Management of Alder; Briggs, D.G., DeBell, D.S., Atkinson, W.A., Eds.; USDA Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1978; pp. 283–306. [Google Scholar]
- Bormann, B.T.; Cromack, K.; Russell, W.O. Influences of Red Alder on Soils and Long-Term Ecosystem Productivity. In The Biology and Management of Red Alder, 1st ed.; Hibbs, D.E., DeBell, D.S., Tarrant, R.F., Eds.; Oregon State University Press: Corvallis, OR, USA, 1994; pp. 47–56. [Google Scholar]
- Bryant, M.D. Changes thirty years after logging in large woody debris, and its use by salmonids. In Riparian Ecosystems and Their Management: Reconciling Conflicting Uses; Johnson, R.R., Ziebell, C.D., Patton, D.R., Folliott, P.F., Hamre, R.H., Eds.; USDA Forest Service, Rocky Mountain, Forest and Range Experiment Station: Fort Collins, CO, USA, 1985; pp. 329–334. [Google Scholar]
- Bisson, P.A.; Bilby, R.E.; Bryant, M.D.; Dollof, C.A.; Grette, G.B.; House, R.A.; Murphy, M.L.; Koski, K.V.; Sedell, J.R. Large Woody Debris in the Pacific Northwest: Past, Present, and Future. In Streamside Management: Forestry and Fishery Interactions; Salo, E.O., Cundy, T.W., Eds.; Institute of Forest Resources, University of Washington: Seattle, WA, USA, 1987; pp. 143–190. [Google Scholar]
- Minore, D.; Weatherly, H.G. Riparian trees, shrubs and forest regeneration in the coastal mountains of Oregon. New For. 1994, 8, 249–263. [Google Scholar] [CrossRef]
- Nierenberg, T.R.; Hibbs, D.E. A characterization of unmanaged riparian areas in the central Coast Range of western Oregon. For. Ecol. Manag. 2000, 129, 195–206. [Google Scholar] [CrossRef]
- Pabst, R.J.; Spies, T.A. Structure and composition of unmanaged riparian forests in the coastal mountains of Oregon, USA. Can. J. For. Res. 1999, 29, 1557–1573. [Google Scholar] [CrossRef]
- Hennon, P.E.; McClellan, M.H. Tree mortality and forest structure in the temperate rain forests of southeast Alaska. Can. J. For. Res. 2003, 33, 1621–1634. [Google Scholar] [CrossRef]
- Kessler, W.B.; Kogut, T.E. Habitat orientations of forest birds in southeastern Alaska. Northwest Sci. 1985, 59, 58–65. [Google Scholar]
- Schwab, F.E. Effect of Vegetation Structure on Breeding Bird Communities in the Dry Zone Douglas-Fir Forests of Southeastern British Columbia. Master’s Thesis, University of British Columbia, Vancouver, BC, Canada, 1979. [Google Scholar]
- Huff, M.H.; Raley, C.M. Regional Patterns of Diurnal Breeding Bird Communities in Oregon and Washington. In Wildlife and Vegetation of Unmanaged Douglas-Fir Forests; Ruggiero, L.F., Aubry, K.B., Carey, A.B., Huff, M.H., Eds.; USDA Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1991; pp. 177–206. [Google Scholar]
- Gilbert, F.F.; Allwine, R. Spring bird communities in the Oregon Cascade Range. In Wildlife and Vegetation of Unmanaged Douglas-Fir Forests; Ruggiero, L.F., Aubry, K.B., Carey, A.B., Huff, M.H., Eds.; USDA Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1991; pp. 145–160. [Google Scholar]
- McComb, W.C. Red alder: Interactions with wildlife. In The Biology and Management of Red Alder, 1st ed.; Hibbs, D.E., DeBell, D.S., Tarrant, R.F., Eds.; Oregon State University Press: Corvallis, OR, USA, 1994; pp. 131–138. [Google Scholar]
- Kelly, J.P. The effect of nest predation on habitat selection by dusky flycatchers in limber pine-juniper woodland. Condor 1993, 95, 83–93. [Google Scholar] [CrossRef]
- Martin, T.E. Nest predation among vegetation layers and habitat types: Revising the dogmas. Am. Nat. 1993, 141, 898–913. [Google Scholar] [CrossRef] [PubMed]
- Easton, W.E.; Martin, K. Effects of thinning and herbicide treatments on nest-site selection by songbirds in young managed forests. Auk 2002, 119, 685–694. [Google Scholar] [CrossRef]
- Airola, D.A. Foraging and Habitat Relations of Insectivorous Birds in a Managed Sierra Nevada Mixed Conifer Forest. Master’s Thesis, University of California, Berkeley, CA, USA, 1979. [Google Scholar]
- Holmes, R.T. The structure of a temperate deciduous forest bird community: Variability in time and space. In Biogeography and Ecology of Forest Bird Communities, 1st ed.; Keast, A., Ed.; SPB Publishing: Hague, The Netherlands, 1990; pp. 121–139. [Google Scholar]
- Stiles, E.W. Bird community structure in alder forests in Washington. Condor 1980, 82, 20–30. [Google Scholar] [CrossRef]
- Willson, M.F.; Comet, T.A. Bird communities of northern forests: Patterns of diversity and abundance. Condor 1996, 98, 337–349. [Google Scholar] [CrossRef]
- Wipfli, M.S.; Musslewhite, J. Density of red alder (Alnus rubra) in headwaters influences invertebrate and detritus subsidies to downstream fish habitats in Alaska. Hydrobiologia 2004, 520, 153–163. [Google Scholar] [CrossRef]
- Cummins, K.W.; Wilzbach, M.A.; Gates, D.M.; Taliaferri, J.B.; Perry, W.B. Shredders and riparian vegetation: Leaf litter that falls into streams influences communities of stream invertebrates. BioScience 1989, 39, 24–30. [Google Scholar] [CrossRef]
- Wipfli, M.S. Terrestrial invertebrates as salmonid prey and nitrogen sources in streams: Contrasting old-growth and young-growth riparian forests in southeastern Alaska, U.S.A. Can. J. Fish. Aquat. Sci. 1997, 54, 1259–1269. [Google Scholar] [CrossRef]
- Mason, C.F.; Macdonald, S.M. The input of terrestrial invertebrates from tree canopies to a stream. Freshw. Biol. 1982, 12, 305–311. [Google Scholar] [CrossRef]
- Piccolo, J.J.; Wipfli, M.S. Does red alder (Alnus rubra) in upland riparian forests elevate macroinvertebrate and detritus export from headwater streams to downstream habitats in southeastern Alaska? Can. J. Fish. Aquat. Sci. 2002, 59, 503–513. [Google Scholar] [CrossRef]
- Allan, J.D.; Wipfli, M.S.; Caouette, J.P.; Prussian, A.; Rodgers, J. Influence of streamside vegetation on terrestrial invertebrate subsidies to salmonid food webs. Can. J. Fish. Aquat. Sci. 2003, 60, 309–320. [Google Scholar] [CrossRef]
- Luken, J.O.; Fonda, R.W. Nitrogen accumulation in a chronosequence of red alder communities along the Hoh River, Olympic National Park, Washington. Can. J. For. Res. 1983, 13, 1228–1237. [Google Scholar] [CrossRef]
- Fevold, K. Sub-Surface Controls on the Distribution of Benthic Algae in Floodplain Back Channel Habitats of the Queets River. Master’s Thesis, University of Washington, Seattle, WA, USA, 1998. [Google Scholar]
- Benda, L.E.; Dunne, T. Stochastic forcing of sediment routing and storage in channel networks. Water Resour. Res. 1997, 33, 2865–2880. [Google Scholar] [CrossRef]
- Gomi, T.; Sidle, R.C.; Richardson, J.S. Understanding processes and downstream linkages of headwater systems. BioScience 2002, 52, 905–916. [Google Scholar] [CrossRef]
- Gregory, S.V.; Swanson, F.J.; McKee, W.A.; Cummins, K.W. An ecosystem perspective of riparian zones. BioScience 1991, 41, 540–551. [Google Scholar] [CrossRef]
- Swanson, F.J.; Johnson, S.L.; Gregory, S.V.; Acker, S.A. Flood disturbance in a forested mountain landscape. BioScience 1998, 48, 681–689. [Google Scholar] [CrossRef]
- Wipfli, M.S.; Gregovich, D.P. Export of invertebrates and detritus from fishless headwater streams in southeastern Alaska: Implications for downstream salmonid production. Freshw. Biol. 2002, 47, 957–969. [Google Scholar] [CrossRef]
- Hanley, T.A.; Barnard, J.C. Red alder, Alnus rubra, as a potential mitigating factor for wildlife habitat following clearcut logging in southeastern Alaska. Can. Field Nat. 1998, 112, 647–652. [Google Scholar]
- Newton, M.; El Hassan, B.A.; Zavitkovski, J. Role of red alder in western Oregon forest succession. In Biology of Alder; Trappe, J.M., Franklin, J.E., Tarrant, R.E., Hansen, G.M., Eds.; USDA Forest Service, Pacific Northwest Forest and Range Experiment Station: Portland, OR, USA, 1968; pp. 73–84. [Google Scholar]
- Deal, R.L.; Russell, J.M. The potential role of red alder for restoring forest ecosystems in even-aged western hemlock-Sitka spruce stands of southeast Alaska. In Integrated Restoration of Forested Ecosystems to Achieve Multi-Resource Benefits: Proceedings of the 2007 National Silviculture Workshop; Deal, R.L., Ed.; USDA Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 2008; pp. 135–145. [Google Scholar]
- Deal, R.L.; Tappeiner, J.C. The effects of partial cutting on stand structure and growth of western hemlock–Sitka spruce stands in southeast Alaska. For. Ecol. Manag. 2002, 159, 173–186. [Google Scholar] [CrossRef]
- Newton, M.E.; Cole, E.C.; White, D.E. Tall planting stock for enhanced growth and domination of brush in the Douglas-fir region. New For. 1993, 7, 107–121. [Google Scholar] [CrossRef]
- O’Hanlon, R.; Harrington, T.J.; Outerbridge, R.A.; Berch, S.M. High species richness but different communities of macrofungi in Sitka spruce (Picea sitchensis [Bong. Carr.]) forests in its native (British Columbia) and non-native (Ireland and Britain) range. Can. J. For. Res. 2013, 43, 450–458. [Google Scholar] [CrossRef]
- Alaback, P.B. Dynamics of Understory Biomass in Sitka Spruce-Western Hemlock Forests of Southeast Alaska. Ecology 1982, 63, 1932–1948. [Google Scholar] [CrossRef]
- Tappeiner, J.C., II; Alaback, P.B. Early establishment and vegetative growth of understory species in the western hemlock—Sitka spruce forests in southeast Alaska. Can. J. Bot. 1989, 67, 318–326. [Google Scholar] [CrossRef]
- Doerr, J.G.; Sandburg, N.H. Effects of precommercial thinning on understory vegetation and deer habitat utilization on Big Level Island in Southeast Alaska. For. Sci. 1986, 32, 1092–1095. [Google Scholar]
- Parker, K.L.; Gillingham, M.P.; Hanley, T.A.; Robbins, C.T. Energy and protein balance of free-ranging black-tailed deer in a natural forest environment. Wildl. Monogr. 1999, 143, 1–48. [Google Scholar]
- Hanley, T.A.; McKendrick, J.D. Seasonal Changes in Chemical Composition and Nutritive Value of Native Forages in a Spruce-Hemlock Forest, Southeastern Alaska; USDA Forest Service, Pacific Northwest Forest and Range Experiment Station: Portland, OR, USA, 1983; p. 48.
- Hanley, T.A.; Barnard, J.C. Food resources and diet composition in riparian and upland habitats for Sitka mice, Peromyscus keeni sitkensis. Can. Field Nat. 1999, 113, 401–407. [Google Scholar]
- Franklin, J.F. Scientific Basis for New Perspectives in Forests and Streams. In Watershed Management; Naiman, R., Ed.; Springer: New York, NY, USA, 1992; pp. 25–72. [Google Scholar]
- Hernandez, O.; Merritt, R.W.; Wipfli, M.S. Benthic invertebrate community structure is influenced by forest succession after clearcut logging in southeastern Alaska. Hydrobiologia 2005, 533, 45–59. [Google Scholar] [CrossRef]
- Hanley, T.A. Small mammals of even-aged, red alder-conifer forests in southeastern Alaska. Can. Field Nat. 1996, 110, 626–629. [Google Scholar]
- Deal, R.L. Understory Plant Diversity in Riparian Alder-Conifer Stands after Logging in Southeast Alaska; US Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1997; p. 8.
- Pechanec, A.A.; Franklin, J.F. Comparison of vegetation in adjacent alder, conifer, and mixed alder-conifer communities. II. Epiphytic, epixylic, and epilithic cryptogam. In Biology of Alder; Trappe, J.M., Franklin, J.F., Tarrant, R.F., Hansen, G.M., Eds.; USDA Forest Service, Pacific Northwest Forest and Range Experiment Station: Portland, OR, USA, 1968; pp. 85–98. [Google Scholar]
- Beechie, T.J.; Pess, G.; Kennard, P.; Bilby, R.E.; Bolton, S. Modeling Recovery Rates and Pathways for Woody Debris Recruitment in Northwestern Washington Streams. N. Am. J. Fish. Manag. 2000, 20, 436–452. [Google Scholar] [CrossRef]
- Tarrant, R.F.; Bormann, B.T.; DeBell, D.S.; Atkinson, W.A. Managing red alder in the Douglas-fir region: Some possibilities. J. For. 1983, 81, 787–792. [Google Scholar]
- Murai, S. Phytotaxomomical and Geobotanical Studies on Genus Alnus in Japan. III. Taxonomy of Whole World Species and Distribution of Each Sect; Government Forest Experiment Station Bulletin: Tokyo, Japan, 1964; Volume 171, pp. 1–107. [Google Scholar]
- Fahlvik, N.; Agestam, E.; Ekö, P.M.; Linden, M. Development of single-storied mixtures of Norway spruce and birch in Southern Sweden. Scan. J. For. Res. 2011, 26, 36–45. [Google Scholar] [CrossRef]
- Webster, C.R.; Lorimer, C.G. Single-tree versus group selection in hemlock-hardwood forests: Are smaller openings less productive? Can. J. For. Res. 2002, 32, 591–604. [Google Scholar] [CrossRef]
- Boncina, A.; Gaspersic, F.; Diaci, J. Long-term changes in tree species composition in the Dinaric mountain forests of Slovenia. For. Chron. 2003, 79, 227–232. [Google Scholar] [CrossRef]

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deal, R.L.; Orlikowska, E.H.; D’Amore, D.V.; Hennon, P.E. Red Alder-Conifer Stands in Alaska: An Example of Mixed Species Management to Enhance Structural and Biological Complexity. Forests 2017, 8, 131. https://doi.org/10.3390/f8040131
Deal RL, Orlikowska EH, D’Amore DV, Hennon PE. Red Alder-Conifer Stands in Alaska: An Example of Mixed Species Management to Enhance Structural and Biological Complexity. Forests. 2017; 8(4):131. https://doi.org/10.3390/f8040131
Chicago/Turabian StyleDeal, Robert L., Ewa H. Orlikowska, David V. D’Amore, and Paul E. Hennon. 2017. "Red Alder-Conifer Stands in Alaska: An Example of Mixed Species Management to Enhance Structural and Biological Complexity" Forests 8, no. 4: 131. https://doi.org/10.3390/f8040131
APA StyleDeal, R. L., Orlikowska, E. H., D’Amore, D. V., & Hennon, P. E. (2017). Red Alder-Conifer Stands in Alaska: An Example of Mixed Species Management to Enhance Structural and Biological Complexity. Forests, 8(4), 131. https://doi.org/10.3390/f8040131