
Look Down to See What’s Up: A Systematic Overview of Treefall Dynamics in Forests



Abstract
:1. Introduction
2. Methodology
3. Treefall Literature: Current Knowledge
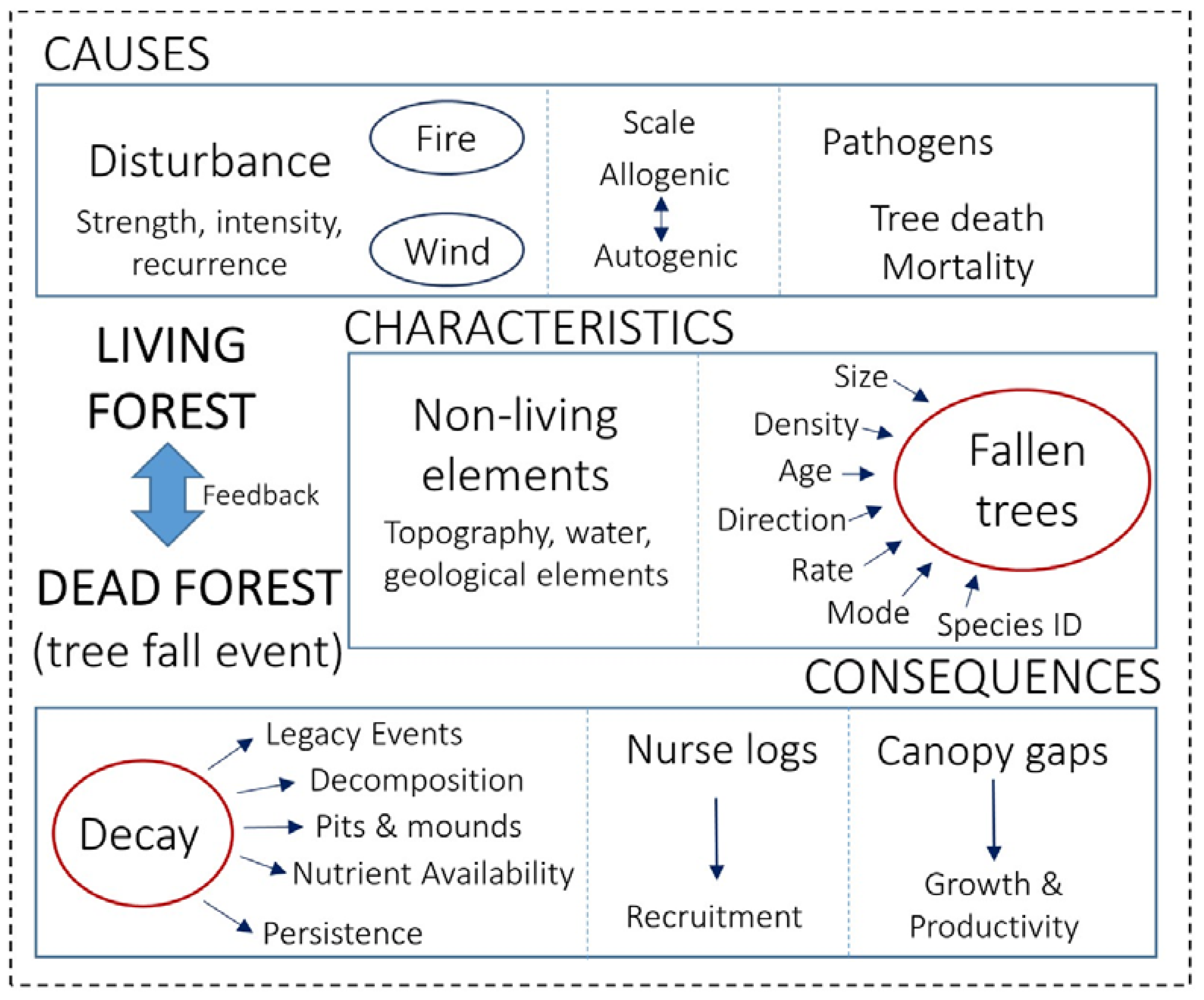
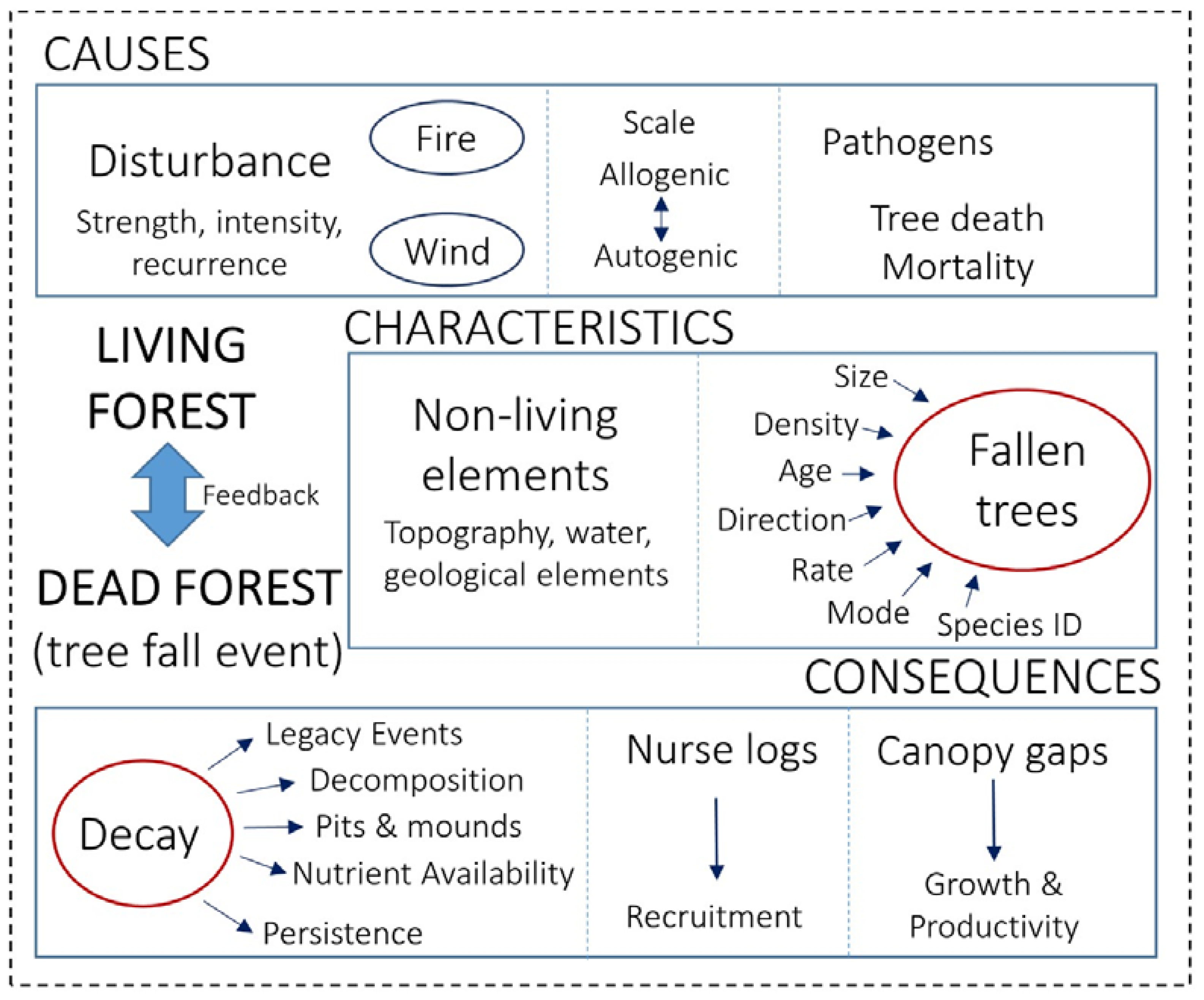
3.1. Causes
3.2. Characteristics
3.3. Consequences
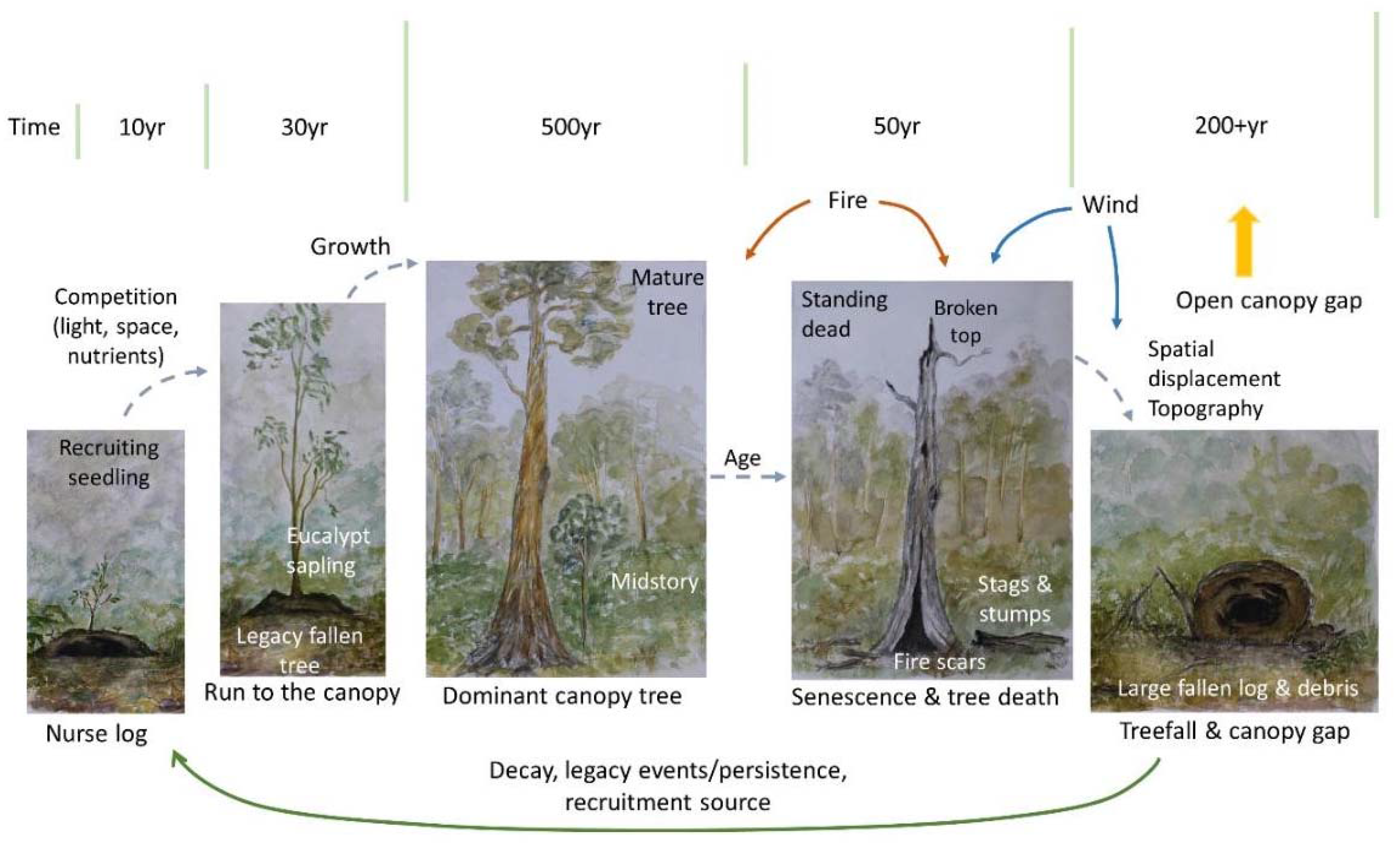
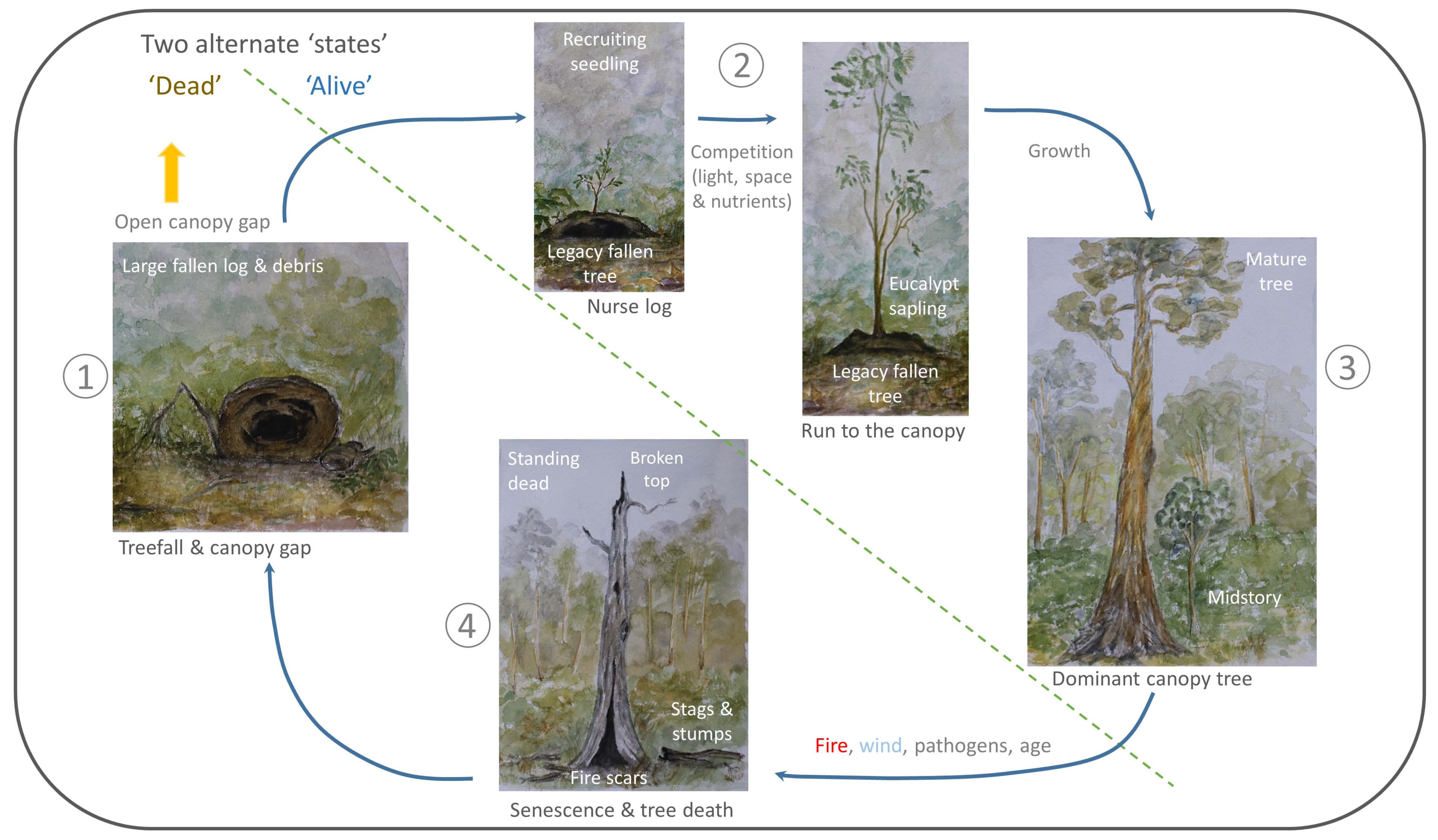
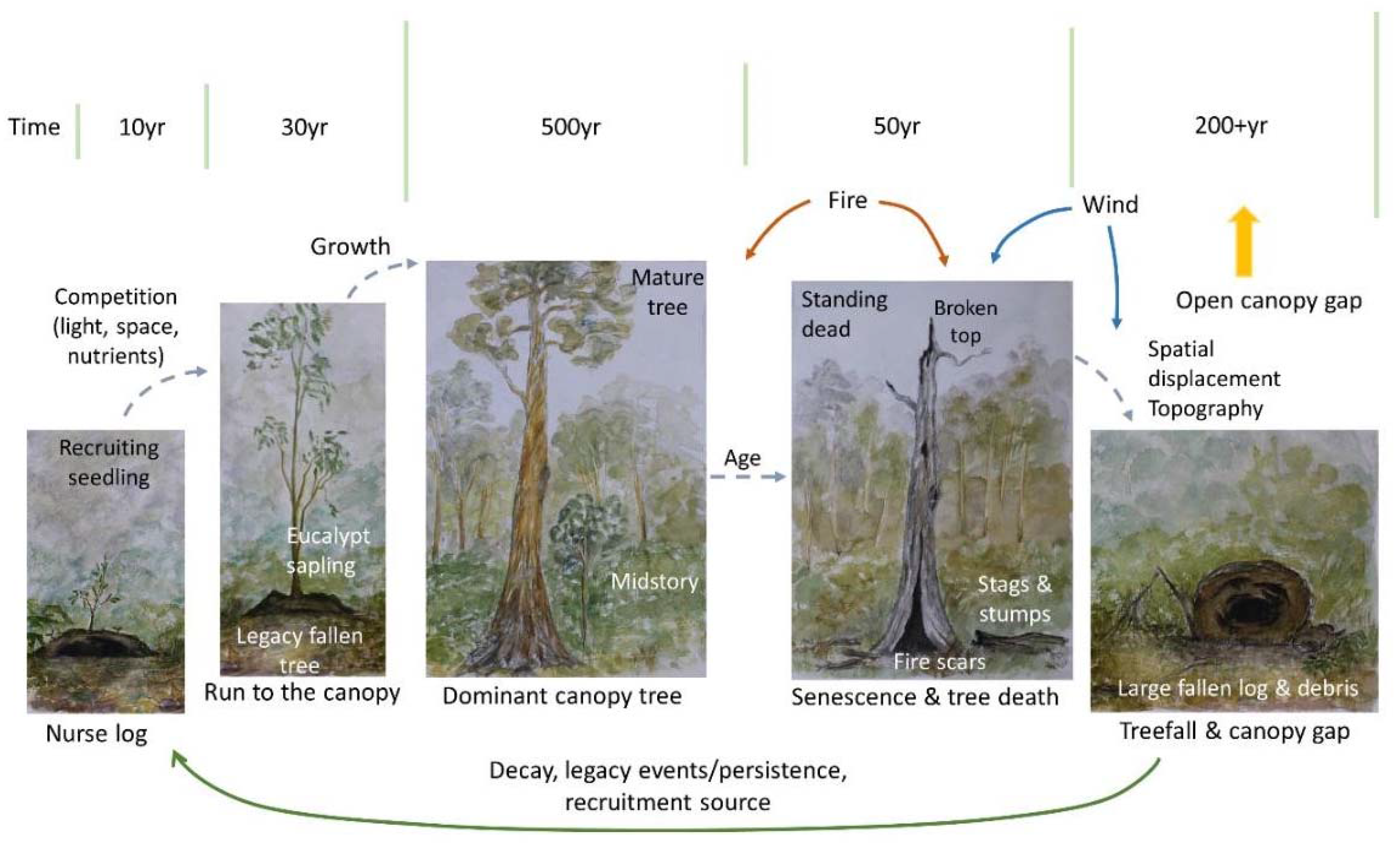
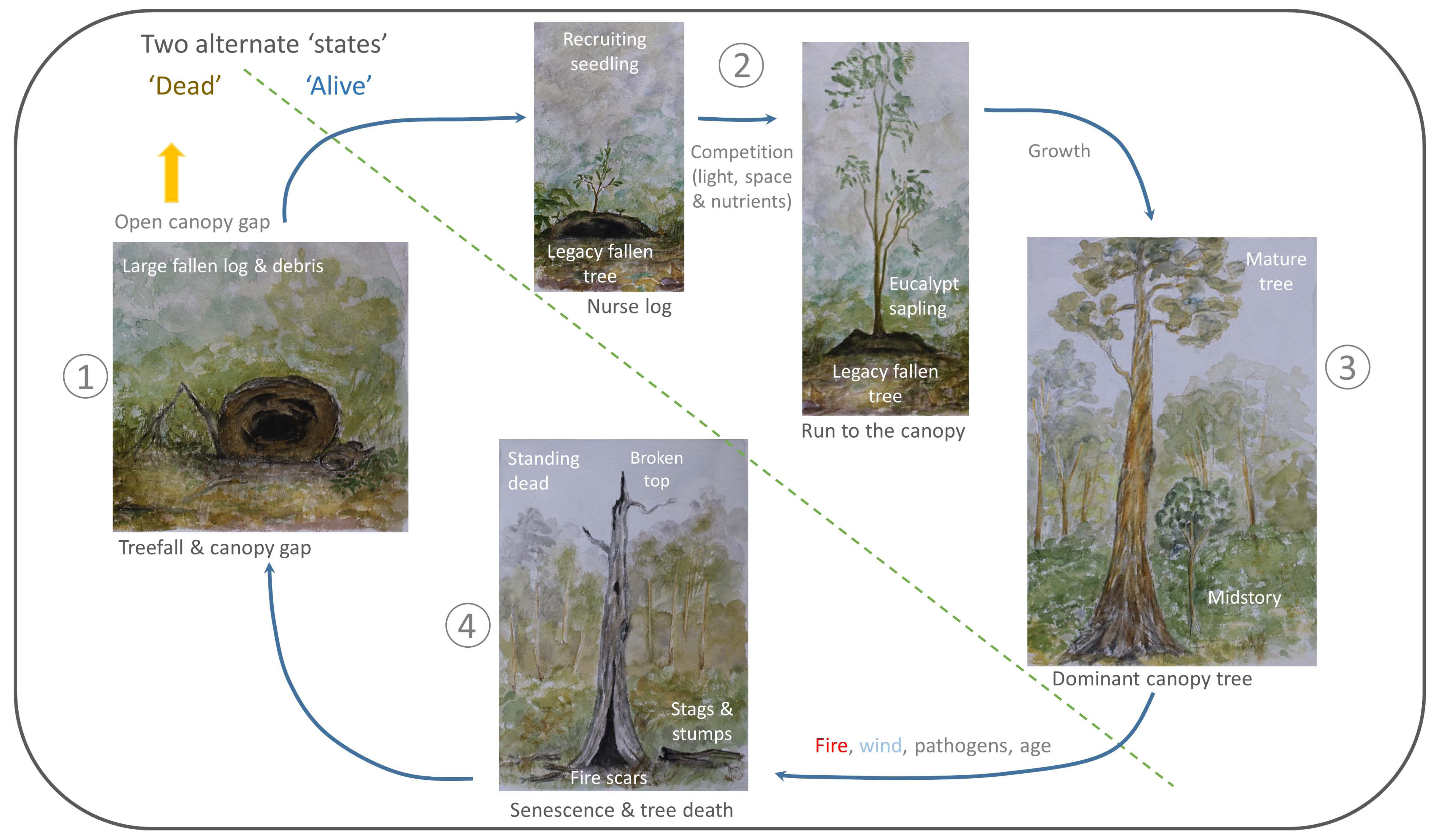
4. Living-Forest Dynamics
5. A Treefall’s Eye-View of a FOREST—What Is Next?
6. Future Directions
7. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Siitonen, J. Forest management, coarse woody debris and saproxylic organisms: Fennoscandian boreal forests as an example. Ecol. Bull. 2001, 49, 11–41. [Google Scholar]
- Fridman, J.; Walheim, M. Amount, structure, and dynamics of dead wood on managed forestland in Sweden. For. Ecol. Manag. 2000, 131, 23–36. [Google Scholar] [CrossRef]
- Müller, J.; Bütler, R. A review of habitat thresholds for dead wood: A baseline for management recommendations in European forests. Eur. J. For. Res. 2010, 129, 981–992. [Google Scholar] [CrossRef]
- Ganey, J.L.; Bird, B.J.; Baggett, L.S.; Jenness, J.S. Density of large snags and logs in northern Arizona mixed-conifer and ponderosa pine forests. For. Sci. 2015, 61, 353–362. [Google Scholar] [CrossRef]
- Rogers, P. Disturbance Ecology and for. Management: A Review of the Literature; US Department of Agriculture, Forest Service, Intermountain Research Station: Ogden, UT, USA, 1996.
- Stachowicz, J.J. Mutualism, facilitation, and the structure of ecological communities: Positive interactions play a critical, but underappreciated, role in ecological communities by reducing physical or biotic stresses in existing habitats and by creating new habitats on which many species depend. BioScience 2001, 51, 235–246. [Google Scholar]
- Brooker, R.W.; Maestre, F.T.; Callaway, R.M.; Lortie, C.L.; Cavieres, L.A.; Kunstler, G.; Liancourt, P.; Tielbörger, K.; Travis, J.M.; Anthelme, F. Facilitation in plant communities: The past, the present, and the future. J. Ecol. 2008, 96, 18–34. [Google Scholar] [CrossRef]
- Flower, C.E.; Gonzalez-Meler, M.A. Responses of temperate forest productivity to insect and pathogen disturbances. Annu. Rev. Plant Biol. 2015, 66, 547–569. [Google Scholar] [CrossRef] [PubMed]
- Orwin, K.H.; Wardle, D.A.; Greenfield, L.G. Context-dependent changes in the resistance and resilience of soil microbes to an experimental disturbance for three primary plant chronosequences. Oikos 2006, 112, 196–208. [Google Scholar] [CrossRef]
- Filotas, E.; Parrott, L.; Burton, P.J.; Chazdon, R.L.; Coates, K.D.; Coll, L.; Haeussler, S.; Martin, K.; Nocentini, S.; Puettmann, K.J. Viewing forests through the lens of complex systems science. Ecosphere 2014, 5, 1–23. [Google Scholar] [CrossRef]
- McIntire, E.J.B.; Fajardo, A. Beyond description: The active and effective way to infer processes from spatial patterns. Ecology 2009, 90, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Franklin, J.F.; Shugart, H.H.; Harmon, M.E. Tree death as an ecological process. BioScience 1987, 37, 550–556. [Google Scholar] [CrossRef]
- Ulanova, N.G. The effects of windthrow on forests at different spatial scales: A review. For. Ecol. Manag. 2000, 135, 155–167. [Google Scholar] [CrossRef]
- Siitonen, J.; Martikainen, P.; Punttila, P.; Rauh, J. Coarse woody debris and stand characteristics in mature managed and old-growth boreal mesic forests in southern Finland. For. Ecol. Manag. 2000, 128, 211–225. [Google Scholar] [CrossRef]
- You, H.; He, D.; You, W.; Xiao, S.; Hong, W. Spatial distribution pattern of coarse woody debris (cwd) in two typical forest types in Tianbaoyan National Nature Reserve. Proceedings of 2012 World Automation Congress (Wac), Puerto Vallarta, Mexico, 24–28 June 2012; pp. 1–8. [Google Scholar]
- Soderberg, U.; Wulff, S.; Stahl, G. The choice of definition has a large effect on reported quantities of dead wood in boreal forest. Scand. J. For. Res. 2014, 29, 252–258. [Google Scholar] [CrossRef]
- Guby, N.A.B.; Dobbertin, M. Quantitative estimates of coarse woody debris and standing dead trees in selected Swiss forests. Glob. Ecol. Biogeogr. Lett. 1996, 5, 327–341. [Google Scholar] [CrossRef]
- Brokaw, N.V.L. Gap-phase regeneration in a tropical forest. Ecology 1985, 66, 682–687. [Google Scholar] [CrossRef]
- Watt, A.S. Pattern and process in the plant community. J. Ecol. 1947, 35, 1–22. [Google Scholar] [CrossRef]
- Uhl, C.; Clark, K.; Dezzeo, N.; Maquirino, P. Vegetation dynamics in Amazonian treefall gaps. Ecology 1988, 69, 751–763. [Google Scholar] [CrossRef]
- Muscolo, A.; Bagnato, S.; Sidari, M.; Mercurio, R. A review of the roles of forest canopy gaps. J. For. Res. 2014, 25, 725–736. [Google Scholar] [CrossRef]
- Zhu, J.; Lu, D.; Zhang, W. Effects of gaps on regeneration of woody plants: A meta-analysis. J. For. Res. 2014, 25, 501–510. [Google Scholar] [CrossRef]
- Canham, C.D.; Loucks, O.L. Catastrophic windthrow in the presettlement forests of Wisconsin. Ecology 1984, 65, 803–809. [Google Scholar] [CrossRef]
- Attiwill, P.M. The disturbance of forest ecosystems: The ecological basis for conservative management. For. Ecol. Manag. 1994, 63, 247–300. [Google Scholar] [CrossRef]
- Šamonil, P.; Král, K.; Hort, L. The role of tree uprooting in soil formation: A critical literature review. Geoderma 2010, 157, 65–79. [Google Scholar] [CrossRef]
- Bassett, M.; Chia, E.K.; Leonard, S.W.J.; Nimmo, D.G.; Holland, G.J.; Ritchie, E.G.; Clarke, M.F.; Bennett, A.F. The effects of topographic variation and the fire regime on coarse woody debris: Insights from a large wildfire. For. Ecol. Manag. 2015, 340, 126–134. [Google Scholar] [CrossRef]
- Lugo, A.E.; Scatena, F.N. Background and catastrophic tree mortality in tropical moist, wet, and rain forests. Biotropica 1996, 28, 585–599. [Google Scholar] [CrossRef]
- Sollins, P. Input and decay of coarse woody debris in coniferous stands in western Oregon and Washington. Can. J. For. Res. 1982, 12, 18–28. [Google Scholar] [CrossRef]
- Weedon, J.T.; Cornwell, W.K.; Cornelissen, J.H.; Zanne, A.E.; Wirth, C.; Coomes, D.A. Global meta-analysis of wood decomposition rates: A role for trait variation among tree species? Ecol. Lett. 2009, 12, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Cousins, S.J.M.; Battles, J.J.; Sanders, J.E.; York, R.A. Decay patterns and carbon density of standing dead trees in California mixed conifer forests. For. Ecol. Manag. 2015, 353, 136–147. [Google Scholar] [CrossRef]
- Maser, C. The Seen and Unseen World of the Fallen Tree; General Technical Report; USDA Forest Service, Pacific Northwest Forest and Range Experiment Station: Portland, OR, USA, 1984; p. 56.
- Harmon, M.E.; Franklin, J.F.; Swanson, F.J.; Sollins, P.; Gregory, S.; Lattin, J.; Anderson, N.; Cline, S.; Aumen, N.; Sedell, J. Ecology of coarse woody debris in temperate ecosystems. Adv. Ecol. Res. 1986, 15, 302. [Google Scholar]
- Woldendorp, G.; Keenan, R.J. Coarse woody debris in Australian forest ecosystems: A review. Aust. Ecol. 2005, 30, 834–843. [Google Scholar] [CrossRef]
- Oberle, B.; Milo, A.; Myers, J.A.; Young, D.F.; Walton, M.L.; Zanne, A.E. Direct estimates of downslope deadwood movement over 30 years in a temperature forest illustrate impacts of treefall on forest ecosystem dynamics. Cana. J. For. Res. 2015. [CrossRef]
- Lorimer, C.G. Methodological considerations in the analysis of forest disturbance history. Can. J. For. Res. 1985, 15, 200–213. [Google Scholar] [CrossRef]
- Schliemann, S.A.; Bockheim, J.G. Methods for studying treefall gaps: A review. For. Ecol. Manag. 2011, 261, 1143–1151. [Google Scholar] [CrossRef]
- Fisher, B.L.; Howe, H.F.; Wright, S.J. Survival and growth of Virola surinamensis yearlings—Water augmentation in gap and understory. Oecologia 1991, 86, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Christensen, M.; Hahn, K.; Mountford, E.P.; Ódor, P.; Standovár, T.; Rozenbergar, D.; Diaci, J.; Wijdeven, S.; Meyer, P.; Winter, S.; et al. Dead wood in European beech (Fagus sylvatica) forest reserves. For. Ecol. Manag. 2005, 210, 267–282. [Google Scholar] [CrossRef]
- Jonsson, B.G.; Dynesius, M. Uprooting in boreal spruce forests: Long-term variation in disturbance rate. Can. J. For. Res. 1993, 23, 2383–2388. [Google Scholar] [CrossRef]
- Everham, E.M.; Brokaw, N.V.L. Forest damage and recovery from catastrophic wind. Bot. Rev. 1996, 62, 113–185. [Google Scholar] [CrossRef]
- Rich, R.L.; Frelich, L.E.; Reich, P.B. Wind-throw mortality in the southern boreal forest: Effects of species, diameter and stand age. J. Ecol. 2007, 95, 1261–1273. [Google Scholar] [CrossRef]
- Peterson, C.J.; Pickett, S.T.A. Treefall and resprouting following catastrophic windthrow in an old-growth hemlock-hardwoods forest. For. Ecol. Manag. 1991, 42, 205–217. [Google Scholar] [CrossRef]
- Peterson, C.J.; Carson, W.P.; McCarthy, B.C.; Pickett, S.T.A. Microsite variation and soil dynamics within newly created treefall pits and mounds. Oikos 1990, 58, 39–46. [Google Scholar] [CrossRef]
- Sobhani, V.M.; Barrett, M.; Peterson, C.J. Robust prediction of treefall pit and mound sizes from tree size across 10 forest blowdowns in eastern north America. Ecosystems 2014, 17, 837–850. [Google Scholar] [CrossRef]
- Zanne, A.E.; Oberle, B.; Dunham, K.M.; Milo, A.M.; Walton, M.L.; Young, D.F. A deteriorating state of affairs: How endogenous and exogenous factors determine plant decay rates. J. Ecol. 2015, 103, 1421–1431. [Google Scholar] [CrossRef]
- Canham, C.D.; Denslow, J.S.; Platt, W.J.; Runkle, J.R.; Spies, T.A.; White, P.S. Light regimes beneath closed canopies and tree-fall gaps in temperate and tropical forests. Can. J. For. Res. 1990, 20, 620–631. [Google Scholar] [CrossRef]
- Whitmore, T. Canopy gaps and the two major groups of forest trees. Ecology 1989, 70, 536–538. [Google Scholar] [CrossRef]
- Arevalo, J.R.; Fernandez-Palacios, J.M. Treefall gap characteristics and regeneration in the laurel forest of Tenerife. J. Veg. Sci. 1998, 9, 297–306. [Google Scholar] [CrossRef]
- Bowman, D.; Kirkpatrick, J. Establishment, suppression and growth of Eucalyptus delegatensis R.T. Baker in multiaged forests. iii. Intraspecific allelopathy, competition between adult and juvenile for moisture and nutrients, and frost damage to seedlings. Aust. J. Bot. 1986, 34, 81–94. [Google Scholar] [CrossRef]
- Attiwill, P.M. Ecological disturbance and the conservative management of eucalypt forests in Australia. For. Ecol. Manag. 1994, 63, 301–346. [Google Scholar] [CrossRef]
- Van Der Meer, P.J.; Dignan, P.; Saveneh, A.G. Effect of gap size on seedling establishment, growth and survival at three years in mountain ash (Eucalyptus regnans F. Muell.) forest in Victoria, Australia. For. Ecol. Manag. 1999, 117, 33–42. [Google Scholar] [CrossRef]
- Harmon, M.E.; Hua, C. Coarse woody debris dynamics in two old-growth ecosystems. BioScience 1991, 41, 604–610. [Google Scholar] [CrossRef]
- Zielonka, T. When does dead wood turn into a substrate for spruce replacement? J. Veg. Sci. 2006, 17, 739–746. [Google Scholar] [CrossRef]
- Chećko, E.; Jaroszewicz, B.; Olejniczak, K.; Kwiatkowska-Falińska, A.J. The importance of coarse woody debris for vascular plants in temperate mixed deciduous forests 1. Can. J. For. Res. 2015, 45, 1154–1163. [Google Scholar] [CrossRef]
- Laiho, R.; Prescott, C.E. Decay and nutrient dynamics of coarse woody debris in northern coniferous forests: A synthesis. Can. J. For. Res. 2004, 34, 763–777. [Google Scholar] [CrossRef]
- Harmon, M.E.; Franklin, J.F. Tree seedlings on logs in picae-tsuga forests of Oregon and Washington. Ecology 1989, 70, 48–59. [Google Scholar] [CrossRef]
- Van Der Meer, P.J.; Bongers, F. Patterns of tree-fall and branch-fall in a tropical rain forest in French Guiana. J. Ecol. 1996, 84, 19–29. [Google Scholar] [CrossRef]
- Boswijk, G.; Fowler, A.; Palmer, J.; Fenwick, P.; Hogg, A.; Lorrey, A.; Wunder, J. The late Holocene kauri chronology: Assessing the potential of a 4500-year record for palaeoclimate reconstruction. Quat. Sci. Rev. 2014, 90, 128–142. [Google Scholar] [CrossRef]
- Swetnam, T.W. Fire history and climate change in giant sequoia groves. Science 1993, 262, 885–889. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, T.; Jeltsch, F.; Hanski, I.; Grimm, V. Using pattern-oriented modeling for revealing hidden information: A key for reconciling ecological theory and application. Oikos 2003, 100, 209–222. [Google Scholar] [CrossRef]
- Grimm, V.; Revilla, E.; Berger, U.; Jeltsch, F.; Mooij, W.M.; Railsback, S.F.; Thulke, H.-H.; Weiner, J.; Wiegand, T.; DeAngelis, D.L. Pattern-oriented modeling of agent-based complex systems: Lessons from ecology. Science 2005, 310, 987–991. [Google Scholar] [CrossRef] [PubMed]
- Rademacher, C.; Neuert, C.; Grundmann, V.; Wissel, C.; Grimm, V. Reconstructing spatiotemporal dynamics of central European natural beech forests: The rule-based forest model before. For. Ecol. Manag. 2004, 194, 349–368. [Google Scholar] [CrossRef]
- Food and Agriculture Organisation (FAO) of the United Nations. Forest Resources Assessment: Terms and Definitions of Forests; FAO Forestry Department: Rome, Italy, 2012. [Google Scholar]
- Wing, B.M.; Ritchie, M.W.; Boston, K.; Cohen, W.B.; Gitelman, A.; Olsen, M.J. Prediction of understory vegetation cover with airborne lidar in an interior ponderosa pine forest. Remote Sens. Environ. 2012, 124, 730–741. [Google Scholar] [CrossRef]
- Vanderwel, M.C.; Malcolm, J.R.; Smith, S.M. An integrated model for snag and downed woody debris decay class transitions. For. Ecol. Manag. 2006, 234, 48–59. [Google Scholar] [CrossRef]
- Sollins, P.; Cline, S.P.; Verhoeven, T.; Sachs, D.; Spycher, G. Patterns of log decay in old-growth douglas-fir forests. Cana. J. For. Res. 1987, 17, 1585–1595. [Google Scholar] [CrossRef]
- Larson, A.J.; Lutz, J.A.; Donato, D.C.; Freund, J.A.; Swanson, F.J.; HilleRisLambers, J.; Sprugel, D.G.; Franklin, J.F. Spatial aspects of tree mortality strongly differ between young and old-growth forests. Ecology 2015, 11, 2855–2861. [Google Scholar] [CrossRef]
- Muir, P.S. Disturbance effects on structure and tree species composition of Pinus contorta forests in western Montana. Can. J. For. Res. 1993, 23, 1617–1625. [Google Scholar] [CrossRef]
- Aakala, T.; Kuuluvainen, T.; De Grandpre, L.; Gauthier, S. Trees dying standing in the northeastern boreal old-growth forests of Quebec: Spatial patterns, rates, and temporal variation. Can. J. For. Res. 2007, 37, 50–61. [Google Scholar] [CrossRef]
- Duyker, E.; Duyker, M. Bruny D’entrecasteaux: Voyage to Australia and the Pacific 1791–1793; Miegunyah/Melbourne University Press: Melbourne, Australia, 2006; p. 392. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
| Author(s) | Forest Type | Topic of Paper | Study Impact * Key Development/Finding Type ^ | Cites | Terminology | ||
|---|---|---|---|---|---|---|---|
| CONSEQUENCES Canopy gaps and gap dynamics | |||||||
| Brokaw, 1985 [18] | Tropical | An assessment of Watt’s [19] description of mature forests as shifting mosaics and gap size dependence of regrowth using periodic observations at multiple sites. | FM | P | Accordance with Watt (1947): Gaps of different sizes and stages of regrowth are an important source of heterogeneity in composition and dynamics of tropical forests. | 821 | Treefall gaps |
| Uhl et al., 1988 [20] | Amazon forests | Effect of gap microhabitats on nutrient availability and regeneration, and the role of gap size in influencing regeneration within a gap within 4 years after gap establishment. | FE and FM | C | (i) Treefall gaps principally benefit pre-established seedlings and saplings; (ii) gap size has little effect on plant density, establishment and mortality; and (iii) gap microhabitats have negligible effects on vegetation dynamics and nutrient availability. | 398 | Treefall gaps |
| Muscolo et al., 2014 [21] | All forest types | A review of the roles of forest canopy gaps. | R | P | To further understand the impact of gaps on the forest as a whole, further investigation needs to be done on the below-ground communities (including: soil processes, organic matter trends, composition and activity of the microbial biomass, and soil characteristics). | 3 | Treefall gap |
| Zhu et al., 2014 [22] | All forest types | Meta-analysis on the effect of gaps on woody-plant regeneration. | M | E | Overall, forest gaps enhance woody plant regeneration, and the effects of gaps varies with forest type, gap characteristics, environmental factors and plant traits. | 8 | Forest gaps and treefall gaps |
| CAUSES Disturbance (Extreme weather events (fire and wind), uprooting) | |||||||
| Canham and Loucks, 1984 [23] | Hardwoo d forest | Assessment of the frequency and extent of catastrophic windthrow, and identification of the mechanisms. | FM | P | Return time for a catastrophic windthrow in these forests is 1210 years with catastrophic thunderstorms as the principle mechanism for large-scale blowdown. | 379 | Blowdown and windthrow |
| Attiwill, 1994 [24] | All forest types | A review of the literature on natural disturbances in forests. | R | P | An ecological framework of natural disturbances and its component processes and effects is synthesised and developed in this review. Provides the basis for sustainable forest management. | 798 | Tree fall |
| Ulanova, 2000 [13] | Boreal forest | A review of the literature on the ecological effects of windthrow and its effects on forest structure and composition at differing spatial scales. | R | L and C | The creation of gaps and microsites by windfall disturbances are two related major environmental components determining the regeneration niche of forest plant species. Windfall disturbances also increase biodiversity (for further information on how, see the paper). | 310 | Gap-phase dynamics, windthrow and fallen tree |
| Šamonil et al., 2010 [25] | All forest types | Investigation of the reported roles of tree uprooting in soil formation. | R | E | Knowledge of the effect of tree uprooting on soil formation is well understood across multiple scales, but there is limited quantitative data to date supporting much of the theory. | 55 | Tree uprooting |
| Bassett et al., 2015 [26] | Eucalypt forest | Development of a conceptual model of CWD dynamics pre- and post-fire to predict how topography, fire severity, and fire history interact to affect the availability of CWD in forests. | M | L | Both fire severity and fire frequency influenced CWD availability in gullies, where CWD on slopes was not. Gullies generally supported more logs than slopes, but longer inter-fire intervals in gullies may allow fuel loads to accumulate and lead to comparatively larger fire impacts. | 2 | CWD and logs and dead trees |
| CAUSES Tree mortality and Standing dead | |||||||
| Franklin et al., 1987 [12] | All forest types | Description of tree death as an ecological process. | R | I to P | Tree death demonstrates key principles of ecological process, and defining the spatial and temporal context of the study is important. | 683 | Tree death |
| Fridman and Walheim, 2000 [2] | All forest types | Evaluation of the dead-wood inventory in Sweden. | FM/M | C | Logs were more decayed than stags. Forest management affects the amount of dead wood present, less dead wood using clear-cut and thinning methods. Inventories are important and are usable as a tool for assessments of dead wood. | 403 | Dead wood, standing dead |
| Lugo and Scatena, 1996 [27] | Rainforest | Causes and consequences of tree mortality. | R | C | Tree mortality events vary extensively across time and space. These events often differ depending on stand conditions, and stem densities (higher mortality with higher densities). Ecological impacts of a sudden tree mortality event contrast with gradual background mortality. | 28 | Tree mortality, tree fall gaps |
| Soderberg et al., 2014 [16] | Boreal forest | Assessment of the choice of definition on the amount of dead wood that is reported in the literature. | FM | C | The different definitions of ‘dead wood’ used in studies, substantially affects the quantity of dead wood that is reported –in most cases leading to underestimates of dead wood volume. | 1 | Dead wood |
| CONSEQUENCES Decay and nurse logs (recruitment) | |||||||
| Sollins, 1982 [28] | Douglas-fir forest | Decay rates and turnover in an oldgrowth forest, assessment of prior measurements that may have been misleading. | FM | C | Densities of fallen boles were lower than previously reported due to methodological and field measurement differences. Highlights the value of permanent plots (undisturbed) for accurate representation of decomposition and nutrient dynamics. | 330 | Tree mortality, fallen boles, fallen and standing dead woody material |
| Siitonen et al., 2000 [14] | Norway spruce forest | Differences in stand structure between managed and unmanaged stands (comparing mature and oldgrowth). | FM | C | Average volume of CWD was much higher in old-growth (managed) than mature (managed) and over mature stands. Logs contributed the most to CWD volume. | 376 | Coarse woody debris (CWD), living trees, logs, dead standing trees |
| Weedon et al., 2009 [29] | All forest types | A global meta-analysis testing the hypothesis that interspecific differences in wood traits affect decomposition of woody debris. | R | E | Found support for their hypothesis. Gymnosperm wood decomposes more slowly that angiosperm, and key nutrients such as nitrogen and phosphorus correlate with decomposition of angiosperm woody debris. | 152 | Woody debris |
| Cousins et al., 2015 [30] | Mixed conifer forest | Developing an understanding of decay rates of standing dead (SD) trees and the implications for carbon accounting in forests. | FM | C | Carbon density of the most decayed SD trees was 60% that of live trees. Species identity, surface area:volume ratio and relative position within the tree are all important characteristics that explained the SD patterns. | 1 | Standing dead (SD) trees, woody debris, deadwood |
| CHARACTERISTICS Non-living and Structural Elements | |||||||
| Maser and Trappe, 1984 [31] | All forest types | Synthesis of the available data on fallen trees in unmanaged forests with the aim of highlighting research needs and knowledge gained. | R | E | We know a lot about fallen trees their importance. Future research should focus on the effect of changes in and around the fallen tree on the overall functioning of forest ecosystems. Soil-log interface may be important. | 327 | Fallen trees, wood, woody debris |
| Harmon et al., 1986 [32] | Temperate forests | Describes CWD and its flow/movement into, from and within an ecosystem. | R | E | Rates of input/accumulation of CWD in forests ranges from 0.12 to 30 Mg/year. This input depends largely on the size of the tree (rate of decomposition), and frequency of disturbances (particularly big ones!). CWD mortality is expected to be aggregated (due to the processes governing tree death such as wind, pathogens affecting multiple individuals of the one area). | 3166 | Woody debris, coarse woody debris (CWD), dead trees, downed boles, logs |
| Woldendor p and Keenan, 2005 [33] | Australian forest | Assessment and literature review of CWD in Australian forests. | R | E | CWD can be high in exotic pine plantations if substantial amounts remain from the once native forest. There were differences in CWD quantity depending on stand age; young <20 years = lowest percentage of CWD, older >70 years = highest CWD and stag biomass. Tree size influences CWD amounts (i.e., tall open forests CWD much greater because of taller trees). | 67 | Coarse woody debris (CWD), standing and fallen dead wood, snags |
| Oberle et al., 2015 [34] | Temperate forest | Importance and movement of deadwood after treefall. | FM | C | While logs can fall in many different directions; snags, logs and branch average direction was consistent with downhill deadwood movement as trees fall. Relationships between log and landscape attributes (movement, shape and topography) suggest that downhill rotation during treefall drives most of the deadwood distribution over their study site. | 1 | Logs, deadwood, snags |
| APPLICATION Modelling and For. Management | |||||||
| Lorimer, 1985 [35] | All forest types | How to infer past disturbance dynamics without using destructive techniques and using more than just age of trees. | R/M | P | Currently, it is difficult to identify past disturbance (severity) from the distribution of tree among age-classes. They advocate that to study disturbance history, random dispersal of plots of various sizes, across a large land area is recommended. | 280 | |
| Siitonen, 2001 [1] | Boreal forest | Exploring the relationships between CWD, forest management (intensively vs. unmanaged), and saproxylic species. | R | E | In managed forests, the average amount of CWD in the landscape has reduced by 90–98%. General species area relationships suggest that a reduction in available habitat (i.e., as logs), might lead to a reduction of >50% saproxylic species in the long term. It is important to assess how much CWD should be retained in managed forests. | 904 | Coarse woody debris (CWD), dead tree, decaying wood |
| Schliemann and Bockheim, 2011 [36] | All forest types | Review of the inconsistencies in gap terminology, and the methods and modelling used to investigate treefall gaps and the influence of gaps in a forest system. | R | E | Recommends a range of standard protocols when studying treefall gaps. For example the size of treefall gaps varied across studies; this study suggested a maximum gap size of 1000 m2 (gap size can influence the results due to scale dependencies of processes). Gap shape varies and should be determined through extensive field survey. | 70 | Treefall gaps, canopy gap |
| Fischer et al., 2016 [37] | All forest types | Description of the development of the individual-based and process-based forest gap model FORMIND and its potential application to tropical forests. | M | E | Long-term modelling projects not only provide understanding of forest systems, but also provide benefits for ecological theory and empirical study design. They are powerful tools, and are becoming increasingly valuable in today’s research. | 0 | Forest gap |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buettel, J.C.; Ondei, S.; Brook, B.W. Look Down to See What’s Up: A Systematic Overview of Treefall Dynamics in Forests. Forests 2017, 8, 123. https://doi.org/10.3390/f8040123
Buettel JC, Ondei S, Brook BW. Look Down to See What’s Up: A Systematic Overview of Treefall Dynamics in Forests. Forests. 2017; 8(4):123. https://doi.org/10.3390/f8040123
Chicago/Turabian StyleBuettel, Jessie C., Stefania Ondei, and Barry W. Brook. 2017. "Look Down to See What’s Up: A Systematic Overview of Treefall Dynamics in Forests" Forests 8, no. 4: 123. https://doi.org/10.3390/f8040123
APA StyleBuettel, J. C., Ondei, S., & Brook, B. W. (2017). Look Down to See What’s Up: A Systematic Overview of Treefall Dynamics in Forests. Forests, 8(4), 123. https://doi.org/10.3390/f8040123