1. Introduction
Tree species are generally expected to expand their ranges to higher latitudes and elevations to keep pace with anthropogenic warming [
1,
2,
3]. The upper range edge of trees is the treeline ecotone and it is considered a sensitive biological indicator of climatic warming due to its strong, world-wide association with growing season temperature [
4,
5,
6,
7,
8]. Future treeline is generally expected to shift higher in elevation by as much as 700 m by the year 2100 [
9,
10], however there is considerable variability in the speed of treeline response to warming in mountains world-wide [
11]. This variation is likely due to several mitigating factors including geomorphology, lags in population changes, and dispersal limitations [
6,
12,
13,
14]. Soil type in particular may affect the ability of trees to establish above current treeline. Properties of soil such as water holding capacity and albedo are likely major factors in successful recruitment of sub-alpine forests. Despite treeline being generally limited by cold temperatures [
7], seedling establishment both at and above the treeline ecotone can be more immediately limited by low water availability and high temperatures [
15,
16,
17,
18].
In the Great Basin of the western United States, sub-alpine forests are predominantly composed of Great Basin bristlecone pine (
Pinus longaeva DK Bailey) and limber pine (
Pinus flexilis James). Sub-alpine stands of Engelmann spruce (
Picea engelmannii Parry) and quaking aspen (
Populus tremuloides Michaux) can also be found on wetter slopes in the eastern Great Basin, and whitebark pine (
Pinus albicaulis Engel) stands are found in parts of the northern and eastern Great Basin as well as on the eastern slope of the Sierra Nevada. Recent studies of Great Basin sub-alpine forests have shown that treeline is advancing through upslope establishment of young trees, but that this establishment has occurred only very recently and in infrequent pulses [
19]. In addition, it appears that even though bristlecone pine is usually the dominant adult tree at treeline where it is found, typically downslope limber pine is responsible for the majority of treeline advance in these stands, with young trees “leap-frogging” over treeline adult bristlecone pine [
19,
20]. This mismatch is especially acute on certain soil types where bristlecone pine adults dominate the sub-alpine forests, but the vast majority of upslope establishment is limber pine [
20]. Upslope establishment by young trees is important in predicting future forests because limber pine and bristlecone pine seedlings have very high survival, even in early life stages [
21]. Germination and establishment in these trees is likely the most significant bottleneck to long-term survival; understanding the limitations on these earliest life stages will be useful in predicting future forest composition.
There are three main soil types in the Great Basin. Granitic soil is weathered from intrusive plutonic igneous bedrock. These soils are generally grey-pink in color and are relatively course-grained (
Table 1). Quartzite soil is weathered metamorphic sandstone rock. It is generally relatively finely grained and dark grey in color. Dolomitic soil is from weathered carbonate rock with many properties similar to limestone. It is medium- to fine-grained and light grey to white in color [
22]. All three soil types are weathered to shallow, rocky sandy loams with a high proportion of surface cover by coarser rock. The percentage of surface rock in the three soil types was previously reported as dolomite 77%, quartzite 84%, and granite 27% [
22]. In the western Great Basin both bristlecone pine and limber pine are found across all three Great Basin soil types, allowing for an explicit comparison of soil preference at the leading edge of treeline species.
Where bristlecone pine is present, it is typically the dominant tree at treeline with the exception being on granitic soils where limber pine typically dominates [
19,
20]. Bristlecone pine is especially dominant on dolomite soils where nearly pure stands are often found to treeline [
23]. This association has been attributed to the high-pH, high-magnesium, and low-phosphorus soil properties excluding other species [
22,
24,
25]. The assumption is that bristlecone pine may prefer other soil types but is a poor competitor and that it thrives on dolomite because other species cannot tolerate the soil. However, bristlecone pine has recently been shown to have higher germination success on dolomite soil compared with quartzite and granite soils in the White Mountains with germination success being positively associated with cooler soil temperatures and higher soil moisture [
21]. If bristlecone pine prefers dolomite soil and if other species are also excluded from dolomite, bristlecone pine may have an even greater advantage on dolomite.
This study examined the preconception that limber pine, a potential competitor of bristlecone pine, is less able to establish on dolomite soil relative to other soil types, especially at the most vulnerable germination and establishment stage. Since adult limber pines are relatively rare on dolomite soil and relatively common on granite soils, germination and establishment should be higher on granite soils. However, given the recent evidence for limber pine establishing above bristlecone pine treeline on dolomite, an alternative hypothesis is that limber pine prefers soil types on which it is rarely found as an established adult [
19,
20]. This study first examines the relative abilities of different soil types to retain water over shorter (days) and longer (months) time scales. It then examines how temperature varies in the different soil types. Finally, it examines differential germination and establishment on the different soil types in an effort to test the hypothesis that limber pine prefers granitic soil where limber pine adults are largely found.
2. Materials and Methods
2.1. Soil Collection and Preparation
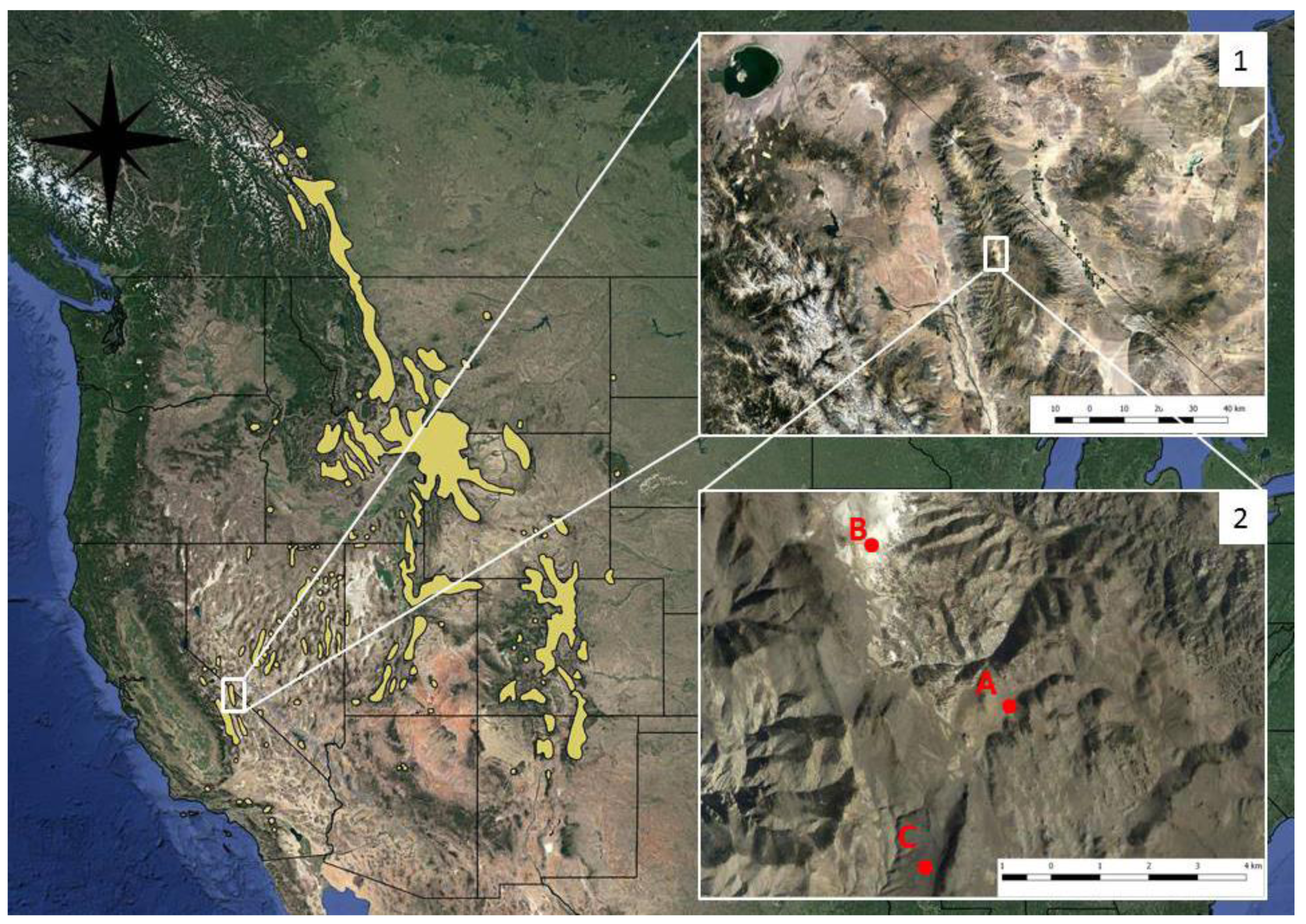
The White Mountains are the northern range of the contiguous White-Inyo Mountains and extend just over the border into Nevada at the extreme northern terminus of the range. The White Mountains are a high desert range parallel to and east of the central Sierra Nevada of California (
Figure 1) reaching a maximum elevation of 4319 m on White Mountain Peak. I collected soil samples of granite, dolomite, and quartzite soils from one location per soil type in the White Mountains, CA from road cuts to minimize disturbance and the chances of contamination by natural seeds (
Figure 1). I collected granite soil near the Crooked Creek Research Center, quartzite soil from the base of County Line Hill, and dolomite soil from Sheep Pass and then passed the soil through a 1-cm
2 screen to remove rocks. Fist-sized rocks were collected separately for experimental reasons (see below). The soil was collected in buckets and transported to the White Mountains Research Center, Crooked Creek Station at 3094 m elevation where I randomly placed each soil type in rectangular 900 cm
2 wooden containers. Soil depth in each container was 15 cm, and containers were arrayed in 3 rows of 12 for a total of 36 containers, allowing for 12 replicates for each soil type. I collected all soil on the same day in late October and sowed the seeds the day after soil was collected.
2.2. Soil Water Retention
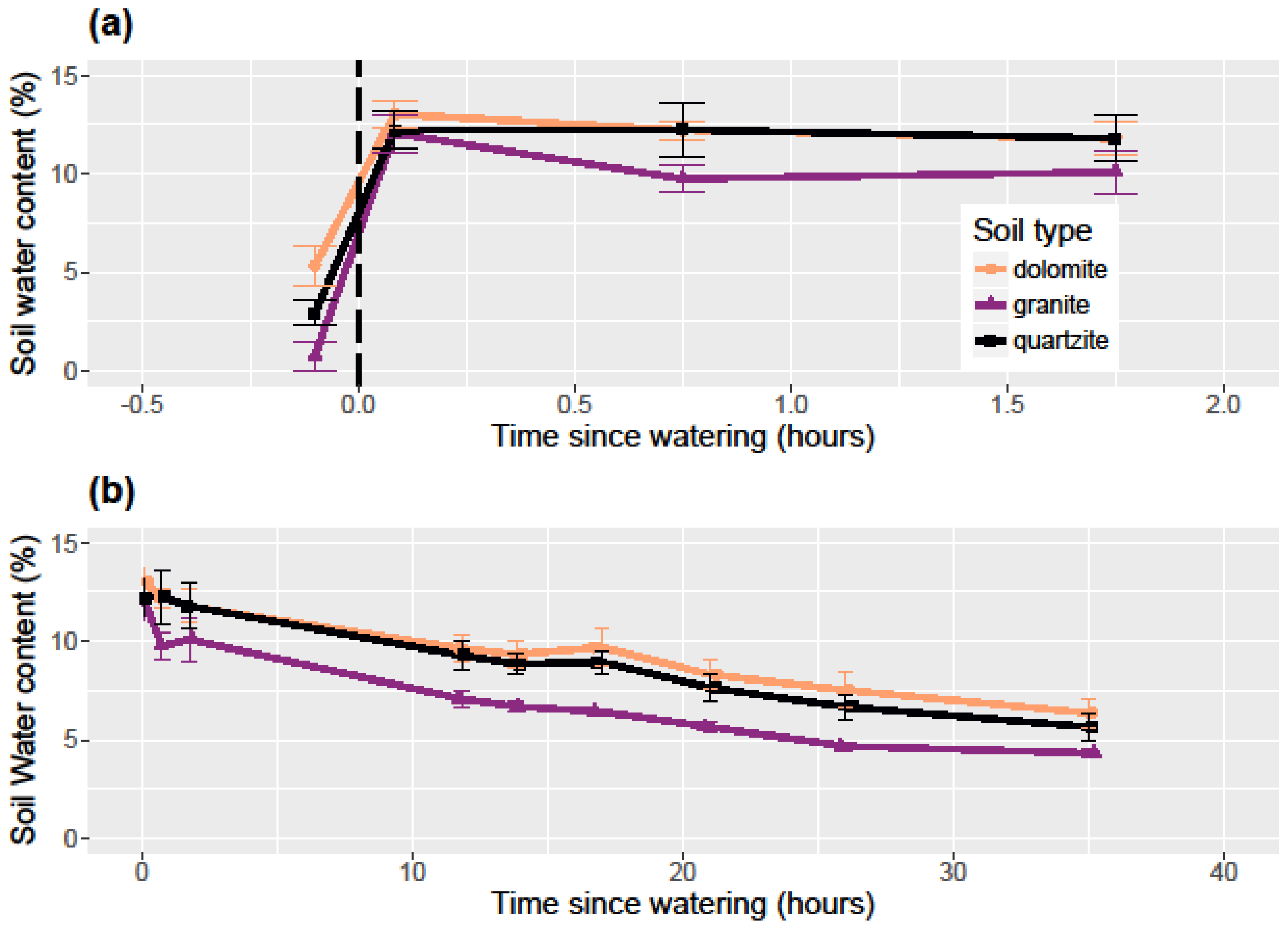
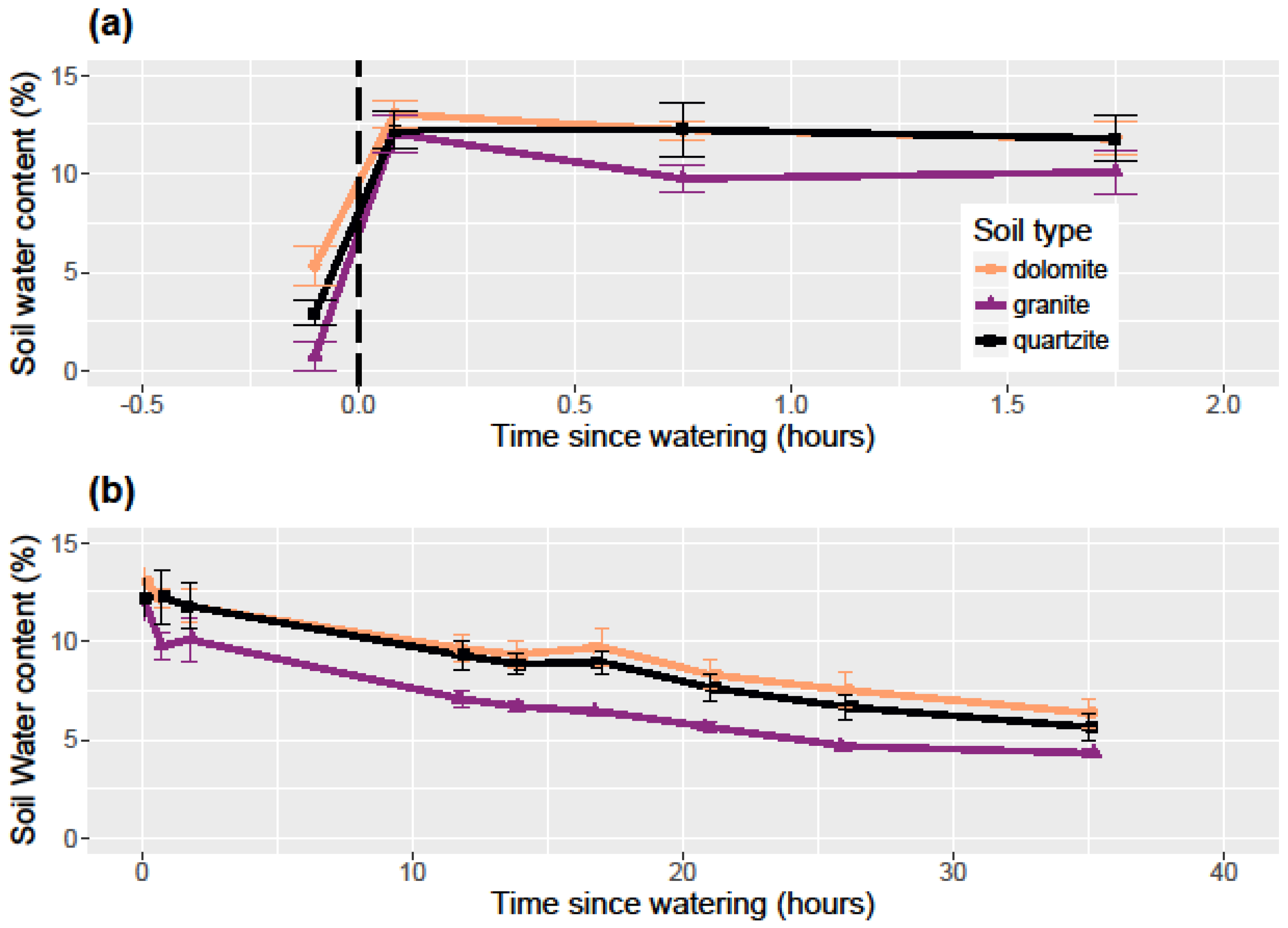
To examine the differential ability of each soil type to retain water, I measured the pre-watered soil moisture in each container and then thoroughly watered each container to saturation on July 1, 2017. July generally represents the start of the warmest and driest period in the growing season when seedlings are at their most vulnerable and when water availability is at its most limiting. I measured soil water content immediately after watering and for approximately the next 2 days at the following intervals: 45 min, 1:45, 11:50, 13:50, 17:00, 21:00, 26:00, 35:00, and 46:00. I watered at 7:30 p.m. to approximate a late-evening thunderstorm which is a somewhat common weather event in this area. All soil water content measurements were taken with a FieldScout TDR-100 Soil Moisture Meter (Spectrum Technologies, Inc., Aurora, IL, USA) with 12 cm probes. The FieldScout TDR measures volumetric water content integrated over the length of the probes and so gives average water content over the entire 12 cm probe length. The TDR was calibrated with deionized (DI) water for the 12 cm probes immediately prior to the experiment. Since a brief natural rain shower occurred at around 45:00 from the experimental watering, analysis excludes this last measurement and the experiment was ended.
2.3. Seed Germination and Seedling Survival
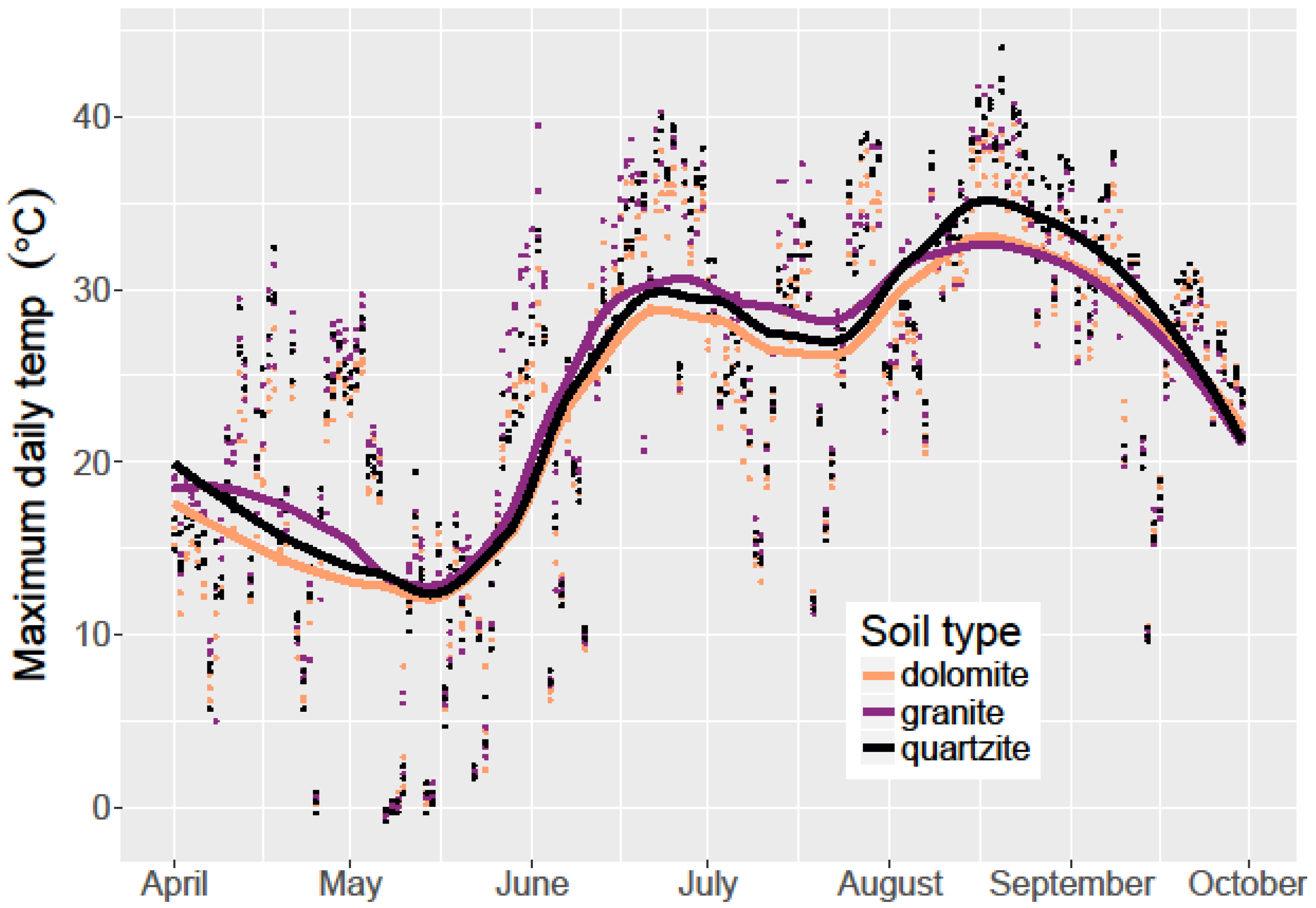
I sowed limber pine seeds in three treatments in October 2014: on the soil surface, under a fist-sized rock, and 5 cm below the soil surface. Seeds were planted in the fall to allow for natural cold stratification, a requirement for these seeds to germinate. In 9 randomly selected containers (three per soil type), I placed ibutton thermochrons (Maxim Integrated) to record soil surface temperature every 4 h. I placed the thermochrons inside a white polyvinyl chloride (PVC) joint for shielding of direct sun and to allow protection from water. Each row of containers was covered with a 1 cm2 wire mesh to exclude seed predators.
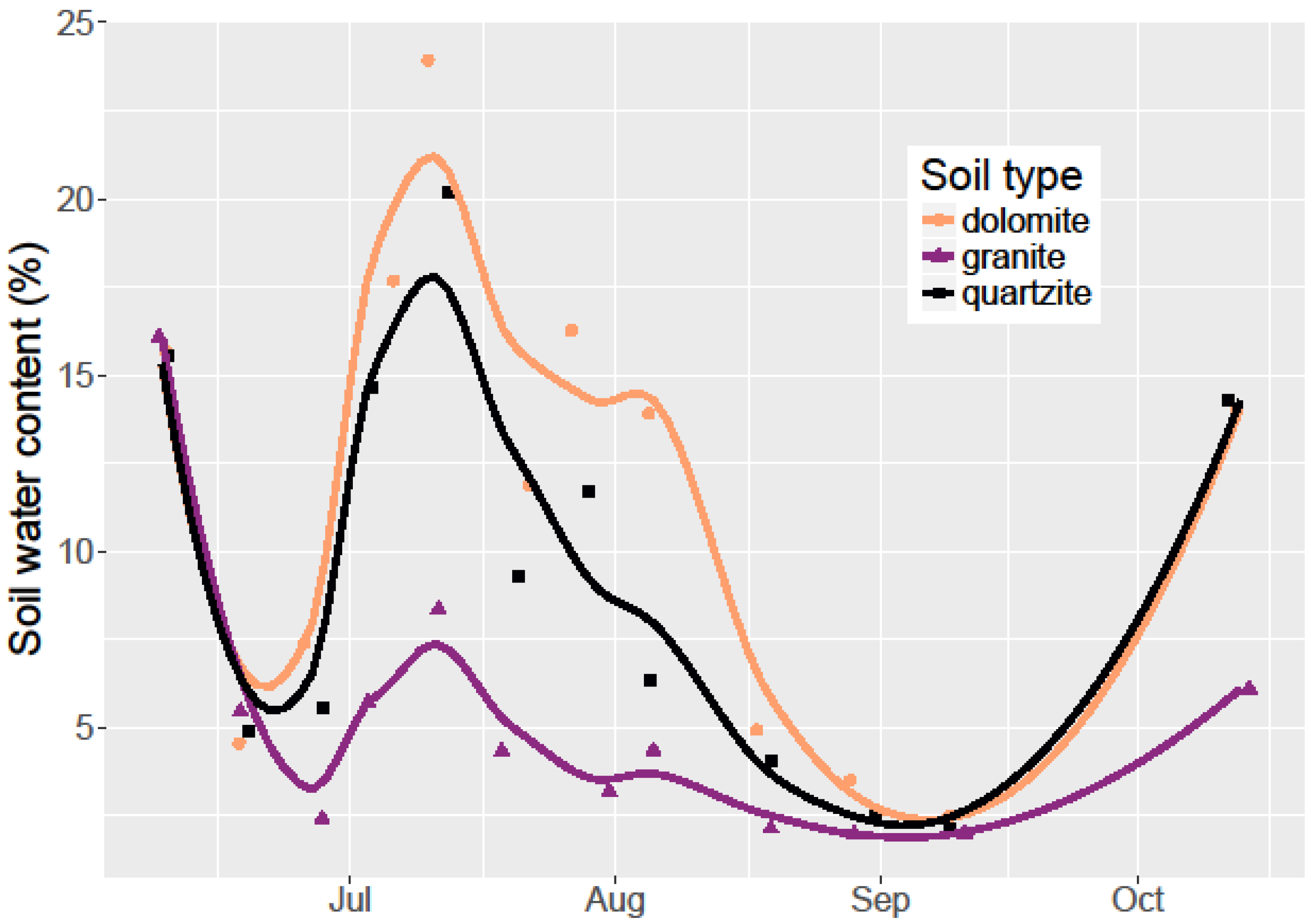
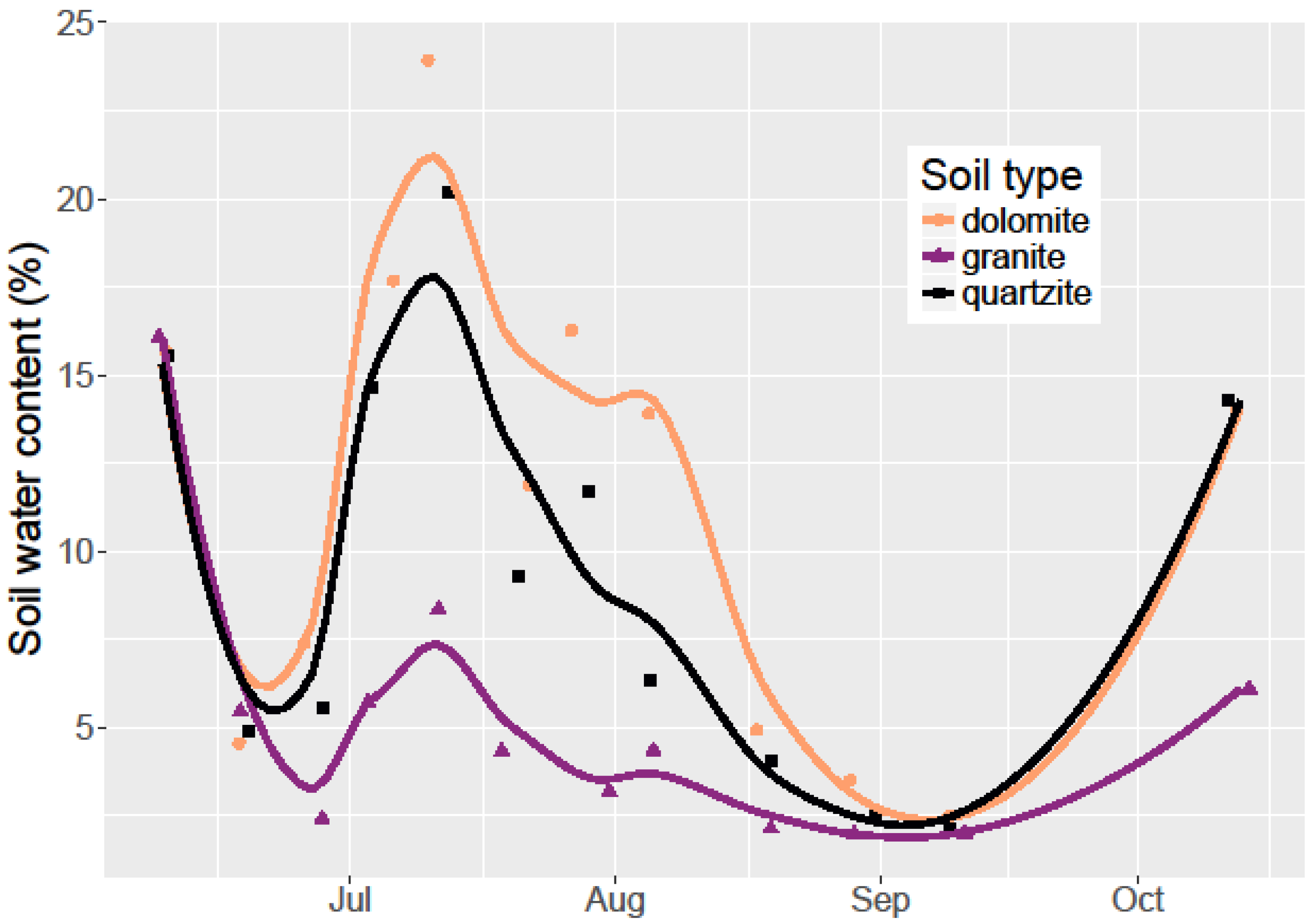
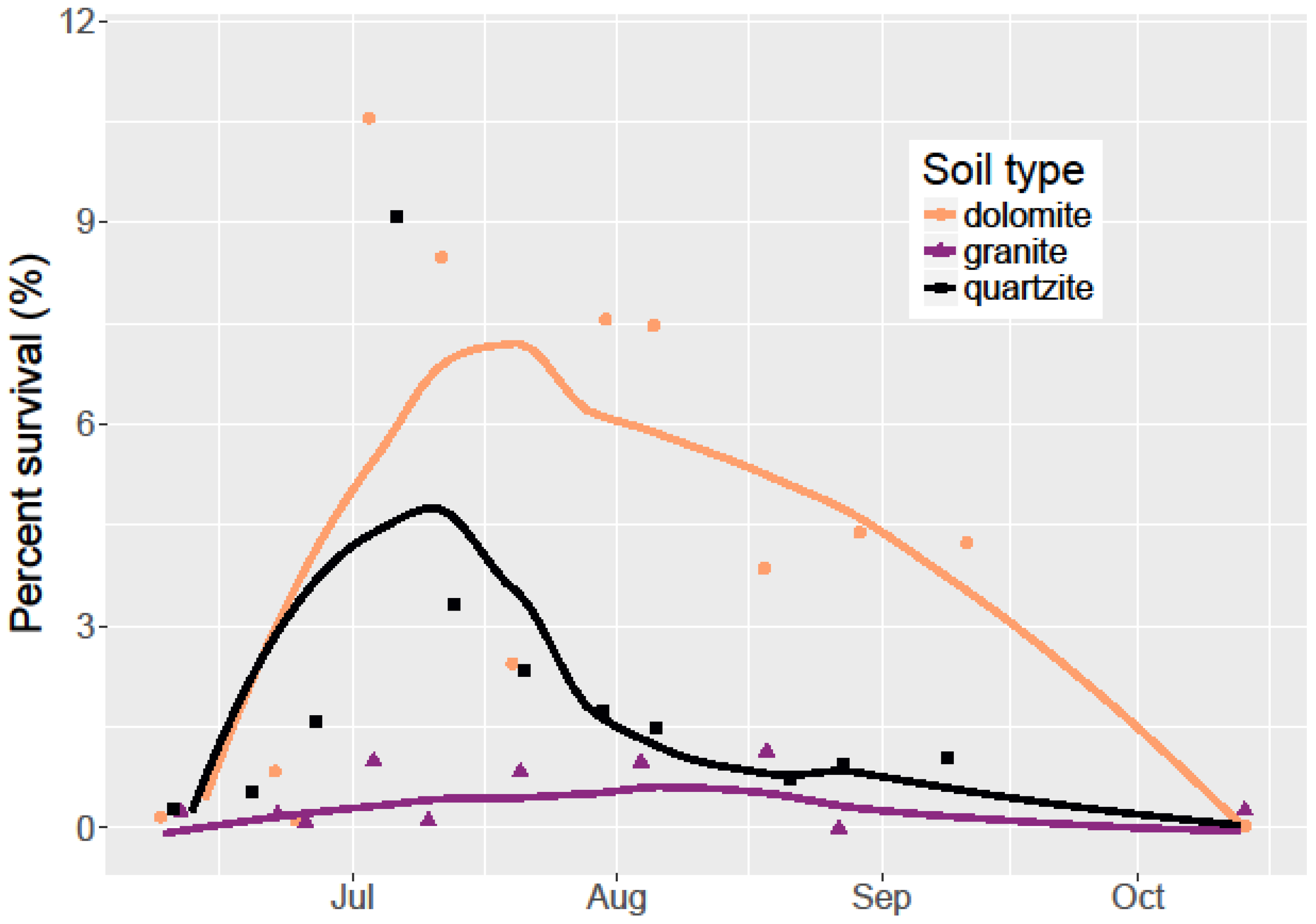
I monitored seed germination and survival by counting emerged seedlings on a weekly basis throughout the 2015 field season (June–October). Since all seeds that germinated did so by mid-July before any seedling mortality, a simple count of living seedlings was sufficient to account for both germination and seedling survival. While monitoring seedling survival, I also measured the soil water content using a FieldScout TDR-100 Soil Moisture Meter with calibrated 12 cm probes. I collected water content measurements weekly in each container.
2.4. Data Analysis
In the soil water retention experiment, I used a one-way ANOVA to examine the differences in each soil type’s relative ability to retain water after experimental watering. At each time step, I performed an ANOVA among the soil types with soil water content being the response variable. Where significance was found, I used Tukey’s honest significant difference (HSD) post-hoc tests to determine which pairs of soil types had statistically different soil water content. To examine the effect of soil type on soil water content and temperature for the 2015 growing season, I used linear regression models.
I also used linear regression models to examine the effects of temperature and water volume on seedling germination and survival. I defined germination as the period starting before seeds had germinated and the period after seedling emergence stopped in mid-July. I defined August 31st as the end of the period denoting survival and so any seedlings which lived until that date were determined to have “survived.” While this does not mark the end of mortality, cooler temperatures and increased precipitation in September resulted in another cohort of germination confounding the survival signal. To examine the effect of soil type on seedling survival over the growing season, I used an ANOVA and Tukey’s HSD tests. All analyses were done in the
R Project for Statistical Computing [
26].
4. Discussion
Limber pine germination rates were low at 9.2% overall. These same seeds planted in a greenhouse study after laboratory cold stratification germinated at an 86% rate (unpublished data). Despite these low germination rates, I found limber pine survival to be higher on dolomite soil than in quartzite soil and much higher than on granitic soil. Among the three soil types, dolomite soil had the highest soil water retention and lowest temperatures during the hottest part of the growing season. This combination resulted in dolomite soil having the most available water for an establishing seedling. Dolomite soil is associated with cooler temperatures and higher water content, but given the lack of a significant relationship between temperature and seedling survival, and the meager relationship between water content and seedling survival, there must be other dolomite soil factors that are beneficial to seedling germination and survival not measured here. Alternatively, it is possible that at higher overall soil water contents and germination rates, the relationship between soil water content and germination would be stronger. The lack of a significant effect of soil type on germination points to the relative equality in the soil type environment during the early season when conditions are cooler and wetter.
When ecologists first turned their attention to the White Mountains in the 1960s, they noted vastly different plant communities on the various soil types, especially the dominance of bristlecone pine forest on dolomite above about 3000 m (10,000 ft) elevation [
22,
27]. This dominance was initially assumed to be due to the harsh soil being inhospitable to potential competitors, especially sagebrush (
Artemisia sp.) and limber pine [
22]. The assumption followed that bristlecone pine might thrive on other soil types (granite and quartzite) but, since it is out-competed there, it is relegated to poorer dolomite soils. Recent research showed that bristlecone pine prefers dolomite soil to quartzite and granite [
21], indicating that bristlecone pine’s presence on dolomite soils is a function of individuals growing better there than elsewhere and not necessarily due to the lack of competition.
This study examines whether other species, specifically limber pine, are actually excluded on dolomite soil. These results show that despite the high pH, high magnesium, and low phosphorus properties of dolomite soils (
Table 1), there must be at least some positive attributes of dolomite for limber pine establishment, namely water holding capacity and temperature, which may trump the negative aspects of dolomite. In recent studies of western North American sub-alpine forests establishing at and above treeline, drought stress was found to be a very strong limiter of establishment, especially for limber pine [
15,
16,
17,
18]. At least in the first year of life, the positive attributes of dolomite translate into positive effects on limber pine germination and establishment. Limber pine seedlings appear to even prefer dolomite to granite and quartzite, two soil types where limber pine adults are more commonly found.
There are some important limitations to this study that may affect the interpretation of these results. While this study was carried out within the elevational and geographical range of limber pine, this experiment is not strictly in situ. Due to the removal of rocks from the soil, soil texture heterogeneity was lost. There are likely differences in soil particle size heterogeneity among soil types that would provide differential germination and establishment. For example, while quartzite soil holds water well relative to granite and dolomite, much of the surface on quartzite soils is made of up of small and large rocks that would make germination difficult but also might provide some protection for establishing seedlings [
22]. In addition, all of the soil samples came from one location per soil type. This limits the ability to extrapolate these findings to other sites in the White Mountains and Great Basin, and limits inference from these results to those areas from which the soil was taken. None of the seedlings survived beyond the first year, so the results of this study are also limited to the earliest life stages. However, since germination and the seedling stage are likely the most limiting life stages [
20,
21], focusing on the limitations of first-year seedlings can still be informative about this important bottleneck. It is also possible that the negative effects of dolomite soil on limber pine are not apparent until later life stages in which limber pine may ultimately be excluded.
Given these limitations, the results of this study along with previous studies, suggest that dolomite is the preferred soil type for both limber pine and bristlecone pine in the earliest life stages [
19,
20,
21]. In a recent study, limber pine was found to be establishing in high density above treeline throughout the Great Basin, especially on dolomite soils where limber pine trees between the ages of 15–30 years of age were common [
20]. This suggests that dolomite soils continue to be a good growth medium for limber pine well after the first year. In these same treeline forests on dolomite, adult cone-bearing, limber pine adults are very rare. The question then becomes: If limber pine prefers dolomite soil for establishment in the early life stages and is common on newly available habitat upslope of treeline on dolomite soil, why are there so few adult limber pines in mature treeline forests on dolomite? It is possible that until very recently, cold temperatures limited limber pine in these high altitude forests, although there are numerous examples of treeline forests on granite soil in the Great Basin that are dominated by limber pine [
19,
20,
23]. On the other hand, treelines on granitic soil dominated by limber pine may be somewhat lower than treelines on quartzite and dolomite (unpublished data). Another possibility is that while bristlecone pine appears to be recruiting much more slowly above treeline than limber pine, given its famously long life spans [
28], bristlecone pine may be able to out-compete limber pine over much longer time scales. While individual bristlecone pines can live to at least 5000 years, 1500 year old limber pines have also been documented [
29]. If this is the case, ecologists would be forced to call the very slow-growing limber pine an early-succession species relative to the extremely slow-growing bristlecone pine.
These results point to the importance of soil type on early life stages, which has important ramifications on tree species range shifts in response to climate change. Whether a species is able to move to new climatically available habitat is a function not only of its dispersal, but also its ability to survive in the conditions, such as the soil type, where the climate is now suitable for the species. In montane systems, and especially sub-alpine forests, this is especially important as species move upslope to potentially novel soil types on generally decreasing surface areas. While both limber pine and bristlecone pine are likely to persist at higher elevations in response to climate change, how the species ranges move in relation to each other will be an interesting development to watch. However, we will have have to watch for a very long time since it will take thousands of years to see the end result.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}