Abstract
Altitude significantly affects vegetation growth and distribution, including the developmental stages of a forest. We used shotgun Illumina sequencing to analyze microbial community composition and functional potential in melojo-oak (Quercus pyrenaica Willd.) rhizospheric soil for three different development stages along an altitudinal gradient: (a) a low altitude, non-optimal site for forest maintenance; (b) an intermediate altitude, optimal site for a forest; and (c) a high altitude, expansion site with isolated trees but without a real forest canopy. We observed that, at each altitude, the same microbial taxa appear both in the taxonomic analysis of the whole metagenome and in the functional analysis of the methane, sulfur and nitrogen metabolisms. Although there were no major differences at the functional level, there were significant differences in the abundance of each taxon at the phylogenetic level between the rhizospheres of the forest (low and intermediate altitudes) and the expansion site. Proteobacteria and Actinobacteria were the most differentially abundant phyla in forest soils compared to the expansion site rhizosphere. Moreover, Verrucomicrobia, Bacteroidetes and Nitrospirae phyla were more highly represented in the non-forest rhizosphere. Our study suggests that rhizospheric microbial communities of the same tree species may be affected by development stage and forest canopy cover via changes in soil pH and the C/N ratio.
1. Introduction
Forest soil is a highly complex ecosystem, due to both physical and chemical heterogeneity and strong nutritional influences both from falling leaves and root exudates [1,2]. The microbial communities of these soils have an essential role in nutrient cycles and ecosystem functionality [3,4] and are influenced by a large number of biotic and abiotic factors [5,6]. Habitat and soil type have been observed to have a stronger impact on microbial communities than the individual plant species [5], and soil pH, temperature, NH4+–N and vegetation diversity have been suggested to account for the majority (81.4%) of microbial community variations [6]. In Tibetan grasslands, evidence suggests that climate changes are likely to change soil microbial community functional structure [6].
Enhanced knowledge of biological altitudinal diversity patterns could facilitate a comprehensive understanding of the influence of climate change over ecosystems [7]. That is to say, the expansion pattern of plants along the altitudinal gradient and composition of microbial communities reflect the developmental stage and may mimic the biogeochemical dynamics of organic matter and ecosystem response to environmental changes [2]. In spite of the strong functional stability of soil microbial communities [8], they are able to respond more rapidly than plant communities to environmental changes [9]. Thus, it is of interest to integrate the knowledge of microbial community dynamics into the study of ecological succession. There are some studies that focus on trends of microbial richness or diversity along altitudinal gradients, which found that pH [10,11] and both pH and vegetation diversity [6,12] are the dominant factors influencing microbial community structure and composition. Moreover, tree composition has a great influence on microbial communities due to shifts in soil pH [13] produced by falling leaves and root exudates. But, the microbial functionality and phylogenetic relationships along the distinct development stages of a forest are poorly understood. In spite of the biases of the metagenomics approach by shotgun sequencing due to the presence of sequenced genomes in the databases only from cultivated bacteria [14], there is evidence for a positive link between soil microbial phylogenetic diversity and functional gene diversity in forest soils and also between the different microbial compositions in different forest types [15].
The Mediterranean region is a transition area between the arid climate of North Africa and the temperate and humid climate of Central Europe. In this ecotone, each small climate variation can lead to substantial changes, making the Mediterranean mountains a region potentially vulnerable to climate change [16]. The higher temperatures and reduced precipitation predicted for this region [17,18] will lead to significant changes in biogeographical vegetation patterns [19]. A protected area of Sierra Nevada, sited in the South-east of the Iberian Peninsula, is part of the UNESCO-MAB GLOCHAMOST project (Global and Climate Change in Mountain Sites). Mountainous regions harbor nearly 50 percent of the diversity hotspots of the planet [20]. These ecosystems present a high diversity of environmental conditions due to strong climatic gradients over a small spatial scale. Studies on the effect of climate change can be made via a strategy of space-for-time substitution as the climate variations with altitude are similar to latitudinal gradients [2,6] and can also reflect the development stages of the forest. The prediction of forest migration to higher altitudes forces us to have a better knowledge of ecosystem interactions and biochemical cycles of nutrients. The acquisition of this knowledge could be useful to design biological treatments to increase survival or improve development of trees at higher altitudes. It should be pointed out that there could be differences in microbial communities between optimal sites for forest development and non-optimal sites.
Notably, the melojo oak (Quercus pyrenaica Willd.) forest within the Sierra Nevada National Park has developed symptoms of a progressive migration along the slope, which may be due to limitations of water resources and elevated temperatures, especially at the lower altitude of the forest [21]. These oak forests show an advanced state of degradation [22], which could even be part of a general well-documented oak decline worldwide, spreading through Europe, Asia, North and South America [23]. Some recent studies indicate that there will be a decrease in the distribution area and an increase in altitudinal expansion [24], due to a predicted average temperature increase of about 4.8 °C by the end of the XXI century [24]. The work of Leverkus and colleagues [25], also performed in Sierra Nevada, supports this hypothesis of altitudinal expansion, since they found a higher survival and better growth of Q. pyrenaica and Quercus ilex L. seedlings above the upper forest limit than at its current site. However, a similar analysis of the tree- or seedling-associated microbial communities has not been done, though effective management of plant-microbe interactions could be useful to improve the survival and growth of Q. pyrenaica trees in future afforestation work.
In the present study, we conduct an analysis of rhizosphere melojo-oak microbial community structure, composition and functionality at three different altitudes in the National and Natural Park of Sierra Nevada. The Melojo-oak trees sampled were in different stages of forest development corresponding to: (a) lower altitude, possibly with incipient stress problems; (b) currently optimal mid-level altitude; and (c) higher altitude, possibly the future optimal area of distribution, where currently there are only some isolated or small clusters of trees amongst padded brushwood (Genista versicolor Boiss.) vegetation. Our study is based on metagenomic data obtained by Illumina shotgun sequencing. We analyzed the microbial groups and the main metabolic processes that might be occurring in this ecosystem within the biogeochemical cycles of methane, sulfur and nitrogen metabolism.
2. Materials and Methods
2.1. Site Selection and Sample Collection
Soil samples were collected on the same sunny day in May 2009 along an altitudinal gradient in the distribution area of melojo oak (Quercus pyrenaica Willd.) in the Sierra Nevada Natural and National Park (South-east Spain). Three study sites were chosen with respect to the degree of forest development in the municipality of Cañar (Granada, Spain): the lowest altitude site of the forest (LAF), where there was detected water stress on the oak forest [21]; the highest altitudinal limit of the forest (HAF); and the expansion zone of forest (XZF), where there are isolated or small groups of melojo oak trees within a padded brushwood (Genista versicolor) vegetation (Figure S1). These correspond to altitudes of around 1482, 1823 and 1945 m above sea level for LAF, HAF and XZF, respectively. Three sampling plots were randomly chosen within each study site along transects of 1.0 km length. In each plot, three trees were sampled, each tree with a trunk diameter of at least 15 cm at breast height, and separated by at least 5 m from the others. The sampled trees were marked, and all locations were recorded with a Global Positioning System (Figure S1).
The rhizospheric samples were collected according to Cobo-Díaz et al. [26] by following the tree’s main roots until young, cork-free roots were found at a distance of less than 50 cm from the trunk, where we collected soil that was attached to the roots. These samples were used for DNA extraction. For physico-chemical analyses of soils, an extra 500 g of soil was obtained in the depth range from 5 to 25 cm close to the sampling point and under the influence of tree roots. In both cases, the soils were stored immediately at 4 °C. The elimination of the first 5 cm of the soil allowed us to discard minor roots from herbaceous plants and leaf litter, which ensured the influence of the tree root on the sample. The soil samples were sieved through a 2-mm mesh. About two kilograms of a homogenous mix of soils from each study site were processed for physico-chemical analyses with standardized procedures (Table S1) at the Laboratorio Agroalimentario from the regional government of Junta de Andalucía at Atarfe (Granada, Spain).
2.2. Sequencing of Environmental DNA
Extraction of environmental DNA from each rhizospheric sample (0.25 g of soil) was carried out by a direct method using the PowerSoilTM DNA Isolation Kit from MoBio Laboratories Inc. (Carlsbad, CA, USA), following the manufacturer’s recommendations, within 24 h after the sample collection. For shotgun sequencing, DNAs from all nine samples (3 plots per site and 3 trees per plot) of each study site were pooled together [8,27,28] in order to obtain a homogeneous representation of each site (circa 20 ng·μL−1 of DNA from each extraction, with a total of 100 μL), and the DNA was concentrated in a speed vacuum centrifuge (3000 rpm for 30 min) to a final volume of 10 μL. The quantity and quality of the DNA were evaluated by agarose gel electrophoresis and with a NanoDrop® ND-1000 spectrophotometer (Thermo Scientific, Wilmington, DE, USA).
A total of 5 μg of each pooled DNA, from 9 rhizospheres per site, was sequenced using an Illumina HiSeq2000 at the Beijing Genomics Institute (Tai Po, Hong Kong, China) on one lane of paired-end 100 bp reads in order to obtain around 40 Gbp of information from each sampled site with an insert size of 170 bp. Sequences obtained were trimmed for quality using the Import Illumina Data of the GALAXY/JGI web server with the following parameters: MinSeqSize = 50 bp, and cutoff-quality = Q30. Paired sequences filtered were merged using the FLASH v1.0.3 web-tool from the GALAXY/JGI web server (https://galaxy.jgi-psf.org/) using default parameters. The obtained sequences were assembled using the SOAPdenovo program [29] using the k-mer sizes of 71, 75, 79, 83, 87 and 91, and performing a merger of the six assemblies. Non-redundant contigs obtained were uploaded to IMG/JGI [30] and MG-RAST [31] servers and made publicly accessible on MG-RAST under the codes 4508410.3, 4508411.3, and 4508412.3 for soil samples HAF, LAF, and XZF, respectively. Those contigs with similarity to prokaryotes were extracted and further processed.
2.3. Phylogenetic Assignment and Functional Analysis of Metagenomic Sequences
Phylogenetic assignment of contigs obtained was performed using BLASTX against the IMG genome database of IMG/JGI web server, using a cut-off e-value of 1e−10. The functional analysis was performed using BLASTX against the KEGG database of MG-RAST web server using a cut-off e-value of 1e−10. Annotation data files were processed with homemade scripts on Ruby with the following parameters: alignment length > 15 amino acids (aa), percentage of identity > 60, e-value < 10−5.
Statistical analysis of metagenomic profiles (STAMP) bioinformatics software v 1.0 [32] was used to compare community composition and functional gene abundance between the rhizospheric soil samples. This analysis was performed using pair to pair metagenomic samples (comparing one site to another individually), and statistical significance of the differences between samples was calculated using two-sided Fisher’s exact test, Newcombe-Wilson confidence interval method with 95% confidence interval, and Storey’s FDR method was used for multiple test correction. Biologically significant taxa were selected by filtering based on a q-value (0.05) and using only those categories that had at least a value of 1 for the difference between proportions or more than 2-fold ratio between proportions.
To compare the libraries, the ‘Vegan’ package of R (R Development Core Team 2016, http://www.R-project.org/) was used for a UPGMA clustering with the Euclidean distance matrix of the relative abundance of each taxonomic or functional group for each sample.
2.4. Taxonomic Classification of 16S rRNA Metagenomic Sequences
Perl homemade scripts were used for the taxonomic annotation of curated sequences obtained before the assembly. The rdp_classifier v2.3 (http://rdp.cme.msu.edu/classifier/classifier.jsp) with a pre-computed SILVA phylogenetic tree was used for the taxonomic annotation of the 16S rRNA sequences [33].
3. Results
3.1. Physico-Chemical Properties of the Soils
The soils from the study sites were sandy-loam (LAF and XZF) or loam (HAF), classified as haplic phaeozems of siliceous origin (Table S1). The pH was slightly acid in the expansion zone soil (XZF), while it was around neutral in forest soils. The HAF sample had the highest available water, and although the three samples were poor with respect to the organic matter content, HAF had a value twofold higher (Table S1). There were no significant differences in total nitrogen (N) between the sampled soils, but the carbon to nitrogen (C/N) ratio was significantly lower in the expansion zone soil (XZF) than in forest soils. HAF and XZF samples showed higher values of assimilable phosphorus compared with LAF (Table S1).
3.2. Taxonomic and Phylogenetic Analyses of the Metagenomes
Around 173, 171 and 192 million read pairs with an average length of 170 bp after merging were obtained for LAF, HAF and XZF samples, respectively. After a search for the SSU rRNA gene, 71,178 sequences were obtained from LAF reads, 65,286 from HAF and 135,237 from XZF, of which 59.6% to 65.7% were classified at family level at the RDP web-server. A total of 31 phyla was detected with the SSU rRNA sequences, although only 12 phyla had a relative abundance higher than 1% in one of the data sets (Figure 1a). Proteobacteria and Actinobacteria, the most abundant phyla, had significantly greater representation in forest soil samples (LAF and HAF) than in the soil sample from the expansion zone, while there were 4 phyla (Bacteroidetes/Chlorobi, Nitrospirae, Fusobacteria and Euryarchaeota) over-represented in the latter soil sample. Acidobacteria were more abundant in LAF than in XZF, and Planctomycetes in HAF than in XZF.
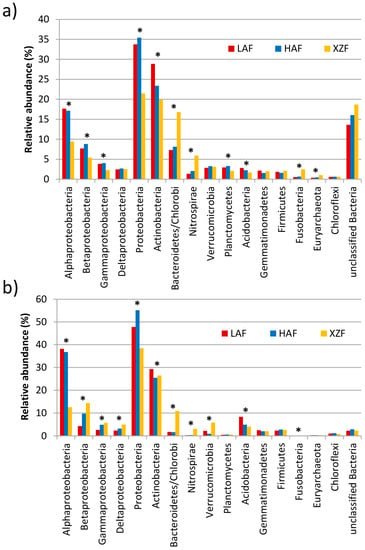
Figure 1.
Taxonomic and phylogenetic assignment of reads at the phylum and class levels. (a) Taxonomic classification of SSU rRNA sequences by BLASTN against the SILVA database using Classifier v2.3 software; (b) Phylogenetic classification of the total contigs datasets annotated on the KO database, performed by BLASTX analysis against the Greengenes genome database using the IMG/JGI web server. The Y axis represents the relative abundance of each taxon expressed as a percentage. Phyla and Proteobacteria classes with more than 1% relative abundance on at least one of the datasets are plotted. Taxonomical groups with statistically significant differences between samples are marked with an asterisk.
Assembly of the reads generated 205,779, 205,095 and 246,663 contigs for LAF, HAF and XZF, respectively (Table 1). That is to say, there were 20% more contigs produced from the XZF sample than from the forest samples, possibly due to the larger number of reads obtained for XZF. The contig mean lengths were 320, 351 and 343 bp for LAF, HAF and XZF, respectively, while the longest contig of each sample was 6579, 18,521 and 91,563 bp, respectively. The annotated contigs against KEGG database were 184,352, 184,881 and 193,696 for HAF, XZF and LAF samples, respectively. From the sequences annotated, 70,027, 64,778 and 66,320 contigs for LAF, HAF and XZF, respectively, passed the quality filter of the annotation (KEGG Orthology annotation with e-value < 10−5, % identity > 60 and alignment length > 15 bp). Similar patterns were observed for the phylogenetic annotation of the total contigs with consistent homology to at least one gene of the KEGG Orthology (KO) database (data not shown). All the phyla, except Verrucomicrobia, Planctomycetes and Euryarchaeota, showed the same differences (Figure 1b) between samples for SSU rRNA data and for total KO data obtained, although some differences for abundances within a phylum were observed depending on the data set analyzed (Figure 1). Within the phylum Proteobacteria, class Alphaproteobacteria had greater abundance in forest soil (LAF and HAF) than in the expansion zone sample (XZF), both for SSU rRNA and total KO datasets. The same occurred with Betaproteobacteria and Gammaproteobacteria, but only for total SSU rRNA datasets, since Betaproteobacteria had higher abundance in HAF and XZF than in LAF for KO datasets (Figure 1a). Thus, the taxonomic (SSU rRNA gene) and phylogenetic analyses of the sequences and contigs showed a similar distribution pattern of phyla.

Table 1.
Main values of the assembly process with the number of reads, contigs and annotated genes. LAF, lowest altitudinal forest; HAF, high altitudinal forest; and XZF, expansion zone forest.
3.3. Functional Analysis of the Metagenomes
Metabolism was the most abundant functional group, at level 1 of hierarchical classification using the KEGG Orthology database (Data not shown), with similar values for the 3 samples: 37.83%, 37.63% and 37.93% of the total contigs annotated for LAF, HAF and XZF, respectively. Within level 1 of the Metabolism functional group, the level 2 most abundant groups were Amino acid metabolism, Carbohydrate metabolism and Energy metabolism (Figure 2a). In addition, within this latter group, there were significant differences for relative abundances of contigs for Nitrogen metabolism (p < 0.01) and Sulfur metabolism (p < 0.001) between HAF and XZF, and for Methane metabolism (p < 0.001) between the expansion zone soil (XZF) sample and forest rhizosphere soil samples (Figure 2b). Thus, these metabolic pathways with statistically significant differences were further analyzed.
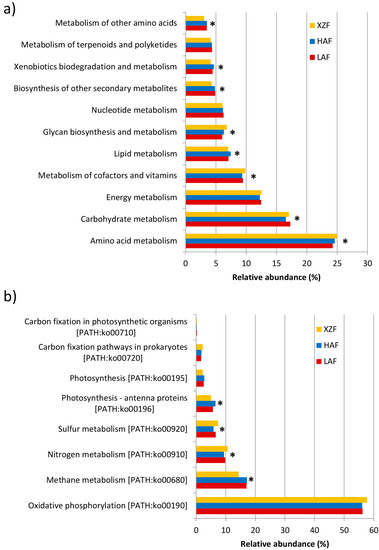
Figure 2.
Percentage of contigs for functional groups within level 1—Metabolism (a) and Level 2—Energy metabolism (b), according to the KEGG Orthology classification of the MG-RAST web-server. Functional groups with statistically significant differences between samples are marked with asterisks.
3.4. Functional Analysis of Methane, Sulfur and Nitrogen Metabolism Genes
A total of 64 genes related to methane metabolism was detected (Figure S2), with formate dehydrogenase major subunit as the most abundant and with significant differences between HAF and XZF (Figure 3); and acetyl-CoA synthetase the second most abundant (10.33–10.58%) but without significant differences among samples. Significant differences between samples were observed for 23 genes (Figure 3). The four subunits of both formate dehydrogenase and carbon-monoxide dehydrogenase had greater representation in rhizospheric forest soil samples (LAF and HAF) than in the rhizospheric samples of the expansion area, while Pyruvate ferredoxin oxidoreductase, also formed by four subunits, and methane/ammonia monooxygenase showed the opposite pattern, with more abundance in the expansion area sample than in rhizospheric forest soil samples (Figure 3). An overall view of methane metabolism showed that formate oxidation (formate → carbon monoxide) accounted for the majority of sequences related to methane metabolism, and this metabolic pathway was over-represented in forest soil samples (Figure 3 and Figure 4a).
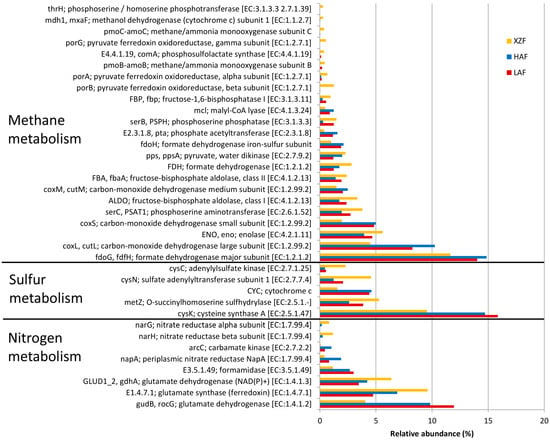
Figure 3.
Percentage of genes involved in methane, sulfur and nitrogen metabolism with statistically significant differences (p < 0.05) between sampled sites.
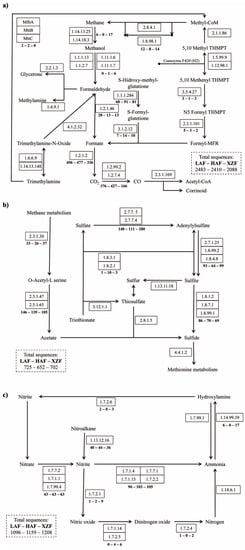
Figure 4.
Partial pathways of methane metabolism (a), sulfur metabolism (b) and nitrogen metabolism (c). Total sequences per sample are indicated by discontinuous line boxes. Sequences obtained for each protein or protein group are indicated under the continuous line boxes, separated between dashes for each sample (always in the order LAF–HAF–XZF).
A total of 26 genes related to sulfur metabolism were detected, with major representation of those related to assimilatory sulfate reduction, although the most abundant gene for all the samples was cysteine synthase A (9.54–15.86%), related to cysteine and methionine metabolism (Figure S3). There were 5 sulfur metabolism genes with significant differences between samples, always between XZF and forest soil samples. Cytochrome c and cysteine synthase A had greater representation in both forest soil samples while sulfate adenylyltransferase subunit 1 and o-succinylhomoserine sulfhydrylase were more abundant in XZF than HAF and adenylylsulfate kinase in non-forest soil sample than in forest soil samples (Figure 3). Sulfate assimilation (sulfate → adenylylsulfate → sulfite → sulfide) accounted for the most sequences within sulfur metabolism, followed by genes for the production of acetate from methane (Figure 4b). However, because no sequences were obtained for the 2.8.1.5 (thiosulfate-dithiol sulfurtransferase) enzyme mediating the conversion of thiosulfate to sulfide, the relationship between the two pathways in these communities is unclear.
A total of 32 genes related to nitrogen metabolism was detected (Figure S4) with an abundance of genes from the ammonification and ammonium assimilation, related to glutamate metabolism, such as glutamine synthetase (29.47%–30.13%), glutamate synthase (NADPH/NADH) large chain (11.76%–15.51%), glutamate dehydrogenase (4.06%–11.95%), glutamate synthase (ferredoxin) (4.74%–9.60%) and glutamate synthase (NADPH/NADH) small chain (6.39%–7.70%). Significant differences between samples were detected for 8 nitrogen metabolism genes, always between the rhizosphere of forest soil sample (LAF or HAF) and the expansion zone sample (XZF, Figure 3). These findings indicate that there were differences in ammonia assimilation, ammonification and nitrification genes between forest soil samples and the expansion area sample, while there were no significant differences between the two forest soils. With respect to the inorganic part of nitrogen metabolism, the nitrate assimilation (NO3− → NH4+) pathway had the highest number of sequences, followed by the nitrite obtained from nitroalkane. These pathways of N assimilation were significantly increased in the XZF sample. However, there were few sequences for other metabolic functions such as denitrification and nitrification, and nitrogen fixation was not detected (Figure 4c). Also, it should be noted that while nitrate assimilation had around 10% of the sequences related to nitrogen metabolism, the ammonia assimilation and ammonification together accounted for around 70% of sequences, being the most represented function for the nitrogen cycle (Figure S4).
3.5. Phylogenetic Analysis of Methane, Sulfur and Nitrogen Metabolism Sequences
The phylogenetic affiliation of the contigs related to methane, sulfur and nitrogen metabolism showed similar patterns of abundances within each metabolic pathway analyzed at the phylum level (Figure 5). This analysis showed that Proteobacteria and Actinobacteria were the most represented phyla for the three metagenomes and for the three metabolic cycles. Proteobacteria had significantly higher abundance in forest soil samples than in the expansion area soil for all three metabolisms, while Actinobacteria had greater representation in LAF and XZF than in HAF, but it was only significant for methane metabolism (Figure 5a). There were significant differences between samples in the other 8 phyla for methane metabolism, 7 for sulfur metabolism and 6 for nitrogen metabolism, considering only those phyla with abundances higher than 1%. Acidobacteria had higher abundance in LAF than in the other samples for the three metabolic cycles, while Chloroflexi were greater in HAF than in XZF. Some phyla showed higher representation in the oak rhizosphere of the expansion area, including Verrucomicrobia, Bacteroidetes and Nitrospirae, for methane, sulfur and nitrogen metabolism. Finally, Gemmatimonadetes were more abundant in forest soil samples for methane metabolism sequences, while Firmicutes had higher representation in XZF than LAF for sulfur metabolism (Figure 5b). Within the phylum Proteobacteria, there were some significant differences at the class level between the expansion area and the two (LAF and HAF) forest soil samples (Figure 5), and for all three metabolic cycles studied. Alphaproteobacteria were more abundant in the oak rhizosphere forest soil than in the expansion area sample, while the opposite occurs with Betaproteobacteria, Gammaproteobacteria and Deltaproteobacteria, which had more representation in the XZF rhizospheric soil sample than in forest soil samples (Figure 5).
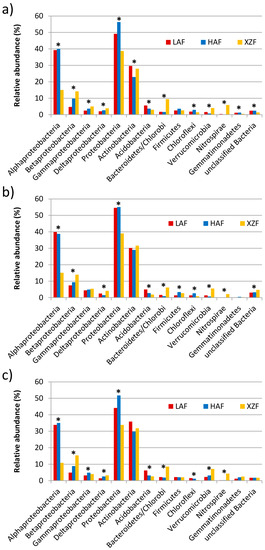
Figure 5.
Phylogenetic classification and relative abundance at the phyla level for methane metabolism (a), sulfur metabolism (b) and nitrogen metabolism (c) sequences, performed by BLASTX analysis against the Greengenes genome database using the IMG/JGI web server. The Y axis represents the relative abundance of each taxon expressed as a percentage. Phylogenetic classification of the Proteobacteria is also shown at the class level. Functional groups with statistically significant differences between samples are marked with asterisks.
The assembled sequences annotated on KEGG Orthology database for methane, sulfur and nitrogen metabolism were phylogenetically assigned to the level of family. Bradyrhizobiaceae (Alphaproteobacteria) and Mycobacteriaceae (Actinobacteria) were the main detected families among the sequences, with significant differences between forest soil samples and the expansion site (Figure 6). For the three metabolic cycles, Bradyrhizobiaceae was relatively more abundant in LAF and HAF samples compared to Mycobacteriaceae, which was more abundant in the XZF soil sample. Moreover, another 36, 22 and 19 families had significant differences for methane metabolism, sulfur metabolism and nitrogen metabolism, respectively, among the three sampled sites. There were families with higher representation in the LAF sample than in the other samples, such as Pseudonocardiaceae, Acidobacteriaceae, Micromonosporaceae and Caulobacteraceae for the three metabolic cycles, Xanthobacteraceae for methane metabolism only, and Sphingomonadaceae for nitrogen metabolism. Nocardioidaceae were also over-represented in HAF for all the metabolic cycles presented in this study, while Methylobacteriaceae was for sulfur metabolism. Others families had higher relative abundance in HAF and XZF than in LAF, as Burkholderiaceae for methane metabolism sequences. Conversely, Burkholderiaceae had greater representation in the non-forest soil sample for nitrogen metabolism but no significant difference in sulfur metabolism. Another family over-represented in the non-forest soil sample XZF was Oxalobacteraceae for methane metabolism. Conversely, the family Conexibacteraceae was more abundant in forest soil samples for methane and sulfur metabolism (Figure 6).
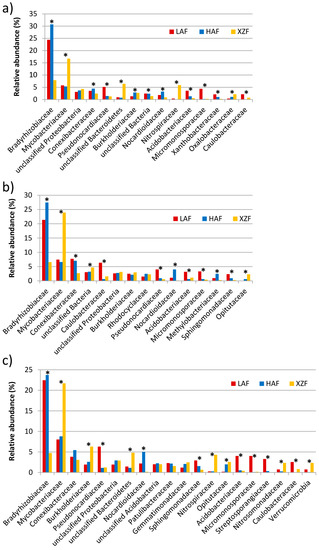
Figure 6.
Phylogenetic classification at the family level for methane metabolism (a), sulfur metabolism (b) and nitrogen metabolism (c) sequences, performed by BLASTX analysis against the Greengenes genome database using the IMG/JGI web server. The Y axis represents the relative abundance of each taxon expressed as a percentage. Those families with more than 2% abundance in at least one of the datasets are shown. Functional groups with statistical significant differences between samples are marked with asterisk.
The analysis by agglomerative hierarchical clustering with Euclidean distance of these sequences at the family level showed remarkable differences depending on the use of a presence/absence (P/A) data matrix versus a relative abundance data matrix. The use of a P/A matrix resulted in clustering primarily by the type of metabolism rather than by sampling site (Figure 7a). However, the use of a relative abundance matrix resulted in clustering by the sampling site (Figure 7b), showing that the forest soil samples were closer to each other than to the XZF soil sample. The same results were obtained with Bray-Curtis dissimilarity (data not shown). This means that the same microbial families are involved in each metabolic cycle across all sites, but the environment of the sampling site determines the relative abundance of each taxa, which could reflect the relative importance of each metabolic cycle at each specific sampling site.
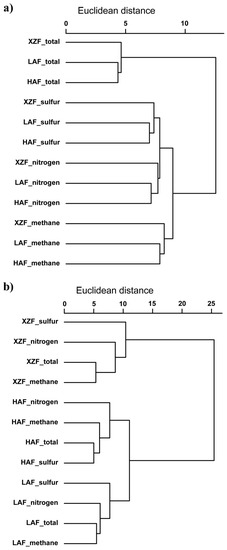
Figure 7.
Agglomerative hierarchical clustering for the presence/absence (a) and relative abundance (b) of sequences grouped by phylogenetic families for both the total and the metabolic cycles analyzed of each sampled site. Euclidean distance matrix and UPGMA algorithm were used for this analysis.
4. Discussion
Major differences in biological variables were found between the expansion area (XZF) and the other (LAF and HAF) soil samples, which suggests that the altitudinal gradient is not the main cause of these differences, as opposed to the developmental status of melojo-oak forest, the tree canopy coverage, climate conditions and/or some other biotic or abiotic parameters included or not included in the sampling procedure. The physico-chemical analyses of soils showed lower values of pH and the C/N ratio at the highest sampled site XZF, where there are only some isolated or small clusters of trees within padded brushwood vegetation but without a real forest canopy. In mountains ecosystems, the highest areas have less developed soils with shallower horizons, which together with the absence of tree canopy, could affect the nutrients available in the rhizosphere. An increase in the C/N ratio has been previously correlated with a decrease in nitrogen decomposition and mineralization [34], meaning that forest soil works better as a carbon and nitrogen reservoir than non-forest soil, despite being under the influence of the same tree species, as in the case of XZF. However, microbial community structure and composition have also been correlated with physico-chemical parameters, such as pH [6,10,11,35,36] and the C/N ratio [36,37], rather than the altitudinal gradient per se. Therefore, the differences in microbial community composition and functionality between forest soils and XZF (non-forest) may be caused by the distinct pH and C/N values existing for these soils.
Differences due to methodology, such as the database used [38], must be taken into account when comparing taxonomical data between different studies, or even between 16S rRNA gene sequences and all the annotated sequences [38,39] for the same dataset. It is also important to note that taxonomic assignment based solely on protein encoding genes currently generates an artificial bias against groups with few sequenced genomes, due to the uneven taxonomic composition of protein and nucleotide databases [14], as can be observed in the results of Figure 1. This likely explains the differences in abundance of unclassified sequences between the 16S rRNA set and the phylogenetic analysis of the annotated sequences set. Another kind of bias could be due to the sequencing of single samples. The deep-sequencing of 16S rRNA amplicons from oak rhizosphere in another study showed that 1 out of 3 samples could offer a dissimilar result [40], but the conclusions obtained with the full set of sequences were similar to the results of the individual analysis of the majority of the samples. Thus, the authors suggested that a single pyrosequencing run would be needed for a comprehensive description of the bacterial communities [40]. The sequencing of 9 tree samples per site, with 20 million reads per rhizosphere, would have allowed us intra-site comparisons but with a very low coverage and scarce representation of the metabolic pathways. Therefore, one single run of 180 million reads with DNA obtained by the pooling of 9 samples from 3 plots per site was the best option in order to obtain a homogeneous representation of the oak rhizosphere in the context of the altitude, diversity of soils, geographic distribution and minimizing the effect of spot perturbations, and in agreement with current methodology [8,27,28].
The high abundance of sequences associated with amino acid, carbohydrate and energy metabolism, and also metabolism of cofactors and vitamins, indicates the presence of essential features for competitive microbial life within the forest soil [41,42]. It has been suggested that the terrestrial CH4 flux could be controlled by microorganisms and that methanotroph microorganisms are particularly active in temperate forest soils [43]. Formate degradation to carbon monoxide makes up the highest percentage of sequences for methane metabolism, and its increase in forest soils could reflect a trend of higher methane oxidation in this kind of soil as compared to the XZF soils. For example, the carbon-monoxide dehydrogenase is over 15% of total methane sequences in forest soils, while in XZF soils, it is below 8% (Figure 3). Previous studies showed that afforestation or reforestation of aerobic soils can enhance soil methane oxidation by influencing the composition and activity of soil methanotrophs [44]. Thus, this result suggests that the forest soil may be a better methane sink than the shrubland (XZF) soil with isolated trees, in agreement with previous results [44]. On the other hand, the major abundance of sequences for pyruvate synthesis and minor for formate degradation to carbon monoxide in the XZF soil sample could be an indication of an alternative pathway for carbon fixation, which supplements the scant carbon supply by litter decomposition and from methane.
Sulfate assimilation (from sulfate till sulfide) is the most prominent pathway for sulfur metabolism in all three soils. The difference in the number of sequences of the sulfate adenylyltransferase could reflect a real incorporation of sulfur from the environment in the XZF rhizospheric soil. Ammonium assimilation and nitrate ammonification are the main functional groups for nitrogen metabolism, meaning that it is a developed ecosystem for the nitrogen cycle [45], because the ammonia generated by degradation of organic compounds is probably taken up, preventing its loss. That is to say, there is a strong use of organic sources of nitrogen at the three sampling sites mainly by the EC: 1.4.1.2 (glutamate dehydrogenase) enzyme in forest soils, and by enzymes EC: 1.4.1.3 [glutamate dehydrogenase (NAD(P)+)] and EC: 1.4.7.1 (glutamate synthase) in the XZF soil. Moreover, in XZF soil, the higher amount of sequences for nitrate reductase (narG and narH genes) could reflect an extra incorporation of N from inorganic sources. Taken together, and in spite of the similar metabolic pathways, it seems that in the XZF soil, there is a higher use of inorganic N and S sources.
Proteobacteria and Actinobacteria were the dominant phyla in the SSU sequence analyses, as is usually the case for soil microbial communities [14,46,47,48]. The same result was obtained for total protein-coding sequences within the three metabolic pathways analyzed. Proteobacteria play a key role in C, N and S cycling [49], and their dominance in rhizospheric forest samples shows the copiotrophic conditions therein. The high presence of Actinobacteria in all the sampled sites may be an indicator of the presence of aromatic compounds in oak leaves [50] or of low carbon availability, as Actinobacteria are described to be adapted to these environmental features [51]. Verrucomicrobia, Bacteroidetes and Nitrospirae were more abundant in the XZF sample than in forest soils for both SSU rRNA sequences and for methane, sulfur and nitrogen metabolism protein-coding sequences. Verrucomicrobia and Bacteroidetes have been described as oligotrophic phyla [52], and a decline of Verrucomicrobia sequences due to nitrogen fertilization has been observed elsewhere [53], thereby highlighting the oligotrophic features of this phylum. Therefore, the over-representation of these phyla in XZF soil could be an indicator of oligotrophic conditions, due in part to a lower intake of nutrients through leaf fall. The higher relative abundance of Bacteroidetes, Betaproteobacteria and Gammaproteobacteria in the XZF sample is similar to that observed in another study on Retama sphaerocarpa shrubs [54], suggesting the influence of the surrounding padded brushwood vegetation on the rhizospheric microbial communities of the isolated trees. The greater abundance of Acidobacteria at the low elevation forest sample agrees with some studies of altitudinal gradient [10,35], although the opposite has been seen in other studies [48,55]. In some cases, the relative abundance of Acidobacteria is negatively correlated with pH [10] or positively with carbon inputs [55]. Fierer et al. [56] observed that the differences along an altitudinal gradient are due more to changes in pH than the altitudinal gradient itself. It appears that, as was observed by Defossez et al. [57], abiotic factors have a strong influence on microbial communities, which at the same time have an influence on and are influenced by the plant community. In the XZF area, the edge effect of a forest ecosystem is strong since it receives more light and wind, producing drier conditions and more extreme temperatures than found in forest interiors [58]. Therefore, changes in the soil microbial community structure and activity can be expected, and these changes may influence litter decomposition rates and nutrient cycling in the ecosystem [59].
At the class level, the relative increases in Beta- and Gammaproteobacteria, and decreases in Alphaproteobacteria, along the altitudinal gradient for SSU sequences correlates with another study that demonstrated the influence of pH and vegetation type on bacterial community structure and composition along an altitudinal gradient on Changbai Mountain [35], as pH and the C/N ratio were also lower in the padded brushwood vegetation without a forest canopy. The more represented families for protein-coding sequences were Bradyrhizobiaceae (phyla Proteobacteria) and Mycobacteriaceae (phyla Actinobacteria), which have also been two of the most represented families in other forest soils [46,47]. But in our case, there was a higher representation of Bradyrhizobiaceae in forest soils, while Mycobacteriaceae were more abundant in the expansion site. Among the Bradyrhizobiaceae is the genus Bradyrhizobium, a well-known Nitrogen-fixing bacteria within root nodules of leguminous plants, including trees [60] and Tardiphaga, which has also been isolated from nodules of tree roots [61]. Thus, different genera of this family may interact with tree roots and may be involved in the N cycle, both in plant symbiosis and in association to the roots. Other genera in the Bradyrhizobiaceae family, such as Bosea (involved in the sulfur cycle), Nitrobacter (oxidation of nitrite) or Oligotropha (fixation of carbon monoxide), could be important in the described metabolic pathways. However, the deep sequencing approach used in this study, with short reads, does not allow us to know the exact genera that are involved in these processes. The Mycobacteriaceae family is composed of genera Mycobacterium and Hoyosella. Mycobacteria are generally free-living saprophytes but are very well known as agents of human diseases [62]. However, there is accumulating evidence to suggest that the genus Mycobacterium can be associated with forest soils [46,62,63]. Unfortunately, information about its functional role in these ecosystems is minimal since these studies were focused on natural reservoirs of human pathogens. Based on recent evidence, the role of Mycobacteria in wild soils and as plant-associated bacteria should be investigated.
5. Conclusions
This study has shown that there are phylogenetic differences in the rhizospheric microbial communities of the same tree species, which may depend on the existence of forest canopy as well as the soil pH and C/N ratio. Furthermore, despite the existence of some taxonomic specificity for methane, sulfur and nitrogen metabolism, the same taxa were represented in each of these metabolic pathways. In addition, the major difference was due to the relative abundance of each taxon at the different altitudinal points without a dramatic change in the nature of the microbial community. Future research with more extensive sampling and replication, greater depth of sequencing and direct assessment of metabolic rates is necessary to have a better knowledge of the functioning of these ecosystems.
Supplementary Materials
The following are available online at www.mdpi.com/1999-4907/8/10/390/s1, Figure S1: Description of the sampled areas, Figure S2: Relative abundance of the sequences obtained for methane metabolism, Figure S3: Relative abundance of the sequences obtained for sulfur metabolism, Figure S4, Relative abundance of the sequences obtained for nitrogen metabolism, Table S1: Chemical and physical properties of soils.
Acknowledgments
This work was supported by research grants including ERDF (European Regional Development Funds): P08-CVI-03549 from Consejería de Innovación, Ciencia y Empresa from Junta de Andalucía, OAPN 021/2007 from Organismo Autónomo Parques Nacionales (Spanish Ministry of Environment) and 20134R069-RECUPERA 2020 from the Spanish Ministerio de Economía y Competitividad and CSIC. We would like to thank the authorities of the Sierra Nevada National Park for the access, facilities and soil sampling, and also Edward Kirton (JGI, California) for technical assistance with bioinformatics. JFCD was awarded a postdoctoral contract from Junta de Andalucía, and AJFG was awarded a postdoctoral contract from RECUPERA 2020. The work of the U.S. Department of Energy Joint Genome Institute, a DOE Office of Science User Facility, is supported by the Office of Science of the U.S. Department of Energy under Contract No. DE-AC02-05CH11231.
Author Contributions
F.L., S.T. and N.T. conceived and designed the experiments; C.D., F.G. and P.V. performed the experiments: C.D. and F.G. analyzed the data; F.L. and C.D. wrote the original draft; F.L., N.T. and S.T. wrote, reviewed and edited the paper.
Conflicts of Interest
The authors declare no conflict of interest. The founding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
References
- Bardgett, R.D.; Mawdsley, J.L.; Edwards, S.; Hobbs, P.J.; Rodwell, J.S.; Davies, W.J. Plant species and nitrogen effects on soil biological properties of temperate upland grasslands. Funct. Ecol. 1999, 13, 650–660. [Google Scholar]
- Bragazza, L.; Bardgett, R.D.; Mitchell, E.A.D.; Buttler, A. Linking soil microbial communities to vascular plant abundance along a climate gradient. New Phytol. 2015, 205, 1175–1182. [Google Scholar] [CrossRef] [PubMed]
- Van der Heijden, M.G.A.; Bardgett, R.D.; van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [PubMed]
- Strickland, M.S.; Lauber, C.; Fierer, N.; Bradford, M.A. Testing the functional significance of microbial community composition. Ecology 2009, 90, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.K.; Nazaries, L.; Munro, S.; Anderson, I.; Campbell, C.D. Use of multiplex terminal restriction fragment length polymorphism for rapid and simultaneous analysis of different components of the soil microbial community. Appl. Environ. Microbiol. 2006, 72, 7278–7285. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Gao, Y.; Wang, S.; Xu, D.; Yu, H.; Wu, L.; Lin, Q.; Hu, Y.; Li, X.; He, Z.; et al. The microbial gene diversity along an elevation gradient of the Tibetan grassland. ISME J. 2014, 8, 430–440. [Google Scholar] [PubMed]
- Malhi, Y.; Silman, M.; Salinas, N.; Bush, M.; Meir, P.; Saatchi, S. Introduction, elevation gradients in the tropics, laboratories for ecosystem ecology and global change research. Glob. Chang. Biol. 2010, 16, 3171–3175. [Google Scholar] [CrossRef]
- Curiel-Yuste, J.; Fernandez-Gonzalez, A.J.; Fernandez-Lopez, M.; Ogaya, R.; Peñuelas, J.; Sardans, J.; Lloret, F. Strong functional stability of soil microbial communities under semiarid Mediterranean conditions is subjected to long-term shifts in baseline precipitation. Soil Biol. Biochem. 2014, 69, 223–233. [Google Scholar]
- Cong, J.; Yang, Y.; Liu, X.; Lu, H.; Zhou, J.; Li, D.; Yin, H.; Ding, J.; Zhang, Y. Analyses of soil microbial community compositions and functional genes reveal potential consequences of natural forest succession. Sci. Rep. 2015, 5, 10007. [Google Scholar] [PubMed]
- Bryant, J.A.; Lamanna, C.; Morlon, H.; Kerkhoff, A.J.; Enquist, B.J.; Green, J.L. Microbes on mountainsides: Contrasting elevational patterns of bacterial and plant diversity. Proc. Nat. Acad. Sci. USA 2008, 105, 11505–11511. [Google Scholar] [PubMed]
- Singh, D.; Shi, L.; Adams, J.M. Bacterial diversity in the mountains of South-West China: Climate dominates over soil parameters. J. Microbiol. 2013, 51, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Lee-Cruz, L.; Kim, W.S.; Kerfahi, D.; Chun, J.H.; Adams, J.M. Strong elevational trends in soil bacterial community composition on Mt. Halla, South Korea. Soil Biol. Biochem. 2014, 68, 140–149. [Google Scholar] [CrossRef]
- Landesman, W.J.; Nelson, D.M.; Fitzpatrick, M.C. Soil properties and tree species drive β-diversity of soil bacterial communities. Soil Biol. Biochem. 2014, 76, 201–209. [Google Scholar] [CrossRef]
- Urich, T.; Lanzén, A.; Qi, J.; Huson, D.H.; Schleper, C.; Schuster, S.C. Simultaneous assessment of soil microbial community structure and function through analysis of the meta-transcriptome. PLoS ONE 2008, 3, e2527. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cong, J.; Lu, H.; Yang, C.; Yang, Y.; Zhou, J.; Li, D. An integrated study to analyze soil microbial community structure and metabolic potential in two forest types. PLoS ONE 2014, 9, e93773. [Google Scholar] [CrossRef] [PubMed]
- Lionello, P.; Malanotte-Rizzoli, P.; Boscolo, R.; Alpert, P.; Artale, V.; Li, L.; Luterbacher, J.; May, W.; Trigo, R.; Tsimplis, M.; et al. The Mediterranean climate: An overview of the main characteristics and issues. In Mediterranean Climate Variability; Lionello, P., Malanotte-Rizzoli, P., Boscolo, R., Eds.; Elsevier: Amsterdam, The Netherlands, 2006; pp. 1–26. [Google Scholar]
- Giorgi, F.; Lionello, P. Climate change projections for the Mediterranean region. Glob. Planet. Chang. 2008, 63, 90–104. [Google Scholar] [CrossRef]
- Felicísimo, A.M.; Muñoz, J.; Villalba, C.J.; Mateo, R.G. Impactos, vulnerabilidad y adaptación al cambio climático de la biodiversidad española. In Flora y Vegetación; PNACC; Ministerio de Medio Ambiente y Medio Rural y Marino: Madrid, Spain, 2011; Volume 1. [Google Scholar]
- Ruiz-Labourdette, D.; Schmitz, M.F.; Pineda, F.D. Changes in tree species composition in Mediterranean mountains under climate change: Indicators for conservation planning. Ecol. Indic. 2013, 24, 310–323. [Google Scholar] [CrossRef]
- Kohler, T.; Werhli, A.; Jurek, M. (Eds.) Mountains and climate change: A global concern. In Sustainable Mountain Development Series; Centre for Development and Environment (CDE), Swiss Agency for Development and Cooperation (SDC) and Geographica Bernensia: Berna, Switzerland, 2014. [Google Scholar]
- Leal-García, J. Análisis del Crecimiento Vegetativo e Inversión Reproductiva de Quercus pyrenaica Willd. en Tres Localidades de Sierra Nevada. Master’s Thesis, University of de Granada, Granada, Spain, 2013. [Google Scholar]
- Camacho-Olmedo, M.T.; García-Martínez, P.; Jiménez-Olivencia, Y.; Menor-Toribio, J.; Paniza-Cabrera, A. Dinámica evolutiva del paisaje vegetal de la Alta Alpujarra granadina en la segunda mitad del siglo XX. Cuadernos Geográficos de la Universidad de Granada 2002, 32, 25–42. [Google Scholar]
- Ragazzi, A.; Moricca, S.; Dellavalle, I.; Turco, E. Italian expansion of oak decline. In Decline of Oak Species in Italy; Problems and Perspectives; Ragazzi, A., Dellavalle, I., Eds.; Universita di Firenze, Concilio Nazionale di Ricerca, Accademia Italiana di Scienze Forestali: Firenze, Italy, 2000; pp. 39–75. [Google Scholar]
- Benito, B.; Lorite, J.; Peñas, J. Simulating potential effects of climatic warming on altitudinal patterns of key species in Mediterranean-alpine ecosystems. Clim. Chang. 2011, 108, 471–483. [Google Scholar] [CrossRef]
- Leverkus, A.B.; Castro, J.; Delgado-Capel, M.J.; Molinas-González, C.; Pulgar, M.; Marañón-Jiménez, S.; Delgado-Huertas, A.; Querejeta, J.I. Restoring for the present or restoring for the future: Enhanced performance of two sympatric oaks (Quercus ilex and Quercus pyrenaica) above the current forest limit. Restor. Ecol. 2015, 23, 936–946. [Google Scholar] [CrossRef]
- Cobo-Díaz, J.F.; Fernández-González, A.J.; Villadas, P.J.; Robles, A.B.; Toro, N.; Fernández-López, M. Metagenomic assessment of the potential microbial nitrogen pathways in the rhizosphere of a Mediterranean forest after a wildfire. Microb. Ecol. 2015, 69, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Sheik, C.S.; Beasley, W.H.; Elshahed, M.S.; Zhou, X.; Luo, Y.; Krumholz, L.R. Effect of warming and drought on grassland microbial communities. ISME J. 2011, 5, 1692–1700. [Google Scholar] [CrossRef] [PubMed]
- Yergeau, E.; Bokhorst, S.; Kang, S.; Zhou, J.; Greer, C.W.; Aerts, R.; Kowalchuk, G.A. Shifts in soil microorganisms in response to warming are consistent across a range of Antarctic environments. ISME J. 2012, 6, 692–702. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhu, H.; Ruan, J.; Qian, W.; Fang, X.; Shi, Z.; Li, Y.; Li, S.; Shan, G.; Kristiansen, K.; et al. De novo assembly of human genomes with massively parallel short read sequencing. Genome Res. 2010, 20, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Markowitz, V.M.; Chen, I.A.; Chu, K.; Szeto, E.; Palaniappan, K.; Pillay, M.; Ratner, A.; Huang, J.; Pagani, I.; Tringe, S.G.; et al. IMG/M 4 version of the integrated metagenome comparative analysis system. Nucl. Acids Res. 2014, 42, D568–D573. [Google Scholar] [CrossRef] [PubMed]
- Meyer, F.; Paarmann, D.; D’souza, M.; Olson, R.; Glass, E.M.; Kubal, M.; Paczian, T.; Rodriguez, A.; Stevens, R.; Wilke, A.; et al. The metagenomics RAST server—A public resource for the automatic phylogenetic and functional analysis of metagenomes. BMC Bioinf. 2008, 9, 386. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Beiko, R.G. Identifying biologically relevant differences between metagenomic communities. Bioinformatics 2010, 26, 715–721. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucl. Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Pérez, C.A.; Carmona, M.R.; Fariña, J.M.; Armesto, J.J. Selective logging of lowland evergreen rainforests in Chiloé Island, Chile: Effects of changing tree species composition on soil nitrogen transformations. For. Ecol. Manag. 2009, 258, 1660–1668. [Google Scholar] [CrossRef]
- Shen, C.; Xiong, J.; Zhang, H.; Feng, Y.; Lin, X.; Li, X.; Liang, W.; Chu, H. Soil pH drives the spatial distribution of bacterial communities along elevation on Changbai Mountain. Soil Biol. Biochem. 2013, 57, 204–211. [Google Scholar] [CrossRef]
- Xu, M.; Li, X.; Cai, X.; Gai, J.; Li, X.; Christie, P.; Zhang, J. Soil microbial community structure and activity along a montane elevational gradient on the Tibetan Plateau. Eur. J. Soil Biol. 2014, 64, 6–14. [Google Scholar] [CrossRef]
- Zhang, B.; Liang, C.; He, H.; Zhang, X. Variations in soil microbial communities and residues along an altitude gradient on the Northern slope of Changbai Mountain, China. PLoS ONE 2013, 8, e66184. [Google Scholar] [CrossRef] [PubMed]
- Delmont, T.O.; Prestat, E.; Keegan, K.P.; Faubladier, M.; Robe, P.; Clark, I.M.; Pelletier, E.; Hirsch, P.R.; Meyer, F.; Gilbert, J.A.; et al. Structure, fluctuation and magnitude of a natural grassland soil metagenome. ISME J. 2012, 6, 1677–1687. [Google Scholar] [CrossRef] [PubMed]
- Uroz, S.; Ioannidis, P.; Lengelle, J.; Cébron, A.; Morin, E.; Buée, M.; Martin, F. Functional assays and metagenomic analyses reveals differences between the microbial communities inhabiting the soil horizons of a Norway spruce plantation. PLoS ONE 2013, 8, e55929. [Google Scholar] [CrossRef] [PubMed]
- Uroz, S.; Buée, M.; Murat, C.; Frey-Klett, P.; Martin, F. Pyrosequencing reveals a contrasted bacterial diversity between oak rhizosphere and surrounding soil. Environ. Microbiol. Rep. 2010, 2, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Andreote, F.D.; Jiménez, D.J.; Chaves, D.; Días, A.C.F.; Luvizotto, D.M.; Dini-Andreote, F.; Fasanella, C.C.; Lopez, M.V.; Baena, S.; Taketani, R.G.; et al. The Microbiome of Brazilian mangrove sediments as revealed by metagenomics. PLoS ONE 2012, 7, e38600. [Google Scholar] [CrossRef] [PubMed]
- Thompson, C.; Beys-Da-Silva, W.; Santi, L.; Berger, M.; Vainstein, M.; Guimarães, J.; Vasconcelos, A.T. A potential source for cellulolytic enzyme discovery and environmental aspects revealed through metagenomics of Brazilian mangroves. AMB Express 2013, 3, 65. [Google Scholar] [CrossRef] [PubMed]
- Nazaries, L.; Tate, K.R.; Ross, D.J.; Singh, J.; Dando, J.; Saggar, S.; Baggs, E.M.; Millard, P.; Murrell, J.C.; Singh, B.K. Response of methanotrophic communities to afforestation and reforestation in New Zealand. ISME J. 2011, 5, 1832–1836. [Google Scholar] [CrossRef] [PubMed]
- Tate, K.R. Soil methane oxidation and land-use change–from process to mitigation. Soil Biol. Biochem. 2015, 80, 260–272. [Google Scholar] [CrossRef]
- Ollivier, J.; Töwe, S.; Bannert, A.; Hai, B.; Kastl, E.M.; Meyer, A.; Su, M.X.; Kleineidam, K.; Schloter, M. Nitrogen turnover in soil and global change. FEMS Microbiol. Ecol. 2011, 78, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Uroz, S.; Tech, J.J.; Sawaya, N.A.; Frey-Klett, P.; Leveau, J.H.J. Structure and function of bacterial communities in ageing soils: Insights from the Mendocino ecological staircase. Soil Biol. Biochem. 2014, 69, 265–274. [Google Scholar] [CrossRef]
- Xu, Z.; Hansen, M.A.; Hansen, L.H.; Jacquiod, S.; Sørensen, S.J. Bioinformatic approaches reveal metagenomic characterization of soil microbial community. PLoS ONE 2014, 9, e93445. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cong, J.; Lu, H.; Li, G.; Qu, Y.; Su, X.; Zhou, J.; Li, D. Community structure and elevational diversity patterns of soil Acidobacteria. J. Environ. Sci. 2014, 26, 1717–1724. [Google Scholar] [CrossRef] [PubMed]
- Kersters, K.; de Vos, P.; Gillis, M.; Swings, J.; Vandamme, P.; Stackebrandt, E. Introduction to the Proteobacteria. In The Prokaryotes; Dwarkin, M., Falkow, S., Rosenberg, E., Schleifer, K.H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; Volume 5, pp. 3–37. [Google Scholar]
- Salminen, J.P.; Roslin, T.; Karonen, M.; Sinkkonen, J.; Pihlaja, K.; Pulkkinen, P. Seasonal variation in the content of hydrolyzable tannins, flavonoid glycosides, and proanthocyanidins in oak leaves. J. Chem. Ecol. 2004, 30, 1693–1711. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Schimel, J.P.; Holden, P.A. Variations in microbial community composition through two soil depth profiles. Soil Biol. Biochem. 2003, 35, 167–176. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Cederlund, H.; Wessén, E.; Enwall, K.; Jones, C.M.; Juhanson, J.; Pell, M.; Philippot, L.; Hallin, S. Soil carbon quality and nitrogen fertilization structure bacterial communities with predictable responses of major bacterial phyla. Appl. Soil Ecol. 2014, 84, 62–68. [Google Scholar] [CrossRef]
- Hortal, S.; Bastida, F.; Armas, C.; Lozano, Y.M.; Moreno, J.L.; García, C.; Pugnaire, F.I. Soil microbial community under a nurse-plant species changes in composition, biomass and activity as the nurse grows. Soil Biol. Biochem. 2013, 64, 139–146. [Google Scholar] [CrossRef]
- Wang, J.; Soininen, J.; Zhang, Y.; Wang, B.; Yang, X.; Shen, J. Contrasting patterns in elevational diversity between microorganisms and macroorganisms. J. Biogeogr. 2011, 38, 595–603. [Google Scholar] [CrossRef]
- Fierer, N.; Mccain, C.M.; Meir, P.; Zimmermann, M.; Rapp, J.M.; Silman, M.R.; Knight, R. Microbes do not follow the elevational diversity patterns of plants and animals. Ecology 2011, 92, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Defossez, E.; Courbaud, B.; Marcais, B.; Thuiller, W.; Granda, E.; Kunstler, G. Do interactions between plant and soil biota change with elevation? A study on Fagus sylvatica. Biol. Lett. 2011, 7, 699–701. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Franklin, J.F.; Spies, T.A. Contrasting microclimates among clearcut, edge and interior of old-growth Douglas-fir forest. Agric. For. Meteorol. 1993, 63, 219–237. [Google Scholar] [CrossRef]
- Pennanen, T. Microbial communities in boreal coniferous forest humus exposed to heavy metals and changes in soil Ph—A summary of the use of phospholipid fatty acids, Biolog® and 3H-thymidine incorporation methods in field studies. Geoderma 2001, 100, 91–126. [Google Scholar] [CrossRef]
- Villadas, P.J.; Fernández-López, M.; Ramírez-Saad, H.; Toro, N. Rhizosphere-bacterial community in Eperua falcata (Caesalpiniaceae) a putative Nitrogen-fixing tree from French Guiana rainforest. Microb. Ecol. 2007, 53, 317–327. [Google Scholar] [CrossRef] [PubMed]
- De Meyer, S.E.; Coorevits, A.; Willems, A. Tardiphaga robiniae gen. nov., sp. nov., a new genus in the family Bradyrhizobiaceae isolated from Robinia pseudoacacia in Flanders (Belgium). Syst. Appl. Microbiol. 2012, 35, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Klenk, H.P.; Clément, C.; Ouhdouch, Y.; van Wezel, G.P. Taxonomy, physiology, and natural products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 1–43. [Google Scholar] [CrossRef] [PubMed]
- Hruska, K.; Kaevska, M. Mycobacteria in water, soil, plants and air: A review. Vet. Med. 2012, 57, 623–679. [Google Scholar]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).