1. Introduction
Tropical dry forests which exhibit diverse patterns of leaf lifespan among dominant species provide interesting systems to investigate how interactions of seasonal environmental conditions influence leaf structural traits, gas exchange, and water relations. Unlike temperate forests where leaf phenology and longevity are strongly synchronized by predictable seasonal progressions of temperature and major changes in photoperiod, tropical dry forests with 4–6 months or more of drought commonly lack such cues [
1]. Moreover, both deciduous and evergreen species often co-occur as dominants. The longer leaf lifespan in evergreens has an advantage in that it enables carbon assimilation to begin quickly in the wet season in tropical dry forests without the investment of carbon cost for the construction of new leaves. Drought-deciduous trees, however, have an advantage if they are able to maintain hydraulic capacity in stems during the dry season and quickly produce new leaves with high photosynthetic capacity with the onset of the wet season or even earlier in the late dry season.
There has been an expanding research interest in comparing and contrasting the ecophysiology of evergreen and deciduous leaf phenologies in dry tropical forests [
2]. Much of this focus has centered on divergent strategies of hydraulics and seasonal gas exchange. Evergreen trees in neotropical dry forests typically have low maximum hydraulic efficiency coupled with safety from xylem embolism in contrast to drought-deciduous trees which maximize carbon gain with associated water loss for a short period in the wet season with an efficient vascular system [
3,
4]. Detailed studies comparing plant water relations and hydraulics of evergreen and drought deciduous trees have been carried out in dry rainforests in Queensland [
5] and dry forests in Costa Rica [
6]. More recently, there have been studies comparing and contrasting the ecophysiology of evergreen trees from semi-evergreen forests and deciduous trees from dry forests in Thailand [
7,
8,
9]. These latter studies have shown differences in ecophysiological adaptations of evergreen compared to drought deciduous trees in Thai forests. The upper canopy leaves of evergreen trees possess the ability to use water more conservatively compared with drought-deciduous trees because of significant differences in the photoprotective mechanisms [
8,
9]. However, differing modes of adaptation among drought deciduous tree species have not been reported.
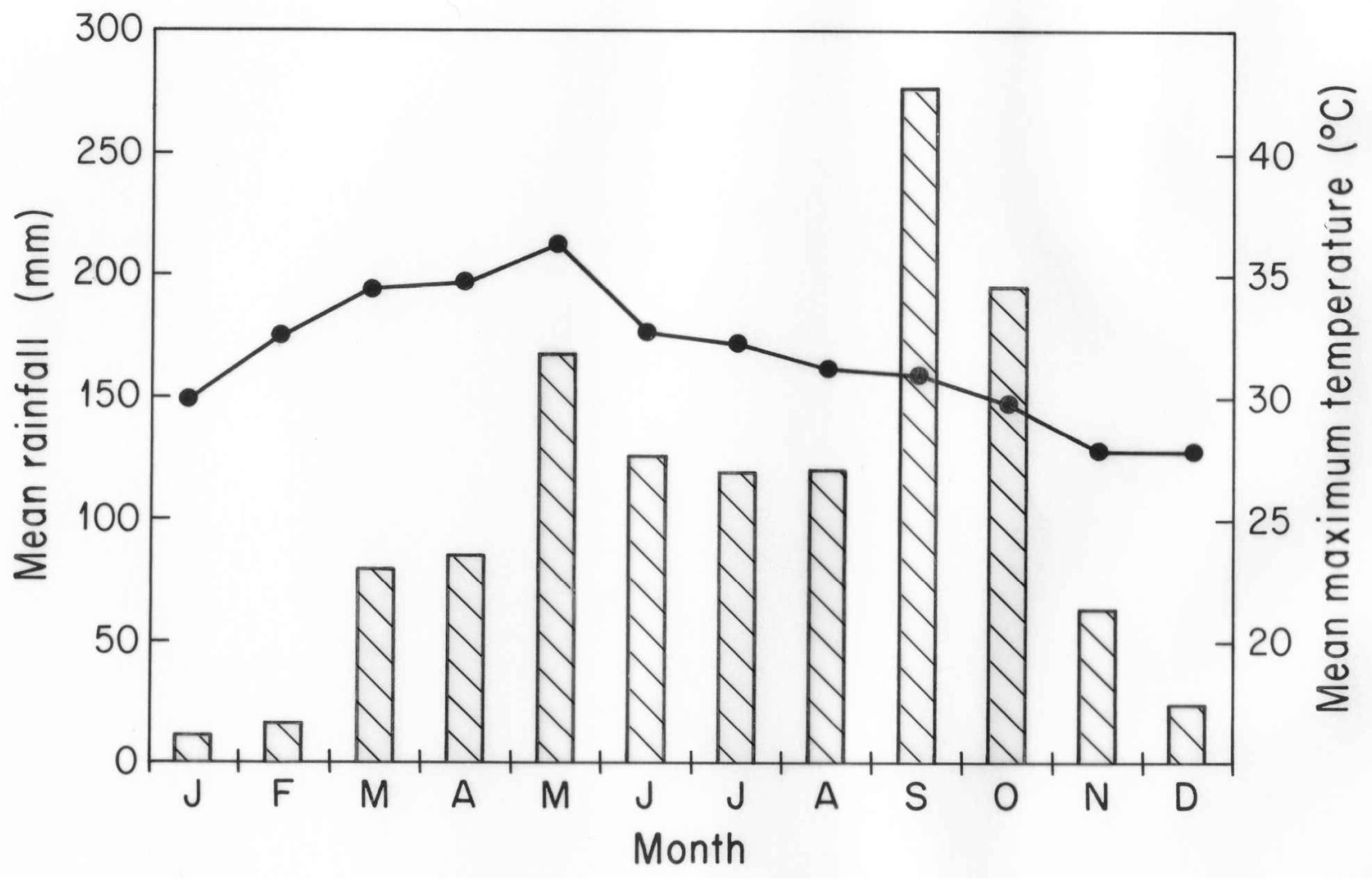
Deciduous dipterocarp forests represent a widespread forest type in Southeast Asia, covering extensive areas from northwestern India and Myanmar across Thailand to the Mekong River drainage of Laos, Cambodia and Vietnam [
10,
11,
12]. Seen late in the dry season in March or April, this forest community is virtually 100% deciduous. The climatic conditions which characterize deciduous dipterocarp forests present severe environmental stresses for plant growth, with drought stress as a major issue. Mean annual rainfall is about 1000–1300 (1500) mm over the range of this community, but strong monsoon influences provide 5–7 months of drought conditions from late November to late April or early May in which no more than 20% of this annual total falls. With high daily temperatures, potential evapotranspiration may exceed precipitation for up to nine months of the year [
12]. Furthermore, soils are characteristically shallow and rocky, providing limited water storage capacity [
12,
13]. Given this severe potential for drought stress, it is not surprising that the community is deciduous. Leaf fall is not synchronous, however, with some species beginning to drop leaves in December and others not beginning leaf fall until March or April [
12,
14]. This is not surprising as leaf abscission in tropical deciduous forests is not always driven by extensive cavitation of xylem vessels [
5,
15]. Hydraulic traits associated with leaf shedding in the dry season have been shown to be variable and more dependent on species than on leaf phenology [
8,
9].
Most species in deciduous dipterocarp forest are leafless in February and March, but new leaves form in April well before the end of the dry season [
1,
12]. It is this surprising presence of new leaves late in the dry season with high ambient temperatures that presents a second environmental stress. Seasonal movements of the Intertropical Convergence Zone bring unusual warm temperatures to Southeast Asia in April and May. Daily mean maximum air temperatures in these months reach 38–40 °C, providing severe heat stress under continuing drought conditions [
12]. There is additional concern for the future of this and other forest communities in Southeast Asia because of potential impacts of increases in mean and extreme temperatures that are predicted to occur in the future under models of global change [
16,
17].
Our objectives in this paper have been to compare and contrast leaf morphology, phenology, and ecophysiological traits of seasonal water relations, stomatal conductance, and leaf temperature balance which may relate to ecological success in members of the Dipterocarpaceae and Fabaceae which form the great majority of tree biomass and canopy cover in deciduous dipterocarp forests. Responses to seasonal drought in this community have been observed to differ between these two families. We compared leaf traits and environmental response of new mature leaves formed in the late dry season to gas exchange, water relations, and associated leaf temperature conditions. We focused attention on two species in each family: Shorea siamensis Miq. and S. obtusa Wall. in the Dipterocarpaceae and Xylia xylocarpa Craib and Hutch. and Dalbergia oliveri Prain in the Fabaceae. We additionally include a limited survey of leaf traits in other tree species in these two families from the same deciduous dipterocarp forest community to determine if a phylogenetic pattern of adaptive traits exist.
3. Results
Leaf traits compared and contrasted between dipterocarp and legume species in the deciduous dipterocarp forest at Sakaerat display many similarities, but also two significant patterns of difference. Previous research has shown that leaf longevity of tree species exhibit significant variability between species and within individual species on an interannual basis, and appear to lack a phylogenetic relationship [
24]. This longevity may range from 7 months in
Shorea obtusa and as long as 10 months in
S. siamensis, while the legume
Sindora siamensis retains leaves on average for 11 months (
Table 1). There were significant differences in individual leaf area of four dipterocarp species compared to the range of individual leaflet area, the functional photosynthetic unit, in legumes, but specific leaf weight was not significantly different. The two dominant
Shorea species at Sakaerat have large leaves with
S. siamensis averaging 180 cm
2 in area, while leaf blades of
S. obtusa had a mean area of 112 cm
2, far larger than that of any of the legume leaflets.
Xylia kerrii and
Dalbergia oliveri had leaflets averaging 33 and 14 cm
2 in area, respectively.
Mean specific leaf weights (SLW) were 130 and 118 g m
−2 for the two
Shorea species compared to the legumes
Xylia xylocarpa and
Dalbergia oliveri at 105 and 58 g m
−2, respectively. A broad range of SLW was present in each family with no significant difference in mean values over this range. Additional data on leaf area and specific leaf weight for two additional dipterocarps (
Shorea roxburghii and Dipterocarpus intricatus) and legume species (
Dalbergia lanceolaria, Sindora siamensis, Albizzia odoratissima, and Erythrophleum succirubrum) was consistent with the pattern of small leaflet size but insignificant differences as a group in SLW compared to the dipterocarps (
Table 1).
Leaf concentrations of nitrogen show a phylogenetic signal with higher concentrations in legume species than dipterocarps, consistent with the likelihood of active nitrogen fixation by root nodules. The four dipterocarp species had mean nitrogen concentrations ranging from 17.1 to 19.8 μg g
−1, compared to 24.6 to 30.7 μg g
−1 if
Sindora, a genus not known to fix nitrogen, is excluded (
Table 1).
Mean values of stable carbon isotope ratios (δ13C), a measure of integrated water use efficiency over the formation of that tissue, was not significantly different between dipterocarps and legumes. Both families exhibited a fairly narrow diversity of mean species values that ranged from −28.7 to −27.0 o/oo.
Measurements of late morning rates of photosynthesis of newly mature leaves in late April 1993 did not show a significant difference in rates between the two families, with a range of mean values from 11.2–14.4 μmol m
−2 s
−1 for the four dipterocarps and 8.0–13.0 μmol m
−2 s
−1 in four legumes (
Table 1). Mean rates of stomatal conductance under these measurements were also not significant between the families, with a range of 143–383 mmol m
−2 s
−1 for the dipterocarps and 98–293 mmol m
−2 s
−1 for the legumes.
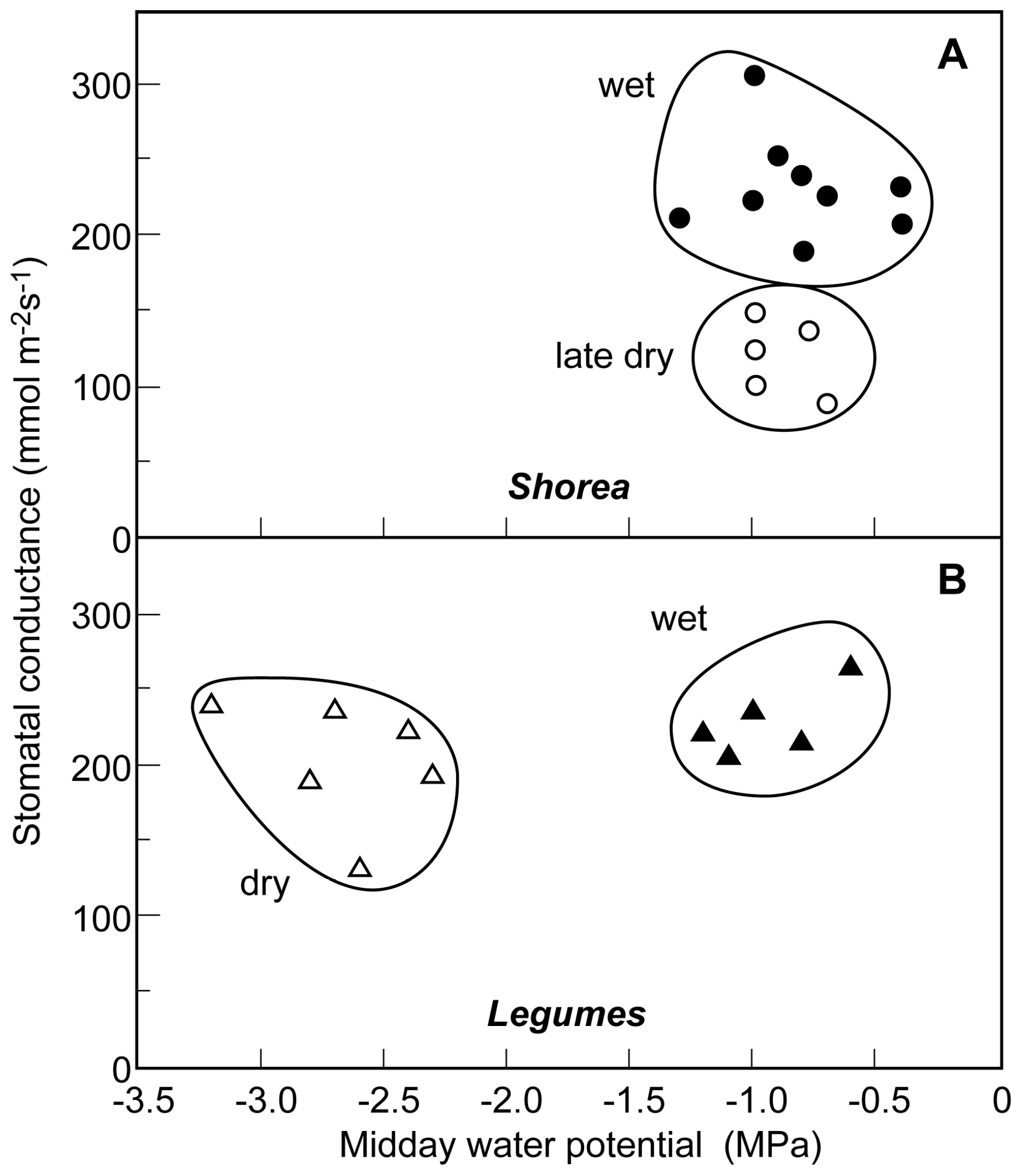
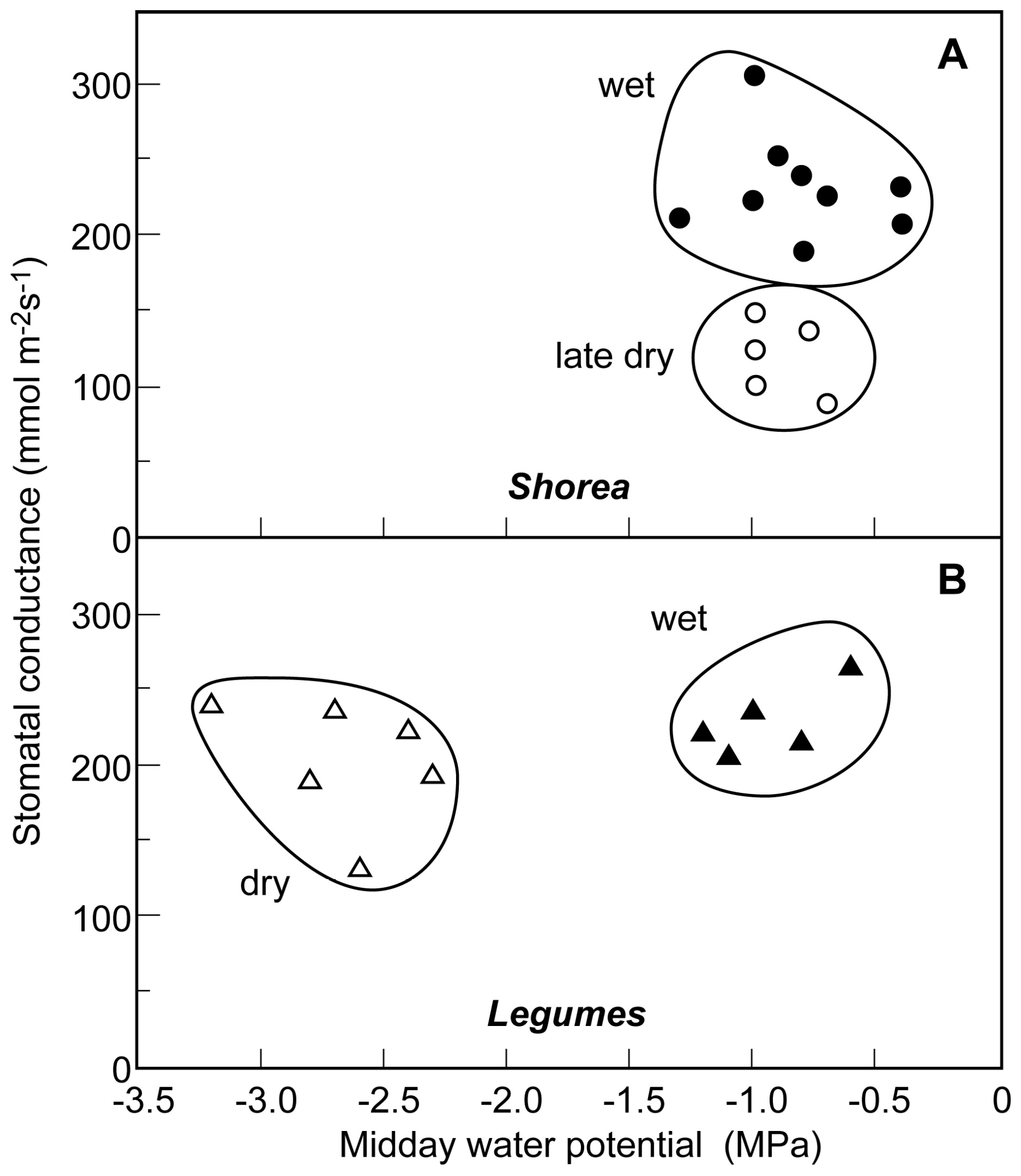
Seasonal studies of leaf water potential and stomatal conductance in the four study species demonstrated divergent patterns of seasonal water relations in the two groups.
Shorea siamensis and
S. obtusa showed little pattern of seasonal change in minimum midday water potentials. Predawn water potentials never dropped below −0.5 MPa in these species and only rarely in the dry season did midday water potentials drop below −1.0 MPa (
Figure 2A). Stomatal conductance remained high through eight months of the year with typical values of 200–300 mmol m
−2 s
−1. Late dry season stomatal response in
Shorea was very different, however. Although tissue water potentials remained high as in the other months of the year, mean maximum stomatal conductance dropped to levels half that typical of the wet and early dry seasons (
Figure 2A).
Data for the legumes
Xylia kerrii and
Dalbergia oliveri showed a very different pattern of seasonal water relations. Wet season conditions of water potential in the legumes in December were quite similar to those found in the two
Shorea species with midday water potentials of −0.6 to −1.2 MPa (
Figure 2B). Mean maximum stomatal conductances in the wet season ranged from 200–265 mmol m
−2 s
−1. Dry season drought stress in late April, however, showed a pronounced effect on patterns of water relations as midday values of leaf water potential dropped to −2.4 to −3.2 MPa. Maximum stomatal conductance was generally somewhat reduced in these months with rates of 130–230 mmol m
−2 s
−1 (
Figure 2B). There was no clear separation of early dry season rates of stomatal conductance (December and January) from late dry season values (April and May) as were present in
Shorea siamensis and
S. obtusa.
Despite the apparently greater water availability for
Shorea siamensis and
S.
obtusa in the late dry season when new leaves were formed, stable carbon isotope analyses of leaf tissues suggested that these species had a similar integrated water use efficiency (WUE) than that present in the two woody legume species, as indicated by leaf δ
13C values. Mean values for these species were −27.0 and −28.6 o/oo, respectively, compared to −27.8 and −28.5 o/oo each for the two legume species, with more negative values indicating lower WUE (
Table 1). These differences are not significant, and a sampling of more species of Dipterocarpaceae and Fabaceae in this community found no pattern of significant difference in values of δ
13C for the two families (
Table 1).
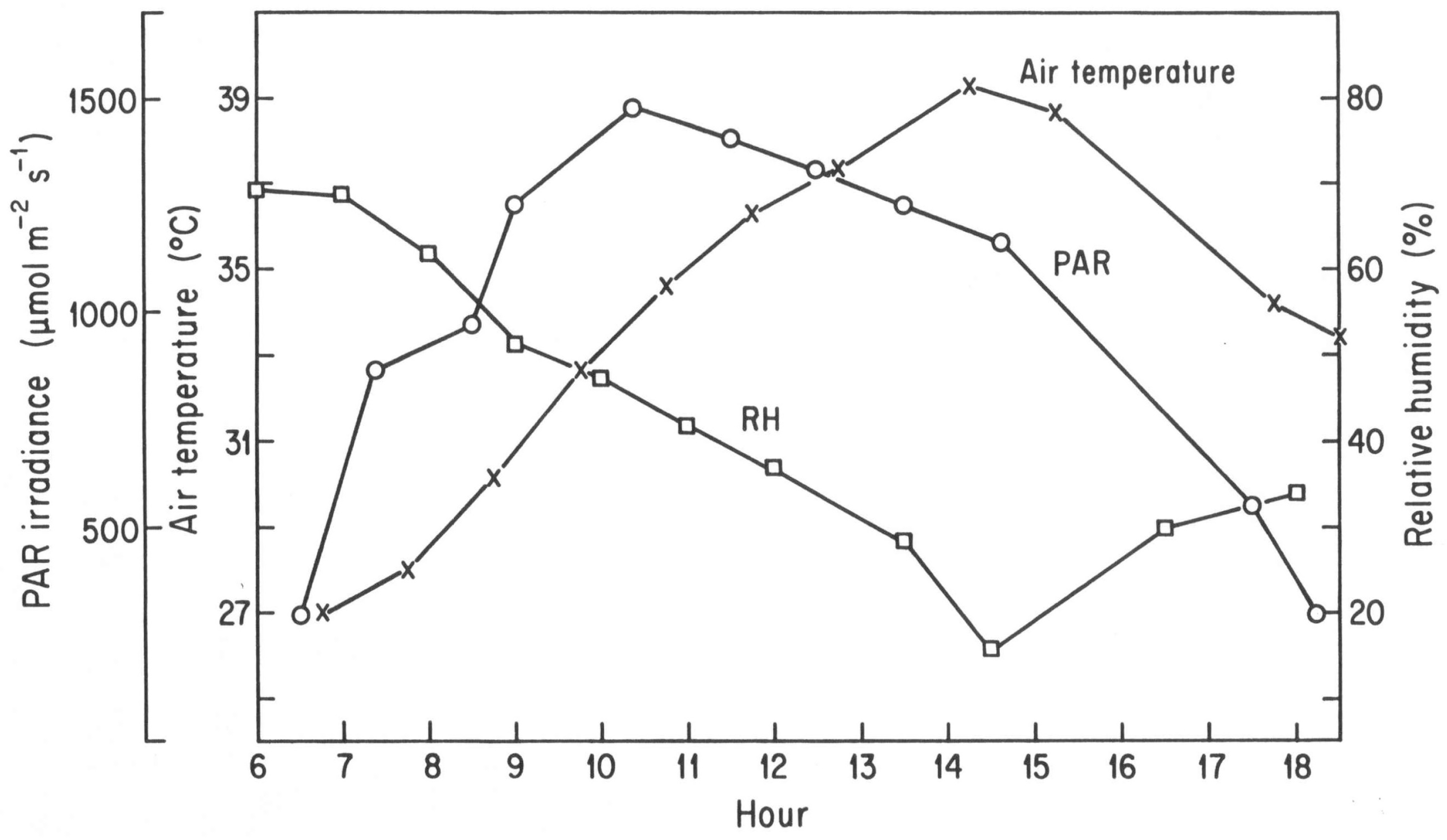
While stomatal conductances of the two
Shorea species commonly peaked at midday in the wet and early dry seasons, the highest conductance rates typically occurred in midmorning in the late dry season. Although solar PAR irradiance peaked at midday, peak ambient air temperatures occurred two hours later. Maximum stomatal conductance occurred at about 09:00 h, associated with increasing ambient air temperatures which peaked at 39 °C at 14:15 h (
Figure 3) and the associated increase in vapor pressure gradient (VPG) which reached 1.01 kPa at this hour.
The response of Shorea species to the high ambient temperatures of 35–39 °C in the late dry season was very different from those of the leaflets of the legume species. Woody legumes including our two study species as well as others such as Dalbergia lanceolaria, Dalbergia conchinchinesis, Albizzia odoratissima, and Erythrophleum succirubrum were commonly wilted by early to mid-morning despite only moderate temperatures, thereby sharply reducing their foliar exposure to solar radiation. Shorea siamensis and S. obtusa did not wilt, however, and changed their mean leaf angles only a few degrees over the course of morning to afternoon.
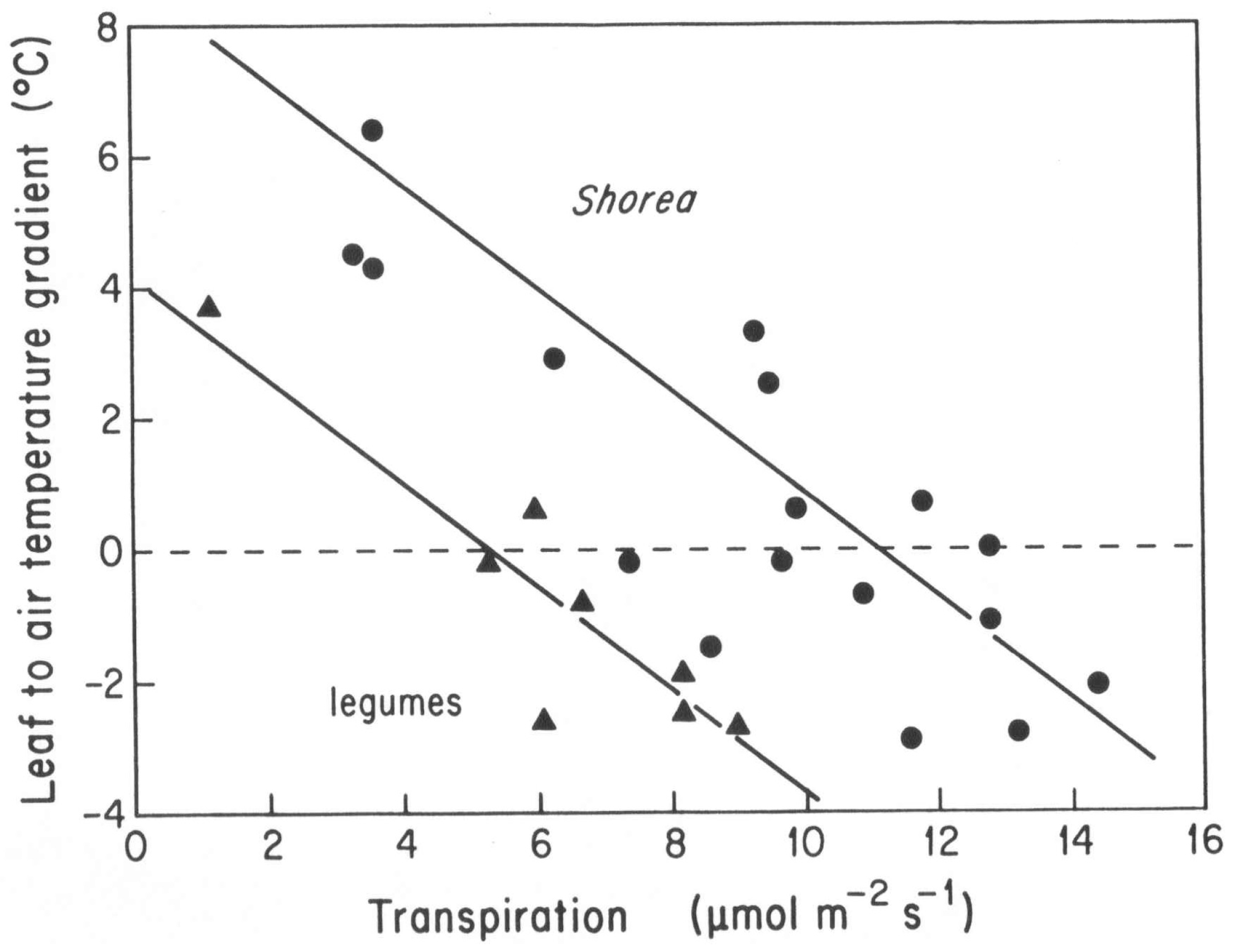
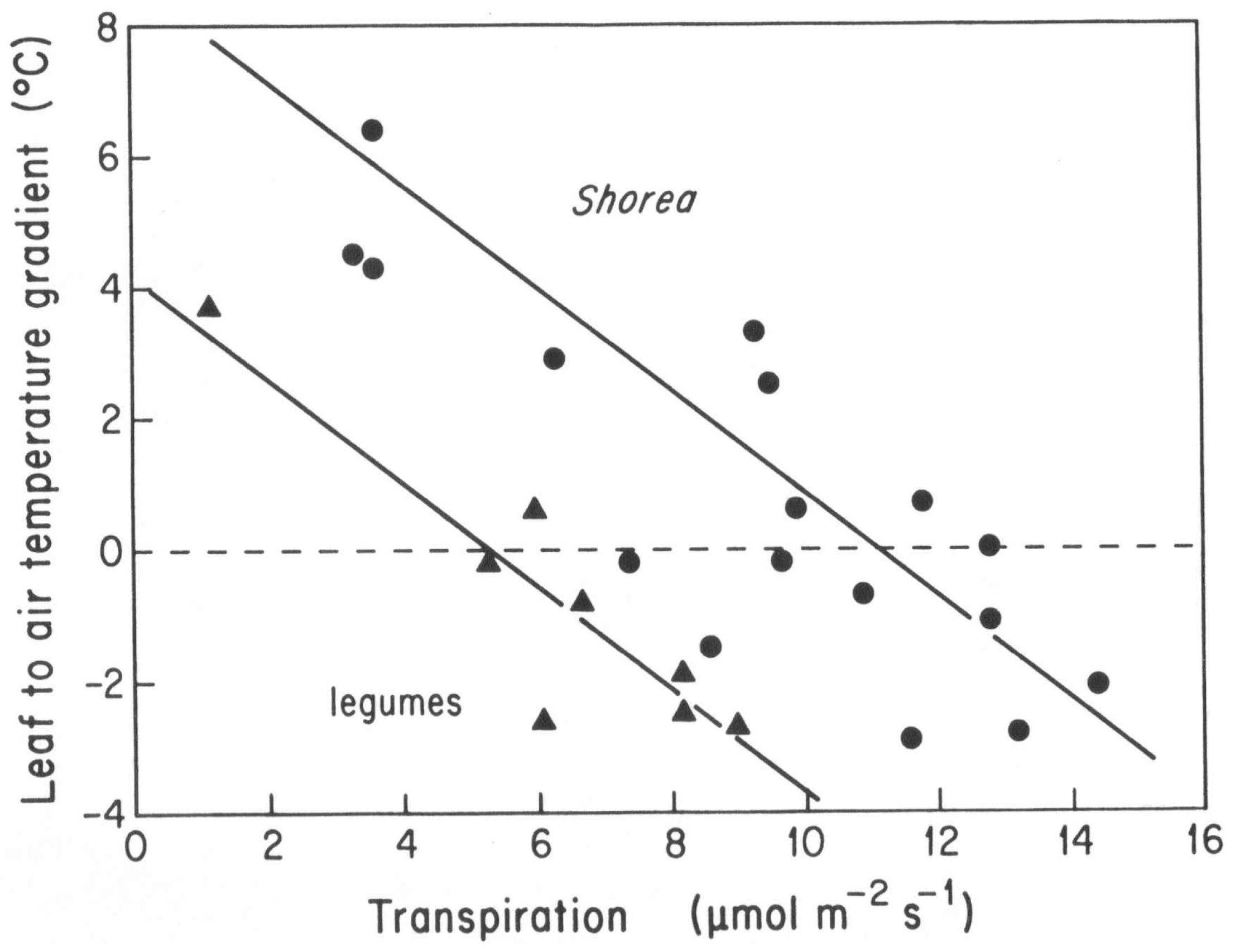
Transpirational cooling has a significantly different effect in the two plant groups. Field studies carried out in late morning and midday on 3–6 May 1993, under full sun and ambient air temperatures above 33 °C found that
Shorea species required transpiration rates around 12 μmol m
−2 s
−1 to prevent heating above ambient, while the smaller legume leaves could maintain the same ambient temperatures with a transpiration rate only half as high (
Figure 4). Extrapolation of these data to the fitted lines indicate that
Shorea without transpirational cooling would reach excess leaf temperatures as high as 7–8 °C over ambient air temperatures compared to only about 4 °C over temperature in
Xylia and
Dalbergia.
Experimental studies with seedlings of Shorea siamensis grown under greenhouse conditions established the interconnection of stomatal conductance in this species with high leaf temperatures. Maximum rates of stomatal conductance occurred at air temperatures from 28–30 °C, but fell sharply at both higher and lower temperatures. At 35 °C, conductance rates dropped to 70% of their maximum levels, and to less than 40% of this level at 40 °C. Stomatal conductance at 45 °C was just 10% of maximum rates. Although the maximum stomatal conductance measured in these well-watered seedlings was only about half maximum values observed with mature trees of this species in the field, the pattern of temperature response of stomatal conductance fitted observed field data, and thus appears to be representative.
4. Discussion
Seasonal changes in canopy stomatal conductance to water vapor and CO2 result from feed-back and/or feed-forward mechanisms driven by diverse signals originated from leaves and/or from roots at each moment. It is not surprising then that the nature of optimal stomatal regulation controlling CO2 assimilation versus water vapor loss can be complex and variable between genotypes and species. Plants face a compromise in reducing transpiration to increase water use efficiency under high temperatures, with an associated impact on assimilation, while at the same time transpiration is a major component of the leaf energy balance.
Adaptive strategies of dealing with high temperatures and photoprotection under extreme dry season temperatures have been addressed for dry evergreen forest trees in Southeast Asia. Photosynthetic depression is present from mid-morning on in the dry season for evergreen dipterocarp species in southern China [
25]. Trees are able to regulate light energy allocation dynamically between photochemistry and heat dissipation during the day, with reduced actual photochemistry and increased heat dissipation in the dry season. The sustained photosynthetic depression in the uppermost-canopy leaves of these species acts as a protective response to prevent excessive water loss and consequent catastrophic leaf hydraulic dysfunction. However, there are differing adaptive responses to high temperatures in drought deciduous tree species [
8,
9]. Two species of drought-deciduous trees from deciduous dipterocarp forests,
Shorea siamensis and
Xylia kerrii included in our study, have been shown to dissipate a large proportion of electron flow through photosynthesis or alternative pathways. In contrast, evergreen trees in adjacent dry evergreen forests employ more conservative water use, reduced assimilation and electron transport rate, and an enhanced capacity for thermal dissipation of excess light energy in photoprotection [
9].
Our comparison of mean species leaf traits between dipterocarp and legume species in deciduous dipterocarp forest found significant differences in leaf/leaflet size and in leaf nitrogen content. However, other leaf morphological and ecophysiological leaf traits such as leaf longevity, specific leaf weight, δ13C, and mean maximum rates of net photosynthesis were not significantly different. Higher leaf nitrogen contents in the legume species were not found to be associated with higher rates of net photosynthesis. The similar small range of values in δ13C suggests that both groups exhibit similar levels of integrated water use efficiency in their growth.
The dipterocarp species in our study present a classic example of an isohydric strategy of adaptation where stomatal regulation maintains a relatively stable minimum water potential over the course of the year compared to the legumes which exhibit anisohydric behavior in which water potential decrease over the dry season as evaporative demand increases [
26,
27]. This effect of stomatal regulation of seasonal patterns of gas exchange is critical in understanding the ability of tree species and genotypes to respond to environmental stress [
28,
29,
30].
No consistent pattern of change in either predawn or midday water potential was present in
Shorea siamensis and
S. obtusa from wet through dry season. In contrast, woody legumes demonstrated a clear seasonal pattern of changing water potential. The lack of an observed seasonal pattern in water availability in
Shorea also differs from the seasonality in water potential present in another Dipterocarpaceae,
Hopea ferrea, which dominates adjacent dry evergreen forests at Sakaerat [
31]. Root excavations have not been carried out in dry dipterocarp forests to determine if
Shorea species are able to exploit fractures in parent rock material to reach ground water pools, but deep root systems have been shown to provide an effective means of avoiding water stress in isohydric species in seasonal mediterranean climates where drought periods may last six months or more [
27,
32].
Drought deciduous trees in Thailand present special challenges for understanding adaptive strategies for new leaf production, which has been widely observed to typically occur a few weeks before the onset of the wet season [
1]. The production of new leaves under the extreme temperatures in the late dry season in Thailand seems counter-intuitive to expected behavior and has led to speculation on what factors act as the trigger of leaf phenological development. Both soil water availability and photoperiod appear to be environmental cues in leaf bud break, but with no common pattern to explain differences between species [
1,
33]. Similar bud break for the production of new leaves in the late dry season has also been reported in India [
34], Australia [
35], Brazil [
36], and Costa Rica [
36,
37]. In most of these observations, the leafless period is variable between species and the synchrony of the timings of leaf fall and bud break among tree species is low. While the selective forces responsible for this seemingly counterproductive phenological characteristic have not been carefully studied, they may well relate to reduced pressure from insect herbivores on sensitive young leaves during the late dry season.
The production of new leaves in the dry season in Thailand exposes these leaves to high ambient air temperatures of 38–40 °C with even higher extremes. Woody legumes responded to this stress by restricting their photosynthetic activity to early morning hours. As higher air temperatures, higher VPG, and lower water potentials increased stress, these species temporarily wilted and folded their leaflets, thereby reducing their heat load from solar irradiance. Legume leaflets under such conditions closely tracked ambient air temperature, or even maintained slightly lower temperatures if stomata remain partially open to allow some generally much lower stomatal conductance.
High levels of irradiance and lowered stomatal conductance in Shorea siamensis and S. obtusa exposed leaves to much higher temperatures. Moderate transpiration rates in the morning allowed these species to maintain leaf temperatures at or below ambient air temperatures. As solar irradiance increased and air temperatures rose, however, these large leaves experienced significant problems maintaining ambient temperatures. Leaf wilting did not occur in these leaves and thus solar energy input remained high. While soil moisture availability for Shorea species might have been sufficient to maintain high rates of stomatal conductance and transpiration in the morning hours, and thus allow leaf cooling below ambient, the high afternoon air temperatures acted to lower rates of stomatal conductance with a consequent negative feedback and lowered transpiration.
Under such afternoon conditions, we have commonly observed Shorea leaf temperatures to reach 44–46 °C as stomatal conductance was reduced. Irregular afternoon winds reduced boundary layer resistances when canopy air was stirred, and afternoon leaf temperatures dropped intermittently to 40–42 °C. A lowering of stomatal conductance by about 50% from wet season levels would be expected to result in leaf temperatures of 38–40 °C, which were characteristic in Shorea over much of the day during the late dry season in April and May before the arrival of monsoon rains. Thus, lowered stomatal conductance was a direct factor promoting high leaf temperatures and thus reduced photosynthetic capacity in Shorea siamensis and S. obtusa during the late dry season.
What is the ecological and ecophysiological advantage of large leaves in
Shorea siamensis and
S.
obtusa? Such an advantage may not exist. Large leaves are characteristic of the Dipterocarpaceae in general and the genus
Shorea specifically. Leaves of both species fall within the normal range of size for
Shorea in seasonal and nonseasonal tropical forests of Southeast Asia [
38]. It seems likely that large leaf size is a genetically canalized trait in this group which is centered in lowland rainforest and montane forests of the Indo-Malaysian region where water stress is not a strong selective factor. Three species of
Dipterocarpus which are characteristic of deciduous dipterocarp forests in Southeast Asia also have large leaves, with mean leaf lengths of 150–250 mm. It seems likely, therefore, that large leaf size is a genetic trait in these two genera, and that it is compensated for in the seasonal monsoon forests of Southeast Asia by alternative strategies of adaptation that may include reduced rates of photochemistry with increased heat dissipation or other biochemical means of increased heat tolerance. It is interesting to note that the five species of Dipterocarpaceae characteristic of deciduous dipterocarp forests (
Shorea siamensis,
S. obtusa,
Dipterocarpus obtusifolius,
D.
tuberculatus, and
D. intricatus) are the only fully deciduous taxa among the more than 550 tree species in this family.
Large leaf sizes also occurred in tree species within several other families in the deciduous dipterocarp forest at Sakaerat. These included Dillenia ovata Wall. ex Hook. f. and Thomson (Dilleniaceae) and Mitragyna rotundifolia (Roxb.) Kuntze (Rubiaceae). Although we have not investigated seasonal patterns of water relations in these species, their diurnal cycles of stomatal physiology in the late dry season appears to parallel those reported here for Shorea species. As with the Dipterocarpaceae, these taxa have evolved from Indo-Malaysian ancestors growing in nonseasonal rainforests. The great majority of woody legumes in the deciduous dipterocarp forest at Sakaerat appear to have adaptive strategies of seasonal water relations similar to that found here for Xylia kerrii and Dalbergia oliveri. These genera are more characteristic of seasonal tropical forest of Southern and Southeastern Asia and do not show a strong Indo-Malaysian ancestry (Larsen 1984). One common legume, however, is an exception to this pattern. Sindora siamensis has large, relatively sclerophyllous leaflets, and does not wilt during the high temperatures of the late dry season. This genus has clear Indo-Malaysian ancestry. While Sindora siamensis has not been studied, it may well have evolved a strategy more like that of the Dipteocarpaceae than other woody legumes.


{kind=link}
{kind=link}
{kind=link}
{kind=link}