Abstract
More intensive removal of woody biomass for the bio-economy will disrupt litter and succession cycles. Especially at risk is the retention of fine and coarse woody debris (FWD and CWD), crucial factors in forest biodiversity and nutrient cycling. However, to what extent CWD affects soil functioning remains unknown, and is seldom considered. From 32 paired test–reference points in eight Fagus sylvatica (L.) stands throughout Southwest Germany, CWD significantly increased soil C/N ratios, base saturation, and possibly pH. CWD-induced changes in soil porosity, available water capacity, and total organic carbon depended on site and CWD characteristics. As such, CWD can be viewed as a “pedogenic hot-spot” of concentrated biogeochemical and -physical processes with outsized effects on soil functioning and development. CWD management for soil functioning should consider site and tree species specific volume thresholds, timed rotations, and spatial densities, but appropriate implementation requires further research to define best management practices. If successful, overall forest resilience as well as soil functioning and productivity can be improved.
1. Introduction
Forests provide numerous functions and services. These are of growing public and scientific interest, as reflected by bio-economy initiatives where biomass is produced and converted into higher-value products [1]. Yet removing biomass from forests is an ecological disturbance. Not only is it a disturbance in the physical sense, but also in the counterfactual sense that when more biomass is removed, there is less potential bio- and necromass in both present and future forest states.
As a disturbance, removing bio- and necromass affects forest ecosystems. Extensive whole-tree harvesting can deplete forest nutrient stocks [2], while an absence of deadwood can lead to lower biodiversity [3]. The extent, however, to which a forest ecosystem is affected by disturbances depends on its resilience; i.e., its ability to absorb disturbance-induced changes while maintaining similar functions, structures, and feedbacks [4,5]. Forest management can be tailored to use forest services while preserving resilience [6], but such steps are wholly dependent on the objectives and ecological consequences of chosen management practices.
Balance between disturbances and ecosystem functioning is illustrated by the effects that removing fine and coarse woody debris (FWD and CWD)—the counterpart of removing more biomass—has on soil functioning. Centuries of litter raking in Central Europe had observable detrimental consequences for soil and forest productivity through nutrient removal [7]. In comparison, the extent to which CWD affects soil ranges from minimal to extensive, depending on species and decay processes [8]. Consequently, retaining harvest residues (i.e., FWD) for soil productivity is recommended [9], while CWD guidelines often concentrate on wildlife conservation without addressing soil protection despite uncertain dynamics for soil organic matter [10] and soil functioning (e.g., Alt-und Totholz Konzept Baden-Württemberg [11]).
In this short paper, we investigate whether CWD influences soil functioning in terms of soil organic matter (SOM), nutrient availability, pH, porosity, and available water capacity (AWC) in eight European beech (Fagus sylvatica L.) stands, and if so, to what extent. We propose that CWD can be considered “pedogenic hot-spots” in forest ecosystems, and as such we outline potential management strategies and additional research necessary for their implementation.
2. Materials and Methods
Mineral soil samples (0–10 cm) came from eight European beech stands at least 90 years old in the Swabian Jura, the Black Forest, the Palatinate Forest, and the Schurwald in SW Germany; coordinates, site characteristics, and soil types are listed in Table 1. A total of 32 pairs of test and reference points (Deadwood and Control points, respectively) were sampled from four pieces of downed beech CWD at each of the eight stands. Deadwood points were immediately adjacent to the middle of CWD, while Control points were 2–3 m away perpendicularly. Selected CWD was parallel to the slope, ≥15 cm in diameter (at point of sampling), ≥1.8 m in length, and spanned three decay classes based on penetrability from Lachat et al. [12] (classes 2–4); exact age was unknown.

Table 1.
Coordinates, underlying bedrock, forest floor type, and soil reference group for the eight study sites. Mull and moder are differentiated by the absence or development of an Oa horizon, respectively.
Bagged samples from each of the sampling points were dried at 40 °C, mixed, sieved <2 mm (A1.3.2 [14]), and aliquoted to measure residual water content after drying at 105 °C. Additional subsamples were milled ≈10 m, dried at 105 °C, and combusted in Sn-foil caps at 1150 °C for total carbon and nitrogen concentrations [15]. Total organic C (OC) was assumed to be total C, except for calcareous sites Siebeter Fuss and Teutschbuch, where it was calculated as C loss upon ignition at 550 °C. Potential cation exchange capacity (CEC; NH4-Acetate & KCl; Lakuvich 1981 [16]) and pH (H2O; A3.1.1.1 [14]) were measured from un-milled aliquots corrected for residual water; analytic equipment is listed in the Appendix. Porosity [17] and available water capacity (AWC, [18]) were measured from 100 cm, structured soil rings through vacuum pycnometry at field moisture content (θ), complete saturation with gypsum-treated water, desorption at 300 hPa in a pressure pot to an equilibrium θ (pores retaining water are 10–0.2 m in diameter), subsequent desorption at 15,000 hPa of a 1 cm aliquot to another equilibrium θ (pores <0.2 m, or dead water), and complete drying at 105 °C.
As in Stutz et al. [8], differences between paired points were calculated as:
where Δ is the difference between Deadwood (test) and Control (reference) points in absolute terms for x soil property, and is the difference between Deadwood and Control points relative to Control points. Significance of differences between paired points was tested with linear mixed effects (LME) models with each site and piece of CWD set as nested random factors. Stepwise linear regressions of best fit using ANOVA checked whether Δ depended on site or CWD characteristics. Statistics were done with R 3.2.3 (2015).
3. Results
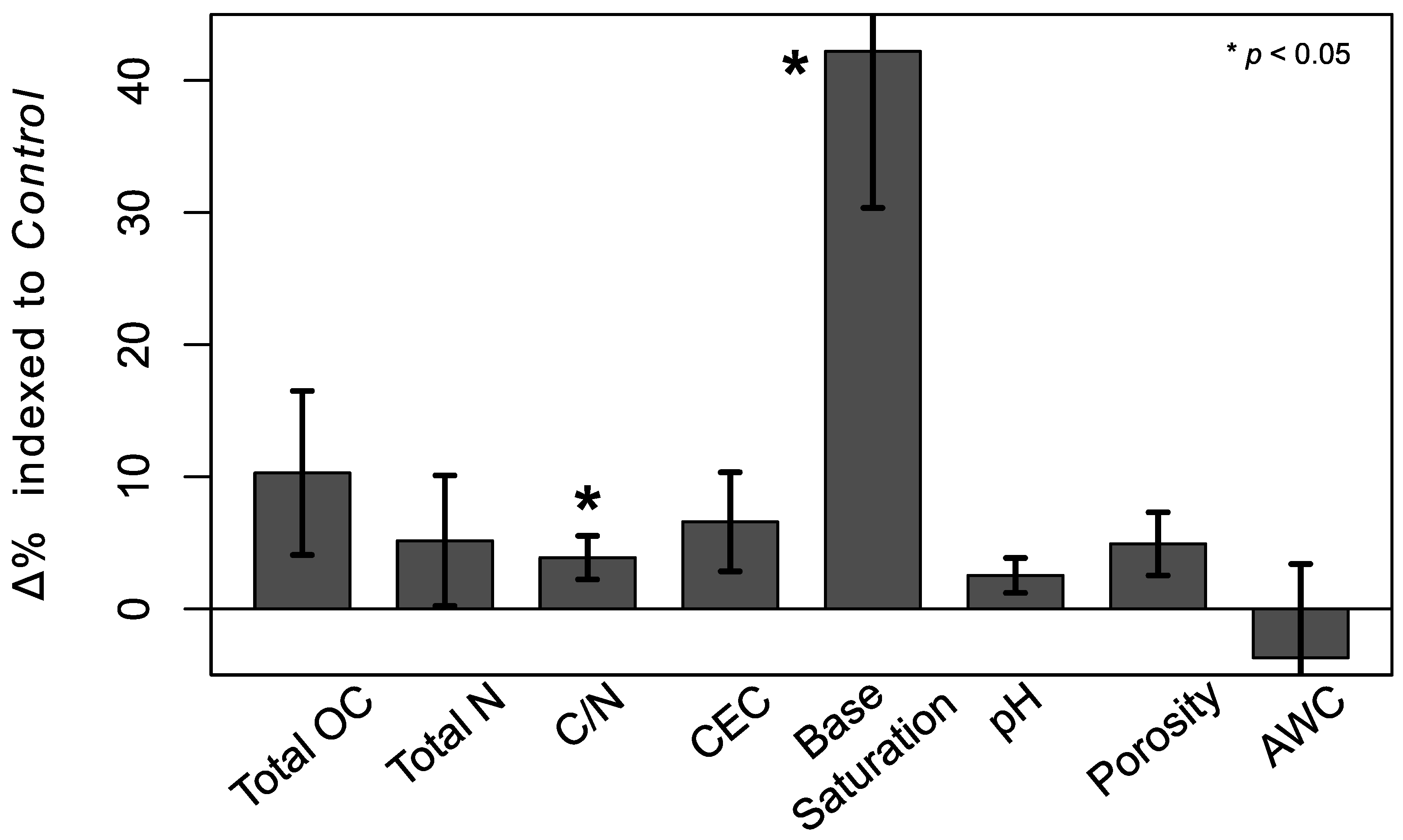
Despite no changes in total OC and N concentrations between all 32 pairs of Deadwood and Control points, C/N ratios did change significantly (Figure 1). Likewise total CEC did not increase significantly, while base saturation did and pH tended to (LME models, p < 0.1). Neither total porosity nor AWC differed significantly between paired points.
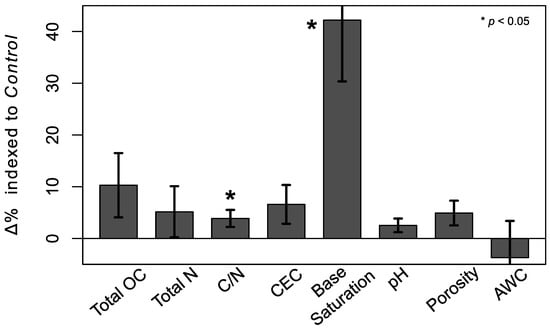
Figure 1.
Mean (Equation (2)) for each soil property. Stars indicate significant differences between Deadwood and Control points (linear mixed effects models, p < 0.05). Total organic carbon (OC), N, cation exchange capacity (CEC), and pH are based on concentrations, while C/N, base saturation, porosity, and available water capacity (AWC) are based on ratios. Error bars are one standard error of mean .
However, changes between paired points depended on differing factors for the investigated aspects of soil functioning (Δ, Equation (1)). Δ total OC depended significantly on the state of wood decay (stepwise regression, p < 0.05); total N tended to as well (p < 0.1). In comparison, Δ porosity depended on forest floor type (p < 0.05), while Δ AWC depended on both underlying bedrock (calcareous or silicate, p < 0.01) and CWD diameter (p < 0.05), without a significant interaction between the two factors. Similarly, Δ base saturation tended to depend on CWD diameter (p < 0.1).
4. Discussion
Increased C/N ratios, base saturation, and possibly pH at Deadwood points would result from an influx of organic matter and nutrients that were quantitatively and chemically different to leaf litter and bulk SOM. Biological communities at various scales would consequently react to that influx, leading to both mineralized organic matter and the growth of assorted organisms including fungi that are actively decomposing CWD. Through direct and indirect processes, such biological activity would disrupt or form micro- and macroaggregates, modify pore structures, and thus alter soil aeration, water holding capacity, and structural stability.
However, no significant changes to soil porosity or AWC were found. Likewise, none were found for total OC, N, or CEC. This lack of changes for all 32 pairs reflect that other factors play confounding roles, as indicated by the stepwise regression analysis. Forest floor type—which corresponds to meso- and macrofauna activity—significantly affected Δ porosity, while the acidity of the underlying bedrock and the diameter of CWD significantly affected Δ AWC; diameter of CWD may also have influenced Δ base saturation. Similarly, the extent of CWD decay influenced Δ total OC and possibly Δ total N, which would have consequences for SOM and nutrient availability. It should also be noted that significant differences between paired points imply that some direct influences of CWD on mineral soil are spatially limited. In contrast, non-significant differences may be due to either spatial influences greater than 2–3 m, site-CWD characteristics, or no influence at all.
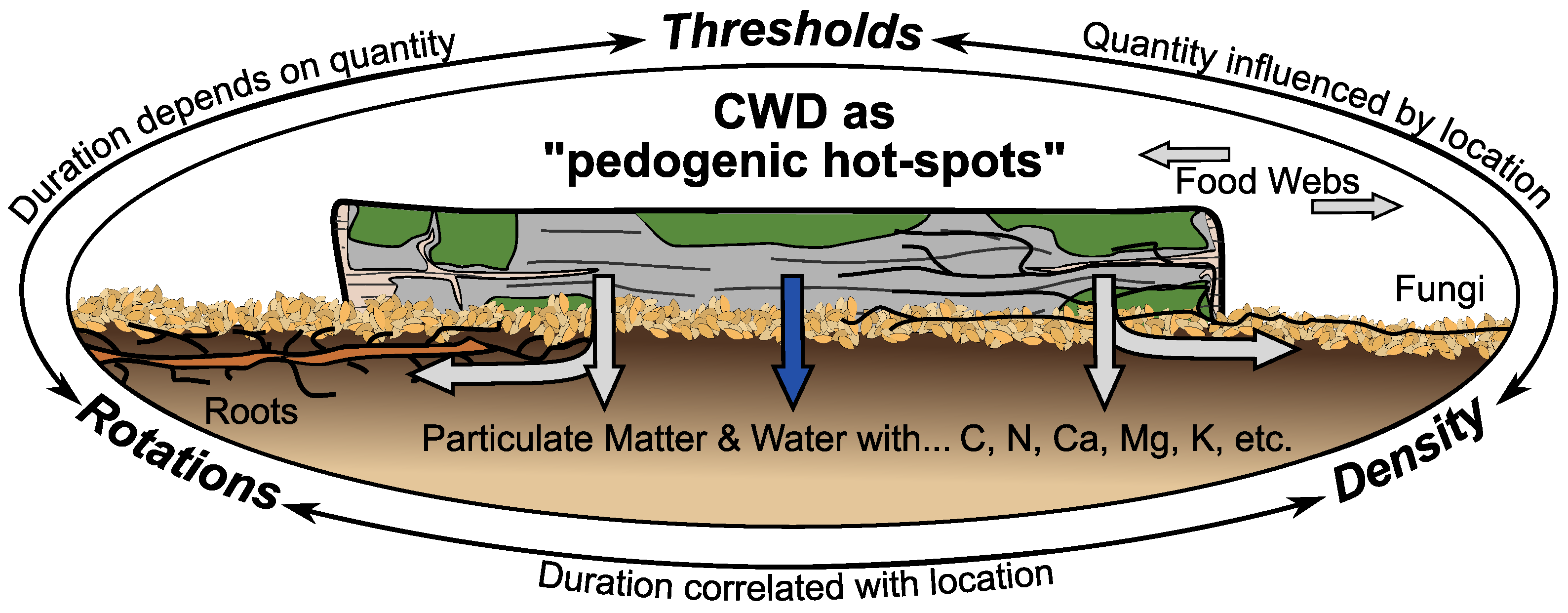
Together these results suggest that CWD are transient and spatially defined centers of concentrated biogeochemical and -physical processes that influence soil functioning. Such properties are characteristic features of both microbial hot-spots as defined by Kuzyakov and Blagodatskaya [19], and general biogeochemical hot-spots as defined by McClain et al. [20]. Consequently, CWD warrants being designated and considered as “pedogenic hot-spots” at the scale of forest stands (Figure 2). Additional (microbial) hot-spots likely occur within and underneath CWD, but the mixture of differing process rates, concentrated nutrients, and affected biological communities is nevertheless spatiotemporally distinguishable to surrounding forest litter and soil.
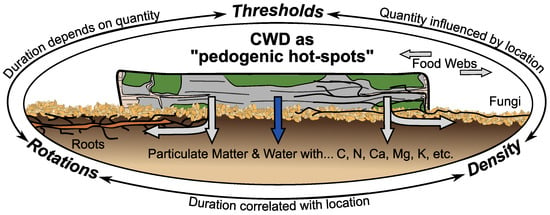
Figure 2.
Coarse woody debris (CWD) are concentrated centers of biogeochemical and -physical processes in space and time—i.e., “pedogenic hot-spots”—which have larger effects on forest and soil ecosystems through roots, fungi, and other biological networks. Minimum thresholds, rotation cycles, and spatial densities are potential management parameters, but specifics are unknown and interrelated.
The role of hot-spots and thus CWD in soil and forest ecosystems are not limited to their immediate surroundings and moment in time. Fungal growth alone has the potential to move nutrients at m scales between individual trees [21]; likewise with tree roots, which probably respond to CWD. Similarly CWD’s role for biodiversity includes habitat niches and communities that are integrated into entire forest (and soil) ecosystems [22]. We would also like to emphasize that CWD not only contributes C quantitatively to soils, but also changes the compositional quality of surrounding SOM. Previous analyses at the study site Conventwald indicated that the action of wood-decaying fungi control SOM and the properties of soil affected by deadwood [8]. These spots of biogeochemical processes and biological refugia would logically contribute to soil and forest buffering capacity against stresses and disturbances, which are crucial parts of forest resilience and health [23]. Altogether this implies that the presence of CWD—the counterfactual to removing more biomass—would have consequences not only for forest and soil functioning, but also for their resilience and development.
If so, managing CWD for soil functioning, resilience, and development is possible. Within the context of silviculture, three broad parameters can be defined: thresholds, rotations, and densities (Figure 2). Minimum thresholds are already used for biodiversity, and as such could be easily adjusted for soil functioning. Stand rotations could also incorporate rotations of CWD in various states of decay. And spatial densities of CWD could fit into similar planning for felling, regeneration, and recruitment.
However, successfully managing CWD for soil functioning through thresholds, rotations, and densities requires currently-unknown answers to three questions: (i) “how much CWD is necessary to influence soil functioning?”; (ii) “for how long does CWD influence soil functioning?”; and (iii) “to what distance does CWD influence soil functioning?” (Figure 2). These questions are pertinent research questions in their own right, but they are also interrelated. Thresholds entail a minimum amount that will last a certain duration and have a limited spatial effect. Rotations rely on the duration of effects, but that is influenced by the quantity and density. Densities rely on the spatial extent to which CWD influences soil functioning, but that depends on the quantity and duration CWD is present. Even then, one additional overall question remains: “what are the site, species, and management properties that control the answers to the above-mentioned questions?”.
Even with such knowledge gaps, adopting these soil management objectives and parameters goes some way in transitioning to a more holistic management of CWD, soils, and forests envisioned in Harmon [24] and Janzen [25]. Yet integrating such objectives and concerns with already-existing ones will not always be straightforward and compatible. Biodiversity stands to benefit from more CWD in most situations, while risks of fire and disease often lead to less CWD. Likewise “old-growth” silviculture incorporates higher stocks of CWD, but minimizes managerial activities to reduce disturbance [26], which is at odds with the outlined potential management parameters. Still, the outcomes of managing CWD for soils and forests ought to be considered, and can provide synergies in more complex, resilient, multi-aged silvicultural systems and adaptively-managed protected areas [6,27]. For example, large retention patches in spruce boreal forests recruited CWD at levels similar to post-fire patches [28], which would benefit both soils and biodiversity.
5. Conclusions
To summarize, European beech CWD from eight stands in SW Germany influenced mineral soil C/N ratios, base saturation, and possibly pH. Additionally, CWD-induced changes in soil porosity, AWC, total OC, and possibly total N and base saturation depended on the type of forest floor, underlying bedrock, CWD diameter, and CWD decay. Altogether, these results imply that CWD are transient and spatially-limited centers of biogeochemical and -physical processes that influence soil functioning. This warrants CWD being designated as “pedogenic hot-spots” in forest ecosystems.
In conclusion, quantitative thresholds, rotation cycles, and spatial densities are potential parameters to manage CWD for soil functioning. However, open interrelated questions on underlying processes and specific values remain to be answered, as well as how to incorporate other management objectives. If done successfully, soil and forest functioning can be improved while maintaining forest resilience through more complete ecosystems, which has consequences for both soil development and forest use.
Acknowledgments
We wish to thank Daniel Dann, Petra Grossmann, Jaane Krüger, Christine Petschke, Angela Thiemann, Paula Vollmer, and Janna Wambsganss for their expertise and assistance; Morgan Barnes for her helpful suggestions; and the two anonymous reviewers for their supportive comments. We also thank the Forstliche Versuchs-und Forschungsanstalt Baden Württemberg, the Forschungsanstalt für Waldökologie und Forstwirtschaft Rheinland Pfalz, ForstBW, and the Landesforsten RLP for providing access to study sites. This work was supported by a grant from the Ministry of Science, Research and the Arts of Baden-Württemberg (Az: 33-7533-10-5/81) to Kenton Stutz and Friederike Lang. The fee necessary for open-access was awarded to Kenton Stutz for his oral presentation titled “Soil Ecological Challenges for Foresty in the Bioeconomy” at the Forstwissenschaftliche Tagung 2016 in Freiburg, from which this paper was written.
Author Contributions
Kenton Stutz and Friederike Lang conceived and designed the experiments, and formulated the discussion; Kenton Stutz performed the experiments, analyzed the data, and wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest. The funding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| FWD | Fine woody debris |
| CWD | Coarse woody debris |
| SOM | Soil organic matter |
| OC | Organic carbon |
| CEC | Cation exchange capacity |
| AWC | Available water capacity |
| LME | Linear mixed effects |
Appendix A
Equipment for laboratory analyses: total carbon and nitrogen, Elementar Vario EL Cube (Langenselbold, Germany); potential CEC, Spectro Ciros CCD ICP Side-on Plasma Optical Emission Spectrometer (Kleve, Germany), Skalar Sanplus system (Breda, Netherlands); pH, Metrohm Titrino 751 GPD meter (Herisau, Switzerland).
References
- Ronzon, T.; Santini, F.; M’Barek, R. The Bioeconomy in the European Union in Numbers. Facts and Figures on Biomass, Turnover and Employment; European Commission, Join Research Center, Institute for Prospective Technological Studies: Seville, Spain, 2015. [Google Scholar]
- Achat, D.L.; Deleuze, C.; Landmann, G.; Pousse, N.; Ranger, J.; Augusto, L. Quantifying consequences of removing harvesting residues on forest soils and tree growth—A meta-analysis. For. Ecol. Manag. 2015, 348, 124–141. [Google Scholar] [CrossRef]
- Albrecht, L. The importance of dead woody material in forests. Forstwiss. Cent. Ver. Mit Tharandter Forstl. Jahrb. 1991, 110, 106–113, (In German, with English Abstract). [Google Scholar] [CrossRef]
- Holling, C.S. Resilience and stability of ecological systems. Annu. Rev. Ecol. Syst. 1973, 4, 1–23. [Google Scholar] [CrossRef]
- Walker, B.; Holling, C.S.; Carpenter, S.R.; Kinzig, A. Resilience, adaptability and transformability in social-ecological systems. Ecol. Soc. 2004, 9, 5. [Google Scholar] [CrossRef]
- DeRose, R.J.; Long, J.N. Resistance and resiliance: a conceptual framework for silviculture. For. Sci. 2014, 60, 1205–1212. [Google Scholar]
- Glatzel, G. The impact of historic land use and modern forestry on nutrient relations of Central European forest ecosystems. Fertil. Res. 1991, 27, 1–8. [Google Scholar] [CrossRef]
- Stutz, K.P.; Dann, D.; Wambsganss, J.; Scherer-Lorenzen, M.; Lang, F. Phenolic matter from deadwood can impact soil properties. Geoderma 2017, 288, 204–212. [Google Scholar] [CrossRef]
- Vance, E.D.; Aust, W.M.; Strahm, B.D.; Froese, R.E.; Harrison, R.B.; Morris, L.A. Biomass harvesting and soil productivity: Is the science meeting our policy needs? Soil Sci. Soc. Am. J. 2014, 78, S95–S104. [Google Scholar] [CrossRef]
- Magnússon, R.I.; Tietema, A.; Cornelissen, J.H.C.; Hefting, M.M.; Kalbitz, K. Tamm review: Sequestration of carbon from coarse woody debris in forest soils. For. Ecol. Manag. 2016, 377, 1–15. [Google Scholar] [CrossRef]
- ForstBW. Alt-und Totholzkonzept Baden-Württemberg; ForstBW: Stuttgart, Germany, 2015. (In German) [Google Scholar]
- Lachat, T.; Brang, P.; Bolliger, M.; Bollmann, K.; Brändli, U.B.; Bütler, R.; Hermann, S.; Schneider, O.; Wermelinger, B. Totholz im Wald. Entstehung, Bedeutung und Förderung. Merkbl. Prax. 2014, 52, 12S. (In German) [Google Scholar]
- WRB, I.W.G. World Reference Base for Soil Resources 2014. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; Number 106 in World Soil Resources Reports; FAO: Rome, Italy, 2014. [Google Scholar]
- Gutachterausschuss Forstliche Analytik. Handbuch Forstliche Analytik: Eine Loseblatt-Sammlung der Analysemethoden im Forstbereich; Bundesministerium für Verbraucherschutz, Ernährung und Landwirtschaft: Bonn, Germany, 2014. (In German) [Google Scholar]
- Nelson, D.W.; Sommers, L.E. Total Carbon, Organic Carbon, and Organic Matter. In Methods of Soil Analysis. Part 3. Chemical Methods; Sparks, D.L., Ed.; Soil Science Society of America: Madison, WI, USA, 1996; pp. 961–1010. [Google Scholar]
- Hendershot, W.H.; Lalande, H.; Duquette, M. Ion Exchange and Exchangeable Cations. In Soil Sampling and Methods of Analysis, 2nd ed.; Carter, M.R., Gregorich, E.G., Eds.; CRC Press: Boca Raton, FL, USA, 2006; p. 1262. [Google Scholar]
- Danielson, R.E.; Sutherland, P.L. Porosity. In Methods of Soil Analysis: Part 1—Physical and Mineralogical Methods, 2nd ed.; Klute, A., Ed.; Number 9 in Agronomy, Soil Science Society of America, American Society of Agronomy: Madison, WI, USA, 1986; pp. 443–461. [Google Scholar]
- Cassel, D.K.; Nielsen, D.R. Field Capacity and Available Water Capacity. In Methods of Soil Analysis: Part 1—Physical and Mineralogical Methods, 2nd ed.; Klute, A., Ed.; Number 9 in Agronomy, Soil Science Society of America, American Society of Agronomy: Madison, WI, USA, 1986; pp. 901–926. [Google Scholar]
- Kuzyakov, Y.; Blagodatskaya, E. Microbial hotspots and hot moments in soil: Concept & review. Soil Biol. Biochem. 2015, 83, 184–199. [Google Scholar]
- McClain, M.E.; Boyer, E.W.; Dent, C.L.; Gergel, S.E.; Grimm, N.B.; Groffman, P.M.; Hart, S.C.; Harvey, J.W.; Johnston, C.A.; Mayorga, E.; et al. Biogeochemical hot spots and hot moments at the interface of terrestrial and aquatic ecosystems. Ecosystems 2003, 6, 301–312. [Google Scholar] [CrossRef]
- Courty, P.E.; Bée, M.; Diedhiou, A.G.; Frey-Klett, P.; le Tacon, F.; Rineau, F.; Turpault, M.P.; Uroz, S.; Garbaye, J. The role of ectomycorrhizal communities in forest ecosystem processes: New perspectives and emerging concepts. Soil Biol. Biochem. 2010, 42, 679–698. [Google Scholar] [CrossRef]
- Harmon, M.E.; Franklin, J.F.; Swanson, F.J.; Sollins, P.; Gregory, S.V.; Lattin, J.D.; Anderson, N.H.; Cline, S.P.; Aumen, N.G.; Sedell, J.R.; et al. Ecology of coarse woody debris in temperate ecosystems. Adv. Ecol. Res. 1986, 15, 133–302. [Google Scholar]
- Trumbore, S.; Brando, P.; Hartmann, H. Forest health and global change. Science 2015, 349, 814–818. [Google Scholar] [CrossRef] [PubMed]
- Harmon, M.E. Moving towards a new paradigm for woody detritus management. Ecol. Bull. 2001, 49, 269–278. [Google Scholar]
- Janzen, H.H. Beyond carbon sequestration: soil as conduit of solar energy. Eur. J. Soil Sci. 2015, 66, 19–32. [Google Scholar] [CrossRef]
- Bauhus, J.; Puettmann, K.; Messier, C. Silviculture for old-growth attributes. For. Ecol. Manag. 2009, 258, 525–537. [Google Scholar] [CrossRef]
- O’Hara, K.L.; Ramage, B.S. Silviculture in an uncertain world: utilizing multi-aged management systems to integrate disturbance. Forestry 2013, 86, 401–410. [Google Scholar] [CrossRef]
- Moussaoui, L.; Fenton, N.J.; Leduc, A.; Bergeron, Y. Deadwood abundance in post-harvest and post-fire residual patches: an evaluation of patch temporal dynamics in black spruce boreal forest. For. Ecol. Manag. 2016, 368, 17–27. [Google Scholar] [CrossRef]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).