
Emerging Needle Blight Diseases in Atlantic Pinus Ecosystems of Spain

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area, Field Observations, and Sampling
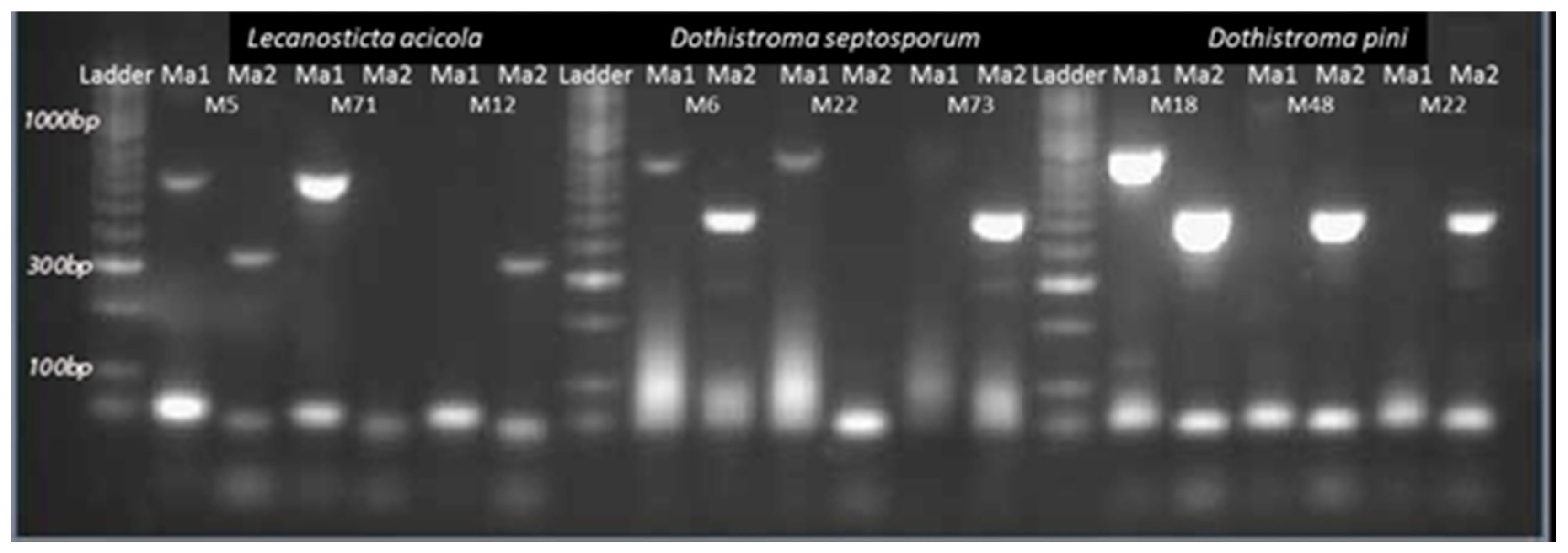
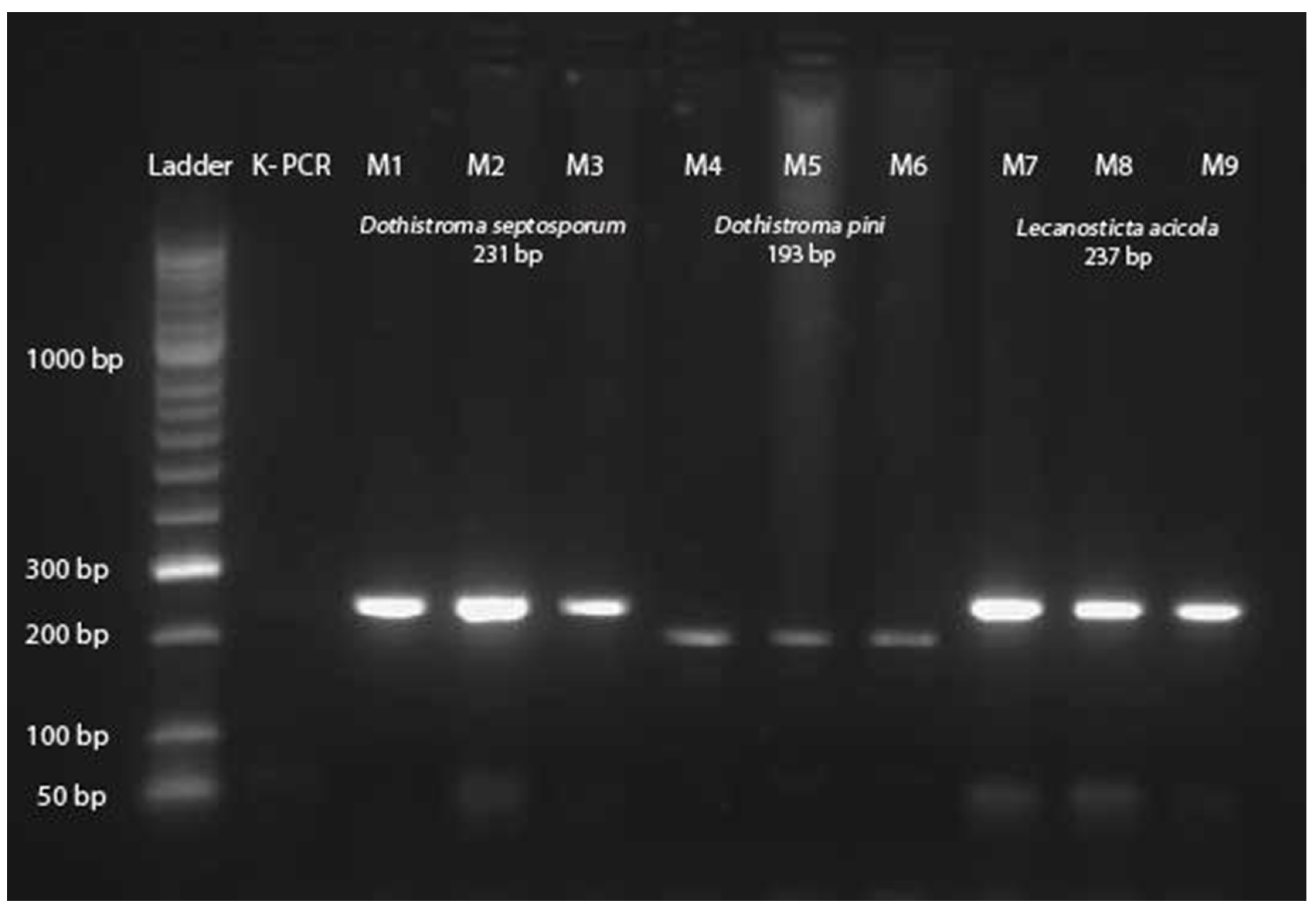
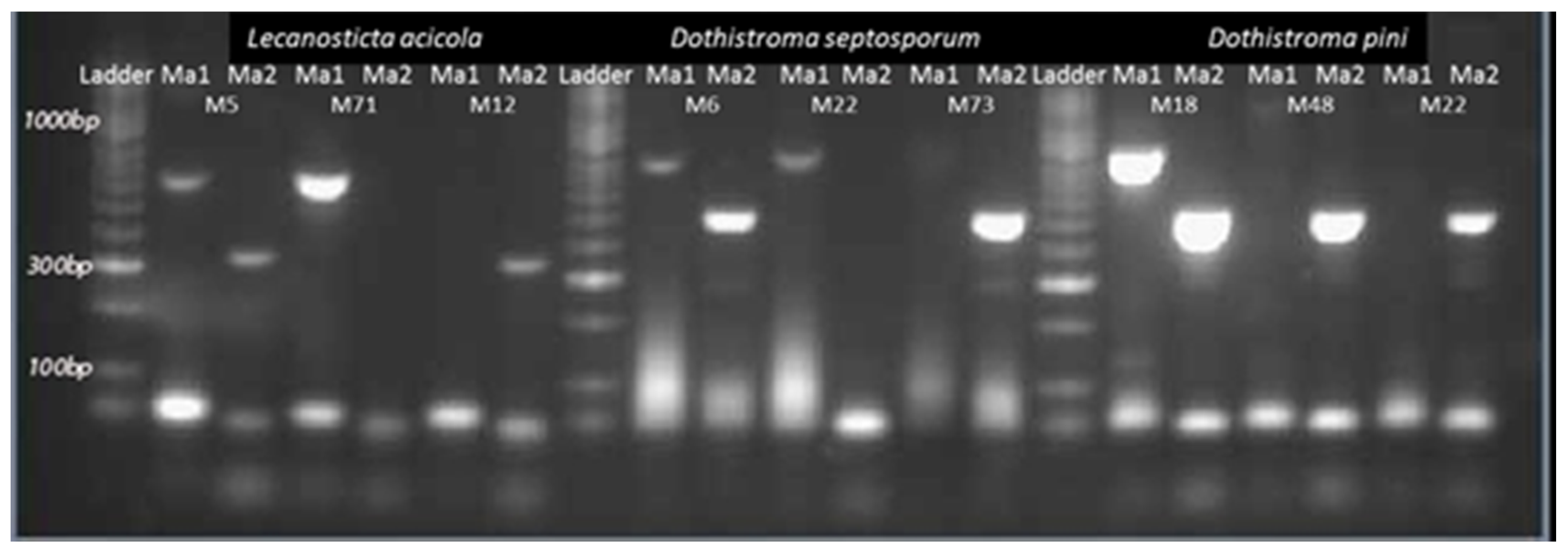
2.2. Laboratory Analyses
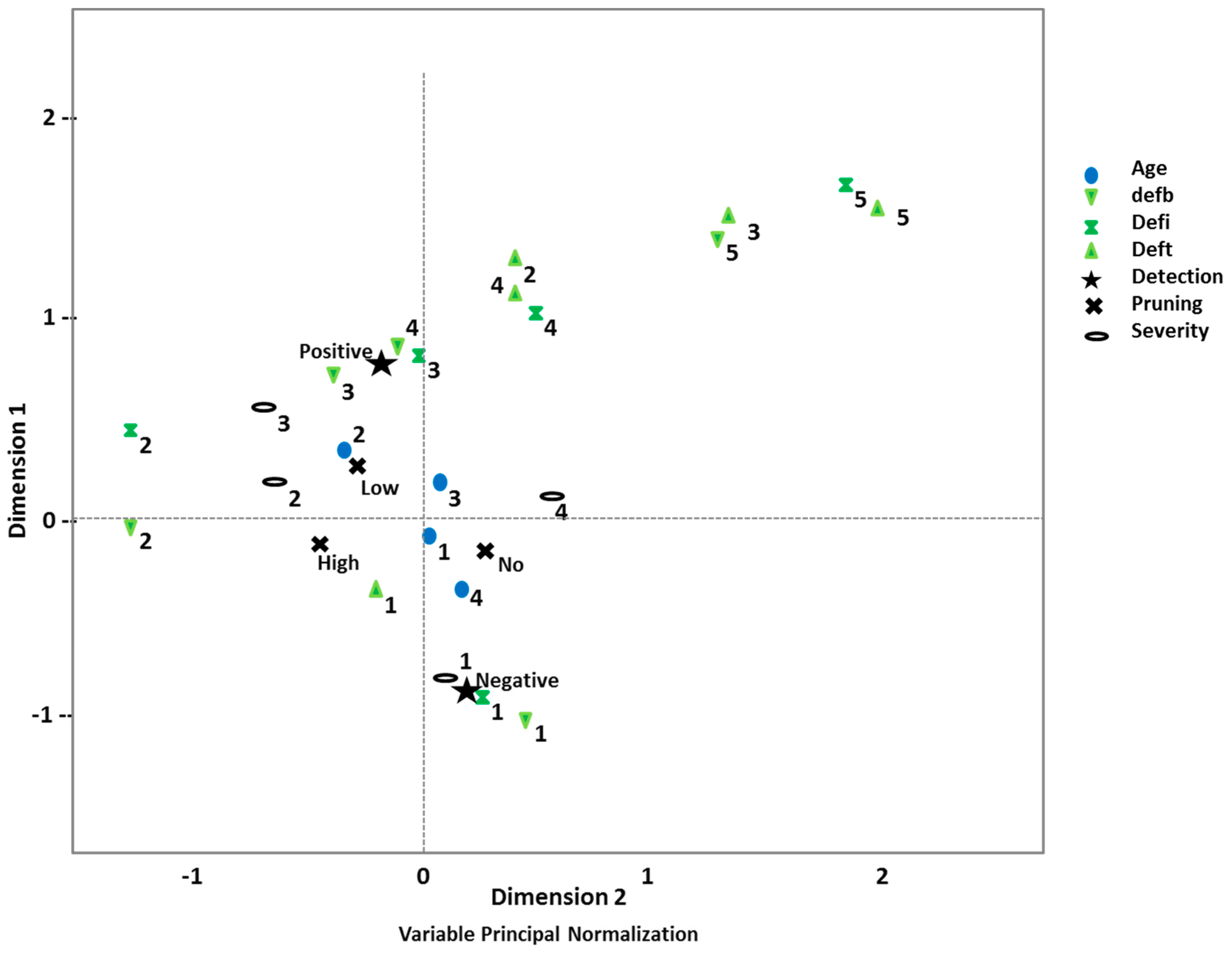
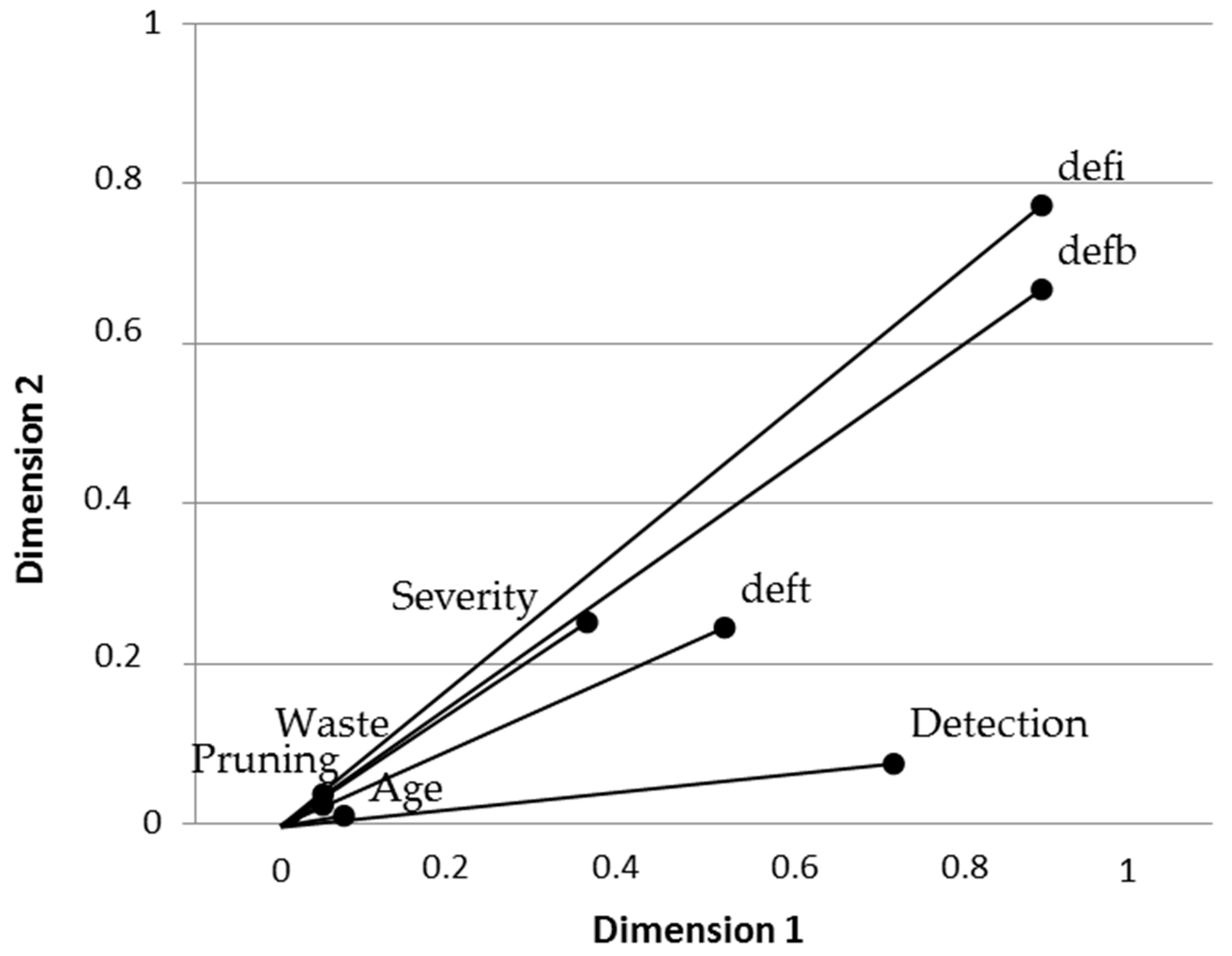
2.3. Statistical Methods
3. Results
3.1. Fungal Species, Host Distribution and Mating Type Detection in the Studied Area
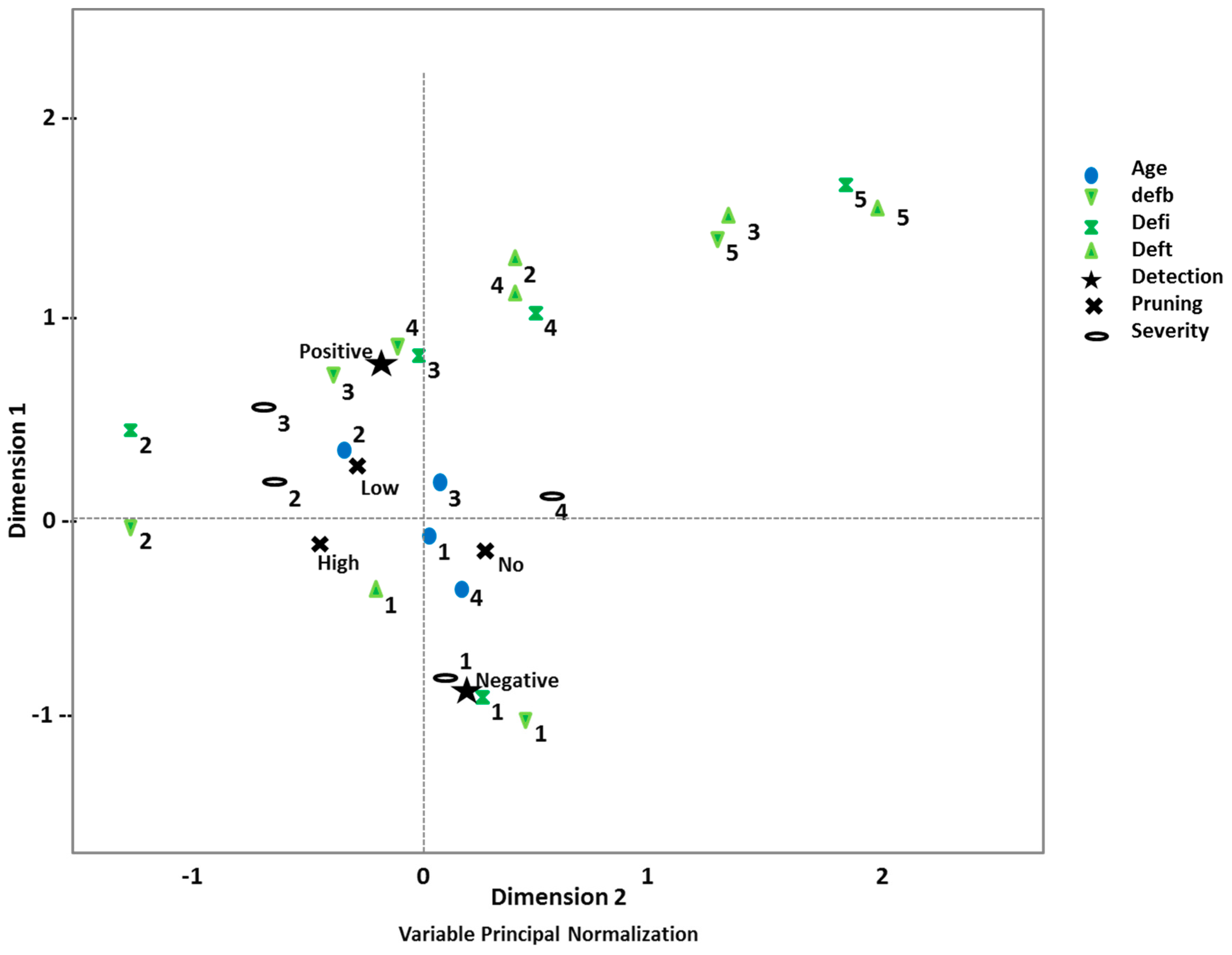
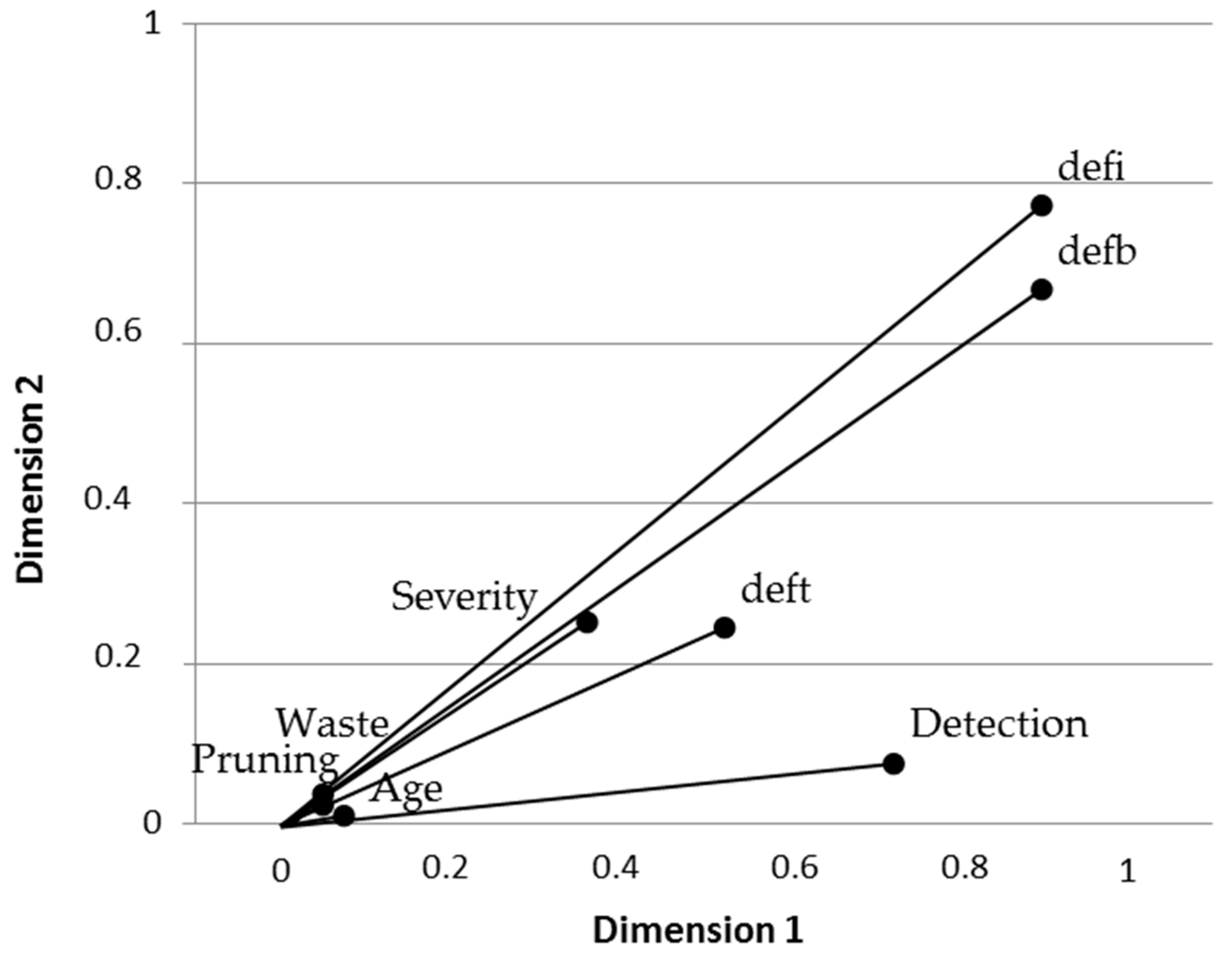
3.2. Disease Impact and Its Connection with Implemented Silvicultural Management
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Drenkhan, R.; Tomešová-Haataja, V.; Fraser, S.; Bradshaw, R.E.; Vahalík, P.; Mullett, M.S.; Martín-García, J.; Bulman, L.S.; Wingfield, M.J.; Kirisits, T.; et al. Global geographic distribution and host range of Dothistroma species: A comprehensive review. For. Pathol. 2016, 46, 408–442. [Google Scholar] [CrossRef]
- Gibson, I.A.S. Pests and diseases of pines in the tropics. For. Pathol. 1979, 9, 126–127. [Google Scholar]
- Gibson, I.A.S. Impact and control of Dothistroma blight of pines. For. Pathol. 1974, 4, 89–100. [Google Scholar] [CrossRef]
- Bulman, L.; Ganley, R.J.; Dick, M. Needle Diseases of Radiata Pine in New Zealand; Scion Client Report No. 13010; Forest Biosecurity Research Council: Rotorua, New Zealand, 2008. [Google Scholar]
- Bradshaw, R.E. Dothistroma (red-band) needle blight of pines and the Dothistromin toxin: A review. For. Pathol. 2004, 34, 163–185. [Google Scholar] [CrossRef]
- Woods, A.J. Species diversity and forest health in northwest British Columbia. For. Chron. 2003, 79, 892–897. [Google Scholar] [CrossRef]
- Woods, A.J.; Coates, K.D.; Hamann, A. Is an unprecedented Dothistroma needle blight epidemic related to climate change? Bioscience 2005, 55, 761–769. [Google Scholar] [CrossRef]
- Brown, A.; Webber, J. Red band needle blight of conifers in Britain; Forestry Commission: Edinburgh, UK, 2008; pp. 1–8.
- Piou, D.; Ioos, R. First report of Dothistroma pini, a recent agent of the Dothistroma needle blight (DNB), on Pinus radiata in France. Plant Dis. 2014, 98, 841. [Google Scholar] [CrossRef]
- Bulman, L.S.; Bradshaw, R.E.; Fraser, S.; Martín-García, J.; Barnes, I.; Musolin, D.L.; La Porta, N.; Woods, A.J.; Diez, J.; Koltay, A.; et al. A worldwide perspective on the management and control of Dothistroma needle blight. For. Pathol. 2016, 46, 472–488. [Google Scholar] [CrossRef]
- Gibson, I.A.S. Dothistroma blight of Pinus radiata. Annu. Rev. Phytopathol. 1972, 10, 51–72. [Google Scholar] [CrossRef]
- Ana Magan, F.J.F. Red band disease of Pinus radiata. Comun. I.N.I.A. Prot. Veg. 1975, 3, 1–16. [Google Scholar]
- Van der Pas, J.B. Reduced early growth rate of Pinus radiata caused by Dothistroma pini. N. Zeal. J. For. Pathol. 1981, 11, 210–220. [Google Scholar]
- Evans, H.C. The Genus Mycosphaerella and Its Anamorphs Cercoseptoria, Dothistroma and Lecanosticta on Pines; Mycology Paper No. 153; Commonwealth Mycological Institute: Kew, Surrey, UK, 1984; pp. 1–102. [Google Scholar]
- Drenkhan, R.; Hantula, J.; Vuorinen, M.; Jankovsky, L.; Müller, M.M. Genetic diversity of Dothistroma septosporum in Estonia, Finland and Czech Republic. For. Pathol. 2012, 136, 71–85. [Google Scholar] [CrossRef]
- Jactel, H.; Brockerhoff, E.; Duelli, P. A test of the biodiversity-stability theory: Meta-analysis of tree species diversity effects on insect pest infestations, and re-examination of responsible factors. In Forest Diversity and Function—Temperate and Boreal Systems; Scherer-Lorenzen, M., Körner, C., Schulze, E.-D., Eds.; Springer: Heidelberg/Berlin, Germany, 2005; Volume 176, pp. 235–262. [Google Scholar]
- Groenewald, M.; Barnes, I.; Bradshaw, R.E.; Brown, A.V.; Dale, A.; Groenewald, J.Z.; Lewis, K.J.; Wingfield, B.D.; Wingfield, M.J.; Crous, P.W. Characterization and distribution of mating type genes in the Dothistroma needle blight pathogens. Phytopathology 2007, 97, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Woods, A.J.; Martín-García, J.; Bulman, L.; Vasconcelos, M.W.; Boberg, J.; La Porta, N.; Peredo, H.; Vergara, G.; Ahumada, R.; Brown, A.; et al. Dothistroma needle blight, weather and possible climatic triggers for the disease’s recent emergence. For. Pathol. 2016, 46, 443–452. [Google Scholar] [CrossRef]
- Jankovsky, L.; Palovcikova, D.; Dvorak, M.; Tomsovsky, M. Records of brown spot needle blight related to Lecanosticta acicola in the Czech Republic. Plant Prot. Sci. 2009, 45, 16–18. [Google Scholar]
- European and Mediterranean Plant Protection Organization (EPP/EPPO). Exigences Spécifiques de Quarantaine; EPPO Technical Documents; EPPO: Paris, France, 1990. [Google Scholar]
- European and Mediterranean Plant Protection Organization (OEPP/EPPO). Mycosphaerella Dearnessii and Mycosphaerella Pini.; Bulletin 38; EPPO: Paris, France, 2008; pp. 349–362. [Google Scholar]
- Brown, A.; Clayden, H. Time for action: Dothistroma (red band) needle blight in Scotland. Forestry 2012, 18, 16–17. [Google Scholar]
- Barnes, I.; Walla, J.A.; Bergdahl, A.; Wingfield, M.J. Four new host and three new state records of Dothistroma Needle Blight caused by Dothistroma pini in the United States. Plant Dis. 2014, 98, 1443. [Google Scholar] [CrossRef]
- Bradshaw, R.E.; Bhatnagar, D.; Ganley, R.J.; Gillman, C.J.; Monahan, B.J.; Seconi, J.M. Dothistroma pini, a forest pathogen, contains homologs of aflatoxin biosynthetic pathway genes. Appl. Environ. Microb. 2002, 68, 2885–2892. [Google Scholar] [CrossRef]
- Bulman, L.S.; Gadgil, P.D.; Kershaw, D.J.; Ray, J.W. Assessment and Control of Dothistroma Needle-Blight; Forest Research Bulletin No. 229; Forest Research: Rotorua, New Zealand, 2004; pp. 1–48. [Google Scholar]
- Ioos, R.; Fabre, B.; Saurat, C.; Fourrier, C.; Frey, P.; Marcais, B. Development, comparison, and validation of real-time and conventional PCR tools for the detection of the fungal pathogens causing brown spot and red band needle blights of pine. Phytopathology 2010, 100, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Janoušek, J.; Krumböck, S.; Kirisits, T.; Bradshaw, R.E.; Barnes, I.; Jankovský, L.; Stauffer, C. Development of microsatellite and mating type markers for the pine needle pathogen Lecanosticta acicola. Australas. Plant Pathol. 2014, 43, 161–165. [Google Scholar] [CrossRef]
- Greenacre, M.; Hastie, T. The Geometric Interpretation of Correspondence Analysis. J. Am. Stat. Assoc. 1987, 82, 437–447. [Google Scholar] [CrossRef]
- Paoletti, M.; Buck, K.W.; Brasier, C.M. Selective acquisition of novel mating type and vegetative incompatibility genes via interspecies gene transfer in the globally invading eukaryote Ophiostoma novo-ulmi. Mol. Ecol. 2006, 15, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Ivory, M.H. Resistance to Dothistroma needle blight induced in Pinus radiata by maturity and shade. Br. Mycol. Soc. 1972, 59, 205–212. [Google Scholar] [CrossRef]
- Garcia, J.; Kummerow, J. Infection of Monterey Pine graftings with Dothistroma pini. Plant Dis. Rep. 1970, 54, 403–404. [Google Scholar]
- Power, A.B.; Dodd, R.S. Early differential susceptibility of juvenile seedlings and more mature stecklings of Pinus radiata to Dothistroma pini. N. Z. J. For. Sci. 1984, 14, 223–228. [Google Scholar]
- Mead, D.J. Sustainable Management of Pinus Radiata Plantations; FAO Forestry Paper No. 170; FAO: Rome, Italy, 2013. [Google Scholar]
- Emmingham, W.; Fitzgerald, S. Pruning to Enhance Tree and Stand Value; Extension Service. Publication number EC 1457; Oregon State University: Orvallis, OR, USA, 1995; pp. 1–12. [Google Scholar]
- O’Hara, K. Pruning Wounds and Occlusion: A Long-Standing Conundrum. J. For. 2007, 105, 131–138. [Google Scholar]
- Gadgil, P.D. Dothistroma Needle Blight; Forest Pathology in New Zealand No. 5; Forest Research Institute: Rotorua, New Zealand, 1984; pp. 1–8. [Google Scholar]
- Mullett, M.S.; Tubby, K.V.; Webber, J.F.; Brown, A.V. A reconsideration of natural dispersal of the pine pathogen Dothistroma septosporum. Plant Pathol. 1984. [Google Scholar] [CrossRef]
- Scott, C.A. The Influence of Low Pruning on Dothistroma Pini Infection in Pinus radiata in Kaingaroa Forest. Forest Pathology Report No.39; Forest Research Institute: Rotorua, New Zealand, 1973; pp. 1–21. [Google Scholar] [CrossRef]
- Hood, I.A.; Ramsden, M. Dothistroma Needle Blight on Pinus Radiata at Gambubal Forest, QFRI Disease Management Research Trials; Interim Report; Queensland Forest Research Institute: Indooroopilly, Austarlia, 1996. [Google Scholar]
- Bulman, L.S.; Dick, M.A.; Ganley, R.J.; McDougal, R.L.; Schwelm, A.; Bradshaw, R.E. Dothistroma needle blight; Gonthier, P., Nicolotti, G., Eds.; CABI: Boston, MA, USA, 2013; pp. 436–457. [Google Scholar]
- Gibson, I.A.S.; Christensen, P.S.; Munga, F.M. First observations in Kenya of a foliage disease of Pines caused by Dothistroma pini Hulbary. Commonw. For. Rev. 1964, 43, 31–48. [Google Scholar]
- Sutherst, R.W.; Maywald, G.F.; Kriticos, D.J. CLIMEX Version 3: User’s Guide. Available online: http://www.hearne.software/getattachment/0343c9d5-999f-4880-b9b2-1c3eea908f08/Climex-User-Guide.aspx (accessed on 27 December 2016).
- Watt, M.S.; Kriticos, D.J.; Alcaraz, S.; Brown, A.V.; Leriche, A. The hosts and potential geographic range of Dothistroma needle blight. For. Ecol. Manag. 2009, 257, 1505–1519. [Google Scholar] [CrossRef]
- EFSA Panel on Plant Health (PLH). Scientific Opinion on the risk to plant health posed by Dothistroma septosporum (Dorog.) M. Morelet (Mycosphaerella pini E. Rostrup, syn. Scirrhia pini) and Dothistroma pini Hulbary to the EU territory with the identification and evaluation of risk. EFSA J. 2013, 11, 173. [Google Scholar] [CrossRef]
- Hepting, G.H. Climate and forest diseases. Annu. Rev. Phytopathol. 1963, 1, 31–50. [Google Scholar] [CrossRef]
- Dale, V.H.; Joyce, L.A.; McNulty, S.; Neilson, R.P.; Ayres, M.P.; Flannigan, M.D.; Hanson, P.J.; Irland, L.C.; Lugo, A.E.; Peterson, C.J.; et al. Climate change and forest disturbances. BioScience 2001, 51, 723–734. [Google Scholar] [CrossRef]
- Moncho, R.; Chust, G.; Caselles, V. Análisis de la precipitación del País Vasco en el período 1961–2000 mediante reconstrucción espacial. Nimbus 2009, 23, 149–170. [Google Scholar]
- Brown, A.; Green, S.; Hendry, S. Needle diseases of pine. Forestry Commission: Edinburgh, UK, 2005; pp. 1–12. [Google Scholar]
- Guernier, V.; Hochberg, M.E.; Guegan, J.F.O. Ecology drives the worldwide distribution of human diseases. PLoS Biol. 2004, 2, 740–746. [Google Scholar] [CrossRef] [PubMed]
- Koltay, A. Incidence of Dothistroma septospora (Dorog.) Morlet in the Austrian pine (Pinus nigra Arn.) stands in Hungary and results of chemical control trials. Novenytermeles 2001, 37, 231–235. [Google Scholar]
- Forestry Commission. Forests and biodiversity, UK Forestry Standard Guidelines. Forestry Commission: Edinburgh, UK, 2011. Available online: http://www.forestry.gov.uk/PDF/FCGL001.pdf/$FILE/FCGL001.PDF (accessed on 27 December 2016). [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Category Codes | Description of Characteristic | Detection of Needle Blight Species | Number of Plantations | |
|---|---|---|---|---|---|
| Positive | Negative | ||||
| Age (years) | 1 | <15 | 28 | 30 | 58 |
| 2 | 15–<20 | 43 | 22 | 65 | |
| 3 | 20–<30 | 55 | 39 | 94 | |
| 4 | ≥30 | 39 | 55 | 94 | |
| Pruning | 0 | no pruning | 78 | 89 | 167 |
| 1 | low | 72 | 39 | 111 | |
| 2 | high | 17 | 16 | 33 | |
| Severity (%) | 1 | <30 | 28 | 68 | 96 |
| 2 | 30–<60 | 33 | 26 | 59 | |
| 3 | 60–<90 | 51 | 18 | 69 | |
| 4 | ≥90 | 64 | 23 | 87 | |
| Defoliation at the base (defb %) | 1 | <20 | 16 | 99 | 115 |
| 2 | 20–<40 | 37 | 28 | 65 | |
| 3 | 40–<60 | 37 | 6 | 43 | |
| 4 | 60–<80 | 46 | 2 | 48 | |
| 5 | ≥80 | 36 | 4 | 40 | |
| Defoliation at the middle (defi %) | 1 | <20 | 24 | 106 | 130 |
| 2 | 20–<40 | 49 | 15 | 74 | |
| 3 | 40–<60 | 51 | 8 | 59 | |
| 4 | 60–<80 | 18 | 3 | 21 | |
| 5 | ≥80 | 27 | 0 | 27 | |
| Defoliation at the top (deft %) | 1 | <20 | 106 | 125 | 231 |
| 2 | 20–<40 | 36 | 7 | 43 | |
| 3 | 40–<60 | 14 | 0 | 14 | |
| 4 | 60–<80 | 17 | 1 | 18 | |
| 5 | ≥80 | 5 | 0 | 5 | |
| Fungal spp./Hosts | nig 1 | hal 2 | pin 3 | pine 4 | rad 5 | syl 6 | men 7 | Total |
|---|---|---|---|---|---|---|---|---|
| None detected | 44 | 3 | 15 | 1 | 70 | 9 | 1 | 143 |
| D. septosporum | 21 | 0 | 2 | 0 | 0 | 2 | 1 | 26 |
| D. septosporum and D. pini | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 3 |
| D. septosporum and L. acicola | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 2 |
| Lecanosticta acicola | 3 | 0 | 0 | 0 | 134 | 0 | 0 | 137 |
| Number of plots | 72 | 3 | 17 | 1 | 205 | 12 | 2 | 311 |
| Variables | B (S.E.) | Wald | ODDS (C.I. 95%) |
|---|---|---|---|
| Age | 5.37 | ||
| Age (1) | 1.27 (0.70) * | 3.24 | 3.54 (0.89–14.08) |
| Age (2) | 1.50 (0.71) ** | 4.35 | 4.47 (1.09–18.23) |
| Age (3) | 0.64 (0.60) | 0.98 | 1.9 (0.53–6.75) |
| Pruning | ns | 0.91 | |
| Waste | ns | 0.001 | |
| Thining | ns | 0.002 | |
| Severity | 15.98 | ||
| Severity (1) | −1.69 (0.76) ** | 4.96 | 0.18 (0.04–0.81) |
| Severity (2) | −0.81 (0.61) | 1.74 | 0.44 (0.13–1.48) |
| Severity (3) | 1.11 (0.69) | 2.52 | 3.03 (0.77–11.89) |
| defb | 16.07 | ||
| defb (1) | −2.89 (1.33) ** | 4.73 | 0.06 (0.00–0.75) |
| defb (2) | −1.57 (1.14) | 1.89 | 0.21 (0.02–1.95) |
| defb (3) | 0.53 (0.97) | 0.3 | 1.71 (0.25–11.53) |
| defb (4) | 3.28 (1.42) ** | 5.28 | 26.56 (1.62–43.53) |
| defi | ns | 5.71 | |
| deft | ns | 0.82 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ortíz de Urbina, E.; Mesanza, N.; Aragonés, A.; Raposo, R.; Elvira-Recuenco, M.; Boqué, R.; Patten, C.; Aitken, J.; Iturritxa, E. Emerging Needle Blight Diseases in Atlantic Pinus Ecosystems of Spain. Forests 2017, 8, 18. https://doi.org/10.3390/f8010018
Ortíz de Urbina E, Mesanza N, Aragonés A, Raposo R, Elvira-Recuenco M, Boqué R, Patten C, Aitken J, Iturritxa E. Emerging Needle Blight Diseases in Atlantic Pinus Ecosystems of Spain. Forests. 2017; 8(1):18. https://doi.org/10.3390/f8010018
Chicago/Turabian StyleOrtíz de Urbina, Esther, Nebai Mesanza, Ana Aragonés, Rosa Raposo, Margarita Elvira-Recuenco, Ricard Boqué, Cheryl Patten, Jenny Aitken, and Eugenia Iturritxa. 2017. "Emerging Needle Blight Diseases in Atlantic Pinus Ecosystems of Spain" Forests 8, no. 1: 18. https://doi.org/10.3390/f8010018
APA StyleOrtíz de Urbina, E., Mesanza, N., Aragonés, A., Raposo, R., Elvira-Recuenco, M., Boqué, R., Patten, C., Aitken, J., & Iturritxa, E. (2017). Emerging Needle Blight Diseases in Atlantic Pinus Ecosystems of Spain. Forests, 8(1), 18. https://doi.org/10.3390/f8010018