Tree Regeneration Spatial Patterns in Ponderosa Pine Forests Following Stand-Replacing Fire: Influence of Topography and Neighbors


Abstract
:1. Introduction
2. Materials and Methods
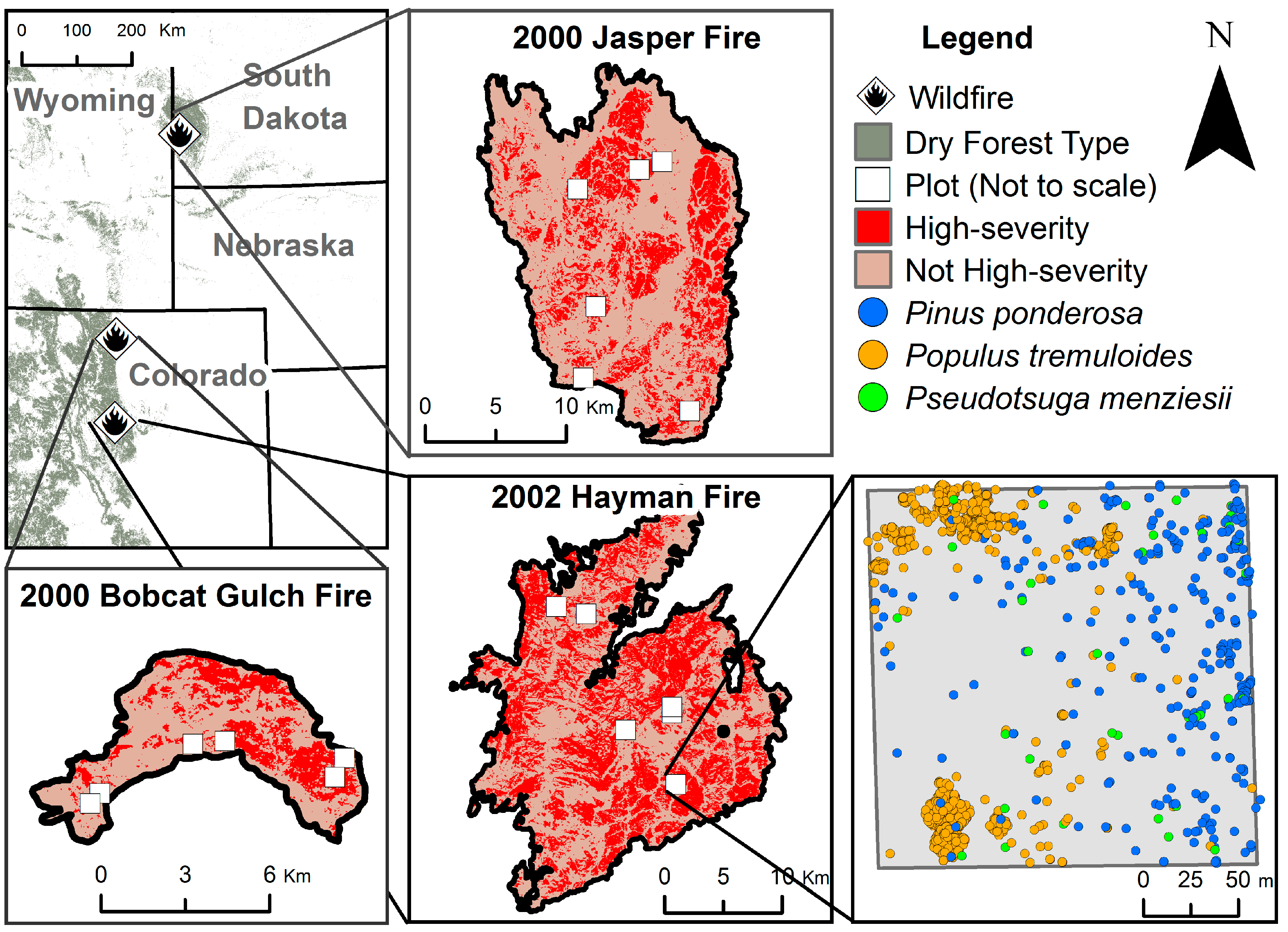
2.1. Data Collection
2.2. Patterns of Regenerating Tree Locations
2.3. Patterns of Regenerating Tree Heights
3. Results
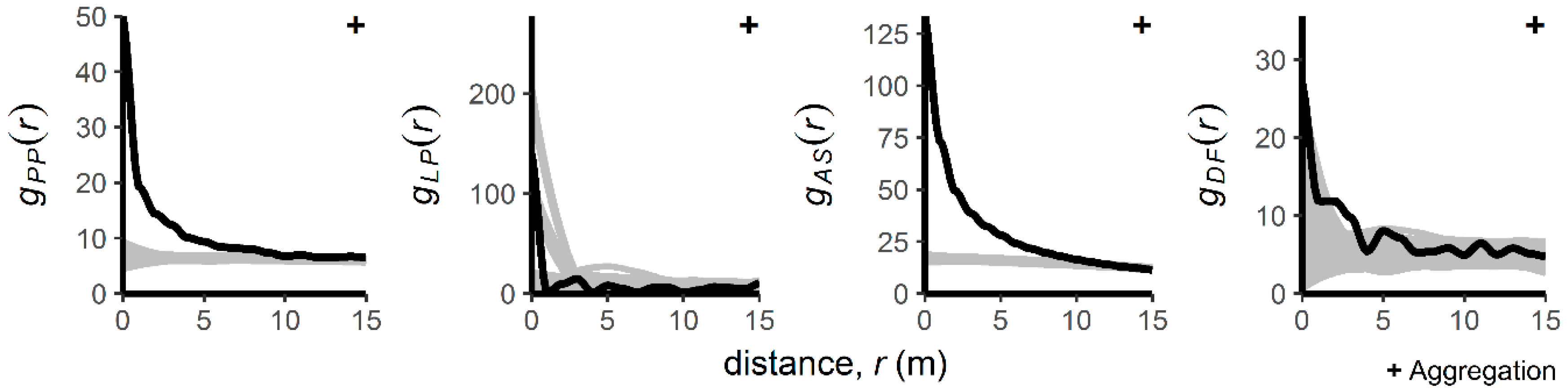
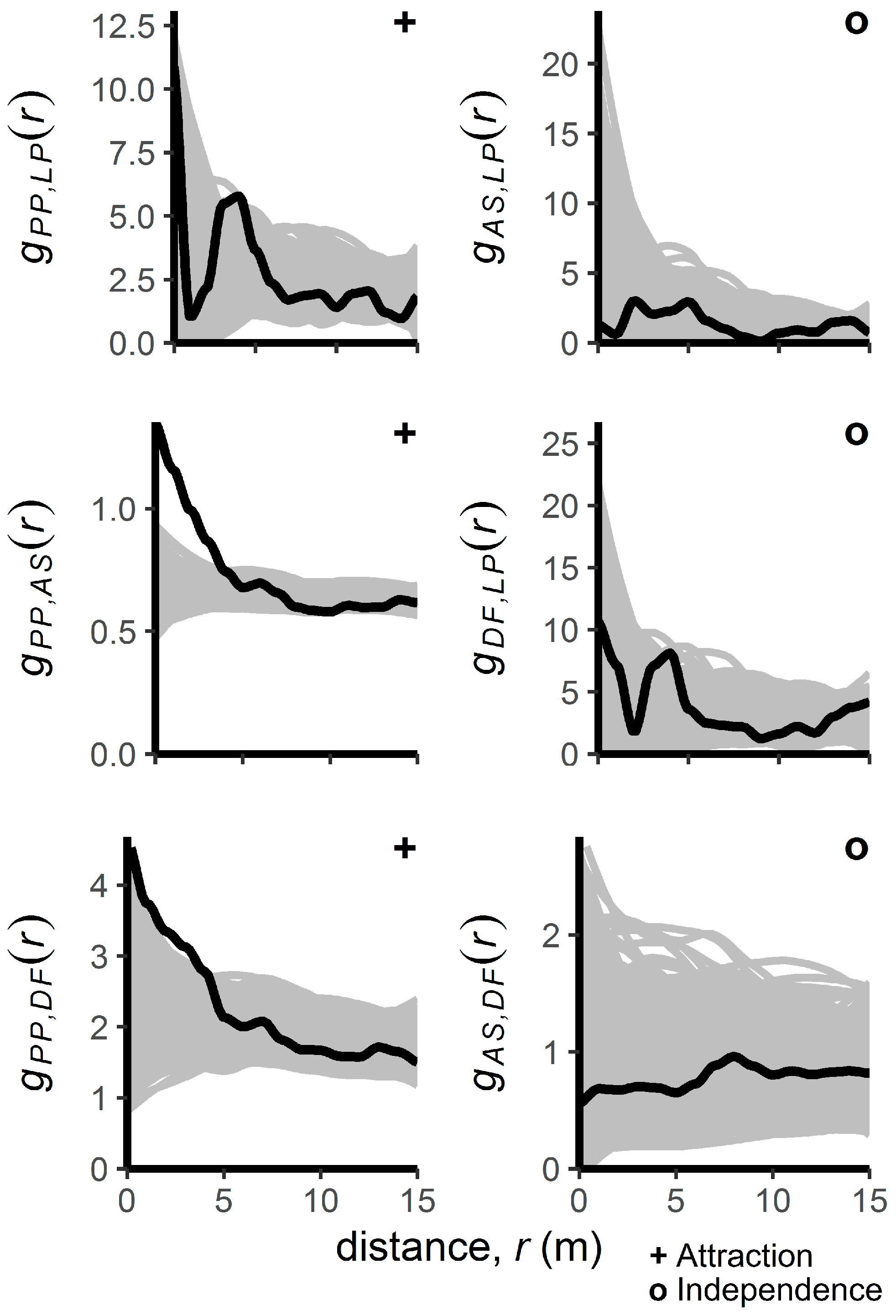
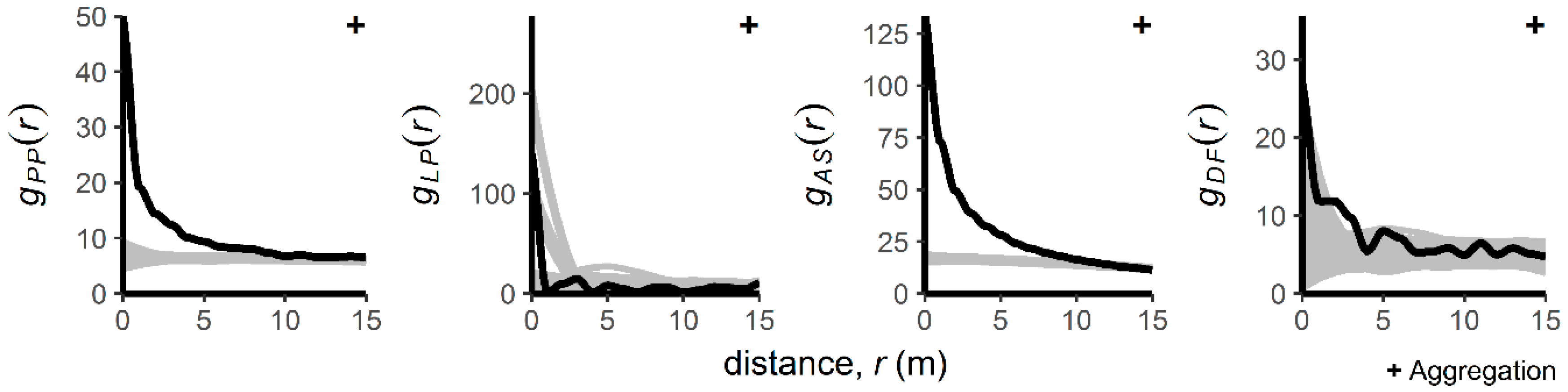
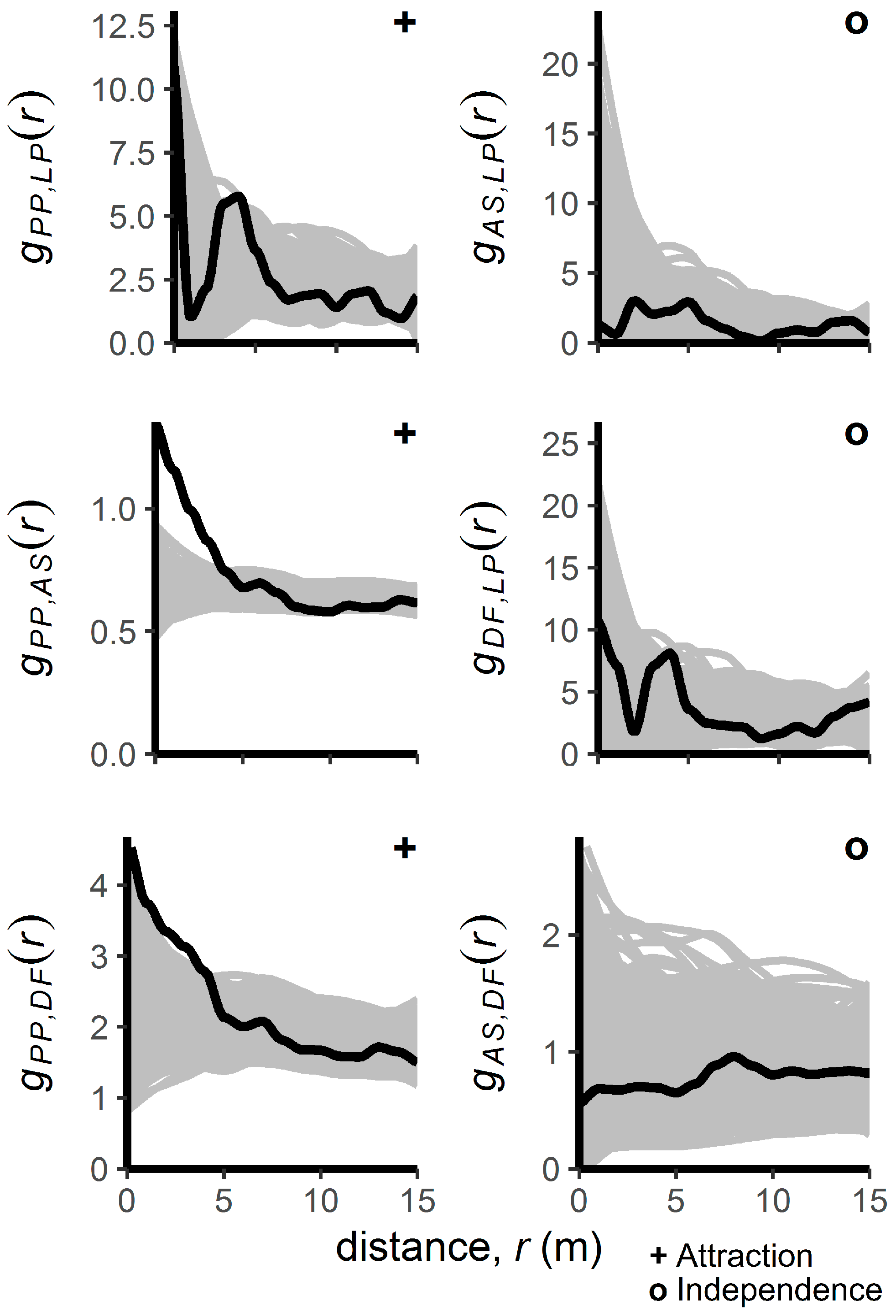
3.1. Patterns of Regenerating Tree Locations
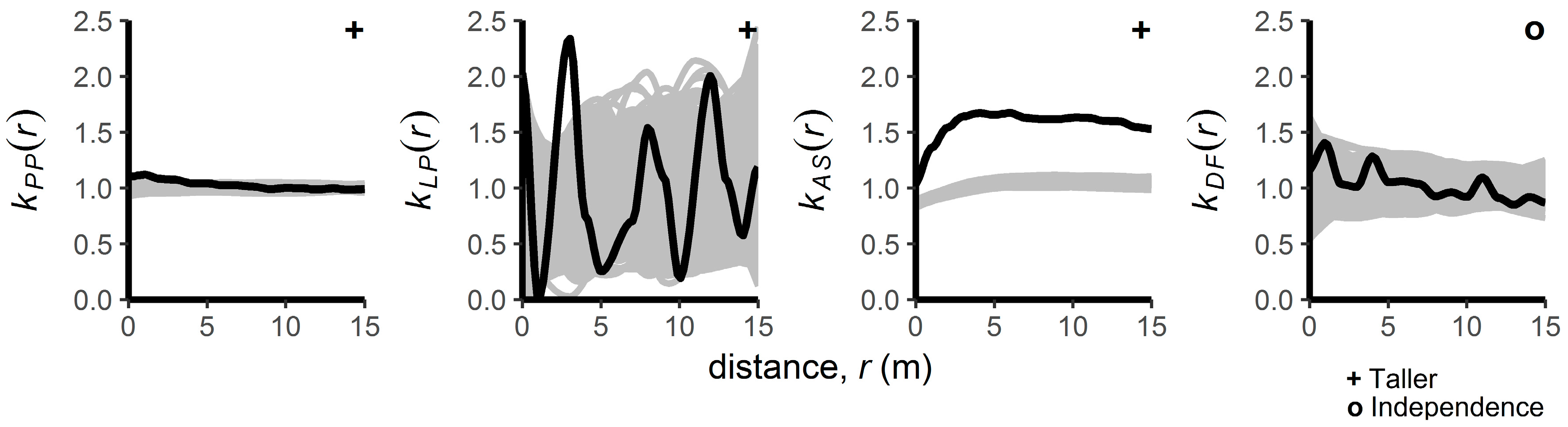
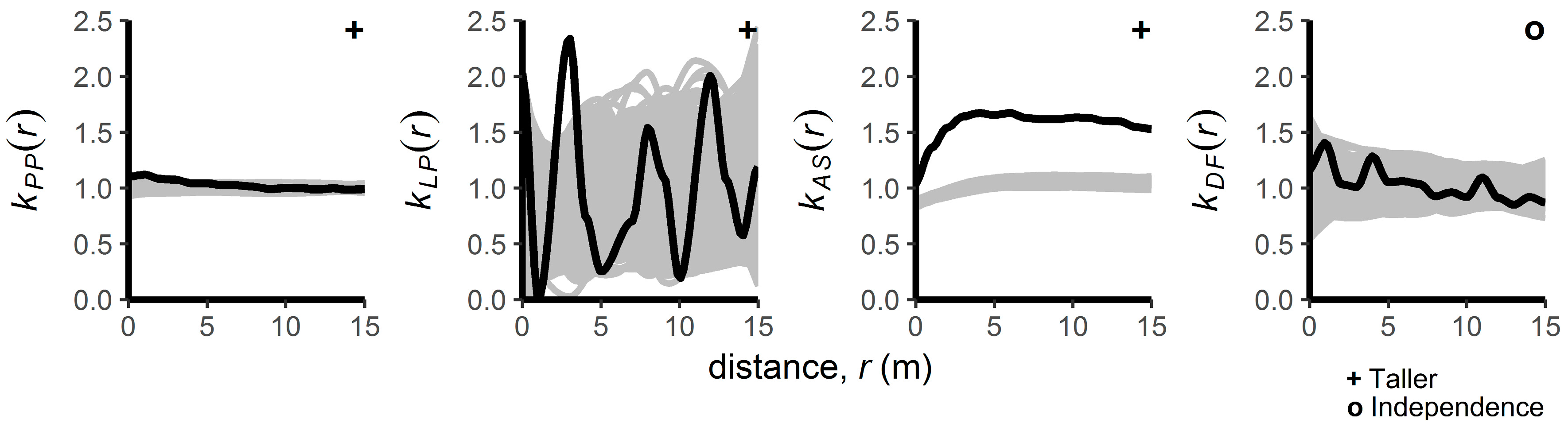
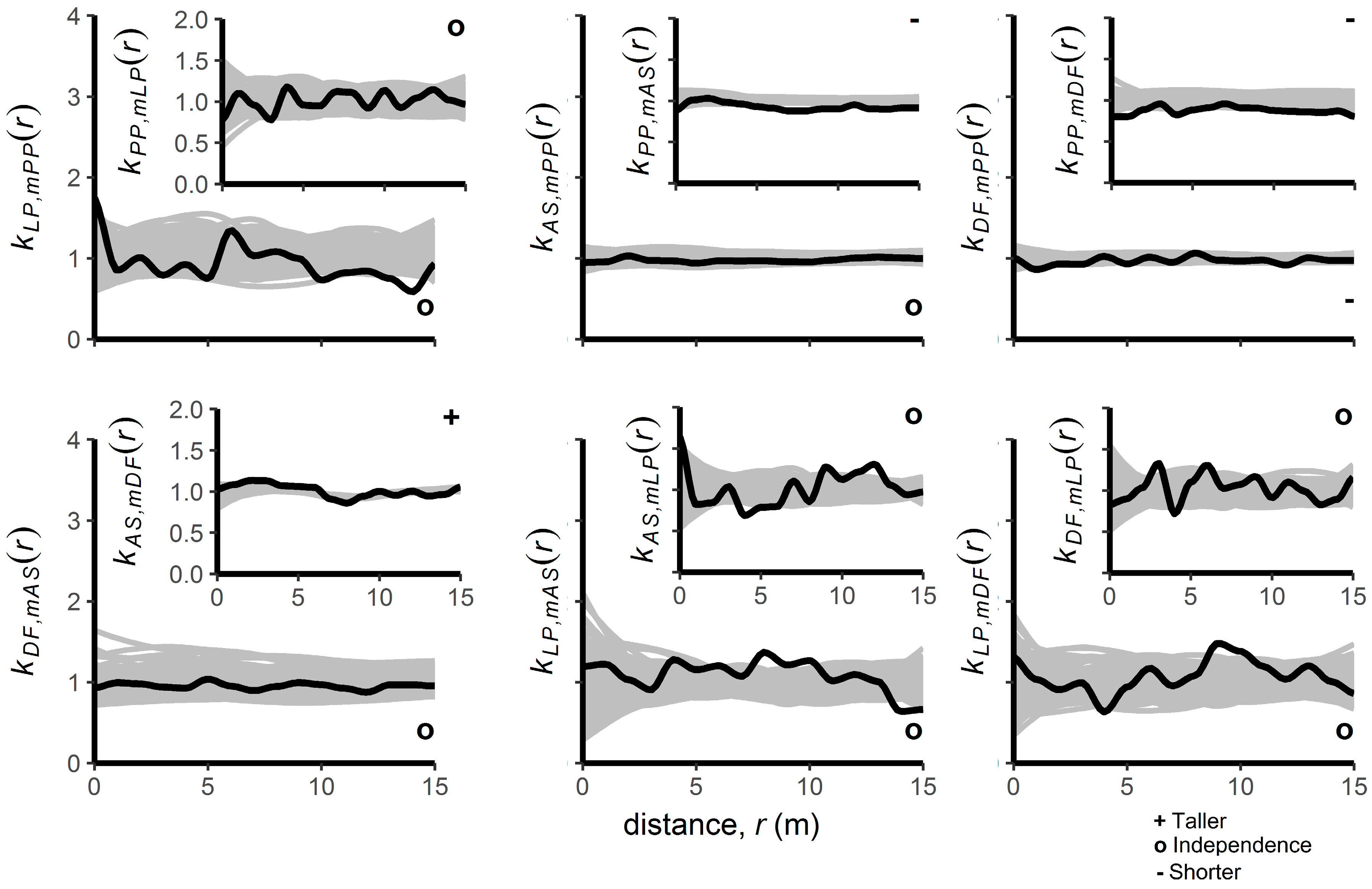
3.2. Patterns of Regenerating Tree Heights
4. Discussion
4.1. Intraspecific Spatial Patterns
4.2. Interspecific Spatial Patterns
4.3. Limitations and Future Research Directions
4.4. Ecological and Management Implications
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Stoyan, D.; Penttinen, A. Recent applications of point process methods in forestry statistics. Stat. Sci. 2000, 15, 61–78. [Google Scholar] [CrossRef]
- Larson, A.J.; Churchill, D. Tree spatial patterns in fire-frequent forests of western North America, including mechanisms of pattern formation and implications for designing fuel reduction and restoration treatments. For. Ecol. Manag. 2012, 267, 74–92. [Google Scholar] [CrossRef]
- Pearson, G. Management of Ponderosa Pine in the Southwest; Monograph 4; USDA Forest Service: Washington, DC, USA, 1950; p. 114.
- Boyden, S.; Binkley, D.; Shepperd, W. Spatial and temporal patterns in structure, regeneration, and mortality of an old-growth ponderosa pine forest in the Colorado Front Range. For. Ecol. Manag. 2005, 219, 43–55. [Google Scholar] [CrossRef]
- Fajardo, A.; Goodburn, J.M.; Graham, J. Spatial patterns of regeneration in managed uneven-aged ponderosa pine/Douglas-fir forests of Western Montana, USA. For. Ecol. Manag. 2006, 223, 255–266. [Google Scholar] [CrossRef]
- Ziegler, J.P.; Hoffman, C.; Battaglia, M.; Mell, W. Spatially explicit measurements of forest structure and fire behavior following restoration treatments in dry forests. For. Ecol. Manag. 2017, 386, 1–12. [Google Scholar] [CrossRef]
- Bače, R.; Svoboda, M.; Janda, P.; Morrissey, R.C.; Wild, J.; Clear, J.L.; Čada, V.; Donato, D.C.; Chen, H.Y.H. Legacy of pre-disturbance spatial pattern determines early structural diversity following severe disturbance in montane spruce forests. PLoS ONE 2015, 10, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Donato, D.C.; Campbell, J.L.; Franklin, J.F. Multiple successional pathways and precocity in forest development: Can some forests be born complex? J. Veg. Sci. 2012, 23, 576–584. [Google Scholar] [CrossRef]
- Hansen, A.J.; Spies, F.J.; Swanson, F.J.; Ohmann, J.L. Conserving biodiversity in managed forests: A lesson from natural forests. Bioscience 1991, 41, 382–392. [Google Scholar] [CrossRef]
- Tepley, A.J.; Swanson, F.J.; Spies, T.A. Post-fire tree establishment and early cohort development in conifer forests of the western Cascades of Oregon, USA. Ecosphere 2014, 5, 1–23. [Google Scholar] [CrossRef]
- Stephens, S.L.; Burrows, N.; Buyantuyev, A.; Gray, R.W.; Keane, R.E.; Kubian, R.; Liu, S.; Seijo, F.; Shu, L.; Tolhurst, K.G.; et al. Temperate and boreal forest mega-fires: Characteristics and challenges. Front. Ecol. Environ. 2014, 12, 115–122. [Google Scholar] [CrossRef]
- Hessburg, P.F.; Spies, T.A.; Perry, D.A.; Skinner, C.N.; Taylor, A.H.; Brown, P.M.; Stephens, S.L.; Larson, A.J.; Churchill, D.J.; Povak, N.A.; et al. Tamm Review: Management of mixed-severity fire regime forests in Oregon, Washington, and Northern California. For. Ecol. Manag. 2016, 366, 221–250. [Google Scholar] [CrossRef]
- Brown, T.J.; Hall, B.L.; Westerling, A.L. The impact of twenty-first century climate change on wildland fire danger in the western United States: An applications perspective. Clim. Chang. 2004, 62, 365–388. [Google Scholar] [CrossRef]
- Chambers, M.E.; Fornwalt, P.J.; Malone, S.L.; Battaglia, M.A. Patterns of conifer regeneration following high severity wildfire in ponderosa pine-dominated forests of the Colorado Front Range. For. Ecol. Manag. 2016, 378, 57–67. [Google Scholar] [CrossRef]
- Rother, M.T.; Veblen, T.T. Limited conifer regeneration following wildfires in dry ponderosa pine forests of the Colorado Front Range. Ecosphere 2016, 7. [Google Scholar] [CrossRef]
- DellaSala, D.A.; Bond, M.L.; Hanson, C.T.; Hutto, R.L.; Odion, D.C. Complex early seral forests of the Sierra Nevada: What are they and how can they be managed for ecological integrity? Nat. Areas J. 2014, 34, 310–324. [Google Scholar] [CrossRef]
- Pielou, E.C. The use of plant-to-neighbour distances for the detection of competition. J. Ecol. 1962, 50, 357–367. [Google Scholar] [CrossRef]
- Das, A.; Battles, J.; Stephenson, N.L.; Van Mantgem, P.J. The contribution of competition to tree mortality in old-growth coniferous forests. For. Ecol. Manag. 2011, 261, 1203–1213. [Google Scholar] [CrossRef]
- Redmond, M.D.; Cobb, N.S.; Clifford, M.J.; Barger, N.N. Woodland recovery following drought-induced tree mortality across an environmental stress gradient. Glob. Chang. Biol. 2015, 21, 3685–3695. [Google Scholar] [CrossRef] [PubMed]
- Getzin, S.; Wiegand, T.; Wiegand, K.; He, F. Heterogeneity influences spatial patterns and demographics in forest stands. J. Ecol. 2008, 96, 807–820. [Google Scholar] [CrossRef]
- Bonnet, V.H.; Schoettle, A.W.; Shepperd, W.D. Postfire environmental conditions influence the spatial pattern of regeneration for Pinus ponderosa. Can. J. For. Res. 2005, 35, 37–47. [Google Scholar] [CrossRef]
- Urza, A.K.; Sibold, J.S. Climate and seed availability initiate alternate post-fire trajectories in a lower subalpine forest. J. Veg. Sci. 2017, 28, 1–14. [Google Scholar] [CrossRef]
- Vander Wall, S.B. Effects of seed size of wind-dispersed pines (Pinus) on secondary seed dispersal and the caching behavior of rodents. Oikos 2003, 100, 25–34. [Google Scholar] [CrossRef]
- Alexander, R.R. Establishment of Lodgepole Pine Reproduction after Different Slash Disposal Treatments; Research Note RMRS-RN-62; USDA Forest Service: Fort Collins, CO, USA, 1966; p. 4.
- Shepperd, W.D. Initial Growth, Development, and Clonal Dynamics of Regenerated Aspen in the Rocky-Mountains; Research Paper RMRS-RP-312; USDA Forest Service: Fort Collins, CO, USA, 1993; p. 8.
- PRISM Climate Group. Available online: prism.oregonstate.edu (accessed on 1 December 2016).
- Peet, R.K. Forest vegetation of the Colorado Front Range. Vegetatio 1981, 45, 3–75. [Google Scholar] [CrossRef]
- Brown, P.M.; Sieg, C.H. Historical variability in fire at the ponderosa pine—Northern Great Plains prairie ecotone, southeastern Black Hills, South Dakota. Ecoscience 1999, 6, 539–547. [Google Scholar] [CrossRef]
- Brown, P.M.; Kaufmann, M.R.; Shepperd, W.D. Long-term, landscape patterns of past fire events in a montain ponderosa pine forest of central Colorado. Landsc. Ecol. 1999, 14, 513–532. [Google Scholar] [CrossRef]
- Fornwalt, P.J.; Huckaby, L.S.; Alton, S.K.; Kaufmann, M.R.; Brown, P.M.; Cheng, A.S. Did the 2002 Hayman Fire, Colorado, USA burn with uncharacteristic severity? Fire Ecol. 2016, 12, 117–132. [Google Scholar] [CrossRef]
- Fule, P.Z.; Swetnam, T.W.; Brown, P.M.; Falk, D.A.; Peterson, D.L.; Allen, C.D.; Aplet, G.H.; Battaglia, M.A.; Binkley, D.; Farris, C.; et al. Unsupported inferences of high-severity fire in historical dry forests of the western United States: Response to Williams and Baker. Glob. Ecol. Biogeogr. 2014, 23, 825–830. [Google Scholar] [CrossRef]
- Shepperd, W.D.; Edminster, C.B.; Mata, S.A. Long-term seedfall, establishment, survival, and growth of natural and planted ponderosa pine in the Colorado Front Range. West. J. Appl. For. 2006, 21, 19–26. [Google Scholar]
- The National Map. Available online: https://nationalmap.gov (accessed on 1 December 2016).
- Tagil, S.; Jenness, J. GIS-based automated landform classification and topographic, landcover and geologic attributes of landforms around the Yazoren Polje, Turkey. J. Appl. Sci. 2008, 8, 910–921. [Google Scholar] [CrossRef]
- Cushman, S.A.; Gutzweiler, K.; Evans, J.S.; McGarigal, K. The Gradient Paradigm: A conceptual and analytical framework for landscape ecology. In Spatial Complexity, Informatics, and Wildlife Conservation; Huettmann, F., Ed.; Springs: New York, NY, USA, 2010; pp. 83–108. [Google Scholar]
- Wiegand, T.; Moloney, K.A. Handbook of Spatial Point-Pattern Analysis in Ecology; Chapman and Hall/CRC Press: Boca Raton, FL, USA, 2014; ISBN 9781420082555. [Google Scholar]
- Baddeley, A.; Rubak, E.; Turner, R. Spatial Point Patterns: Methodology and Applications with R; Chapman and Hall/CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Goreaud, F.; Pelissier, R. Avoiding misinterpretation of biotic interactions with the intertype population independence vs. random labelling hypotheses. J. Veg. Sci. 2012, 14, 681–692. [Google Scholar] [CrossRef]
- Suzuki, S.N.; Kachi, N.; Suzuki, J.I. Development of a local size hierarchy causes regular spacing of trees in an even-aged Abies forest: Analyses using spatial autocorrelation and the mark correlation function. Ann. Bot. 2008, 102, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Donato, D.C.; Fontaine, J.B.; Campbell, J.L.; Robinson, W.D.; Kauffman, J.B.; Law, B.E. Conifer regeneration in stand-replacement portions of a large mixed-severity wildfire in the Klamath–Siskiyou Mountains. Can. J. For. Res. 2009, 39, 823–838. [Google Scholar] [CrossRef]
- Pauchard, A.; Escudero, A.; Garcia, R.A.; de la Cruz, M.; Langdon, B.; Cavieres, L.A.; Esquivel, J. Pine invasions in treeless environments: Dispersal overruns microsite heterogeneity. Ecol. Evol. 2016, 6, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.H.; Uria-Diez, J.; Wiegand, K. Spatial distribution and association patterns in a tropical evergreen broad-leaved forest of north-central Vietnam. J. Veg. Sci. 2016, 27, 318–327. [Google Scholar] [CrossRef]
- Oliver, W.W.; Ryker, R.A. Pinus ponderosa Dougl. ex Laws. In Silvics of North America, Volume 1, Conifers; Agricultural Handbook 654; Burns, R.M., Honkala, B.H., Eds.; USDA Forest Service: Washington, DC, USA, 1990; pp. 413–424. [Google Scholar]
- Stoll, P.; Prati, D. Intraspecific aggregation alters competitive interactions in experimental plant communities. Ecology 2001, 82, 319–327. [Google Scholar] [CrossRef]
- Losapio, G.; Cruz, M.D.; Escudero, A.; Schmid, B.; Schöb, C. Positive interactions support the formation of complex spatial networks. bioRxiv 2017, 1–21. [Google Scholar] [CrossRef]
- Ehle, D.S.; Baker, W.L. Disturbance and stand dynamics in ponderosa pine forests in Rocky Mountain National Park, USA. Ecol. Monogr. 2003, 73, 543–566. [Google Scholar] [CrossRef]
- Weiner, J.; Solbrig, O.T. The meaning and measurement of size hierarchies in plant populations. Oecologia 1984, 61, 334–336. [Google Scholar] [CrossRef] [PubMed]
- Pielou, E.C. A single mechanism to account for regular, random and aggregated populations. J. Ecol. 1960, 48, 575–584. [Google Scholar] [CrossRef]
- Callaway, R.M. Positive Interactions and Interdependence in Plant Communities; Springer: Dordrecht, The Netherlands, 2007; ISBN 9781402062247. [Google Scholar]
- Wild, J.; Kopecký, M.; Svoboda, M.; Zenáhlíková, J.; Edwards-Jonášová, M.; Herben, T. Spatial patterns with memory: Tree regeneration after stand-replacing disturbance in Picea abies mountain forests. J. Veg. Sci. 2014, 25, 1327–1340. [Google Scholar] [CrossRef]
- Vakili, E.; Hoffman, C.M.; Keane, R.E.; Tinkham, W.T.; Dickinson, Y. Spatial variability of surface fuels in treated and untreated ponderosa pine forests of the southern Rocky Mountains. Int. J. Wildl. Fire 2016. [Google Scholar] [CrossRef]
- Holmgren, M.; Scheffer, M. Strong facilitation in mild environments: The stress gradient hypothesis revisited. J. Ecol. 2010, 98, 1269–1275. [Google Scholar] [CrossRef]
- Wright, A.; Schnitzer, S.A.; Reich, P.B. Living close to your neighbors: The importance of both competition and facilitation in plant communities. Ecology 2014, 95, 2213–2223. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.J.; Squierst, E.R. An unexpected change in spatial pattern across 10 years in an aspen-white pine forest. J. Ecol. 1995, 83, 847–855. [Google Scholar] [CrossRef]
- Brown, P.M.; Cook, B. Early settlement forest structure in Black Hills ponderosa pine forests. For. Ecol. Manag. 2006, 223, 284–290. [Google Scholar] [CrossRef]
- Brown, P.M.; Battaglia, M.A.; Fornwalt, P.J.; Gannon, B.; Huckaby, L.S.; Julian, C.; Cheng, A.S. Historical (1860) forest structure in ponderosa pine forests of the northern Front Range, Colorado. Can. J. For. Res. 2015, 45, 1462–1473. [Google Scholar] [CrossRef]
- Turner, M.G.; Romme, W.H.; Tinker, D.B. Surprises and lessons from the 1988 Yellowstone fires. Front. Ecol. Environ. 2003, 1, 351–358. [Google Scholar] [CrossRef]
- Beschta, R.L.; Rhodes, J.J.; Kauffman, J.B.; Gresswell, R.E.; Minshall, G.W.; Karr, J.R.; Perry, D.A.; Hauer, F.R.; Frissell, C.A. Postfire management on forested public lands of the western United States. Conserv. Biol. 2004, 18, 957–967. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Statistic | Density (Trees ha−1) * | Height (m) | Distance (m) * | Aspect (Unitless) | Slope (%) | TPI |
|---|---|---|---|---|---|---|
| Ponderosa pine (Pinus ponderosa) | ||||||
| Mean | 8.6 | 0.8 | 76.2 | 0.8 | 9.3 | 0.1 |
| Std. dev. | 14.5 | 0.5 | 134.3 | 0.8 | 8.7 | 0.4 |
| Range | 1.0–260.0 | 0.1–3.0 | 0.0–758.8 | 0.0–2.0 | 1.1–51.6 | −2.1–3.2 |
| Lodgepole pine (Pinus contorta) | ||||||
| Mean | 0.7 | 1.2 | 150.6 | 1.6 | 39.2 | 0.2 |
| Std. dev. | 0.5 | 0.6 | 119.4 | 0.5 | 8.3 | 0.7 |
| Range | 0.0–5.8 | 0.3–3.0 | 17.7–457.9 | 0.1–2.0 | 16.6–54.4 | −1.5–1.8 |
| Quaking aspen (Populus tremuloides) | ||||||
| Mean | 43.5 | 1.4 | 338.5 | 1.2 | 9.6 | −0.4 |
| Std. dev. | 66.1 | 1.0 | 194.5 | 0.8 | 7.4 | 0.8 |
| Range | 0.0–414.3 | 0.1–4.0 | 10.6–759.0 | 0.0–2.0 | 1.0–49.9 | −3.3–2.8 |
| Douglas-fir (Pseudotsuga menziesii) | ||||||
| Mean | 1.5 | 0.5 | 107.9 | 1.6 | 17.1 | 0.0 |
| Std. dev. | 2.1 | 0.3 | 85.1 | 0.6 | 12.9 | 0.6 |
| Range | 0.0–30.3 | 0.1–1.6 | 3.2–453.5 | 0.0–2.0 | 1.5–53.4 | −2.1–2.7 |
| All | ||||||
| Mean | 43.0 | 1.2 | 358.9 | 1.1 | 9.9 | −0.2 |
| Std. dev. | 61.2 | 0.9 | 205.1 | 0.8 | 8.5 | 0.8 |
| Covariate | Β | SE | t | p | Relative Magnitude |
|---|---|---|---|---|---|
| Ponderosa pine (Pinus ponderosa) | |||||
| (Int.) | −3.549 | 0.083 | −42.798 | <0.001 | |
| Distance | −0.006 | <0.001 | −17.769 | <0.001 | 119.744 |
| Aspect | −0.317 | 0.058 | −5.504 | <0.001 | 1.793 |
| Slope | −0.059 | 0.006 | −10.266 | <0.001 | 13.348 |
| TPI | −0.050 | 0.070 | −0.709 | 0.478 | 1.587 |
| Lodgepole pine (Pinus contorta) | |||||
| (Int.) | −12.536 | 1.478 | −8.483 | <0.001 | |
| Distance | −0.004 | 0.002 | −1.855 | 0.064 | 24.294 |
| Aspect | 0.669 | 0.500 | 1.339 | 0.181 | 3.811 |
| Slope | 0.102 | 0.029 | 3.474 | 0.001 | 630.665 |
| TPI | 0.110 | 0.327 | 0.336 | 0.737 | 2.706 |
| Quaking aspen (Populus tremuloides) | |||||
| (Int.) | −5.637 | 0.492 | −11.453 | <0.001 | |
| Distance | 0.001 | <0.001 | 3.656 | <0.001 | 2.220 |
| Aspect | 0.193 | 0.062 | 3.101 | 0.002 | 1.471 |
| Slope | −0.026 | 0.011 | −2.306 | 0.021 | 5.172 |
| TPI | −0.883 | 0.057 | −15.495 | <0.001 | 2955.452 |
| Douglas-fir (Pseudotsuga menziesii) | |||||
| (Int.) | −9.674 | 1.474 | −6.564 | <0.001 | |
| Distance | −0.007 | 0.001 | −6.108 | <0.001 | 265.845 |
| Aspect | 0.580 | 0.190 | 3.048 | 0.002 | 3.190 |
| Slope | 0.094 | 0.018 | 5.107 | <0.001 | 380.372 |
| TPI | −0.116 | 0.138 | −0.845 | 0.398 | 2.857 |
| Covariate | Std. Err. | t | p | Rel. Mag. | |
|---|---|---|---|---|---|
| Ponderosa pine (Pinus ponderosa); (marginal-R2 = 0.021) | |||||
| Intercept | −0.760 | 0.151 | −5.030 | <0.001 | |
| Distance | 0.001 | <0.001 | 3.110 | 0.002 | 1.595 |
| Aspect | −0.026 | 0.026 | −0.998 | 0.318 | 1.054 |
| Slope | −0.006 | 0.004 | −1.565 | 0.118 | 1.352 |
| TPI | 0.048 | 0.025 | 1.930 | 0.054 | 1.294 |
| Lodgepole pine (Pinus contorta); (marginal-R2 = 0.271) | |||||
| Intercept | −0.178 | 0.671 | −0.264 | 0.793 | |
| Distance | 0.003 | 0.001 | 1.995 | 0.054 | 3.652 |
| Aspect | −0.153 | 0.273 | −0.562 | 0.578 | 1.348 |
| Slope | −0.004 | 0.013 | −0.327 | 0.745 | 1.177 |
| TPI | −0.054 | 0.148 | −0.364 | 0.718 | 1.192 |
| Quaking aspen (Populus tremuloides); (marginal-R2 = 0.159) | |||||
| Intercept | −1.153 | 0.210 | −5.502 | <0.001 | |
| Distance | <0.001 | <0.001 | 2.333 | 0.020 | 1.445 |
| Aspect | −0.112 | 0.014 | −8.037 | <0.001 | 1.251 |
| Slope | 0.046 | 0.003 | 17.861 | <0.001 | 9.648 |
| TPI | −0.143 | 0.011 | −13.118 | <0.001 | 2.397 |
| Douglas-fir (Pseudotsuga menziesii); (marginal-R2 = 0.021) | |||||
| Intercept | −1.319 | 0.127 | −10.349 | <0.001 | |
| Distance | 0.001 | <0.001 | 1.656 | 0.099 | 1.421 |
| Aspect | 0.233 | 0.067 | 3.497 | 0.001 | 1.592 |
| Slope | −0.005 | 0.003 | −1.635 | 0.104 | 1.305 |
| TPI | −0.005 | 0.063 | −0.082 | 0.935 | 1.025 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ziegler, J.P.; Hoffman, C.M.; Fornwalt, P.J.; Sieg, C.H.; Battaglia, M.A.; Chambers, M.E.; Iniguez, J.M. Tree Regeneration Spatial Patterns in Ponderosa Pine Forests Following Stand-Replacing Fire: Influence of Topography and Neighbors. Forests 2017, 8, 391. https://doi.org/10.3390/f8100391
Ziegler JP, Hoffman CM, Fornwalt PJ, Sieg CH, Battaglia MA, Chambers ME, Iniguez JM. Tree Regeneration Spatial Patterns in Ponderosa Pine Forests Following Stand-Replacing Fire: Influence of Topography and Neighbors. Forests. 2017; 8(10):391. https://doi.org/10.3390/f8100391
Chicago/Turabian StyleZiegler, Justin P., Chad M. Hoffman, Paula J. Fornwalt, Carolyn H. Sieg, Mike A. Battaglia, Marin E. Chambers, and Jose M. Iniguez. 2017. "Tree Regeneration Spatial Patterns in Ponderosa Pine Forests Following Stand-Replacing Fire: Influence of Topography and Neighbors" Forests 8, no. 10: 391. https://doi.org/10.3390/f8100391
APA StyleZiegler, J. P., Hoffman, C. M., Fornwalt, P. J., Sieg, C. H., Battaglia, M. A., Chambers, M. E., & Iniguez, J. M. (2017). Tree Regeneration Spatial Patterns in Ponderosa Pine Forests Following Stand-Replacing Fire: Influence of Topography and Neighbors. Forests, 8(10), 391. https://doi.org/10.3390/f8100391